
Abstract
Background
Jujube (Ziziphus jujuba Mill.) fruit is one of the largest productions in China and its increasing production has drawn considerable attention from researchers. Polysaccharide is one of the most abundant components of jujube, and it represents a major group of biolotegically active constituents. This study intended to investigate the special structure of a homogeneous acidic polysaccharide (PZMP4) produced from Ziziphus Jujuba cv. Muzao fruit using novel methods, including DEAE-Sepharose Fast Flow and Sephacryl S-300 column chromatography.
Results
The structure of PZMP4 was determined via high-performance gel permeation chromatography (HPGPC), gas chromatography (GC), Fourier transform infrared spectroscopy (FT-IR), methylation analysis, nuclear magnetic resonance spectroscopy (NMR), scanning electron microscopy (SEM), and atomic force microscopy (AFM). The results reveal that PZMP4 with a molecular weight of 27.90 kDa was composed of rhamnose, arabinose, mannose, glucose, galactose, and galacturonic acid at a ratio of 2.32:2.21:0.22:0.88:2.08:8.83. Advanced structural analysis revealed a netted structure with molecular aggregates of PZMP4. Structural features demonstrated that the basic backbone of PZMP4 appeared to mainly consist of (1→4)-linked GalpA with three branches bonded to O-3 of (1→3)-linked Araf, (1→2)-linked Rhap, and terminated with GalpA.
Conclusions
PZMP4’s unique structure could imply distinct bioactivities and have considerable utilization in functional food.
Graphic abstract

Highlights
-
Structural characteristics of PZMP4 were analyzed by HPGPC, GC, FT-IR, GC-MS, NMR, SEM and AFM.
-
PZMP4 mainly consisted of (1→4)-linked GalpA with three branches boned to O-3.
-
The branches included (1→3)-linked Araf, (1→2)-linked Rhap, and terminated with GalpA.
Similar content being viewed by others


Background
Jujube, (Ziziphus jujuba Mill.), a member of the family Rhamnaceae, is widely used as a medicinal herb. It is predominant in subtropical Asian and European countries, particularly in China [1, 2]. The Z. jujuba fruit is nutrient-dense, with carbohydrates accounting for the majority (55–85%). It also has moisture (25–30%), crude protein (2.9–6.6%), crude fiber (2.4–8.4%), crude fat (0.4–1.0%), and several other vitamins and minerals [3]. Furthermore, numerous essential functional components present in jujube, including polysaccharides, flavonoids, saponins, and cyclic nucleotides, have been demonstrated to account for a variety of biological activities, such as anti-inflammatory, anti-cancer, and anti-oxidant activities, as well as hematopoietic function, neurobeneficial properties, and intestinal flora regulation [4].
Of the various functional components in Z. jujuba fruit, polysaccharide is especially important because of its bioactivities and large cellular concentrations. It is mainly composed of different ratios of monosaccharides and glycosidic bonds [5]. The activities of polysaccharides of Z. jujuba are determined by their molecular weights and chemical structures. An increased galacturonic acid concentration could result in enhancing antioxidant activity [6]. The structural–physicochemical properties and bioactivities of polysaccharides vary greatly among different Z. jujuba varieties [1]. Our research efforts have contributed to a better understanding of the structural basis of jujube polysaccharides [7,8,9].
In this present study, an acidic polysaccharide (PZMP4) was isolated from Ziziphus Jujuba cv. Muzao and characterized by HPGPC, GC, AFM, SEM, NMR spectroscopy, FT-IR spectroscopy, and methylation analysis.
Materials and methods
Materials
The Z. Jujuba cv. Muzao fruits were supplied by the Jia County of Shaanxi. Sephacryl S-300 and DEAE-Sepharose Fast Flow cellulose were provided by GE Healthcare Life Sciences. Standard monosaccharides were obtained from Sigma Chemical Co. All additional chemicals utilized in the experiments were of analytical grade.
Polysaccharide isolation
The production of crude polysaccharide from Z. Jujuba cv. Muzao (ZMP) from jujube fruit at the red and ripened stage was performed as previously described [10, 11]. After re-dissolution, the ZMP was added to a DEAE-Sepharose FF column equalized with 0.4 M NaCl and a Sephacryl S-300 column balanced with distilled water. It was then gathered, concentrated, and lyophilized for the production of designative PZMP4 purified polysaccharides [7].
General methods
The carbohydrate content was determined by the phenol–sulfuric acid method with glucose as the standard [12]. The Bradford method with bovine serum albumin as the reference was used to assess the protein content [13]. The Folin–Ciocalteu colorimetric method was used to quantify the value of the total phenol content [14].
To identify and quantify PZMP4 monosaccharide, GC analysis was performed as reported previously [15]. HPGPC on an Agilent-LC 1200 instrument equipped with a TSK-gel G3000PWxl (7.8 mm × 300 mm) column was used to analyze the homogeneity and average molecular weight of PZMP4 [7, 16].
The IR spectra of PZMP4 were obtained by the KBr disc method, with 400–4000 cm−1 range for the FT-IR spectrometer. The one- and two-dimensional NMR spectra of PZMP4 were acquired with a Bruker AVIII-600 NMR spectrometer [8].
We first performed carboxyl reduction with 1-ethyl-3-(3-dimethylaminopropyl) carbodiimide and NaBD4, and then further performed hydrolysis with trifluoroacetic acid, reduction with NaBH4, and acetylation with acetic anhydride. The resultant partially methylated alditol acetate derivatives were identified by their retention times and electron ionization spectra on a GCMS-QP2010A instrument equipped with a DB-17MS capillary column (60.0 m × 0.25 mm × 0.25 μm) [8].
The surface morphology of PZMP4 was examined using an S-4800 SEM (Japan) under 10 kV accelerating voltage. PZMP4 was dissolved in distilled water, dropped on the surface of a mica carrier, and then dried at 70 °C under ambient pressure [8]. The AFM images were taken with an Agilent 5500 atomic force microscope (USA) in tapping mode [7].
Results and discussion
Preliminary PZMP4 characterization
ZMP was isolated from Z. Jujuba cv. Muzao fruit and extracted by ultrasonic-assisted extraction, ethanol precipitation, deproteination, dialysis, and lyophilization. Further purification was subsequently done using a DEAE Sepharose Fast Flow column (2.6 cm × 100 cm), which was eluted with phosphate-buffered saline (20 mM, pH 6.0), and 0.4 M NaCl solution at a flow rate of 1.5 mL/min. We determined the elution by observing phenol–sulfuric acid, and then passed it through a Sephacryl S-300 column (2.6 cm × 100 cm) with deionized water for further purification. As a result, a single elution peak named PZMP4 was obtained. The total carbohydrate content of PZMP4 was found to be 92.64%. The protein and total phenol contents of PZMP4 were 3.09%, and 0.95%, respectively, higher than the acidic (PZMP2-2) polysaccharide from Z. Jujuba cv. Muzao [8].
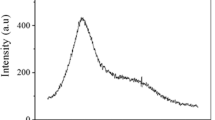
As shown in Fig. 1A, HPGPC revealed that the acidic polysaccharide PZMP4 was homogeneous, with only one symmetrical absorption peak. A standard curve of the logarithm of relative molecular weight associated with elution time (t) was created using a serious detraining, shown as the following: lg Mw = − 0.3259t + 10.9495 (R2 = 99.57%). On the basis of the equation, the average molecular weight of PZMP4 was estimated to be 27.90 kDa with a retention time of 19.983 min. This acidic jujube polysaccharide fraction had a molecular weight similar to HJP-4 (Z. Jujuba cv. Hamidazao polysaccharide) [1].

The physicochemical analysis of PZMP4: A HPGPC spectrum, and B monosaccharide spectrum, scanning electron microscopy (C ×1000, D ×5000, E ×10,000)
GC was used to determine the monosaccharide component of PZMP4. The hydrolysate of PZMP4 consisted of six monosaccharides, including rhamnose, arabinose, mannose, glucose, galactose, and galacturonic acid (Fig. 1B). Their ratios were 2.32:2.21:0.22:0.88:2.08:8.83, suggesting that PZMP4 was an acidic heteropolysaccharide. Furthermore, the results indicated that rhamnose, arabinose, and galacturonic acid accounted for the majority of the total polysaccharide content. However, the result was different for Z. Jujuba cv. Hamidazao polysaccharide fractions; this may be caused by different raw materials, as well as extraction and purification methods [1, 5].
FT-IR spectrum analysis
The functional groups and chemical bonds of PZMP4 were further analyzed by FT-IR (Additional file 1: Fig. S1A). The O–H-stretching vibration and the C–H-stretching vibration were represented by distinct bands in the 3411 and 2937 cm−1 regions, respectively. The absorbances at 1741 and 1244 cm−1 demonstrated the existence of uronic acid, which corroborated the results of the uronic acid assay [8, 17]. The symmetrical C=O-stretching vibrations were verified by the high peak at 1610 cm−1 [18]. The 1415 cm−1 peak represented the characteristic absorption of C-H bands, while the 1099 cm−1 peak indicated the pyranose form’s C–O-stretching vibrations [8]. The slight characteristic absorptions at 800–900 cm−1 may indicate the existence of α- and β-configurations [19]. Consequently, FT-IR analysis of PZMP4 revealed absorption peaks of typical plant polysaccharides.
Methylation analysis
PZMP4 was methylated and reacted with trifluoroacetic acid. The resultant partly methylated alditol acetates were examined by gas chromatography–mass spectrometry (GC–MS). Table 1 shows the linkage types and molar ratio percentages of PZMP4 according to the Complex Carbohydrate Research Center (CCRC) Spectral Database for PMAA. PZMP4 contained four main glycosidic linkages, 1,2-linked Rhap, 1-linked GalpA, 1,3-linked Araf, and 1,3,4-linked GalpA with molar percentage ratios of 1.64:1.00:1.52:4.50, thus, implying that the PZMP4’s backbone could only be of 1,4-linked GalpA residues, with branches connected to the O-3 position for some residues. The structure of PZMP4 was further supported by NMR spectroscopy.
NMR analysis
To determine the detailed structure of PZMP4, one-dimensional and two-dimensional NMR spectra were used for further study. The C/H chemical shifts of several glycosidic bonds were consistent with the previous literature; the data are shown in Fig. 2 and Table 2. The 1H-NMR spectrum of PZMP4 (Fig. 2A) displays four main anomeric proton signals at δ 4.87, 5.00, 4.82, and 5.00/5.05, which were designated as A, B, C, and D, respectively. H-2, H-3, H-4, and H-5 of 1,3,4-linked GalpA residues were responsible for the significant peaks in the 3.74–5.06 ppm range. The 13C NMR spectrum (Fig. 2B) revealed six anomeric signals resonating at 101.89, 70.79/70.93, 79.42, 84.05, 73.41/73.04, and 173.68/173.80 ppm. According to previous results in the literature, the relevant anomeric carbon signals of tagged residues in the 1H and 13C NMR spectra were attributed to data in the 2D NMR spectra [20, 21]. From the chemical shift data in the COSY (Additional file 2: Fig. S2B), NOESY (Additional file 2: Fig. S2C), 1H/1H TOCSY (Additional file 2: Fig. S2D), HSQC (Additional file 2: Fig. S2E), and HMBC spectra (Additional file 2: Fig. S2F), the proton and carbon assignments of four main residues in PZMP4 are presented in Table 2.

The NMR spectra of PZMP4 in D2O. A 1H spectra; B 13C spectra
The signal at δH 5.00 was determined from the chemical shift of the anomeric proton of residue B. The equivalent signal in the anomeric carbon was δC 102.36. The δC 71.45/δH 3.91, δC 70.79/δH 3.63 (3.66), δC 71.59/δH 3.96, δC 74.21/δH 4.32, and δC 178.21 signals were allocated to C-2, C-3, C-4, C-, and C-6 of residue B, respectively [22, 23]. The anomeric proton of residue C had a chemical shift of 4.82 ppm, whereas the anomeric carbon had a chemical shift of 102.36 ppm. The COSY and TOCSY spectra were used to identify the other protons in residue C. According to HSQC, the other comparable carbon and hydrogen signals were 84.19 (4.11/4.04), 71.45 (3.86), 74.85 (3.66), 71.60 (3.69), and 19.44 (1.15) ppm. According to the NMR data, the chemical shifts of this residue were identical to that of α-1,2-linked Rhap [24, 25]. HSQC indicated the other carbon and hydrogen signals with 110.24 (5.00/5.05), 81.61 (4.20), 86.77 (3.94), 83.74 (4.02), and 63.96 (3.62) ppm. From the NMR data, the chemical shifts of this residue were identical to that of α-1,3-linked Galp [26, 27].
HMBC, COSY, and NOESY could determine the glycosidic linkages between sugar residues. Hence, with these techniques, the intra-residue connections were determined and are listed in Table 2. As the HMBC spectrum shows, some inter-residual cross-peaks were identified: A C-3 to D H-1, A C-4 to D H-1, A C-4 to B H-1, D C-3 to D H-1, A C-4 to D H-3, A C-3 to C H-2, and A C-1 to D H-1. In addition, in the COSY spectrum, certain inter-residual cross-peaks were also recognized: C/D H-1 to C/D H-2, A/B H-2 to A/B H-3, A/B/C/D H-3 to A/B/C/D H-4, and C/D H-4 to C/D H-5. A/C H-1 to A H-3 were detected in the NOESY spectrum [28].
According to the monosaccharide composition of PZMP4, combined with the results of FT-IR, GC–MS, 1D and 2D NMR analyses, it was determined that PZMP4 was mainly composed of →4)-GalpA-(1→ backbone, with a branching point at the O-3 position consisting of Araf, Rhap, and GalpA residues.
Morphological properties
Different morphological properties are key components that contribute to the complexity of polysaccharide forms. SEM, as a microscopic-molecular-morphology observation technique, is frequently used to characterize the surface morphology of polysaccharides [29]. The SEM images of PZMP4 demonstrated the disparate morphological characteristics (Fig. 1C–E). PZMP4 had a rough surface and a netted structure due to molecular aggregation, which was most likely induced by a variety of hydroxyl and carboxyl groups [30].
Another important tool is AFM, which could provide both 2D images and 3D surface images of plant polysaccharides. Additional file 1: Figure S1B, C depicted the AFM findings of PZMP4. We observed an irregular special block structure for PMP4 indicating the state of aggregation, which was compatible with the SEM images. Higher hydroxyl and carbonyl group contents of plant polysaccharide chains appeared to strengthen the intermolecular and intramolecular interactions, and thereby resulted in the strong molecular chain aggregation [31]. The molecules of PZMP4 had a tendency to form a stacked and aggregated structure to be stable.
Conclusions
A new acidic polysaccharide (PZMP4) from Ziziphus Jujuba cv. Muzao fruit was successfully purified and characterized. PZMP4 was a homogeneous polysaccharide with a molecular weight of 27.90 kDa. It was mainly composed of rhamnose, arabinose, mannose, glucose, galactose, and galacturonic acid at a molar ratio of 2.32:2.21:0.22:0.88:2.08:8.83. Methylation and NMR spectroscopy suggested that the main backbone of PZMP4 was composed of (1→4)-linked GalpA, with three branches attached to O-3 of (1→3)-linked Araf, (1→2)-linked Rhap, and terminated with GalpA. Notably, the unique structure of PZMP4 indicated that it could possess valuable bioactivities and have wide applications in food industries.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- HPGPC:
-
High-performance gel permeation chromatography
- GC:
-
Gas chromatography
- FT-IR:
-
Fourier transform infrared spectroscopy
- NMR:
-
Nuclear magnetic resonance spectroscopy
- SEM:
-
Scanning electron microscopy
- AFM:
-
Atomic force microscopy
- ZMP:
-
Crude polysaccharide from Zizyphus Jujuba cv. Muzao
- TA:
-
Trifluoroacetic acid
- GC–MS:
-
Gas chromatography–mass spectrometry
References
Yang Y, Qiu Z, Li L, Vidyarthi SK, Zheng Z, Zhang R. Structural characterization and antioxidant activities of one neutral polysaccharide and three acid polysaccharides from Ziziphus jujuba cv. Hamidazao: a comparison. Carbohydr Polym. 2021;261:117879.
Ji XL, Hou CY, Gao YG, Xue YQ, Yan YZ, Guo XD. Metagenomic analysis of gut microbiota modulatory effects of jujube (Ziziphus jujuba Mill.) polysaccharides in a colorectal cancer mouse model. Food Funct. 2020;11:163–73.
Gao QH, Wu CS, Wang M. The jujube (Ziziphus jujuba Mill.) fruit: a review of current knowledge of fruit composition and health benefits. J Agric Food Chem. 2013;61:3351–63.
Chen J, Tsim KWK. A review of edible jujube, the Ziziphus jujuba fruit: a heath food supplement for anemia prevalence. Front Pharmacol. 2020;11:593655.
Ji X, Peng Q, Yuan Y, Shen J, Xie X, Wang M. Isolation, structures and bioactivities of the polysaccharides from jujube fruit (Ziziphus jujuba Mill.): a review. Food Chem. 2017;227:349–57.
Chang SC, Hsu BY, Chen BH. Structural characterization of polysaccharides from Zizyphus jujuba and evaluation of antioxidant activity. Int J Biol Macromol. 2010;47:445–53.
Ji X, Liu F, Peng Q, Wang M. Purification, structural characterization, and hypolipidemic effects of a neutral polysaccharide from Ziziphus Jujuba cv. Muzao. Food Chem. 2018;245:1124–30.
Ji X, Zhang F, Zhang R, Liu F, Peng Q, Wang M. An acidic polysaccharide from Ziziphus Jujuba cv. Muzao: purification and structural characterization. Food Chem. 2019;274:494–9.
Ji X, Yan Y, Hou C, Shi M, Liu Y. Structural characterization of a galacturonic acid-rich polysaccharide from Ziziphus Jujuba cv. Muzao. Int J Biol Macromol. 2020;147:844–52.
Ji X, Peng Q, Li H, Liu F, Wang M. Chemical characterization and antiinflammatory activity of polysaccharides from Zizyphus jujube cv. Muzao. Int J Food Eng. 2017. https://doi.org/10.1515/ijfe-2016-0382.
Qin Z, Liu HM, Cheng XC, Wang XD. Effect of drying pretreatment methods on structure and properties of pectins extracted from Chinese quince fruit. Int J Biol Macromol. 2019;137:801–8.
Dubois M, Gilles KA, Harmilton JK, Rebers PA, Smith F. Colorimetric method for determination of sugars and related substances. Anal Chem. 1956;28:350–6.
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein binding. Anal Biochem. 1976;72:248–54.
Li S, Shah NP. Characterization, antioxidative and bifidogenic effects of polysaccharides from Pleurotus eryngii after heat treatments. Food Chem. 2016;197:240–9.
Wang X, Zhang L, Wu J, Xu W, Wang X, Lu X. Improvement of simultaneous determination of neutral monosaccharides and uronic acids by gas chromatography. Food Chem. 2017;220:198–207.
Liu XX, Liu HM, Yan YY, Fan LY, Qin GY. Structural characterization and antioxidant activity of polysaccharides extracted from jujube using subcritical water. LWT Food Sci Technol. 2019;117:108645.
Chen Y, Xie MY, Nie SP, Li C, Wang YX. Purification, composition analysis and antioxidant activity of a polysaccharide from the fruiting bodies of Ganoderma atrum. Food Chem. 2008;107:231–41.
Vasilieva T, Sigarev A, Kosyakov D, Ul’yanovskii N, Anikeenko E, Chuhchin D. Formation of low molecular weight oligomers from chitin and chitosan stimulated by plasma-assisted processes. Carbohydr Polym. 2017;163:54–61.
Ren G, Xu L, Lu T, Yin J. Structural characterization and antiviral activity of lentinan from Lentinus edodes mycelia against infectious hematopoietic necrosis virus. Int J Biol Macromol. 2018;115:1202–10.
Zhang XH, Liu LN, Lin CW. Structural features, antioxidant and immunological activity of a new polysaccharide (SP1) from sisal residue. Int J Biol Macromol. 2013;59:184–91.
Zhan R, Xia L, Shao JH, Wang C, Chen DF. Polysaccharide isolated from Chinese jujube fruit (Zizyphus jujuba cv. Junzao) exerts anti-inflammatory effects through MAPK signaling. J Funct Foods. 2018;40:461–70.
Hu HB, Liang HP, Wu Y. Isolation, purification and structural characterization of polysaccharide from Acanthopanax brachypus. Carbohydr Polym. 2015;127:94–100.
Zhao ZH, Dai H, Wu XM, Chang HT, Gao XM, Liu MJ. Characterization of a pectic polysaccharide from the fruit of Ziziphus jujuba. Chem Nat Compd. 2007;43(4):374–6.
Li N, Liu X, He XX, Wang SY, Cao SJ, Xia Z. Structure and anticoagulant property of a sulfated polysaccharide isolated from the green seaweed Monostroma angicava. Carbohydr Polym. 2017;159:195–206.
Ji XL, Hou CY, Shi MK, Yan YZ, Liu YQ. An insight into the research concerning Panax ginseng CA Meyer polysaccharides: a review. Food Rev Int. 2020. https://doi.org/10.1080/87559129.2020.1771363.
Cardoso SM, Silva AMS, Coimbra MA. Structural characterisation of the olive pomace pectic polysaccharide arabinan side chains. Carbohydr Res. 2002;337:917–24.
Ji XL, Peng BX, Ding HH, Cui BB, Nie H, Yan YZ. Purification, structure and biological activity of pumpkin (Cucurbita moschata) polysaccharides: a review. Food Rev Int. 2021. https://doi.org/10.1080/87559129.2021.1904973.
Hou CY, Yin MS, Lan P, Wang HR, Nie H, Ji XL. Recent progress in the research of Angelica sinensis (Oliv.) Diels polysaccharides: extraction, purification, structure and bioactivities. Chem Biol Technol Agric. 2021;8:13.
Xu Y, Liu G, Yu Z, Song X, Li X, Yang Y. Purification, characterization and antiglycation activity of a novel polysaccharide from black currant. Food Chem. 2016;199:694–701.
Ji X, Hou C, Yan Y, Shi M, Liu Y. Comparison of structural characterization and antioxidant activity of polysaccharides from jujube (Ziziphus jujuba Mill.) fruit. Int J Biol Macromol. 2020;149:1008–18.
Liu W, Wang H, Yu J, Liu Y, Lu W, Chai Y. Structure, chain conformation, and immunomodulatory activity of the polysaccharide purified from Bacillus Calmette Guerin formulation. Carbohydr Polym. 2016;150:149–58.
Acknowledgements
We are grateful to the platform of College of Food and Bioengineering, Zhengzhou University of Light Industry.
Funding
This work was funded by the Natural Science Foundation of Henan Province (212300410297), Basic Research Plan of Higher Education School Key Scientific Research Project of Henan Province (21A550014), Doctoral Research Foundation of Zhengzhou University of Light Industry (2020BSJJ015), Fourth Chinese Materia Medica Resources Survey (Z135080000022), and Key Research and Development Program of Hebei Province (20370509D).
Author information
Authors and Affiliations
Contributions
Conceptualization and methodology: JX; software and validation: CY; investigation, resources, and data curation: TJ; writing original draft preparation: ZS; writing review and editing: JY; visualization and supervision: JX and SM. All authors have agreed to the published version of the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1
: Figure S1. The physicochemical analysis of PZMP4: (A) FI-IR spectrum, (B) Atomic force microscope planar image, (C) Three-dimensional image of atomic force microscope.
Additional file 2
: Figure S2. The NMR spectra of PZMP4 in D2O. (A) DEPT-135 spectra, (B) COSY spectra, (C) NOESY spectra, (D) 1H/1H TOCSY spectra, (E) HSQC spectra, (F) HMBC spectra.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ji, X., Cheng, Y., Tian, J. et al. Structural characterization of polysaccharide from jujube (Ziziphus jujuba Mill.) fruit. Chem. Biol. Technol. Agric. 8, 54 (2021). https://doi.org/10.1186/s40538-021-00255-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40538-021-00255-2