
Abstract
Background
Wildlife conservation often focuses on establishing protected areas. However, these conservation zones are frequently established without adequate knowledge of the movement patterns of the species they are designed to protect. Understanding movement and foraging patterns of species in dynamic and diverse habitats can allow managers to develop more effective conservation plans. Threatened lemurs in Madagascar are an example where management plans and protected areas are typically created to encompass large, extant forests rather than consider the overall resource needs of the target species.
Methods
To gain an understanding of golden-crowned sifaka (Propithecus tattersalli) movement patterns, including space use and habitat selection across their range of inhabited forest types, we combined behavior data with Dynamic Brownian Bridge Movement Models and Resource Selection Functions. We also examined the influence of abiotic, biotic, and anthropogenic factors on home range size, movement rates, and foraging patterns.
Results
We found that home range size and movement rates differed between seasons, with increased core area size and movement in the rainy season. Forest type also played a role in foraging behavior with sifaka groups in the humid forest avoiding roads in both seasons, groups in the dry deciduous forest avoiding road networks in the rainy season, and groups in the moderate evergreen forest displaying no selection or avoidance of road networks while foraging.
Conclusion
Our study illustrates the importance of studying primate groups across seasons and forest types, as developing conservation plans from a single snapshot can give an inaccurate assessment of their natural behavior and resources needs of the species. More specifically, by understanding how forest type influences golden-crowned sifaka movement and foraging behavior, conservation management plans can be made to the individual forest types inhabited (dry deciduous, moderate evergreen, humid, littoral, etc.), rather than the region as a whole.
Similar content being viewed by others

Background
Conservation biologists have long recognized the importance of establishing protected areas to facilitate population persistence of wildlife in landscapes that are threatened by increasing human encroachment, habitat fragmentation, and habitat loss [1,2,3,4]. However, efforts to conserve wildlife and preserve biodiversity often are based on an incomplete understanding of animal movement as well as variability in movement patterns among groups or populations that the areas are meant to protect [5]. While a number of studies have demonstrated the relevance of incorporating movement, particularly animal foraging and home range size, into protected area design [6,7,8,9], integration between the disciplines of conservation biology and movement (coined “conservation behavior”) is limited [10, 11]. Yet, knowledge of movement behavior, specifically how, when, and where animals move and forage within their habitat, would illuminate how populations navigate and utilize resources within their environment and thus develop better management plans [12, 13]. Specifically, species, populations, or even groups often respond differently to factors such as seasonality, habitat characteristics, and anthropogenic pressures and therefore a better understanding of their role is crucial when developing management plans and establishing protected areas.
In many tropical regions, seasons are often divided into dry and rainy, with primary productivity varying seasonally as a function of rainfall. This seasonality thus influences the distribution and availability of resources on the landscape and as a result animal movement strategies shift to increase foraging efficiency [14,15,16]. For example, the black-fronted titi monkey (Callicebus nigrifons) [17] and collared brown lemur (Eulemur collaris) [18] cope with dry season food shortages by reducing movement rates, while the common bumble bee (Bombus vosnesenskii) [19] and African elephant (Loxodonta africana) [20], respond by increasing foraging and movement rates. Thus, animals are coping with dry season conditions by shifting home range size or location [21] and altering time spent foraging [22]. Understanding how seasonal fluctuations influence movement and foraging patterns in free-living animals can allow managers to more effectively design protected areas and protect critical resources [23].
In addition to abiotic factors, biotic factors such as habitat type, strongly influence animal movement and foraging [24]. Various studies demonstrate that animals adjust their home range size and foraging patterns in response to habitat type and structure (e.g. roe deer (Capreolus capreolus) [25] and coyote (Canis latrans) [26]); indicating that landscape heterogeneity is a key factor influencing the movement of species. While studies of canids, ungulates, and primates have examined the influence of habitat type on home range size, a large proportion of studies are limited to examining metrics of habitat structure (e.g., forest maturity, vegetation density, food scarcity, microhabitat preference) on animal movement and home range size [27, 28]. The benefit of understanding movement behavior across distinct habitat types is that management strategies can be designed for each habitat type a species occupies.
Importantly, anthropogenic influences affecting animal movement behaviors can have deleterious effects on wildlife, and must be considered when establishing protected areas [29, 30]. The presence of human developments and road networks may negatively influence animal movement behavior by increasing human-wildlife interactions (e.g., hunting, poaching, vehicle collisions) and pushing animals out of prime habitat [31, 32]. Large mammals may be especially affected by human encroachment due to their larger home range size, lower population density, more narrow geographic distributions, and large portions of their distributions being shared with humans [33]. For instance, black bears (Ursus americanus) have been found to avoid areas with human development during daylight hours [34] and woodland caribou (Rangifer tarandus caribou) avoid high use roads, mines, and cabins during months of high human activity [35]. Few studies have examined the influence of human infrastructure on primate movement, although they often are strongly affected by anthropogenic activities [36, 37].
The lemurs of Madagascar face significant anthropogenic threats [38]. Between 1953 and 2014, Madagascar lost 44% of its forests due to clearing for slash and burn agriculture, resulting in 46% of the remaining forests being located within 100 m of a forest edge [39]. This high degree of forest destruction and increasing presence of edge forest habitat has influenced lemur behavior and their ability to meet nutritional demands. While our understanding of lemur movement is limited, a few studies have examined lemur home range size [40, 41], dietary flexibility [42], species abundance [43], and reproduction in various forest types [44, 45]. Within the genus Propithecus, groups of diademed sifakas (P. diadema) in humid fragmented habitats had reduced home range size and daily path length and foraged on sub-optimal food items compared to sifaka groups in contiguous forest environments [40]. In contrast, groups of Milne-Edwards’ sifakas (P. edwardsi) inhabiting humid logged forests traveled shorter distances each day to feed in a low-quality food environment, yet maintained larger home ranges than groups in contiguous forests [41]. Further, groups of Verreaux’s sifakas (P. verreauxi) in Madagascar’s dry deciduous forests exhibited significant home range reduction from the rainy to the dry season [46]. While these studies have shed light on Propithecus behavioral responses to abiotic and biotic factors in extremes of the humid-dry forest gradient of forest types, we do not understand how species in the genus Propithecus respond in a moderate forest type. Knowledge of Propithecus movement behavior in regards to these factors would enable us to predict how these lemurs would adapt to changes in forest habitat and design a reserve accordingly.
Golden-crowned sifaka (Propithecus tattersalli) are a critically endangered lemur endemic to naturally fragmented forests of northeastern Madagascar [47]. Unlike the other eight species of sifaka (Propithecus spp.) on Madagascar that are restricted to dry or humid forest types, P. tattersalli inhabit a range of forest types [48, 49]. Variation of forest types they inhabit makes them a unique opportunity to examine the influence of seasonality, forest type, and anthropogenic factors on movement and foraging behavior in a primate. Studies of golden-crowned sifaka have documented a major decline in the overall population in the last decade and studies have informed researchers of the species selective use of forests in a naturally fragmented landscape. However, no studies have examined the influence of movement on space use and foraging tree selection across their range [50, 51]. An understanding of how abiotic, biotic, and anthropogenic factors influence golden-crowned sifaka space use and foraging throughout their range would allow unique management plans to be developed for distinct populations within each particular forest type occupied rather than the species as a whole.
In this study, we analyzed location and foraging behavior of six golden-crowned sifaka groups to evaluate the effects of abiotic (seasonality: rainy and dry season), biotic (forest type: dry deciduous, moderate evergreen, and humid forests), and anthropogenic (disturbance: edge and interior forests) factors on their movement patterns and space use. Approaches to studying nonhuman primate space use typically are limited to examining daily path length and home range overlap through the use of area estimators (minimum convex polygon, line-based kernel density, etc.) [52, 53]. More modern and sophisticated approaches such as Dynamic Brownian Bridge movement models (DBBMM) and Bayesian methods [54, 55] reduce the likelihood of both Type I and Type II errors which can bias our understanding of animal space use and habitat selection [56]. By selecting DBBMM to estimate space use we were able to incorporate both temporal and behavioral characteristics of movement trajectories into estimation of an animal’s home range [57]. To test our first objective, we predicted that seasonal movement rates would be greater in the rainy season, when we expected that sifakas would search out energy rich but spatially limited resources (i.e. fruits). In contrast, we expected that sifakas in the dry season would be conserving limited energy resources and thus restrict their movements. We also expected greater movement rates in more extreme forest types (dry and humid forests) and edge forests compared to moderate evergreen and interior forests for similar energetic reasons [58], as well as differences in sifakas densities between the three forest types [59]. Second, we predicted that home range size and core area range size would be larger in the rainy season compared to the dry season, as sifakas would maximize their foraging area to exploit energy rich, rainy season resources. We also expected home range and core areas to be larger along forest edges, where energy-rich resources may be more limited [58]. Additionally, we predicted that home range and core areas sizes would be larger in the dry and humid forests where golden-crowned sifaka densities are lower, and smaller in the moderate forests where golden-crowned sifaka densities are higher [59]. Third, we predicted that sifakas would select foraging locations with the largest feeding trees within their home ranges, which we expected to have the most available fruits [60], and avoid locations near human settlements or man-made structures, where we expected forest disturbance to be greatest [61].
Methods
Study area
Research was conducted in the Loky-Manambato Protected Area (49°56ʹE, 13°31ʹS) of northeastern Madagascar (Fig. 1). This protected area encompasses a unique biogeographical transition zone from Madagascar’s northern and western dry deciduous forests to southern humid forests. The Loky-Manambato region contains a mosaic of various forest types including dry deciduous, dry evergreen, humid, and littoral forests separated by agricultural areas and savanna [50]. The total forest cover of this protected area is 475.3 km2 and individual forest fragments range from 11.6 to 46.3 km2. The region experiences a 4-month rainy season from December to March followed by an 8-month dry season [62]. The study sites include three distinct forest types: humid forest, moderate evergreen forest, and dry deciduous forest.

Map of the golden-crowned sifaka (Propithecus tattersalli) range within the Loky-Manambato Protected Area in northeastern Madagascar, as indicated in the box on the inset of Madagascar. Different shades of green indicate the three main forest types and hatched black lines indicate the three forest fragments surveyed: dry (light green, Solaniampilana), moderate (green, Bekaroaka), and wet (dark green, Binara). The thin orange line depicts the unpaved national road in the region
Study species and subjects
Golden-crowned sifaka live in semi-cohesive social groups ranging in size from 3 to 12 individuals with one or more adult males, several adult females, and several immature individuals of both sexes. Group members typically travel in a coordinated fashion and generally remain in visual or auditory contact with at least one other group member [62]. Thus, we assume that all animals within a given social group share a home range, and therefore treated each group as a distinct unit of analysis. Golden-crowned sifaka are frugo-folivores, but also consume seeds, petioles, buds, flowers, and bark.
We studied six groups of golden-crowned sifaka distributed across the three distinct forest types (two groups each in dry deciduous, moderate evergreen, and humid forest) in the Loky-Manambato Protected Area. Prior to data collection, all groups were habituated to human presence. Habituation was considered complete when lemurs no longer alarm called, fled from human presence, or moved closer to observers out of curiosity. Similar to studies conducted with other lemur species, this process took less than 2 months [63]. We selected three of the 11 large forest fragments containing golden-crowned sifaka due to their accessibility: Solaniampilana (dry deciduous), Bekaraoka (moderate evergreen), and Binara (humid) (Fig. 1). Sifaka densities within these fragments were variable with 26.7 sifakas/km2 (95% CI 16.2–44.1) in Solaniampilana, 78.17 sifakas/km2 (95% CI 53.1–114.8) in Bekaraoka, and 20.77 sifakas/km2 (95% CI 11.2–38.0) in Binara [59]. Within each forest type, we followed one group in primary forest towards the center of the forest (hereafter interior; characterized by lemurs having a home range at least 300 m from the forest edge) and one group on the edge of the forest fragment (hereafter edge; characterized by having a home range adjacent to the forest edge). Average group size was six individuals and ranged from five to eight (Table 1). Although lemur groups were not marked or collared, we identified individual lemurs by their distinct, permanent physical features (e.g. missing eye, ear cuts, coloration) to ensure we were following the target group.
Group location data
We collected golden-crowned sifaka group location data during two periods, February-April 2019 (rainy season) and June–August 2019 (dry season). Each group was followed for 7–9 consecutive days during the rainy season and the dry season. We followed groups from sleep tree to sleep tree (~ 12 h per day) and collected location data at 15-min intervals. If no animals were visible at the 15-min interval, observers waited to establish visual contact with the social group before recording any locations. During behavioral follows, we maintained a distance of at least 10 m from the lemurs and followed slowly behind the groups in an effort to minimize disrupting their natural behaviors. While the use of telemetry based tracking technology has been shown to reduce disturbances to wildlife behavior, we chose behavioral follows due to a lack of tracking equipment but also to avoid subjecting the animals to the stress of capture and immobilization [64]. In addition to daytime activity, golden-crowned sifaka are known to exhibit nocturnal movements, specifically during periods of bright moon light [65], and thus groups were not always located in the same sleep tree the following morning. In those instances, we reestablished contact with the group as quickly as possible. Group locations were recorded using a GPS receiver (Garmin 64 s), using the Universal Transverse Mercator (UTM) coordinate system (zone 39L), and points were logged at the group’s approximate geometric center.
Foraging and landscape data
We recorded foraging data at the same 15-min intervals using scan sampling to record the behavior, height in the tree, and nearest neighbor of each individual in a group [66]. If an individual was actively feeding during the scan, the plant species and part (e.g., young/mature leaf, leaf petiole, un/ripe fruit, seed, or flower) were identified, GPS location recorded, and data concerning tree species, size, and current phenology collected. In addition to collecting foraging data specific to each of the lemur groups, we also collected general landscape data throughout each of the six lemur home ranges in both the rainy and dry season. We did this by randomly generating forty GPS points within each of the six home ranges (in both rainy and dry seasons) and collected data from potential feeding trees (species, size, phenology) within five m of each location. This allowed us to gain an understanding of the entire landscape of all six home ranges, not just the specific feeding trees utilized by each of the groups.
Home range estimation
Utilization distributions (i.e., 95% isopleth, hereafter home ranges and 50% isopleth, hereafter core area) were estimated for each golden-crowned sifaka group using Dynamic Brownian Bridge Movement Models (DBBMM); [57]. Home range DBBMMs use behavior and movement trajectory data of the animal group that is collected in sequential relocation studies. This method provides a spatially explicit model, which describes the probability of the given animal group occurring in a given location during a specified period. This approach also accounts for temporal autocorrelation, spatial uncertainty, irregularly sampled data, and shifts in an animal’s behavior (resting, foraging, thermoregulating, corridor use, etc.), making it specifically applicable to studies of group living primates [56, 57, 67]. Using DBBMMs to estimate group home ranges requires a Brownian motion variance parameter (σ2, in meters), which quantifies the degree of diffusion or irregularity of an animal’s path [57]. A moving window analysis identifies changes in the movement behavior and estimates σ2 for each step. Because the σ2 parameter is estimated using a “leave-one-method”, the size of the moving window must include an odd number of GPS locations and a margin of greater than three locations bounding each end of the window in which no behavioral changes can occur [57]. We parameterized the DBBMM with a 21-step window size, a 9-step margin size, and a 15 m location error for all lemur groups, as visual inspection indicated these settings were sufficient to identify changes in home range size and overall animal movement [57]. Home ranges were estimated for each lemur group using the DBBMM function in R package ‘move’ [68, 69]. We conducted a three-way analysis of variance (ANOVA) predicting for both home range and core areas sizes, respectively, to determine if season, forest type, interior or edge forests, and the interaction of forest type and season influenced core area and home range size. All analyses were conducted in version 3.6.1 of program R [70]. We used Akaike’s Information Criterion corrected for small samples sizes (AICc) to identify a top model from the set of candidate ANOVA models and the delta of two was as a threshold for equally plausible models [71].
Seasonal core area overlap
To determine the percent of joint home range overlap between the rainy and dry season core areas, we calculated the total area of each home range and then divided the area of overlap between seasons by the total home range size [72].
Areaαβ is the seasonal core area overlap area common to α and β, and core-area α and core-area β are the seasonal core areas of the same group during the rainy and dry season, α and β, respectively. Possible core area overlap ranged from 0% overlap, indicating no shared space use between seasons, to 100% overlap, indicating the dry and rainy season core area ranges overlapped completely. To determine if the core area overlap between the rainy and dry seasons varied as a function of forest type (humid, moderate, or dry), we conducted a one-way ANOVA comparing core area overlap as a function of degree of forest type. All analyses were conducted in version 3.6.1 of program R [70].
Movement rates
We calculated movement rates (meters/monitoring interval [15-min]) for each lemur group using the collected relocation data. The step length (i.e., the distance between sequential recorded locations) was divided by the time elapsed between each sequential location (15-min) to calculate speed for each golden-crowned sifaka group to characterize movement rates. To determine how movement varied across season (rainy and dry), forest disturbance (edge or interior), and forest types (dry, moderate, and humid) we calculated movement rates at both the daily and seasonal scale.
Daily movement rates were bootstrapped to calculate a mean for each observational day. Bootstrapping is a process that involves repeatedly drawing independent samples from a data set (x) to create bootstrap data sets (x1, x2,…, xn). Our samples were performed with replacement which allowed for the same observation to be sampled more than once such that each bootstrapped sample was the same length as our raw lemur speed data (m/15-min). To calculate seasonal movement rates (\(\widehat{SMR}\)), we drew 1000 independent samples (\({\widehat{\alpha }}^{1},{\widehat{\alpha }}^{2},\dots ,{\widehat{\alpha }}^{B})\) to calculate means and standard error (\({\widehat{SE}}_{B}\)), which we then used to generate 95% confidence intervals for comparison of means among seasons and groups,
where \({\widehat{SE}}_{B}\) served as our estimate of the standard error of \(\widehat{\alpha }\) estimated from the raw lemur speed data (m/h). We calculated seasonal movements rates using the bootstrapping approach outlined above but employed the method for each observation season [73, 74].
To determine how environmental variables influenced daily movement rates, we fit LMMs to predict movement rate as a function of all combinations of season (rainy or dry), forest disturbance (edge or interior), and forest type (dry, moderate, and humid), while treating forest type-forest fragmentation per group intercepts as random effects [75]. We used Akaike's Information Criterion corrected for small sample size (AICc) to identify a top model from the set of candidate models [76]. We used the Satterthwaite method to approximate the degrees of freedom and computed p-values for direct effects and interactions using t-statistics [77].
Finally, to determine how environmental variables influenced seasonal movement rates, we conducted a three-way ANOVA of seasonal movement rates as a function of forest type (dry, moderate, and humid), forest disturbance (edge and interior), and season (dry and rainy). We used Akaike's Information Criterion corrected for small sample size (AICc) to identify a top model from the set of candidate models [76]. All analyses were conducted in version 3.6.1 of program R [70].
Habitat selection
To quantify habitat selection of golden-crowned sifaka groups, in relation to tree size and proximity to anthropogenic factors, we fit a Resource Selection Function (RSF) using a use-available design. A RSF is defined as any function producing a value proportional to the probability of selection of a given habitat [78, 79]. Any estimate derived from an RSF is dependent on the definition of available habitats [54, 78, 80]. For our RSF, selection by golden-crowned sifaka availability was considered within home range selection (Johnson’s third order [80]) as defined by a 95% seasonal home range using DBBMMs. Within our seasonal home ranges, we characterized availability by systematically identifying available locations at intervals of 10 m, as this was the spatial resolution of all spatial data used in the RSF [81].
We created our RSFs by fitting generalized linear mixed-effects model (GLMM) with a binomial exponential family and logit link function, which included a group-specific (forest type and disturbance) random intercept term to account for non-independence of habitat associations within groups [82, 83]. For our RSF, we used GPS locations of all feeding trees that golden-crowned sifaka utilized during the rainy and dry field seasons and possible locations within their known home ranges. We extracted tree basal area (cross-sectional area of trees at breast height), Euclidian distance to village, road, and habitat fragment edge for each golden-crowned sifaka feeding tree and each available location. These data were generated using satellite imagery and habitat sampling of resources within each of the six lemur home ranges.
To relate tree basal area and crown volume to lemur GPS location data, we created continuous surfaces of tree basal area and crown volume estimates across our study area by using inverse distance weighting (IDW) interpolation in the package gstat [84]. IDW uses a weighted average of estimates from nearby sampling locations to predict tree basal area and crown volume estimates to the surrounding pixels of a sampling location composed of user-specified areas [85]. Our user-specified areas of inference were 169 m2 because it most closely matched the mean distance between vegetation sampling locations (148.85 m). This interpolation process provided spatially explicit estimates of tree basal area and crown volume estimates which we could then associate with our lemur GPS data.
To examine if lemur habitat selection varied across forest types and seasons, we developed candidate models using various combinations of distance to habitat feature (i.e., village, road, and habitat fragment), and basal area, and used Akaike's Information Criterion (AICc) corrected for small samples sizes to identify a top model from the set of candidate models [71] to determine if (1) differences in habitat selection vary as a function of forest type, and (2) differences in habitat selection vary as a function of season at each site. To account for behavioral differences in lemur groups, we accounted for random effects using an ‘animal ID’ that consisted of each lemur group’s respective forest type (dry, moderate, or humid), forest disturbance classification (edge or interior), and season (rainy or dry). We tested for collinearity and no environmental variables used in model development exhibited high correlation (i.e., |r|> 0.7). All coefficients were estimated using the “lme4” package [70, 75].
Overdispersion of the models was examined by calculating the sum of squared Pearson (SSQ) residuals, the ratio of (SSQ residuals/residual degrees-of-freedom), the residual df, and the p-value based on the appropriate χ2 distribution. Additionally, each individual model was a GLM which allowed us to test overdispersion using a chi-squared comparing the model deviance by the residual degrees-of-freedom [86]. To estimate the explanatory power of the models, we calculated a conditional, marginal, and pseudo R2 for each model [87, 88].
Results
Home range and core area size estimations
Overall, home range sizes for golden-crowned sifaka groups in the Loky-Manambato Protected Area were highly variable, ranging from 2.78 to 31.56 hectares (Table 2). Our top ANOVA model (Table 3), revealed that golden-crowned sifaka core areas varied significantly with season (p = 0.006, F(1,10) = 11.84, residual SE = 0.005) with core areas being larger in the rainy season (average of 1.74 hectares in the rainy season, 0.81 hectares in the dry season). However, while our ANOVA model candidate set for home ranges did include season as a top model (dry or rainy; p = 0.136, F(1,10) = 2.63, SE = 0.07), it was no better than our null model (Table 3).
Seasonal core area overlap
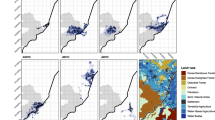
Seasonal core area overlap varied from 16.7 to 53.9% (Table 2; Fig. 2). Core area overlap between the rainy and dry season did not vary with forest type (p = 0.824, F(2.3) = 0.214, SE = 0.072) or forest disturbance (p = 0.617, F(1.4) = 0.343, residual SE < 0.091).

Maps of the Brownian Bridge utilization distributions depicting core area use for golden-crowned sifaka (Propithecus tattersalli) groups in the Loky-Manambato Protected Area of northeastern Madagascar during the dry (June–August 2019; dark gray) and rainy (February-April 2019; light gray) seasons. Overlapping areas were occupied during both seasons. The six boxes display the seasonal home ranges for all six lemur groups followed. Columns indicate forest disturbance classification (interior or edge) and rows indicate the occupied forest type (dry, moderate, or humid). Solid lines correspond to the interior and the hashed lines correspond to the edge in the inset map of each site
Daily and seasonal movement rates
Our top model (Table 4) indicated that seasonal movement rates varied as a function of season (Sum Squares = 0.405, 95% CI [0.13, 0.65], F(1,10) = 11.27, p = 0.007, residual S.E. = 0.04), with higher rates in the rainy season (rainy season: 83.47 m/h; dry season: 56.70 m/h; Fig. 3; Additional file 1: Table S1). When investigating daily movement rates, our LMM analysis supported our seasonal movement results, as the top model included season; however, there was not support for effects of forest type or forest disturbance (β = 0.36, 95% CI [0.11, 0.60]; p = 0.019; Table 4; Additional file 1: Table S2).

Seasonal movement rates (meters/hour) of golden-crowned sifaka (Propithecus tattersalli) groups in the Loky-Manambato Protected Area in northeastern Madagascar. Data was collected during the dry (June–August 2019) and rainy (February-April 2019) seasons using relocation data collected every 15 min. The step length (e.g., the distance between sequential locations) was divided by the time elapsed between each step to calculate speed for each lemur group. Black lines correspond to 95% confidence intervals
Habitat selection
We found that lemur habitat selection varied between humid, moderate, and dry forest types as indicated by AICc (Table 5). We also found that selection by lemurs varied between the wet and dry season, within forest types, as indicated by AICc (Table 6). Given selection varied by forest type and season, we elected to make inferences on selection by forest type and by season. This necessitated we change our models from GLMMs to generalized linear models (GLMs) wherein we dropped the group-specific (forest type and disturbance) random intercept term given our final models retained only two groups [89]. Tests for overdispersion and explanatory power raised no concern (Additional file 1: Tables S3 and S4).
We found that groups in dry deciduous forests selected locations with greater crown volume in the dry season (β = 1.22, S.E. ± 0.11, p < 0.001). During the rainy season, groups in dry forest selected locations with greater crown volume (β = 1.04, S.E. ± 0.12, p < 0.001) and greater tree basal area (β = 2.89, S.E. ± 0.73, p < 0.001), and avoided habitat closer to villages (β = 3.06, S.E. ± 0.74, p < 0.001) and roads (β = 2.42, S.E. ± 0.50, p < 0.001; Fig. 4).

Selection coefficient plot for golden-crowned sifakas (Propithecus tattersalli) in the Loky-Manambato Protected Area in northeastern Madagascar in the dry (June–August 2019) and rainy (February-April 2019) seasons within three forest types (dry, moderate, and wet). This coefficient plot displays beta estimates for tree basal area and tree crown volume and distance to forest edge, roads, and villages. Blue points represent habitat selection during the rainy season and red points represent habitat selection during the dry season. Solid lines above and below each point represent the 95% confidence intervals around each beta estimate
Lemur groups in moderate evergreen forests selected locations with greater crown volume (β = 0.52, S.E. ± 0.07, p < 0.001), greater tree basal area (β = 0.35, S.E. ± 0.17, p = 0.03), and locations farther from villages (β = 1.30, S.E. ± 0.32, p < 0.001) in the dry season. In the rainy season, groups selected feeding locations with greater tree crown volume (β = 1.24, S.E. ± 0.07, p < 0.001) and greater tree basal area (β = 0.55, S.E. ± 0.19, p = 0.003) and avoided habitats closer to villages (β = 1.91, S.E. ± 0.38, p < 0.001; Fig. 4).
Finally, we found that lemur groups in the humid forests selected feeding locations characterized with greater crown volume (β = 0.98, S.E. ± 0.07, p < 0.001), closer to villages (β = -0.76, S.E. ± 0.30, p = 0.01), and closer to the forest edge (β = − 1.04, S.E. ± 0.17, p < 0.001), and avoided locations near roads (β = 2.79, S.E. ± 0.93, p = 0.003) in the dry season. In the rainy season groups in humid forests selected locations with greater tree basal area (β = 0.085, S.E. ± 0.04, p = 0.049) and greater crown volume (β = 1.34, S.E. ± 0.06, p < 0.001) that were closer to villages (β = -1.28, S.E. ± 0.19, p < 0.001), and forest edges (β = − 0.24, S.E. ± 0.05, p < 0.001), and avoided habitats near roads (β = 3.26, S.E. ± 0.73, p < 0.001). While the effects of crown volume, villages, forest edges and roads were the same across seasons, these effects were stronger in the rainy season (Fig. 4).
Discussion
There are three primary results from our study. First, golden-crowned sifaka (Propithecus tattersalli) movement rates are greater in the rainy season and in more humid forests. Second, golden-crowned sifaka core area size is impacted by season, with larger core areas used in the rainy season. Third, golden-crowned sifakas select foraging locations where the largest trees in their home range are located. Similarly, we also detected variation in behavioral responses to villages, road networks, and the forest edge. Golden-crowned sifaka groups in humid and dry deciduous forest fragments specifically avoided foraging locations near road networks in the rainy season, while lemurs in the moderate evergreen forest did not select or avoid locations near road networks. In sum, groups of golden-crowned sifaka showed marked variation in behavioral responses to human disturbance, but for all groups, higher-use zones were characterized by locations having larger trees. Thus, season, forest type, and disturbance all have effects on golden-crowned sifaka space use and ranging behavior.
Across seasons and regardless of forest type or disturbance, golden-crowned sifaka group daily movement rates shifted, with groups moving farther per unit time in the rainy season. This contrasts with previous studies on Milne-Edwards’ sifaka groups that found no seasonal effects on distance moved per day (i.e., daily path length) [41]. Thus, there exists some degree of variability among sifaka species. Movement rates in golden-crowned sifaka groups were also closely linked to home range size in that as home range size increased in the rainy season, so did the average distance moved per hour. This finding is consistent with other studies of highly mobile mammals that found that movement rates and resource availability determined home range size of white-tailed deer (Odocoileus virginianus) and Iberian ibex (Capra pyrenaica) [90, 91].
Golden-crowned sifaka group home range sizes varied between 3 and 32 ha. These home range sizes were smaller than those of diademed sifaka groups in Madagascar’s eastern humid forests, which range from 19 to 79 ha [92], but were larger than those of Verreaux’s sifaka groups in Madagascar’s southern dry forests, which have home range sizes ranging from 5 to 10 ha [93]. Similar to this trend, mouse lemurs (Microcebus spp) inhabiting western dry forests were able to maintain higher population densities than mouse lemur species inhabiting eastern humid forests [94]. Contrary to previous findings, and our predictions, we found that golden-crowned sifaka groups in dry deciduous and humid forests did not occupy significantly larger home range or core area sizes compared to groups living in moderate evergreen forest fragments. This finding was unexpected because golden-crowned sifaka densities were substantially larger in moderate forest fragments compared to dry deciduous and humid forest fragments [59]. In sum, our study was the first determining that the same species of sifaka can inhabit drastically different forest types and display great variation in home range size. However, while animal densities have implications for spatial ecology [95], golden-crowned sifaka densities did not appear to predict home range and core areas sizes [59].
Our prediction that golden-crowned sifaka home range sizes would vary between the rainy season and the dry season was partially supported. While home range sizes were not significantly different between seasons, core area size was statistically larger for golden-crowned sifaka groups in the rainy season compared to the same groups’ core area range sizes in the dry season. Similar to findings of Milne-Edwards’ sifaka, we found that golden-crowned sifaka maintained similar home range locations in both seasons, but displayed considerable seasonal shifts in core area locations [41]. This difference was likely due to the non-uniform and seasonal variation in resource distribution that influenced how golden-crowned sifaka distributed their space use to forage efficiently [96]. Surprisingly, the degree of core area overlap we observed did not vary based on the forest type or forest disturbance level occupied.
We predicted that golden-crowned sifaka groups in more degraded, edge, habitats would occupy larger home ranges and have larger core areas than those in interior forests; however, our data did not support that assumption. Previous studies have demonstrated varying effects of disturbance on home range size of eastern sifaka species in rainforest habitats. For instance, diademed sifaka living along forest edges occupied significantly smaller home range sizes than conspecifics in interior, undisturbed forests [40], as is consistent with many other mammalian species [97]. However, Milne-Edwards’ sifaka in disturbed (logged) forests maintained larger home range sizes than those in undisturbed forests [41]. Unfortunately, the majority of lemur studies (87%) examining the effects of forest disturbance on lemur health, genetics, biodiversity, and behavior were conducted in the humid forests of eastern Madagascar [98]. Further, lemur responses to habitat edges in dry forest are often highly variable, with groups avoiding, selecting, or demonstrating no response in regards to feeding along forest edges [98, 99]. As a result, further investigation of home ranges of golden-crowned sifaka and other dry forest lemurs are needed to understand how increasing anthropogenic changes are influencing lemur ecology and conservation.
While we expected golden-crowned sifaka groups to avoid villages and roads, we found a mixed response. In some forest types and seasons, sifaka groups selected for foraging locations near roads, while in other forest types and seasons sifaka groups avoided foraging locations near roads. The same mixed response was found in regard to foraging locations near villages. Other studies have found similar mixed responses to anthropogenic influences [100, 101]. Additionally, the national road that bisects the global range of golden-crowned sifaka is currently being paved to improve access to mineral reserves and transportation through the region. While infrastructure improvement will likely provide much needed improvement to local economies, our results demonstrate that even narrow road networks can restrict suitable sifaka habitat. Improved road access also is likely to increase resource extraction within the region (e.g., selective logging of hardwoods and gold mining), with negative impacts expected for sifakas [38]. The negative impacts of road networks are well documented for mammalian species. For example, elk (Cervus canadensis) and caribou (Rangifer tarandus) tend to avoid road crossings and seek cover when in close proximity to road networks [102]. Road expansion and paving also increases the prevalence of vehicle collisions with wildlife (e.g., Asiatic cheetah (Acinonyx jubatus venaticus) [103] and Florida panther (Puma concolor coryi)) [104]. Importantly, even low vehicle traffic (0–30 vehicles/12 h) caused wolverines (Gulo gulo) to alter their movement patterns and to avoid areas with road networks [105]. Consequently, increasing human activity and road prevalence is likely to impact foraging and space use behavior of wildlife species, with potentially disastrous consequences for already crucially endangered golden-crowned sifaka.
Conclusions
Limitations and future directions
Although our data revealed various patterns in golden-crowned sifaka movement and foraging patterns, we could only establish general conclusions due to our limited monitoring efforts (15–18 days/year for each group) and small sample size (six sifaka groups). Additionally, due to our rotation-based field schedule, we were only able to follow one group of sifaka at a time. Future work with more continuous monitoring, preferably with the use of remote tracking devices, would enable us to better tease apart the relationships between season, forest type, anthropogenic factors, and movement, as well as to increase model robustness. Second, there were limitations regarding the covariates used in the RSF. Specifically, we assumed that all village disturbance was equal. However, each village differed in population size, degree of infrastructure, and distance from the forest edge. As a result, some villages could have had greater anthropogenic influences than others. Third, we could not claim that human disturbance was directly related to golden-crowned sifaka groups selecting for the largest trees within their respective home ranges. Feeding on the largest trees could have been related to increased food availability and security, rather than human driven factors. Fourth, we used simple categories (e.g., ‘rainy’ and ‘dry’) to reflect seasons, forest types, and disturbance levels. While this uncovered broad patterns, more sophisticated resource measurements, such as precipitation amounts or Landsat imagery, would improve the context within which we interpret sifaka movement and resource selection.
Conservation implications
Our study illustrates the complex anthropogenic and ecological processes that influence movement behavior of golden-crowned sifaka groups. We found evidence that human settlements and road networks play an important role in shaping sifaka foraging and ranging behavior. Additionally, ecological factors such as season are drivers of home range size and space use in this species. Our study enforces the importance of studying primate groups in both the rainy and dry seasons to ensure that conservation efforts meet the full range of a species’ movement, home range size, and resource needs. By understanding how forest type influences golden-crowned sifaka movement and foraging behavior, conservation management plans can be appropriately crafted to the unique forest types throughout the Loky-Manambato Protected Area (humid, moderate evergreen, dry deciduous, littoral, etc.), rather than the region as a whole. Our findings can also be used to inform Malagasy infrastructure and road development plans by working with local conservation NGOs, government officials, and construction teams to limit construction nearby lemur home ranges that are most impacted by human activity. We would advise that the national road not be re-routed towards Binara, the humid forest fragment, due to the strong avoidance lemurs display towards existing road networks and the increased movement of lemurs within this forest fragment. We detected the least avoidance of anthropogenic activity for lemurs in the moderate evergreen forest type, suggesting they are more resilient to the negative effects of human infrastructure. Overall, as anthropogenic disturbance continues to alter habitat structure throughout Madagascar, a deeper knowledge of how fragmentation, habitat loss, and infrastructure development influence golden-crowned sifaka space use, density, and population health will be essential for wildlife managers to make well informed decisions that improve conservation plans for at-risk species.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Tabarelli M, Venceslau A, Cezar M, Paul J, Peres CA. Prospects for biodiversity conservation in the Atlantic Forest: lessons from aging human-modified landscapes. Biol Conserv. 2010;143:2328–40.
Allen AM, Singh NJ. Linking movement ecology with wildlife management and conservation. Front Ecol Evol. 2016;3:1–13.
Delciellos AC, Ribeiro SE, Vieira MV. Habitat fragmentation effects on fine-scale movements and space use of an opossum in the Atlantic Forest. J Mammal. 2017;98:1129–36.
Gastón A, Ciudad C, Mateo-Sánchez MC, García-Viñas JI, López-Leiva C, Fernández-Landa A, et al. Species’ habitat use inferred from environmental variables at multiple scales: how much we gain from high-resolution vegetation data? Int J Appl Earth Obs Geoinf. 2017;55:1–8.
Festa-Bianchet M, Apollonio M. Animal behavior and wildlife conservation. Washington, DC: Island Press; 2003.
Merrick MJ, Koprowski JL. Should we consider individual behavior differences in applied wildlife conservation studies? Biol Conserv. 2017;209:34–44.
Grüss A, Kaplan DM, Guénette S, Roberts CM, Botsford LW. Consequences of adult and juvenile movement for marine protected areas. Biol Conserv. 2011;144:692–702.
Angeloni L, Schlaepfer MA, Lawler JJ, Crooks KR. A reassessment of the interface between conservation and behaviour. Anim Behav. 2008;75:731–7.
Choi CY, Peng HB, He P, Ren XT, Zhang S, Jackson MV, et al. Where to draw the line? Using movement data to inform protected area design and conserve mobile species. Biol Conserv. 2019;234:64–71.
Berger-Tal O, Polak T, Oron A, Lubin Y, Kotler BP, Saltz D. Integrating animal behavior and conservation biology: a conceptual framework. Behav Ecol. 2011;22:236–9.
Cooke SJ, Blumstein DT, Buchholz R, Caro T, Fernández-Juricic E, Franklin CE, et al. Physiology, behavior, and conservation. Physiol Biochem Zool. 2014;87:1–14.
Berger-Tal O, Blumstein DT, Carroll S, Fisher RN, Mesnick SL, Owen MA, et al. A systematic survey of the integration of animal behavior into conservation. Conserv Biol. 2016;30:744–53.
Shillinger GL, Palacios DM, Bailey H, Bograd SJ, Swithenbank AM, Gaspar P, et al. Persistent leatherback turtle migrations present opportunities for conservation. PLoS Biol. 2008;6:1408–16.
Reyna-hurtado R, Teichroeb JA, Bonnell TR, Hernández-sarabia RU, Vickers SM, Serio-silva JC, et al. Primates adjust movement strategies due to changing food availability. Behav Ecol. 2018;29:368–76.
Trapanese C, Meunier H, Masi S. What, where and when: spatial foraging decisions in primates. Biol Rev. 2019;94(2):483–502.
Rosenzweig ML. A theory of habitat selection. Ecology. 1981;62:327–35.
Nagy-Reis MB, Setz EZF. Foraging strategies of black-fronted titi monkeys (Callicebus nigrifrons) in relation to food availability in a seasonal tropical forest. Primates. 2017;58:149–58.
Campera M, Serra V, Balestri M, Barresi M, Ravaolahy M, Randriatafika F, et al. Effects of habitat quality and seasonality on ranging patterns of collared brown lemur (Eulemur collaris) in littoral forest fragments. Int J Primatol. 2014;35:957–75.
Pope NS, Jha S. Seasonal food scarcity prompts long-distance foraging by a wild social bee. Am Nat. 2018;191:45–57.
Wato YA, Prins HHT, Heitkönig IMA, Wahungu GM, Ngene SM, Njumbi S, et al. Movement patterns of African Elephants (Loxodonta africana) in a Semi-arid Savanna suggest that they have information on the location of dispersed water sources. Front Ecol Evol. 2018;6:1–8.
Asensio N, Schaffner CM, Aureli F. Variability in core areas of spider monkeys (Ateles geoffroyi) in a tropical dry forest in Costa Rica. Primates. 2012;53:147–56.
Branco PS, Merkle JA, Pringle RM, Pansu J, Potter AB, Reynolds A, et al. Determinants of elephant foraging behaviour in a coupled human-natural system: Is brown the new green? J Anim Ecol. 2019;88:780–92.
Rice MB, Apa AD, Wiechman LA. The importance of seasonal resource selection when managing a threatened species: Targeting conservation actions within critical habitat designations for the Gunnison sage-grouse. Wildl Res. 2017;44:407–17.
Street GM, Fieberg J, Rodgers AR, Carstensen M, Moen R, Moore SA, et al. Habitat functional response mitigates reduced foraging opportunity: implications for animal fitness and space use. Landsc Ecol. 2016;31:1939–53.
Said S, Servanty S. The influence of landscape structure on female roe deer home-range size. Landsc Ecol. 2005;20:1003–12.
Holzman S, Conroy MJ, Pickering J. Home range, movements, and habitat use of coyotes in southcentral Georgia. J Wildl Manage. 1992;56:139–46.
McLean KA, Trainor AM, Asner GP, Crofoot MC, Hopkins ME, Campbell CJ, et al. Movement patterns of three arboreal primates in a Neotropical moist forest explained by LiDAR-estimated canopy structure. Landsc Ecol. 2016;31:1849–62.
Campos FA, Bergstrom ML, Childers A, Hogan JD, Jack KM, Melin AD, et al. Drivers of home range characteristics across spatiotemporal scales in a Neotropical primate, Cebus capucinus. Anim Behav. 2014;91:93–109.
Haddad NM, Brudvig LA, Clobert J, Davies KF, Gonzalez A, Holt RD, et al. Habitat fragmentation and its lasting impact on Earth’s ecosystems. Sci Adv. 2015;1(2):e1500052.
Menchaca A, Rossi NA, Froidevaux J, Dias-freedman I, Caragiulo A, Wultsch C, et al. Population genetic structure and habitat connectivity for jaguar (Panthera onca) conservation in Central Belize. BMC Genet. 2019;20:1–13.
Peaden JM, Nowakowski AJ, Tuberville TD, Buhlmann KA, Todd BD. Effects of roads and roadside fencing on movements, space use, and carapace temperatures of a threatened tortoise. Biol Conserv. 2017;214:13–22.
Rogan MS, Miller JRB, Lindsey PA, Mcnutt JW. Socioeconomic drivers of illegal bushmeat hunting in a Southern African Savanna. Biol Conserv. 2018;226:24–31.
Cardillo M, MacE GM, Gittleman JL, Jones KE, Bielby J, Purvis A. The predictability of extinction: biological and external correlates of decline in mammals. Proc R Soc B Biol Sci. 2008;275:1441–8.
Zeller KA, Wattles DW, Conlee L, Destefano S. Black bears alter movements in response to anthropogenic features with time of day and season. Mov Ecol. 2019;7:1–14.
Polfus JL, Hebblewhite M, Heinemeyer K. Identifying indirect habitat loss and avoidance of human infrastructure by northern mountain woodland caribou. Biol Conserv. 2011;144:2637–46.
Isaac NJB, Cowlishaw G. How species respond to multiple extinction threats. Proc R Soc B Biol Sci. 2004;271:1135–41.
de Almeida-Rocha JM, Peres CA, Oliveira LC. Primate responses to anthropogenic habitat disturbance: a pantropical meta-analysis. Biol Conserv. 2017;215:30–8.
Estrada A, Garber PA, Rylands AB, Roos C, Fernandez-duque E, Di FA, et al. Impending extinction crisis of the world’s primates: Why primates matter. Sci Adv. 2017;3:1–16.
Vieilledent G, Grinand C, Rakotomalala FA, Ranaivosoa R, Rakotoarijaona J, Allnutt TF, et al. Combining global tree cover loss data with historical national forest cover maps to look at six decades of deforestation and forest fragmentation in Madagascar. Biol Conserv. 2018;222:189–97.
Irwin MT. Feeding ecology of Propithecus diadema in forest fragments and continuous forest. Int J Primatol. 2008;29:95–115.
Gerber BD, Arrigo-Nelson S, Karpanty SM, Kotschwar M, Wright PC. Spatial ecology of the endangered Milne-Edwards’ Sifaka (Propithecus edwardsi): do logging and season affect home range and daily ranging patterns? Int J Primatol. 2012;33:305–21.
Erhart EM, Tecot SR, Grassi C. Interannual variation in diet, dietary diversity, and dietary overlap in three sympatric strepsirrhine species in southeastern Madagascar. Int J Primatol. 2018;39:289–311.
Herrera JP, Borgerson C, Tongasoa L, Andriamahazoarivosoa P, Rasolofoniaina BJR, Rakotondrafarasata ER, et al. Estimating the population size of lemurs based on their mutualistic food trees. J Biogeogr. 2018;45(11):2546–63.
Baden AL. A description of nesting behaviors, including factors impacting nest site selection, in black-and-white ruffed lemurs (Varecia variegata). Ecol Evol. 2019;9:1010–28.
Lahann P, Dausmann KH. Live fast, die young: flexibility of life history traits in the fat-tailed dwarf lemur (Cheirogaleus medius). Behav Ecol Sociobiol. 2011;65:381–90.
Norscia I, Carrai V, Borgognini-Tarli SM. Influence of dry season and food quality and quantity on behavior and feeding strategy of Propithecus verreauxi in Kirindy, Madagascar. Int J Primatol. 2006;27:1001–22.
Quéméré E, Champeau J, Besolo A, Rasolondraibe E, Rabarivola C, Crouau-Roy B, et al. Spatial variation in density and total size estimates in fragmented primate populations: the golden-crowned sifaka (Propithecus tattersalli). Am J Primatol. 2010;72:72–80.
Quéméré E, Crouau-Roy B, Rabarivola C, Louis EE, Chikhi L. Landscape genetics of an endangered lemur (Propithecus tattersalli) within its entire fragmented range. Mol Ecol. 2010;19:1606–21.
Goodman SM, Raherilalao MJ, Wohlhauser S. Site 6: Loky Manambato. Terr Prot Areas Madagascar Their Hist Descr Biot. 2018.
Quéméré E, Amelot X, Pierson J, Crouau-roy B, Chikhi L. Genetic data suggest a natural prehuman origin of open habitats in northern Madagascar and question the deforestation narrative in this region. PNAS. 2012;109:13028–33.
Salmona J, Heller R, Quéméré E, Chikhi L. Climate change and human colonization triggered habitat loss and fragmentation in Madagascar. Mol Ecol. 2017;26:5203–22.
Lehmann J, Boesch C. Social influences on ranging patterns among chimpanzees (Pan troglodytes verus) in the Tai National Park, Cote d’Ivoire. Behav Ecol. 2002;14:642–9.
Steiniger S, Hunter AJS. A scaled line-based kernel density estimator for the retrieval of utilization distributions and home ranges from GPS movement tracks. Ecol Inform. 2013;13:1–8.
Avgar T, Potts JR, Lewis MA, Boyce MS. Integrated step selection analysis: bridging the gap between resource selection and animal movement. Methods Ecol Evol. 2016;7:619–30.
Kranstauber B. Modelling animal movement as Brownian bridges with covariates. Mov Ecol. 2019;7:1–10.
Fieberg J, Borger L. Could you please phrase “home range” as a question? J Mammal. 2012;93:890–902.
Kranstauber B, Kays R, Lapoint SD, Wikelski M, Safi K. A dynamic Brownian bridge movement model to estimate utilization distributions for heterogeneous animal movement. J Anim Ecol. 2012;81:738–46.
Wright PC. Lemur traits and Madagascar ecology: coping with an island environment. Yearb Phys Anthropol. 1999;42:31–72.
Semel B, Karpanty SM, Semel MA, Stauffer DF, Quéméré E, Walters JR, et al. Highly variable densities and a decline in critically endangered golden-crowned sifaka (Propithecus tattersalli) population abundance from 2008–2018. Int J Primatol.
Worman COD, Chapman CA. Densities of two frugivorous primates with respect to forest and fragment tree species composition and fruit availability. Int J Primatol. 2006;27:203–25.
Jaeger JAG, Bowman J, Brennan J, Fahrig L, Bert D, Bouchard J, et al. Predicting when animal populations are at risk from roads: an interactive model of road avoidance behavior. Ecol Model. 2005;185:329–48.
Meyers DM. The effects of resource seasonality on behavior and reproduction in the golden-crowned Sifaka (Propithecus tattersalli, Simons, 1988) in three Malagasy forests. Durham, NC: Duke University; 1993.
Rakotonanahary AN, Andriaholinirina NV, Rasoloharijaona S, Rajaonarison JF, Hagenson RA, Sefczek TM, et al. Habituation of greater bamboo Lemurs (Prolemur simus) in the Vatovavy forest, Madagascar. Primate Conserv. 2021;35:117–24.
Hebblewhite M, Haydon DT. Distinguishing technology from biology: a critical review of the use of GPS telemetry data in ecology. Philos Trans R Soc B Biol Sci. 2010;365:2303–12.
Erkert HG, Kappeler PM. Arrived in the light: diel and seasonal activity patterns in wild Verreaux’s sifakas (Propithecus v. verreauxi; Primates: Indriidae). Behav Ecol Sociobiol. 2004;57:174–86.
Altmann J. Observational study of behavior: sampling methods. Behaviour. 1974;49:227–67.
Gurarie E, Andrews RD, Laidre KL. A novel method for identifying behavioural changes in animal movement data. Ecol Lett. 2009;12:395–408.
Horne JS, Garton EO, Krone SM, Lewis JS. Analyzing animal movements using Brownian bridges. Ecology. 2007;88:2354–63.
Kranstauber B, Smolla M, Scharf A. Visualizing and analyzing animal track data. 2013.
R Core Team. Vienna: R Foundation for Statistical Computing; 2020.
Burnham KP, Anderson DR. Model selection and multimodel inference: a practical information-theoretic approach. New York, NY: Springer-Verlag; 2002.
Atwood TC, Weeks HP. Spatial home-range overlap and temporal interaction in eastern coyotes: the influence of pair types and fragmentation. Can J Zool. 2003;81:1589–97.
Booth JG, Hall P, Wood ATA. Balanced importance resampling for the bootstrap. Annu Stat. 1993;21:286–98.
Davison AC, Hinkley DV, Schechtman E. Efficient bootstrap simulation. Biometrika. 1986;73:555–66.
Bates D, Mächler M, Bolker BM, Walker SC. Fitting linear mixed-effects models using lme4. J Stat Softw. 2015;67:1–48.
Hurvich CM, Tsai CL. Regression and time series model selection in small samples. Biometrika. 1989;76:297–307.
Gaylor DW, Hopper FN. Estimating the degrees of freedom for linear combinations of mean squares by Satterthwaite’s formula. Technometrics. 1969;11:691–706.
Lele SR, Merrill EH, Keim J, Boyce MS. Selection, use, choice and occupancy: clarifying concepts in resource selection studies. J Anim Ecol. 2013;82:1183–91.
Manly BF, McDonald L, Thomas D, McDonald TL, Erickson WP. Resource selection by animals. 2nd ed. Dordrecht: Springer; 2002.
Johnson DJ. The comparison of usage and availability measurements for evaluating resource preference. Ecology. 1980;61:65–71.
Benson JF. Improving rigour and efficiency of use-availability habitat selection analyses with systematic estimation of availability. Methods Ecol Evol. 2013;4:244–51.
Gillies CS, Hebblewhite M, Nielsen SE, Krawchuk MEGA, Aldridge CL, Jacqueline L, et al. Application of random effects to the study of resource. J Anim Ecol. 2006;75:887–98.
Hebblewhite M, Merrill E. Modelling wildlife—human relationships for social species with mixed-effects resource selection models. J Appl Ecol. 2008;45:834–44.
Pebesma E, Graeler B, Pebesma ME. Package ‘gstat’. 2019.
De Smith MJ, Goodchild MF, Longley PA. Geospatial analysis: a comprehensive guide to principles techniques and software tools. 6th ed. La Vergne, TN: Ingram Publisher Services; 2018.
Hilbe JM, Robinson AP. Methods of Statistical Model Estimation. 2013.
Nakagawa S, Johnson PCD, Schielzeth H. The coefficient of determination R2 and intra-class correlation coefficient from generalized linear mixed-effects models revisited and expanded. J R Soc Interface. 2017;14:20170213.
Smith TJ, McKenna CM. A comparison of logistic regression pseudo R2 indices. Mult Linear Regres Viewp. 2013;39:17–26.
Bolker BM, Brooks ME, Clark CJ, Geange SW, Poulsen JR, Stevens MHH, et al. Generalized linear mixed models: a practical guide for ecology and evolution. Trends Ecol Evol. 2009;24:127–35.
Rhoads CL, Bowman JL, Eyler B. Home range and movement rates of female exurban white-tailed deer. J Wildl Manage. 2010;74:987–94.
Viana DS, Granados JE, Fandos P, Pérez JM, Cano-manuel FJ, Burón D, et al. Linking seasonal home range size with habitat selection and movement in a mountain ungulate. Mov Ecol. 2018;6:1–11.
Irwin MT. Diademed sifaka (Propithecus diadema) ranging and habitat use in continuous and fragmented forest: higher density but lower viability in fragments? Biotropica. 2008;40:231–40.
Benadi G, Fitchel C, Kappeler P. Intergroup relations and home range use in Verreaux’s Sifaka (Propithecus verreauxi). Am J Primatol. 2008;70:956–65.
Setash CM, Zohdy S, Gerber BD, Karanewsky CJ. A biogeographical perspective on the variation in mouse lemur density throughout Madagascar. Mamm Rev. 2017;47:212–29.
Webber QMR, Vander WE. An evolutionary framework outlining the integration of individual social and spatial ecology. J Anim Ecol. 2018;87:113–27.
Ostro LET, Silver SC, Koontz FW, Young TP. Ranging behavior of translocated and established groups of black howler monkeys Alouatta pigra in Belize, Central America. Biol Conserv. 1999;87:181–90.
Tucker MA, Böhning-gaese K, Fagan WF, Fryxell JM, Van Moorter B, Alberts SC, et al. Moving in the Anthropocene: global reductions in terrestrial mammalian movements. Science (80-). 2018;359:466–9.
Kling KJ, Yaeger K, Wright PC. Trends in forest fragment research in Madagascar: documented responses by lemurs and other taxa. Am J Primatol. 2020;82(4):e23092.
Lehman SM, Rajaonson A, Day S. Edge effects and their influence on lemur density and distribution in southeast Madagascar. Am J Phys Anthropol. 2006;129:232–41.
Gaynor KM, Hojnowski CE, Carter NH, Brashares JS. The influence of human disturbance on wildlife nocturnality. Science (80-). 2018;360:1232–5.
Suraci JP, Clinchy M, Zanette LY, Wilmers CC. Fear of humans as apex predators has landscape-scale impacts from mountain lions to mice. Ecol Lett. 2019;22:1578–86.
Prokopenko CM, Boyce MS, Avgar T. Characterizing wildlife behavioural responses to roads using integrated step selection analysis. J Appl Ecol. 2017;54:470–9.
Mohammadi A, Almasieh K, Clevenger AP, Fatemizadeh F. Road expansion: a challenge to conservation of mammals, with particular emphasis on the endangered Asiatic cheetah in Iran. J Nat Conserv Elsevier. 2018;43:8–18.
Criffield M, Van De Kerk M, Leone E, Cunningham MW, Lotz M, Oli MK, et al. Assessing impacts of intrinsic and extrinsic factors on Florida panther movements. J Mammal. 2018;99:702–12.
Scrafford MA, Avgar T, Heeres R, Boyce MS. Roads elicit negative movement and habitat-selection responses by wolverines (Gulo gulo luscus). Behav Ecol. 2018;29:534–42.
Acknowledgements
We would like to sincerely thank Amidou Souleimany, Aylett Lipford, Giovanni Walters, our local guides (Amadou, Andre, Augiste, Bezily, Christone, Da, Edward, Ishmael, Jaojoby, John, Justin, Lahimena, Laurent, Lucien, Mamoud, Michelle, Moratombo, Patrice, Pierre, Seraphin, Sylvano, Theodore, Thierry, Zoky), cooking staff (Ayati, Fatomia, Francia, Jao Fera, Nicole), and porters, Fanamby (Serge Rajaobelina, Richelin, Narcisse, Tiana Andriamanana, Sylvano Tsialazo), and Madagascar Institute for the Conservation of Tropical Environments (MICET; Benjamin Andriamihaja, Benji Randrianambinina, Claude, Nary) for their assistance with data collection and logistics.
Funding
We acknowledge funding from the Rufford Foundation and the National Science Foundation GRFP (DGE 1651272). Opinions, findings, conclusions, or recommendations expressed are those of the authors and do not necessarily reflect the views of the NSF.
Author information
Authors and Affiliations
Contributions
MS and IM designed the study. MS, BS, and TR conducted the fieldwork. MS, HA, and MC conducted the coding and data analyses. MS wrote the first draft of the manuscript and all authors contributed substantially to revisions. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This research was conducted with permission from the Ministry of Foreign Affairs of Madagascar, Madagascar National Parks, the Ministry of the Environment, Forests, and Tourism (MEFT), and Madagascar Institute for the Conservation of Tropical Environments (MICET). MICET was also instrumental in permit acquisition (N015/19/MEEF/SG/DGF/DSAP/SCB) and overall research coordination. Our animal follow methods were approved by the Virginia Tech Institutional Animal Care and Use Committee (IACUC) office (permit #17-127).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Table S1. Mean seasonal speed for golden-crowned sifaka (Propithecus tattersalli) groups. Table S2. Mean daily speed for golden-crowned sifaka (Propithecus tattersalli) groups. Table S3. GLM of golden-crowned sifaka (Propithecus tattersalli) foraging tree selection. Table S4. Formulation of the resource selection GLM of golden-crowned sifaka (Propithecus tattersalli) groups.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Semel, M.A., Abernathy, H.N., Semel, B.P. et al. Environmental and anthropogenic influences on movement and foraging in a critically endangered lemur species, Propithecus tattersalli: implications for habitat conservation planning. Mov Ecol 10, 20 (2022). https://doi.org/10.1186/s40462-022-00320-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40462-022-00320-x