
Abstract
Background
The effect of past climatic changes on the distribution of organisms is a fertile field of research that has been tackled in many different ways. Because the fossil record provides direct access to the chronological and geographic dimensions of biological events occurred in the past, it can be a useful tool for assessing range contractions and expansions related to climatic changes.
Results
Here, we provide support for the ‘recent’ shrinkage of the range of a frigophilous anuran, the common frog, Rana temporaria Linnaeus, 1758, by analyzing the amphibian fossil assemblage coming from the Grotta di Equi, a Late Pleistocene site (about 45 ka) located in the Apuan Alps (northern Apennine chain, Massa-Carrara Province, Italy). Besides, R. temporaria, the assemblage includes cf. Salamandra salamandra Linnaeus, 1758 and Bufo bufo Linnaeus, 1758.
Conclusions
The presence of R. temporaria in the Apuan Alps during a cold interglacial phase at an elevation that is much lower than the minimum current elevation in the region (about 300 m lower) supports the previously reported hypothesis that the Pleistocene coolings favored the dispersal of this frigophilous species along the Apennine chain and that its current, locally disjointed distribution is the effect of the post Pleistocene raise in temperature. Our results concur in predicting that the current global warming will further affect, negatively, the range of this frigophilous species by further reducing its distribution in the southern sectors of its range.
Similar content being viewed by others

Background
The effect of past climatic changes on the distribution of organisms is a fertile field of research that has been tackled in many different ways. In recent years, phylogeographic studies have used genetic and molecular methods for investigating the history of the distribution of various taxa of amphibians (among others: Duellman [1999]; Riberon et al. [2001]; Zeisset and Beebee [2001]; Bonnet and Chippindale [2004]; Vences et al. [2004]; Alexandrino et al. [2007]; Crottini et al. [2007]; Recuero et al. [2007]; Vences and Wake [2007]; Stöck et al. [2008]; Shepard and Burbrink [2009]; Mattoccia et al. [2011]; Chiari et al. [2012]). As far as Europe is concerned, Rana temporaria has been the subject of several papers (Veith et al. [2003]; Palo et al. [2004]; Gómez and Lunt [2006]; Zeisset and Beebee [2008]; Teacher et al. [2009]; Stefani et al. [2012]), probably because its current broad range (Gasc et al. [1997]; Kuzmin [1999]; Sillero et al. [2014]) and the fact that it is adapted to relatively cold environments render this species a good model for evaluating the range changes that occurred during and after the Pleistocene coolings. The fossil record is only marginally taken into consideration in these papers, but it could potentially provide relevant information thanks to the fact that it represents the only way to directly access to the chronological and geographic dimensions of biological events occurred in the past.
Here, we present data concerning R. temporaria and the associated amphibians from the Grotta di Equi (Figure 1), a Late Pleistocene archeo-palaeontological site, located in the northern sector of the Apennine chain, Italy (De Stefani, [1917]; Ghezzo et al., [2014]). The Equi fossil fauna, though not re-examined in detail in recent times, is generally correlated with the middle of Marine Isotope Stage (MIS) 3, thus approximately in the range of 24 to 56 ka (Ghezzo et al. [2014]). A new excavation survey of the cave confirmed the presence of in situ archaeological deposits with radiometric ages consistent with the biochronological interpretation (radiocarbon ages of 43,700 ± 1,900 and 44,000 ± 2,200 before present; Bigagli et al. [2013]). The site is especially famous for the Mousterian lithics (that testify for the former human frequentation of the cave) and for the richness and diversity of fossil material, in particular, of the cave bear Ursus spelaeus (Regalia [1911]; De Stefani [1917], [1923]; Rellini [1918]; Battaglia [1919]; Branchini [1928]). The presence of anurans in the cave has long been reported (De Stefani [1917]; Battaglia [1919]; Branchini [1928]), even though the authors did not operate any identification on the material or just referred the remains to ‘Bufo vulgaris’ or ‘Rana sp.’ as still indicated in the collection tags.

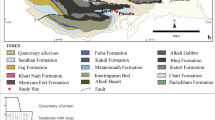
Current Italian distribution of the common frog and the location of the palaeontological sites. Current Italian distribution of the common frog, Rana temporaria (modified from Sindaco et al. [2006]) and location of the palaeontological sites that yielded fossils of this taxon. Note that the Grotta di Equi opens on the slopes of a Tuscan massif, the Apuan Alps, where now the common frog is extinct and that the elevation of the cave is about 300 m lower than the current lower range of the common frog in Tuscany. Italian Pleistocene localities with ascertained presence of the common frog: 1) Grotta di Equi (MS); 2) Grotta della Volpe, Val D’Avesa (VR) + Grotta di Veja (A), S. Anna d’Alfaedo (VR) + Grotta della Fata Nana, Bosco Chiesanuova (VR) + Valdiporro, Bosco Chiesanuova (VR) + Cengelle 1, Monte Tondo, Soave (VR); 3) Grotta Generosa, San Fedele d’Intelvi (CO).
The study of the historical collections and of the remains coming from the recently re-opened excavation has allowed us to better define the composition of the herpetofauna of this cave as preliminary presented in a conference abstract by Bartolini et al. ([2013]). In this paper, we briefly describe the amphibian material found in the Grotta di Equi, discuss its identification and then focus on the common frog, R. temporaria for the possible implications that its discovery in this site could have on the biogeographic history of the taxon in the Apennine area.
Methods
The studied material is part of the historical collection of the Museum of Natural History of the University of Florence, Section of Geology and Palaeontology, under the accession numbers IGF 8239 V-IGF 8602 V. The only exception is represented by a caudal vertebra (US39/001) that was recovered during recently re-opened excavations, and its repository is under the aegis of the ‘Soprintendenza per i Beni Archeologici della Toscana’.
The anatomical description and identification of the fossils recovered from the site were done following Francis ([1934]), Haller-Probst and Schleich ([1994]), Sanchiz ([1998]), Bailon ([1999]), Delfino ([2002]), and Pitruzzella ([2009]). The fossil remains were compared with a reference osteological collection (Massimo Delfino Herpetological Collection - MDHC) stored at the Department of Earth Sciences of the University of Turin. Of particular relevance were the following specimens: Salamandra salamandra: MDHC 23, 124, 205, 212; Triturus carnifex: MDHC 18, 38, 60, 85 to 87, 145; Bufo bufo: MDHC 13, 14, 32, 33, 50, 51. 53, 56, 104, 211; Bufo gr. Bufo viridis: MDHC 35, 36, 58, 123; Pelophylax esculentus skl: MDHC 5, 24, 52, 71; Pelophylax ridibundus: MDHC 1, 4, 206 to 208; Rana arvalis: MDHC 182, 183; Rana dalmatina: MDHC 2, 37, 127, 128, 129, 130, 131; Rana italica: MDHC 31, 64; R. temporaria: MDHC 3, 16, 184 to 186, 210.
Results
Amphibia Linnaeus, 1758
Caudata Fischer von Waldheim, 1813
Salamandridae Golfuss, 1820
Salamandra Garsault, 1764
Salamandra salamandra Linnaeus, 1758
cf. Salamandra salamandra Linnaeus, 1758
Referred material: US39/001, a caudal vertebra
(Figure 2)

cf. Salamandra salamandra Linnaeus, 1758 from the Grotta di Equi. Caudal vertebra (US39/001) in dorsal (a), ventral (b), anterior (c), and left lateral (d) views. Scale bar equals 1 mm. h.ar, haemal arch; n.sp, neural spine; pr.tr, processus trasversus; prezyg, prezygapophyses.
Description
Caudal vertebra: the only caudal vertebra (Figure 2) is poorly preserved but still presents the centrum, the prezygapophyses, the transverse processes, and the neural and haemal arches. The centrum of the vertebra is opisthocoelous and it measures approximately 3 mm, from the preserved portion of the condyle to the ventral edge of the cotyle.
In dorsal view, the vertebra appears quite long with pronounced transverse processes; the prezygapophyses are anterolaterally directed. The neural arch is mostly missing, but a central and fairly short neural spine is still preserved. In ventral view, the haemal arch shows a very little crest; it expands posteriorly beyond the edge of the centrum with a bilobed tip.
Identification
According to Haller-Probst and Schleich ([1994]), Delfino ([2002]), and Pitruzzella ([2009]), the described features of US39/001 identify it as a caudal vertebra because: the neural spine is short and present only in the central portion of the neural arch (missing both in the anterior and posterior parts of it); the neural canal is very small and semicircular; the haemal arch is well developed and bilobed. Moreover, its proportions fit the general morphology of caudal vertebrae. Other characters permit to distinguish US39/001 from other caudates, in particular, Salamandrina, Speleomantes, and Triturus sensu lato (Delfino [2002]; [2004]), and to focus on Salamandra: relative large size, oval-shaped prezygapophyses, opisthocoelous centrum with large cotyle, haemal arch shaped as an irregular semi-cone with long fan-like lateral margins in its ventro-posterior portion. Due to the absence of osteological characters allowing the specific identification of Salamandra spp. (Villa et al. [2014]), the caudal vertebra from the Grotta di Equi is referred to cf. S. salamandra taking into consideration its young age and its geographic provenience (Sindaco et al. [2006]; Vanni and Nistri [2006]).
Anura Fischer von Waldheim, 1813
Bufonidae Gray, 1825
Bufo Garsault, 1764
Bufo bufo Linnaeus, 1758
Referred material (total number of remains: 5): humerus: 2; radio-ulna: 1; tibiofibula: 1; ilium: 1.
(Figure 3)

Bufo bufo Linnaeus, 1758 from the Grotta di Equi. Right humerus IGF 8574 V in dorsal (a) and ventral (b) views; left ilium IGF 8573 V in lateral view (c). Scale bars equal 5 mm. acet, acetabulum; acet.bor, acetabular margin; e.cap, eminentia capitata; e.rad, epicondylus radialis; e.ul, epicondylus ulnaris; tub.sup, tuber superior; vex, pars cylindriformis.
Description
Humerus: the two recovered humeri are only partially preserved as the proximal portion of both is missing (Figure 3a,b). The general aspect of the remains is pretty robust and solid. The diaphysis is slightly curved, and the condyle (eminentia capitata) is displaced in a rather lateral position. No crests are present on these specimens. IGF 8572 V is approximately 17 mm long (from the tip of the condyle to the proximal edge).
Radio-ulna: although broken in its distal part, the main features of IGF 8574 V are still identifiable, as the olecranon and the sulcus longitudinalis. This element is 22 mm long.
Tibiofibula: due to preservational reasons, it is not possible to understand if the groove that marks the point of median fusion of the tibia and the fibula is the sulcus proximalis or rather the sulcus distalis. The general robustness and an evident foramen nutritium further characterize this remain.
Ilium: the ilium is not complete, as the pars ascendens and the pars descendens are partially broken and the anterior part of the ilial shaft (pars cylindriformis) is missing (Figure 3c). However, it is possible to state that there are no dorsal crest, fossae, nor laminar structures (in particular, the ‘calamita ridge’). The tuber superior is clearly evident and triangle-shaped. Furthermore, it has to be underlined that the robustness that characterizes this remain clearly distinguishes it from all the other ilia retrieved from the site.
Ranidae Rafinesque, 1814
Rana Linnaeus, 1758
Rana temporaria Linnaeus, 1758
Referred material (total number of remains 359): angulosplenial: 1; coracoid: 1; humerus: 74; radio-ulna: 24; ilium: 43; ischium: 1; femur: 55; tibiofibula: 129; tarsal: 11; metapodial: 4; vertebra: 5; urostilum: 11.
(Figure 4)

Rana temporaria Linnaeus, 1758 from the Grotta di Equi. Right angulosplenial IGF 8588 V in dorsal view (a); female right humerus IGF 8492 V in dorsal (b) and ventral (c) views; male right humerus IGF 8290 V in dorsal (d) and ventral (e) views; right ilium IGF 8316 V in lateral (f), medial (g), and posterior (h) views. Scale bars equal 5 mm. acet, acetabulum; acet.bor, acetabular margin; cr.dors, crista dorsalis; cr.lat, crista lateralis; cr.med, crista medialis; cr.ven, crista ventralis; e.cap, eminentia capitata; e.rad, epicondylus radialis; e.ul, epicondylus ulnaris; vex, pars cylindriformis.
Description
Angulosplenial: IGF 8588 V (Figure 4a) is the only amphibian cranial element found in the Grotta di Equi (Figure 4a). It is rather long and thin, with a single well-developed processus coronoideus and a crista paracoronoidea, vertically oriented.
Coracoid: IGF 8581 V is a complete coracoid, with a considerably anteroposteriorly elongated pars epicoracoidalis with the margo fenestralis more curved compared to the margo posterior.
Humerus: in dorsal view, this element is rather straight and the eminentia capitata is spherical and aligned to the main axis of the diaphysis; the ventral crest (crista ventralis) is present, but the paraventral crest (crista paraventralis) is missing. Of the two epicondyles, the epicondylus ulnaris is more pronounced than the radialis. Both female (Figure 4b,c) and male (Figure 4d,e) humeri were found in the Grotta di Equi. Male humeri present well-developed crests (crista lateralis and crista medialis), whereas females lack them both. Moreover, in males, the medial crest is dorsally folded and so developed that is present in most of the medio-distal portion of the humerus. IGF 8290 V, a male humerus, is longer and larger than IGF 8492, a female humerus. The first one is approximately 23 mm long, whereas the second one is 16 mm long.
Radio-ulna: the preservation of these skeletal elements is generally good, so that is easy to recognize their main diagnostic features like the olecranon and the sulcus longitudinalis. Their mean length is of approximately 15 mm.
Ilium: the best preserved elements present two expansions near the acetabular fossa: the dorsal acetabular expansion, pars ascendens, and the ventral one, pars descendens, which appears to be lobe-shaped (Figure 4f,g,h). The ilia host two evident structures: a dorsal crest (crista dorsalis) on top of the ilial shaft (pars cylindriformis) and a dorsal tuberosity (tuber superior) joined by a marked dorsal ‘notch’. The dorsal crest in some specimens can be more curved medially than in others. In addition, there is a supracetabular fossa, variably deep, but the preacetabular fossa and the interiliac sulcus are both missing. The ilial shaft is quite slender and slightly curved downward. IGF 8318 V is approximately 20 mm long (from the proximal edge of the acetabulum to the tip of the ilial shaft).
Femur: the femora referred to this taxon share a thin diaphysis characterized by a double curvature in an S shape and by the absence of crests.
Tibiofibula: slender and proportionally long tibiofibulae are quite abundant in the Grotta di Equi. They are quite elongated and clearly show the two sulci, sulcus proximalis and sulcus distalis, and the foramina nutritia.
Tarsals and metapodials: very thin and elongated tarsal bones and metapodials (with and approximate length of 15 mm) have been tentatively referred to this taxon.
Presacral vertebrae: few presacral vertebrae are procoelous and have long transverse processes. IGF 8579 V is well preserved and characterized by a short centrum, about 3 mm long; the neural canal is quite wide compared to the overall height of the vertebra (2.5 mm out of 4 mm). It has two well-developed transverse processes anterolaterally directed. Prezygapophyses are anteriorly oriented whereas postzygapophyses are posterolaterally directed. In anterior view, the neural arch is strongly curved dorsally, with a median notch (more clearly visible in dorsal view).
Urostilum: the two articular fossae are well defined and circular; no transverse processes are present; the dorsal lamina (crista dorsalis) is particularly tall and anteroposteriorly well-developed.
Identification
The anuran remains have been analyzed using mostly the characters described by Bailon ([1999]). The ilium, in particular, proved to be particularly useful for the identification at specific level. Two distinct morphologies of this element have been found in the Grotta di Equi: one is large, stout, and with no crests whereas the other is slender and with an evident dorsal crest. The general features and proportions allowed us to refer the first morphology to B. bufo; Bufo gr. B. viridis and Bufo calamita, both present nowadays in Europe, can be ruled out since neither the preacetabular fossa nor the lamina calamita are present in the material from the Grotta di Equi. On the other hand, the dorsal crest and its peculiar morphology, such as low height and medial inclination, focus the identification on R. temporaria excluding all the other anuran taxa provided with ilial crest. The other skeletal elements (other than the ilia) have been referred to B. bufo or R. temporaria according to their morphology and proportions, following Bailon ([1999]) and with direct comparison with the extant osteological material of these two species.
Discussion
The amphibian assemblage of the Grotta di Equi
The complete collection of fossil amphibians from the Grotta di Equi consists of 359 skeletal elements of R. temporaria; 5 elements of B. bufo; and one vertebra referred to cf. S. salamandra. Although the great disparity in number of specimens per each species, a common characteristic can be outlined: nearly all fossils but one are postcranial elements (the only cranial element is the angulosplenial IGF 8588 V referred to R. temporaria; Figure 4a). Several reasons could have had a role in determining this bias, like the fragility and the low chance of fossilization of the cranial elements compared to the postcranials, the application of partially destructive collecting and sieving techniques, and the little knowledge of these anatomical elements from those who selected the material about a century ago.
The most relevant result of this study is the identification of fossil remains of R. temporaria for the possible implication they could have for the comprehension of the distribution pattern of this extant taxon in the last 50 ka.
Fossils of R. temporaria were already known in Italy for the following Pleistocene palaeontological localities (see Figure 1): Grotta della Volpe, Val D’Avesa (VR); Grotta di Veja (A), S. Anna d’Alfaedo (VR); Grotta della Fata Nana, Bosco Chiesanuova (VR); Valdiporro, Bosco Chiesanuova (VR); Cengelle 1, Monte Tondo, Soave (VR); and Grotta Generosa, San Fedele d’Intelvi (CO) (see Delfino [2002]; Delfino et al. [2008]; Bona et al. [2009]). The identification of R. temporaria at Grotte dei Balzi Rossi (IM) is actually doubtful and should be verified by the revision of the original material - currently not traced - cited by Brocchi ([1879]).
The occurrence of this taxon in an archaeological site as the Grotta di Equi is not unusual: the association of humans and the common frog has been reported for two prehistoric sites of the French department Jura (Bailon [1997]). One of these, Chalain 3 has provided more than 12,000 skeletal elements of R. temporaria in middens, as well as in human and canine coprolites. This extraordinary palaeoherpetological deposit apparently resulted from the consumption of frogs during their reproductive season (Bailon [1997]). It is worth mentioning that unlike these remains from Chalain 3, which present clear evidence of breakage due to chewing and even corrosion as a result of digestion, the fossils from the Grotta di Equi do not have any of these sign on them, so the possibility of human- or animal-mediated accumulation (and potential transportation) of R. temporaria at Equi should be reasonably ruled out.
Ecological preferences
In terms of ecology, the three amphibian taxa identified at the Grotta di Equi share a broad altitudinal range, being currently distributed in Italy from nearly the sea level up to over 2,000 meter above sea level (m a.s.l.) (Sindaco et al. [2006]), but R. temporaria deserves further comments.
Among the various species of genus Rana, R. temporaria is definitively the one which tolerates the lowest temperatures, being active even at few degrees above zero (Savage [1961]; Bernini and Razzetti [2006]; Vanni and Nistri [2006]; Razzetti et al. [2007]). In fact, its reproductive season begins between February and June, depending on elevation of the breeding site and its exposure, and mating usually takes place in water bodies still partially frozen. In addition, the common frog is an anuran well adapted to a great variety of different habitats, like high pastures, peat-bogs, grasslands, beech woods, deciduous forests, and coniferous forests. In any case, essential aspects are humid and cool areas not far from streams and water bodies. When present at low elevations, ‘it is generally only found in the coolest wooded areas adjacent to water courses especially if high-banked’ (Bernini and Razzetti [2006]: 371). Due to these characteristics, R. temporaria is a taxon with a very broad geographic distribution, spanning from very high latitudes in Scandinavia (up to Cape North) to south-central Europe. It is, however, absent from most of the Mediterranean area (central and southern Iberia, central and southern Italy, southern Balkans) (Bernini and Razzetti [2006]; Sillero et al. [2014]). To the broad geographic distribution corresponds a wide altitudinal range, from few meter on the sea level to about 3,000 (Vences et al. [2003]), but low elevations are preferred in northern and central Europe, whereas it is relegated mostly to hills and mountains in the south (Bernini and Razzetti [2006]).
In the Italian peninsula, the common frog is therefore a hill or mountain species, widespread all over the Alps, where some lacunae in the distribution should be considered as missing data more than areas of absence of the species; on the Apennines, instead, its range is much more fragmented and discontinuous, and this is particularly true the more we move southward. The southernmost records of this taxon are in the area of the Forlì-Cesena, Florence, and Arezzo provinces, but it is also known a relict population in the Latium portion of the Monti della Laga (RI) (Bernini and Razzetti [2006]). The Italian altitudinal distribution fits the whole range reported by Vences et al. ([2003]), spanning from 20 m a.s.l. in Liguria (Savona Province) to about 2,760 m a.s.l. in the Parco Nazionale del Gran Paradiso (Sindaco et al. [2006]). In Tuscany, the common frog occurs in a much narrower altitudinal range on the Apennines: all the known localities are located between 640 and 1,800 m a.s.l (Vanni and Nistri [2006]). The Grotta di Equi is approximately 350 m a.s.l, and therefore, it is significantly out of the current altitudinal range of R. temporaria in Tuscany. Moreover, R. temporaria does not inhabit the Apuan Alps anymore.
Climatic oscillation and distributional changes
According to Bologna and Balletto ([2007]), within the biogeographically complex context of the Italian batracofauna, it is possible to identify a Euroasiatic component, made of taxa still widespread all over the western part of the Palearctic region. These species took advantage of the cold climate phases of the Pleistocene to colonize the western part of Europe from refugia located in Southern Europe and Asia. Of all these species, which took advantage of colder climates of stadials or glacials to disperse, some have expanded along the Apennine chain, maintaining nowadays relict populations, with the results of a highly discontinuous distribution. R. temporaria is one of these taxa.
In recent years, several researchers focused their attention on the phylogeographic analysis of amphibian species using molecular techniques. R. temporaria was studied to understand phylogenetic relationship between population and range modification related to climate change (see among others, Palo et al. [2004]; Teacher et al. [2009]; Veith et al. [2003]; Zeisset and Beebee [2008]). Southern Europe has always been considered a fundamental glacial refugium, the Iberian, Italian, and Balkan peninsulas in particular. Taxa like R. temporaria have used these refugia as areas from which expand northward: the complex of the European population seems to be composed of just two lineages, a western lineage and an eastern one (Palo et al. [2004]; Teacher et al. [2009]). Stefani et al. ([2012]) specifically analyzed the phylogeographic history and genetic variability of R. temporaria in Italy and identified in the Italian peninsula the principal refugium of the western lineage; in order to explain the complex genetic pattern of Italian population and the differences between the alpine area and the Apennine one, Stefani and co-workers turned to the refugia-within-refugia model (Gómez and Lunt [2006]).
The materials from the Grotta di Equi are dated at about 45 ka (Bigagli et al. [2013]), corresponding to the MIS 3, an interglacial phase. MIS 3 is, however, a relatively cold interglacial phase (among others, Siddall et al. [2008]; Van Meerbeeck et al. [2009]) in which climate should have been much colder than that of other interglacials. The relatively cold conditions of MIS 3 had probably created a particularly propitious opportunity for cold taxa like the common frog to greatly expand their range, moving southward and even at lower altitudes where they cannot be found nowadays.
Abundant fossils of R. temporaria indicating the presence of a population in an area located about 300 m below the current minimum regional lower elevation and dating back to a moderately cold period (about 45 ka) essentially confirms what speculated by Bologna and Balletto ([2007]) about the broadening of the range of frigophilous taxa during glacials or stadial periods. It is therefore expected that in the period of global warming that we are going to face (Walther et al. [2002]; Deutsch et al. [2008]; Laurance et al. [2010]) climate changes will further affect the range of R. temporaria by shrinking it to levels that could approximate those of the warmest past interglacials. Carroll et al. ([2009]), thanks to nearly 70,000 records collected in 10 years, already reported the effect of the increase of temperature on the anticipation of the first spawning dates of R. temporaria, but to our best knowledge, there are no data for this species about distributional changes occurred in the last few decades. A detailed study of fossil remains, coming from precisely dated sites, and therefore testifying the former distribution of this species in relation to climate, would help in better predicting possible future changes. A few useful data are already available for the Iberian Peninsula (Blain [2005], [2009]; Blain et al. [2008]; López-García et al. [2011], [2014]), but more data are need for the Italian Peninsula on which this paper is focused.
Conclusions
The analysis of the fossil amphibians from the Grotta di Equi, hosted in the collections of the Museum of Natural History of the University of Florence, has led to the description, and re-identification of the anurans remains originally cited by De Stefani ([1917]), Battaglia ([1919]), and Branchini ([1928]) as B. vulgaris and Rana sp. More than 360 skeletal elements have been recovered from the cave, and the majority of them belonging to the common frog R. temporaria, five to B. bufo and just one attributed on a morphological, chronological, and geographic basis to cf. S. salamandra. These results, besides improving the knowledge of the fossil herpetofauna from Tuscany (studied since more than a century, see Rook et al. [2013] and references therein) have a much more general impact on the knowledge of the biogeography of one of the species that were identified. The identification of fossil remains of R. temporaria is in fact the most relevant result, as the taxon is not present nowadays in the area of Equi and in the whole Apuan Alps. More in particular, its current distribution in Tuscany, as well as on the whole Apennine chain, is characterized by being discontinuous and restricted to elevations that are about 300 m higher than that of the Grotta di Equi. The remains described here confirm the hypothesis of cycles of dispersion of cold taxa during glacial or stadial phases, and of a contraction during warmer phases (as the current one), and suggest that the current global warming will further affect, negatively, the range of this frigophilous species. Further studies on material of other caves along the Apennine chain could confirm the conclusions of this study and bring new insights regarding the expansion-contraction dynamics of the range according to climatic oscillation.
References
Alexandrino J, Teixeria J, Arntzen JW, Ferrand N: Historical biogeography and conservation of the golden-striped salamander ( Chioglossa lusitanica ) in northwestern Iberia: integrating ecological, phenotypic and phylogeographic data. In Phylogeography of southern European refugia: evolutionary perspectives on the origins and conservation of European biodiversity. Edited by: Weiss S, Ferrand N. Springer, Dordrecht; 2007:189–205. 10.1007/1-4020-4904-8_6
Bailon S (1997) La grenouille rousse (Rana temporaria). Une source de nourriture pour les habitants de Chalain 3. In: Pétrequin P (ed) Les sites littoraux néolithiques de Clairvaux-les-Lacs et de Chalain (Jura). III: Chalain 3, 3200–2900 av. J.-C, 2nd edn. Éditions de la Maison des sciences de l’homme, Paris, pp 711–716
Bailon S: Différenciation ostéologique des anoures (Amphibia, Anura) de France. In Fiches d’ostéologie animale pour l’Archéologie, Série C, Varia. Edited by: Desse J, Desse-Berset N. Centre de Recherches Archéologiques-CNRS, Valbonne; 1999:1–41.
Bartolini S, Delfino M, Cioppi E, Rook L: Late Pleistocene anurans from ‘Grotta di Equi’ (Fivizzano, Apuane Alps, Tuscany, Italy). In Atti IX Congresso Nazionale della Societas Herpetologica Italica, Bari-Conversano, 26–30 September 2012. Edited by: Scillitani G, Liuzzi C, Lorusso L, Mastropasqua F, Ventrella P. Pineta, Conversano (BA); 2013:104–105.
Battaglia R: La grotta preistorica di Equi nelle Alpi Apuane. Natura, Rivista di Scienze Naturali 1919, 10: 49–59.
Bernini F, Razzetti E: Rana temporaria Linnaeus, 1758. In Atlante degli anfibi e dei rettili d’Italia/Atlas of Italian amphibians and reptiles. Edited by: Sindaco R, Bernini F, Doria G, Razzetti E. Polistampa, Firenze; 2006:368–373.
Bigagli C, Iardella R, Palchetti A: Paribeni E (2013) Fivizzano (MS): saggio nella Grotta della Tecchia. Notiziario della Soprintendenza per i Beni Archeologici della Toscana 2013,8(2012):187–193.
Blain H-A: Contribution de la Paléoherpétofaune (Amphibia et Squamata) à la connaissance de l’évolution du climat et du paisaje du Pliocène supérieur au Pléistocène moyen d’Espagne. Ph.D. Dissertation. Muséum national d’Histoire naturelle, Paris; 2005.
Blain H-A: Contribution de la paléoherpétofaune (Amphibia & Squamata) à la connaissance de l’évolution du climat et du paysage du Pliocène supérieur au Pléistocène moyen d’Espagne. Treballs del Museo de Geología de Barcelona 2009, 16: 39–170.
Blain H-A, Bailon S, Cuenca-Bescós G: Los anfibios y escamosos del Pleistoceno inferior-medio de Gran Dolina (Atapuerca, Burgos, España): una perspectiva paleobiogeográfica. Studia Geologica Salmanticensia 2008, 8: 71–84.
Bologna MA, Balletto E: Biogeografia. In Fauna d’Italia, Amphibia. Edited by: Lanza B, Andreone F, Bologna MA, Corti C, Razzetti E. Calderini, Bologna; 2007:47–56.
Bona F, Laurenti B, Delfino M: Climatic fluctuations during the last glacial in the North-Western lombardian prealps: the upper Pleistocene faunal assemblages of the Caverna Generosa (Como, Italy). Riv Ital Paleontol Stratigr 2009,15(2):253–267.
Bonnet RM, Chippindale PT: Speciation, phylogeography and evolution of life history and morphology in plethodontid salamanders of the Eurycea multiplicata complex. Mol Ecol 2004, 13: 1189–1203. 10.1111/j.1365-294X.2004.02130.x
Branchini I: Esame tipologico dei manufatti della Grotta d’Equi (Lunigiana). Atti della Prima Riunione dell’Istituto Italiano di Paleontologia Umana in Archivio per l’Antropologia e la Etnologia 1928,58(1–4):48–63.
Brocchi P: Note sur des ossements de Batraciens recueillis par M. Riviere dans les grottes de Menton. Bull Soc Philomathique Paris 1879, 7: 223–226.
Carroll EA, Sparks TH, Collinson N, Beebee TJC: Influence of temperature on the spatial distribution of first spawning dates of the common frog ( Rana temporaria ) in the UK. Glob Change Biol 2009, 15: 467–473. 10.1111/j.1365-2486.2008.01726.x
Chiari Y, van der Meijden A, Mucedda M, Lourenço JM, Hochkirch A, Veith M: Phylogeography of Sardinian Cave Salamanders (Genus Hydromantes ) is mainly determined by geomorphology. PLoS One 2012,7(3):e32332. doi:10.1371/journal.pone.0032332 10.1371/journal.pone.0032332
Crottini A, Androne F, Kosuch J, Borkin LJ, Litvinchuk SN, Eggert C, Veith M: Fossorial but widespread: the phylogeography of the common spadefoot toad ( Pelobates fuscus ), and the role of the Po Valley as a major source of genetic variability. Mol Ecol 2007, 16: 2734–2754. 10.1111/j.1365-294X.2007.03274.x
De Stefani C: La grotta preistorica di Equi nelle Alpi Apuane. Archivio per l’Antropologia e la Etnologia 1917, 46: 42–82.
De Stefani C: La Grotta di Equi nelle Alpi Apuane. Bullettino di Paletnologia Italiana 1923,43(2–3):123–129.
Delfino M (2002) Erpetofaune italiane del Neogene e del Quaternario. Dissertation. Università di Modena e Reggio Emilia
Delfino M: Herpetological paleodiversity: general considerations about the Italian fossil record of modern species. Ital J Zoolog 2004,71(S2):13–16. 10.1080/11250000409356600
Delfino M, Kotsakis T, Arca M, Tuveri C, Pitruzzella G, Rook L: Agamid lizards from the Plio-Pleistocene of Sardinia (Italy) and an overview of the European fossil record of the family. Geodiversitas 2008, 30: 641–656.
Deutsch CA, Tewksbury JJ, Huey RB, Sheldon KS, Ghalambor CK, Haak DC, Martin PR: Impacts of climate warming on terrestrial ectotherms across latitude. Proc Natl Acad Sci U S A 2008, 105: 6668–6672. 10.1073/pnas.0709472105
Patterns of distribution of amphibians: a global perspective. Johns Hopkins University Press, Baltimore; 1999.
Francis ETB: The anatomy of the salamander. Claredon Press, Oxford; 1934.
Gasc JP, Cabela A, Crnobrnja-Isailovic J, Dolmen D, Grossenbacher K, Haffner P, Lescure J, Martens H, Martínez Rica JP, Maurin H, Oliveira ME, Sofianidou TS, Veith M, Zuiderwijk A (eds) (1997) Atlas of amphibians and reptiles in Europe. Collection Patrimoines Naturels, 29, Societas Europaea Herpetologica. Muséum National d’Histoire Naturelle and Service du Patrimoine Naturel, Paris
Ghezzo E, Palchetti A, Rook L: Recovering data from historical collections: stratigraphic and spatial reconstruction of the carnivoran record from the Late Pleistocene Equi cave (Apuane Alps, Italy). Quat Sci Rev 2014, 96: 168–179. 10.1016/j.quascirev.2014.03.012
Gómez A, Lunt DH: Refugia within refugia: patterns of phylogeographic concordance in the Iberian Peninsula. In Phylogeography of Southern European Refugia. Edited by: Weiss S, Ferrand N. Springer, Dordrecht; 2006:155–188.
Haller-Probst M, Schleich H-H: Vergleichende osteologische Untersuchungen an einigen Urodelen Eurasiens (Amphibia: Urodela, Salamandridae, Proteidae). Courier Forsch-Inst Senckenberg 1994, 173: 23–77.
Kuzmin SL: The amphibians of the former Soviet Union. Pensoft, Sofia; 1999.
Laurance WF, Useche DC, Shoo LP, Herzog SK, Kessler M, Escobar F, Brehm G, Axmacher JC, Chen I-C, Arellano Gámez L, Hietz P, Fiedler K, Pyrcz T, Wolf J, Merkord CL, Cardelús C, Marshall A, Ah-Peng C, Aplet GH, del Coro AM, Baker WJ, Barone J, Brühl CA, Bussmann RW, Cicuzza D, Eilu G, Favila ME, Hemp A, Hemp C, Homeier J, et al.: Global warming, elevational ranges and the vulnerability of tropical biota. Biol Conserv 2010, 144: 548–557. 10.1016/j.biocon.2010.10.010
López-García JM, Blain H-A, Cuenca-Bescós G, Alonso C, Alonso S, Vaquero M: Small vertebrates (Amphibia, Squamata, Mammalia) from the Late Pleistocene-Holocene of Valdavara-1 cave (Galicia, Northwestern Spain). Geophys J Roy Astron Soc 2011, 44: 253–269.
López-García JM, Blain H-A, Bennàsar M, Alcover JA, Bañuls-Cardona S, Fernández-García M, Fontanals M, Martín P, Morales JI, Muñoz L, Pedro M, Vergés JM: Climate and landscape during the Heinrich Event 3 in south-western Europe: the small-vertebrate association from Galls Carboners cave (Mont-ral, Tarragona, north-eastern Iberia). J Quat Sci 2014, 29: 130–140. 10.1002/jqs.2687
Mattoccia M, Marta S, Romano A, Sbordoni V: Phylogeography of an Italian endemic salamander (genus Salamandrina ): glacial refugia, postglacial expansions, and secondary contact. Biol J Linnean Soc 2011, 104: 903–922. 10.1111/j.1095-8312.2011.01747.x
Palo J, Schmeller D, Laurila A, Primmer C, Kuzmin S, Merila J: High degree of population subdivision in a widespread amphibian. Mol Ecol 2004, 13: 2631–2644. 10.1111/j.1365-294X.2004.02269.x
Pitruzzella G: Osteologia e record fossile del genere Salamandrina . Università di Firenze, Dissertation; 2009.
Razzetti E, Zanghellini S, Bernini F: Rana temporaria . In Fauna d’Italia. Edited by: Lanza B, Andreone F, Bologna MA, Corti C, Razzetti E. Calderini, Bologna; 2007:40–47.
Recuero E, Iraola A, Rubio X, Machordom A, García-París M: Mitochondrial differentiation and biogeography of Hyla meridionalis (Anura: Hylidae): an unusual phylogeographical pattern. J Biogeogr 2007, 34: 1207–1219. 10.1111/j.1365-2699.2007.01688.x
Regalia E: Fauna glaciale in due grotte di Equi (Alpi Apuane). Riv It Paleont 1911, 17: 24–28.
Rellini U: La grotta preistorica di Equi nelle Alpi Apuane. Bullettino di Paletnologia Italiana 1918,5(2):67–74.
Riberon A, Miaud C, Grossenbacher K, Taberlet P: Phylogeography of the Alpine salamander, Salamandra atra (Salamandridae) and the influence of the Pleistocene climatic oscillations on population divergence. Mol Ecol 2001, 10: 2555–2560. 10.1046/j.1365-294X.2001.01373.x
Rook L, Croitor R, Delfino M, Ferretti MP, Gallai G, Pavia M: The Upper Valdarno Plio-Pleistocene vertebrate records: an historical overview, with notes on palaeobiology and stratigraphic significance of some important taxa. It J Geosci 2013, 132: 104–125.
Sanchiz B (1998) Salientia. In: Handbuch der Paläoherpetologie-Encyclopedia of Paleoherpetology. part 4. Verlag Friedrich Pfeil, München
Savage MR: The ecology and life history of the common frog (Rana temporaria temporaria). Pitman & Sons Ltd, London; 1961.
Shepard DB, Burbrink FT: Phylogeographic and demographic effects of Pleistocene climatic fluctuations in a montane salamander, Plethodon fourchensis . Mol Ecol 2009, 18: 2243–2262. 10.1111/j.1365-294X.2009.04164.x
Siddall M, Rohling EJ, Thompson WG, Waelbroeck C: MIS 3 sea-level fluctuations: data synthesis and new outlook. Rev Geophys 2008., 46: doi:10.1029/2007RG000226 10.1029/2007RG000226
Sillero N, Campos J, Bonardi A, Corti C, Creemers R, Crochet PA, Isailović JC, Denoël M, Ficetola GF, Gonçalves J, Kuzmin S, Lymberakis P, de Pous P, Rodríguez A, Sindaco R, Speybroeck J, Toxopeus B, Vieites DR, Vences M: Updated distribution and biogeography of amphibians and reptiles of Europe based on a compilation of countrywide mapping studies. Amphib Reptil 2014, 35: 1–31. 10.1163/15685381-00002935
Atlante degli anfibi e dei rettili d’Italia/Atlas of Italian amphibians and reptiles. Polistampa, Firenze; 2006.
Stefani F, Gentilli A, Sacchi R, Razzetti E, Pellitteri-Rosa D, Pupin F, Galli P: Refugia within refugia as a key to disentangle the genetic pattern of a highly variable species: the case of Rana temporaria Linnaeus, 1758 (Anura, Ranidae). Mol Phylogenet Evol 2012, 65: 718–726. 10.1016/j.ympev.2012.07.022
Stöck M, Sicilia A, Belfiore N, Buckley D, Lo Brutto S, Lo Valvo M, Arculeo M: Post-Messinian evolutionary relationships across the Sicilian channel: mitochondrial and nuclear markers link a new green toad from Sicily to African relatives. BMC Evol Biol 2008, 8: 56–74. 10.1186/1471-2148-8-56
Teacher AGF, Garner TWJ, Nichols RA: European phylogeography of the common frog ( Rana temporaria ): routes of postglacial colonization into the British Isles, and evidence for an Irish glacial refugium. Heredity 2009, 102: 490–496. 10.1038/hdy.2008.133
Van Meerbeeck CJ, Renssen H, Roche DM: How did Marine Isotope Stage 3 and Last Glacial Maximum climates differ? - perspectives from equilibrium simulations. Clim Past 2009, 5: 33–51. 10.5194/cp-5-33-2009
Vanni S, Nistri A: Atlante degli anfibi e rettili della Toscana. Regione Toscana. Museo di Storia Naturale dell’Università degli Studi di Firenze, Firenze; 2006.
Veith M, Kosuch J, Vences M: Climatic oscillations triggered post-Messinian speciation of Western Palearctic brown frogs (Amphibia, Ranidae). Mol Phylogenet Evol 2003, 26: 310–327. 10.1016/S1055-7903(02)00324-X
Vences M, Wake DB: Speciation, species boundaries and phylogeography of amphibians. In Amphibian Biology, Systematics. Edited by: Heatwole H, Tyler M. Surrey Beatty and Sons, Chipping Norton; 2007:2613–2671.
Vences M, Grossenbacher K, Puente M, Palanca A, Vieites D: The Cambale’s fairy tale: elevational limits of Rana temporaria (Amphibia: Ranidae) and other European amphibians revisited. Folia Zool 2003,52(2):189–202.
Vences M, Kosuch J, Rödel M-O, Lötters S, Channing A, Glaw F, Böhme W: Phylogeography of Ptychadena mascariensis suggests transoceanic dispersal in widespread African-Malagasy frog lineage. J Biogeogr 2004, 31: 593–601. 10.1046/j.1365-2699.2003.01031.x
Villa A, Andreone F, Boistel R, Delfino M: Skull and lower jaw osteology of the Lanza’s salamander, Salamandra lanzai (Amphibia, Caudata). In Studia Herpetologica. Edited by: Corti C, Capula M. Studies on amphibians and reptiles in honour of Benedetto Lanza. Latina, Belvedere; 2014:171–200.
Walther G-R, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin J-M, Hoegh-Guldberg O, Bairlein F: Ecological responses to recent climate change. Nature 2002, 416: 389–395. 10.1038/416389a
Zeisset I, Beebee TJC: Determination of biogeographical range: an application of molecular phylogeography of the European pool frog Rana lessonae . Proc R Soc B-Biol Sci 2001, 268: 933–938. 10.1098/rspb.2001.1600
Zeisset I, Beebee TJC: Amphibian phylogeography: a model for understanding historical aspects of species distributions. Heredity 2008, 101: 109–119. 10.1038/hdy.2008.30
Acknowledgements
The authors wish to thank the ‘Soprintendenza per i Beni Archeologici della Toscana’ and E. Paribeni, A. Palchetti, C. Bigagli, and R. Iardella for having granted access to the recently excavated fossil material and for the information provided. Roberto Sindaco and the Societas Herpetologica Italica kindly provided the map of the current distribution of the common frog on which Figure 1 is based. A.H. Blain and an anonymous reviewer improved the quality of the manuscript. The authors would also like to acknowledge R.M. Amores, the copyeditor of this article, for her valuable help in editing this paper. A. Villa took the photos of Figure 2. M. Delfino was supported by Università di Torino (Fondi di Ricerca Locale Università di Torino 2012, 2013), Spanish Ministerio de Economia y Competitividad (CGL2011-28681), and Generalitat de Catalunya (2014 SGR 416 GRC).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
EC and LR manage the project aimed to the re-evaluation and valorization of the Equi historical Palaeontological collections of the Museum of Natural History (Geology and Paleontology Section), University of Florence. MD identified the material. SB, MD, LR, and EC wrote the manuscript. All authors read and approved the final manuscript.
Authors’ original submitted files for images
Below are the links to the authors’ original submitted files for images.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made.
The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this licence, visit https://creativecommons.org/licenses/by/4.0/.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Bartolini, S., Cioppi, E., Rook, L. et al. Late Pleistocene fossils and the future distribution of Rana temporaria (Amphibia, Anura) along the Apennine Peninsula (Italy). Zool. Stud. 53, 76 (2014). https://doi.org/10.1186/s40555-014-0076-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40555-014-0076-5