
Abstract
Background
A better understanding of sea turtle spatial ecology is critical for the continued conservation of imperiled sea turtles and their habitats. For resource managers to develop the most effective conservation strategies, it is especially important to examine how turtles use and select for habitats within their developmental foraging grounds. Here, we examine the space use and relative habitat selection of immature green turtles (Chelonia mydas) using acoustic telemetry within the marine protected area, Buck Island Reef National Monument (BIRNM), St. Croix, United States Virgin Islands.
Results
Space use by turtles was concentrated on the southern side of Buck Island, but also extended to the northeast and northwest areas of the island, as indicated by minimum convex polygons (MCPs) and 99%, 95%, and 50% kernel density estimations (KDEs). On average space use for all categories was < 3 km2 with mean KDE area overlap ranging from 41.9 to 67.7%. Cumulative monthly MCPs and their proportions to full MCPs began to stabilize 3 to 6 detection months after release, respectively. Resource selection functions (RSFs) were implemented using a generalized linear mixed effects model with turtle ID as the random effect. After model selection, the accuracy of the top model was 77.3% and showed relative habitat selection values were highest at shallow depths, for areas in close proximity to seagrass, and in reef zones for both day and night, and within lagoon zones at night. The top model was also extended to predict across BIRNM at both day and night.
Conclusion
More traditional acoustic telemetry analyses in combination with RSFs provide novel insights into animal space use and relative resource selection. Here, we demonstrated immature green turtles within the BIRNM have small, specific home ranges and core use areas with temporally varying relative selection strengths across habitat types. We conclude the BIRNM marine protected area is providing sufficient protection for immature green turtles, however, habitat protection could be focused in both areas of high space use and in locations where high relative selection values were determined. Ultimately, the methodologies and results presented here may help to design strategies to expand habitat protection for immature green turtles across their greater distribution.
Similar content being viewed by others
Background
Marine environments are vulnerable to multiple anthropogenic threats, especially nearshore coastal habitats. In these areas, human activities are increasing and often negatively impacting the ecosystem and dependent marine animals [1], including destructive land use practices [2] pollution [3], overexploitation [4], damaging fishing practices [5], dredging [6], and large-scale oil spills [7]. Coastal habitats can intrinsically be more difficult to manage than terrestrial systems because both stakeholder use and oceanic processes are operating at multiple, often complex temporal and spatial scales [8, 9]. In response to the increasing pressures and associated complications for effective conservation efforts, resource managers have begun to rely heavily on spatial management techniques [10] and the establishment of marine protected areas (MPAs) [8, 11,12,13].
MPAs have been implemented around the world and they vary widely in purpose, size, duration, enforcement, and regulation [14, 15]. In recent years, MPA effectiveness has become questioned; many fail to meet conservation and management goals because they lack local or governmental buy-in, enforcement, or are simply too small to be ecologically effective [16,17,18,19]. In addition, spatial conservation strategies often lack the formal incorporation of animal movement data to inform the size, structure, and ultimately, the effectiveness of the MPA [20, 21]. As such, it is essential to quantify the habitat and space use of marine life to ensure management plans adequately match the spatial ecology of the species they are designed to protect. Ultimately, well-designed MPAs should incorporate animal movement data to allow for resource managers to establish and implement the most efficient, appropriate, and effective management decisions to protect resources [22,23,24].
Sub-tropical and tropical coastal habitats are frequently utilized by immature marine turtles as developmental feeding grounds, primarily for green (Chelonia mydas) and hawksbill (Eretmochelys imbricata) turtles. These turtles can remain in these neritic waters, foraging in benthic habitats (e.g., seagrass, sponges) for up to multiple decades, until reaching sexual maturity [25,26,27]. While conservation and management efforts, such as harvest and bycatch regulations, have led to increasing trends in sea turtle populations around the world [28, 29], evidence indicates that coastal habitats are still threatened [30]. Considering turtles use these nearshore waters for a substantial period of their juvenile life stage, MPAs could be well-suited to protect turtles from direct and indirect take, but also protect their forage and shelter habitats. However, a detailed evaluation of the distribution and use of these areas by juvenile sea turtles is critical to determine the efficacy of an MPA [31] for continued conservation of their populations and essential habitats [31, 32].
Satellite telemetry has been the most commonly used technology to examine turtle movements [33], resulting in novel findings related to their spatial ecology. For example, Scott et al. [34] demonstrated, globally, that 35% of satellite tagged green turtles aggregated within MPAs, while Hart et al. [35] highlighted 82% of satellite tagged green turtles were within local and regional MPAs in Florida. However, while satellite telemetry is effective at examining large-scale movements, this technology generally provides coarse-scale position data, thus limiting the capacity to fully understand fine-scale habitat use for immature turtles. To gain insights into fine-scale marine life movements and space use, acoustic telemetry has become a pervasive tool [33]. While researchers must address caveats unique to acoustic telemetry [36], relative to satellite telemetry, acoustic telemetry uses less expensive transmitters (i.e., allows for higher sample sizes) and provides information at both broad and fine spatial–temporal scales that is meaningful for informing local and regional management [33, 37,38,39]. This technology typically involves the use of stationary hydrophone receivers that can detect hundreds of transmitters enabling researchers to answer new fine-scale ecosystem level questions by tagging multiple species [40]. Further, with the increase of transmitters and receivers deployed over the last 25 years, researchers are now able to track individuals across many hundreds of kilometers through large-scale collaborative acoustic telemetry networks [33, 39, 41]. While this technology can be utilized to examine long-distance movements, the most prevalent and useful application for marine turtles is for the study of immature turtle space use within developmental grounds at a finer resolution. While this technology has proven to be valuable when providing insights on the immature turtle home range, space use, and behaviors [31, 42,43,44,45,46,47], there is still much to gain when it comes to understanding marine turtle ecology.
Here, we quantify the space use and relative habitat selection of immature green turtles within the waters of Buck Island Reef National Monument (BIRNM), St. Croix, United States Virgin Islands (USVI) using acoustic telemetry. Relative habitat selection was examined with respect to habitat variables such as zone type (lagoon, bank/shelf, reef, other), reef type (no reef, patch reef, continuous reef), seagrass proximity, and depth. The objectives of this study were to determine immature green turtle space use and relative habitat selection to further understand their spatial ecology within the MPA. These data and analytical methods are useful for developing spatial management strategies for immature green turtles, plus our study also provides insights on the broader use of acoustic telemetry on sea turtles that typically has limited tag retention rates [48].
Results
Each detection log was examined individually to determine if or when a given transmitter may have fallen off a tagged turtle and, thus, resulting in many false detections. After the data were filtered and potential false detections removed, we examined the detection logs of 58 immature green turtles within the BIRNM using 68 transmitters (due to retagging) deployed across 2012 (n = 38) and 2013 (n = 30) (Additional file 1: Figs. S1, S2, Table S1). Individual size of tagged turtles ranged from 33.8 to 65.5 cm (straight line carapace nuchal [SCLn]; 46.9 ± 7.0 cm). Full minimum convex polygon (MCP) values for detection logs ranged from 0.0 to 4.2 km2 (0.9 ± 0.9 km2). Days at liberty (i.e., tracking duration) ranged from 1 to 505 days (146.3 ± 100.1 days) and residency values (i.e., detection days/days at liberty) ranged from 0.6 to 1 (0.8 ± 0.3). Tracking duration was limited, with only four transmitters remaining active 10 detection months after release. This is likely an effect of transmitter retention due to tear-outs from attachment holes failing and/or tags moving towards the edge of their scutes from growth [48] (Fig. 1). Thus, limited and variable tracking durations also led to variation in individual and overall mean cumulative monthly MCP values and the proportions to their full MCPs (Fig. 1). However, cumulative monthly MCP values began to stabilize around six detection months and proportions to their full MCPs appeared to stabilize around three detection months after release.

a Cumulative minimum convex polygon (MCP km2) boxplots, median shown by black horizontal bar, for turtle detection datasets each detection month after release, b boxplots, median shown by black horizontal bar, showing proportion of monthly cumulative MCPs to full MCPs for each individual and each detection month after release, c mean overall proportion of monthly cumulative MCPs to full MCPs for each detection month after release (loess regression with 95% confidence interval (CI)), d number of transmitters active (i.e., producing MCP values) for each detection month after release (loess regression with 95% CI)
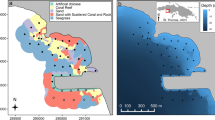
After removing detection logs with less than 100 centers of activity (COAs) (n = 10), we used a total of 97,065 centers of activity to examine the space use of 52 immature green turtles (with 58 transmitters due to retagging) across 2012, 2013, and 2014. Constructed 99% kernel density estimates (KDEs) ranged from 1.2 to 5.9 km2 (2.3 ± 0.8 km2) with a mean overlap of 67.7% (± 19.3%), 95% KDEs range from 0.7 to 3.7 km2 (1.4 ± 0.5 km2) with a mean overlap of 64.5% (± 21.25%), and 50% KDEs ranged from 0.1 to 0.6 km2 (0.3 ± 0.0 km2) with a mean overlap of 41.9% (± 33.6%) (Fig. 2). While 95% and 50% KDEs were largely located on the southern side of Buck Island, some individual 95% KDEs extended around to the northeast and northwest areas of the island as well (Fig. 3).

Boxplots representing the area of turtle kernel density estimations (KDEs) across 50% (yellow), 95% (green), and 99% (red). Original kernel utilization distributions (KUDs) were constructed using a 200-m smoothing parameter. Note, median shown by black horizontal bar

Receivers represented by blue dots, 50% and 95% turtle kernel density estimations represented by yellow and green, respectively. Darker shades of yellow or green indicate higher levels of overlap. Original kernel utilization distributions were constructed using a 200-m smoothing parameter
Resource selection function (RSF) model and relative selection predictions across space
Variance inflation factors revealed reef type and zone type were highly correlated and, thus, we decided to remove reef type from the model. The top RSF generalized linear mixed model (GLMM) (using a random 60% subset of the data) was the full model, as indicated via model selection (Additional file 1: Table S2). This model included diel and depth as an interaction term, diel and distance to seagrass as an interaction term, diel and zone type as an interaction term, and individual and transmitter as the random effects. However, summary outputs for the final model indicated the random effect, year, had a variance component equal to zero, thus was removed in further analysis.
Using the trained RSF GLMM and the holdout dataset (i.e., the random 40% subset of the original data frame), overall model accuracy was 77.3% with predictions of 1 s at 72.2%. While 0 s are considered background points and represent the area that was available (not absences), our model predicted 0 s at 85.2% accuracy. Model performance across the three categorical variables (diel period, and zone type) and their levels ranged from 71.7 to 100% (77.1 ± 11.3, Additional file 1: Table S3).
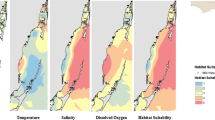
We examined the relative habitat selection of turtles via marginal effects plots as generated from our top model (Fig. 4a). Relative selection by turtles was high in areas 200 m or closer to seagrass in both day and night (Fig. 4a). Shallower depths (less than 10 m) were found to have higher relative selection probabilities both day and night, however, there were higher relative selection probabilities in the day (Fig. 4a). For zone type, turtles had the three highest relative selection probabilities within reef zones at day, reef zones at night, and within lagoon zones at night (Fig. 4a).

a Marginal effects plots for each conditional term in the final model, distance to seagrass (m) at day and night (top), depth (m) at day and night (middle), and zone type at day and night (bottom). All terms not specified in each plot were held constant. Depth and distance to seagrass were held at their means and diel and zone were held at their proportional levels. b Habitat covariates, including distance to seagrass (m, top), depth (m, middle), and zone type (bottom), plotted with the Buck Island Reef National Monument boundary designated by red dashed line
Using the top model to predict across BIRNM (including areas with no detection coverage), the greatest relative selection probabilities for immature green turtles were located within shallow depths near Buck Island and generally decreased farther away from the island at deeper depths (Fig. 5). While turtle KDEs did not perfectly match predicted relative selection probabilities, higher probabilities were located directly adjacent to the island and off the northwestern end the island where lagoon and reef zones exist. Areas in close proximity to seagrass coverage, and lagoon and reef zone habitats produced higher probabilities of relative selection than areas farther away or outside, respectively (Fig. 5).

Predicted relative selection across Buck Island Reef National Monument (BIRNM) during day (above) and night (below) using the top model and its covariates (distance to seagrass, zone, and depth) while taking into account the random effect (turtle ID) variances. The BIRNM boundary designated by red dashed line
Discussion
The combination of acoustic telemetry and RSFs demonstrated that immature green turtles within the BIRNM have small, specific home ranges and core use areas, primarily associated with shallow depths, reef and lagoon zones, and areas of seagrass coverage or areas in close proximity to seagrass. Cumulative space use generally increased with each month passing after release and stabilized around six detection months, while proportions to their full MCPs appeared to stabilize around three detection months following release. Overall, tracking duration times varied among individuals likely due to transmitter retention issues [48], yet our MCP metrics suggested 3 to 6 months of tracking for turtles may provide sufficient amount of data for inferences. RSFs indicated relative habitat selection (i.e., relative to the other habitats available) was high at night and day within reef zones, but only increased within lagoon habitats at night. The highest rates of predicted relative selection were in shallow depths near the island where access to shelter habitats (i.e., reef and lagoon areas) were available. In addition, areas with, or in close proximity to, seagrass had increased relative selection propensities compared to areas farther away from seagrass.
Vulnerable to predation, such as from tiger sharks (Galeocerdo cuvier), immature green turtles must balance foraging opportunities and risk avoidance tactics [49]. Our results support previous findings that turtles forage in productive seagrass habitats and likely utilize physical structure, e.g., lagoon and reef-type structures, for protection [42, 47, 50,51,52]. Furthermore, Casselberry et al. [53] found acoustically tagged tiger sharks within BIRNM were more likely to inhabit depths > 10 m, at which point, relative habitat selection for turtles was very low (< 0.25). The spatial distribution of individual MCPs and 99%, 95%, and 50% KDEs around BIRNM was relatively small and on average < 3 km2. Prior studies have reported similar home range sizes, with the estimated home ranges for immature green turtles in Hawaii being < 3 km2 [54] and in Florida were 3 km2 [42]. Further, turtle core use areas, defined by 50% KDEs, within BIRNM were small (0.3 ± 0.0 km2), similar to that reported by Makowski et al. [42] (0.5 ± 0.4 km2) and Chambault et al. [55] (0.2 ± 0.3 km2). When resource distributions (i.e., food and shelter) were tightly clustered, space use for immature green turtles was constrained [42, 47, 54]. Alternatively, when resources were dispersed, turtles have been shown to have a much larger distribution [56]. For immature green turtles within the BIRNM, the required habitats, food, and shelter are tightly clustered within the shallow waters surrounding Buck Island itself, suggesting this MPA provides all the resources immature green turtles require. In addition, these areas are surrounded by less preferred, more exposed, and potentially more dangerous habitats (depths > 10 m), further suggesting constrained turtle space use. Similarly, immature green turtle space use on the neighboring island, Culebra Island, Puerto Rico (< 100 km), also remained in well-defined areas [47]. However, uncharacteristic of Buck Island, Culebra Island has larger protective embayments that immature turtles use almost entirely, even while suitable reef habitats exist directly adjacent to the bays. In both locations, turtles appear to be selecting for lagoon-type habitats at night, potentially for protection, however, absent of the larger protective embayments characteristic of Culebra, turtles within the BIRNM appear to also utilize reef habitats for possible protection and shelter. Blumenthal et al. [52] noted immature green turtles in the Cayman Islands exhibited a similar pattern of space use across lagoon and reef type habitats at night. While both habitat types within the BIRNM may be providing protection and shelter at night, if some immature green turtles forage at night [51], lagoon-type habitats may also provide low risk (i.e., from predation) foraging opportunities as well.
Considering immature green turtles use and have the highest predicted relative selection values near Buck Island, the BIRNM MPA is providing sufficient protection for developmental grounds. Turtle MCPs and KDEs demonstrated space use was largely located directly south of the island and to the east and west, thus, space use did not perfectly match the predicted relative selection probabilities that were observed across a much broader spatial extent. This may be due to tagging effort being generally located south of the island where green turtle abundances were highest, thus, if immature green turtles exhibit small home ranges as demonstrated here and by Brill et al. [54], Makowski et al. [42], Griffin et al. [47], and Chambault et al. [55], we would expect tagged individuals to remain near their capture locations. Further, the RSF model could have been improved with additional covariates that capture additional environmental characteristics (e.g., fetch, wave energy, seagrass total area), ecological processes (e.g., density dependence, predator landscape metrics, [57]), and green turtle foraging strategies. These additional covariates would likely provide further insight into why relative selection values vary across habitat zones and diel periods (e.g., higher selection values in lagoon zones just at night vs. higher selection values in reef zones at both night and day). Ultimately, RSFs are inherently phenomenological and not mechanistic, meaning while space use is an interaction between movement and habitat selection, these RSF predictions are solely focused on observed habitat selection patterns and do not take into account movement processes (e.g., home ranging behavior). These predicted relative selection probabilities are indicative of important habitats in locations that turtles would theoretically select for. Thus, detailed habitat protection (e.g., no anchor, motor and idle only zones, invasive seagrass control) could be focused in areas of space use and in locations with high relative selection probabilities, as indicated by the RSF model.
Consistent with methods provided by Selby et al. [58], we reveal a framework for examining relative habitat selection of animals using acoustic telemetry and RSFs. This analytical approach includes inferring initial space use with calculated COAs, overlaying both COAs and random points onto aggregated habitat information, and using RSF GLMMs to examine resource selection. We then used the trained RSF GLMM model to determine model accuracy and predict relative selection across the greater study area. While this methodology is useful for acoustic telemetry users that wish to examine relative selection and space use beyond areas of detection coverage, it is critical that acoustic arrays are designed to initially capture the representative habitats to reduce biases. In addition, the scaling and selection of habitat classifications are important and should be considered prior to developing models. Important animal space use-habitat relationships may be lost if cell sizes are set too large since the aggregation process selects the dominant habitat type within a cell for classification. One solution is to use a priori ecological knowledge (in our case, using seagrass proximity) to ensure appropriate habitat variables are included in analyses. Further, these analyses focus on relative selection rather than probability of occurrence, meaning that there may be additional variables to explore to improve the accuracy of interpolated space use across an entire area other than RSFs (e.g., integrated step selection functions). Ultimately, RSFs in combination with more traditional analyses (e.g., KDEs) are useful tools to understand habitat selection, space use, and to develop conservation strategies for a given species.
Conclusion
MPAs are on the forefront of spatial management, however, it is critical to formally incorporate information on animal movements to more effectively implement, evaluate, and improve the use of this conservation tool [20, 59,60,61]. Combined with the appropriate analytical methods, acoustic telemetry has become an increasingly important method for resource managers [33]. This technology combined with RSF GLMMs provides new insights in predicting relative habitat selection and space use of a given species, with this application, resource managers within and outside MPAs can refocus enforcement, habitat protection and restoration in areas of need to reach conservation endpoints.
Methods
Study site
BIRNM, managed by the United States National Park Service, was established in 1961 and expanded in 2001. This protected area, located on the northeastern shelf of St. Croix, is a 73.4-km2 no-take marine reserve with small restricted anchoring zone (Additional file 1: Fig. S3). This MPA is one of the few no-take reserves in the Caribbean and one of the oldest [62]. Uninhabited, Buck Island (0.7 km2) is in the center of BIRNM and 2.5 km northeast of St. Croix, USVI. Buck Island and its surrounding waters serve as foraging and/or nesting areas for green, hawksbill, loggerhead (Caretta caretta), and leatherback (Dermochelys coriacea) sea turtles [63]. Within BIRNM, habitats range widely from shallower water to deeper water habitats. From west to east, a high rugosity linear reef surrounds the south side of the island and wraps around Buck Island towards the northwest corner [58]. This reef system encompasses a 50- to 150-m-wide lagoon. Further, the northwest corner and north of the island are characterized by many patch reefs while the south and southwest areas of Buck Island are characterized by low rugosity sea grass patches intermixed with sand flats [64]. Sea grass beds comprised Thalassia sp., Syringodium sp., Halophila sp.
A passive acoustic receiver array, designed to study multiple species [53, 61, 65,66,67,68], at BIRNM was expanded from six to 141 Vemco VR2W receivers (Vemco Amirix Systems, NS, Canada) between 2011 and 2018. Receivers were strategically placed on sand screws and some on cement block anchors around the island and its various habitats with the majority being deployed in < 15 m. Receiver data downloads and maintenance were performed twice a year.
Turtle capture and tagging
Following established protocols [69], green turtles were captured by hand while freediving or by “turtle jumping” rodeo method (Additional file 1: Fig. S2). After capture, turtles were brought aboard the research vessel, where curved (CCL) and straight carapace nuchal (SCLn) and total (SCLt) lengths, width, height, biological samples, were also taken along with photographs to document any anomalies. Each acoustic transmitter, either V16-4L (16 × 88 mm, 24 g in air, 69 kHz, 152 dB, with 30–90 s delay interval, Vemco Amirix Systems, NS, Canada) or V13-1L (13 × 36 mm, 11 g in air, 60–84 kHz, 147 dB, with 30–90 s delay interval, Vemco Amirix Systems, NS, Canada), was attached to individual turtles via a coated wire secured through the right or left marginal carapace scute and by using half a tube of West Marine epoxy [48]. To reduce tagging effects on turtles (e.g., non-normal swimming behavior due to transmitter package), epoxy around the transmitters were molded for optimal hydrodynamics. Here, we also note V16-4Ls are more powerful than V13-1Ls and, theoretically, are able to be detected at greater distances.
Data analysis
Data filtering and organization
We exported turtle detection data from VUE database (Vemco) and analyzed it the R statistical environment (R Development Core Team 2018). Data were first filtered for background detections using the background_detections function (time threshold set at 2700 s) in the glatos package (see https://gitlab.oceantrack.org/GreatLakes/glatos). Subsequently, detections of an individual were removed if the rate of movement exceeded 10 m/s or had a time difference of less than 30 s from its last detection. Tagged turtles often lose their transmitters prior to the expiration of the transmitter’s battery life [48], thus, if a transmitter falls in vicinity of one or more receivers, many background positive detections may occur. Using the detection_events function, from the glatos package, we identified the last “event” in each turtle’s detection log, in this case, a new event was defined as when two consecutive detections are logged across two different receivers. We also modified this function to incorporate distance, for example, we identified events when turtles exceeded 400 m. By examining event abacus detection plots for each individual, we were able to generate conservative cut-off periods to ensure tag failure was not resulting in background positive detections.
For each individual transmitter dataset, we filtered raw detection data and then transformed it into COAs using the mean position algorithm with 60-min bins [70]. COAs were generated with the VTRACK R package [71]; this algorithm calculates the average receiver location for a specified time (e.g., 60-min bins) to obtain fine resolution movement data while minimizing temporal autocorrelation [43, 46]. Using the calculated COAs, full and cumulative monthly MCPs were constructed for each individual transmitter with an adequate amount of tracking data. To generate cumulative monthly MCPs for each individual, an MCP would first be calculated for the first detection month after release. Calculated MCPs for subsequent detection months would be cumulatively added to the first MCP calculated, thus cumulative monthly MCPs. For example, cumulative monthly MCP values only increase when the subsequent detection month’s MCP also increases. Further, the last cumulative monthly MCP in an individual’s detection log will always be equal to the observed full MCP. We also examined the proportion of monthly cumulative MCPs to full MCPs at the individual level and across all individuals for each detection month after release. MCPs were generated with the VTRACK R package [71]. Further, we examined how many transmitters remained active across detection months after release. Here, monthly MCPs were generated at the detection month level, thus, detection months after release refers to the subsequent months an individual was detected excluding any monthly gaps in detection history. For additional analyses, to ensure enough data to model individual turtle as a random effect and following Selby et al. [58], we eliminated individual transmitter datasets with less than 100 COAs. For each individual transmitter dataset, we used KDEs to represent space use. To do this, we first fitted fixed kernel utilization distributions (KUDs) to the COAs, and subsequently estimated the 99%, 95%, and 50% KDEs from each KUD. The KUD represents a bivariate probability density function of use [72, 73], while the KDE is the vectorized polygon that results from drawing isopleths around a percentage of the cumulative utilization distribution. All KUDs and KDEs were derived using the adehabitatHR package [74] and with a 200 m smoothing parameter. Further, for each KDE, the area and the percent overlap to other KDEs were calculated. Here, percent overlap is defined by the proportion of individual is KDE that is overlapped by individual js KDE [75].
RSFs and GLMMs
An RSF is a statistical model that is proportional to the probability of use by an animal [76]. Relative selection strength and, thus, space use of an animal can often be defined by RSFs [77, 78], making them valuable to developing effective conservation strategies [79,80,81]. RSFs allow us to examine the relative selection of individuals based on a given set of covariates. Subsequently, RSFs can be used to help predict habitat selection probabilities outside the observable area based on the selected covariates. To understand relative habitat selection and space use of immature green turtles in the BIRNM, we applied RSFs using GLMMs with a binomial distribution similar to the approach presented by Selby et al. [58].
The response variable included turtle presence locations (COAs), coded as 1 s, and random background points, coded as 0 s. All COAs and random background points were constrained within the available observable area (i.e., where acoustic receivers provided detection coverage). Random background points were randomly placed at an equal number of random locations per individual’s COAs within the available observable area matching the same number of observed diel periods (day vs. night) and observed points per year. To define our available habitat, i.e., detection coverage, across the study area, we generated a 400-m buffer, at any given receiver at a given year (i.e., 2012, 2013, 2014, Additional file 1: Fig. S3). Following Selby et al. [58], we chose a 400-m buffer based on both previous range-testing of the BIRNM array [82] and to retain higher numbers of calculated COAs. All spatial data management and construction were performed using the raster [83], and sp [84] packages.
The calculated COAs (presences) and random background points were collapsed into 200 × 200 m raster cells, along with habitat and depth. Habitat data were provided by and adapted from the National Oceanic and Atmospheric Administration (NOAA) [64]. Benthic habitat classifications were derived using aerial photographs, light detection ranging imagery, and four types of multibeam echosounder imagery [64]. Using the NOAA-derived habitat classifications and their attributes, we generated three variables of interest, distance to seagrass (seagrass defined as > 50% coverage, Fig. 4b), reef type (no reef, patch reef, continuous reef, Additional file 1: Fig. S4), and zone type (lagoon, bank/shelf, reef, other, Fig. 4b). We used the nearest neighbor technique to recalculate depth (Fig. 4b) into 200 × 200 m raster cells, cells with no data available were replaced with the mean values of adjacent cells. All habitat information (factors) and depth (m) values were assigned to each raster cell, using the rasterize function from the raster package [83], and the corresponding COAs and random background points. In addition, diel period (day and night) were assigned to all points using civil dawn and civil dusk times within the maptools package [85]. Variance inflation factors were assessed to determine if variables were too correlated to include in models.
GLMMs were implemented using random subsets of 60% of the dataset. The remaining 40% of the dataset (holdout dataset) was later used to test model accuracy. The full model included diel and depth as an interaction term, diel and distance to seagrass as an interaction term, diel and zone type as an interaction term, and individual and year as random effects. All continuous variables were standardized (z-standardization). Datasets from individuals that had multiple transmitters attached (due to tag loss and subsequent recapture and retagging) were grouped together, respectively. We fit the full model using the glmmTMB package [86] and used the MuMIn package [87] to run all possible subsets, i.e., combination of coefficients. All models included individual and year as random effects. We then used the Akaike information criteria (AIC) to select the top model. Variance components of random effects (year and individual) were also examined in model selection procedures. Using the proportion of data that was not used in the final model (i.e., the random subset of 40%), we assessed the model’s accuracy. Model accuracy was defined as the top model’s ability to correctly predict the probability of 1 s and 0 s using the 40% data subset. Further, model accuracy was assessed across all categorical variables. Marginal effects plots were generated for each conditional term in the final model using the sjPlot package [88]. All other model terms not specified in the given plot were held constant, the continuous variables, depth and distance to seagrass, were held at their means and all other factor variables were held constant at their proportional level rather than at their reference levels.
To predict the relative selection and space use of turtles across BIRNM, we first aggregated all habitat classifications and depth data from the entire study area into multiple raster layers. Subsequently, using the aggregated raster layers, we used the top model to predict and plot the relative selection probabilities across the entire study area for both day and night.
Availability of data and materials
The dataset contains sensitive data containing locations for a protected species. Data are available from corresponding author on reasonable request.
Abbreviations
- AIC:
-
Akaike information criteria
- BIRNM:
-
Buck Island Reef National Monument
- CCL:
-
Curved carapace length
- GLMM:
-
Generalized linear mixed model
- KDE:
-
Kernel density estimation
- KUD:
-
Kernel utilization distribution
- MPA:
-
Marine protected area
- NOAA:
-
National Oceanic and Atmospheric Administration
- RSF:
-
Resource selection function
- SCLn:
-
Straight carapace nuchal length
- SCLt:
-
Straight carapace total length
- MCPs:
-
Minimum convex polygons
References
Millennium Ecosystem Assessment MA. Ecosystems and human well-being: a framework for assessment. Report of the Conceptual Framework Working Group of the Millennium Ecosystem Assessment. 2003.
Quiros TEAL, Croll D, Tershy B, Fortes MD, Raimondi P. Land use is a better predictor of tropical seagrass condition than marine protection. Biol Conserv. 2017;209:454–63.
Vikas M, Dwarakish GS. Coastal pollution: a review. Aquat Procedia. 2015;4:381–8.
Jackson JBC. Reefs since Columbus. Coral Reefs. 1997;16(1):S32.
Board OS, Council NR. Effects of trawling and dredging on seafloor habitat. National Academies Press; 2002.
Erftemeijer PLA, Lewis IIIRRR. Environmental impacts of dredging on seagrasses: a review. Mar Pollut Bull. 2006;52(12):1553–72.
Silliman BR, van de Koppel J, McCoy MW, Diller J, Kasozi GN, Earl K, et al. Degradation and resilience in Louisiana salt marshes after the BP-Deepwater Horizon oil spill. Proc Natl Acad Sci USA. 2012;109(28):11234–9.
Gleason M, McCreary S, Miller-Henson M, Ugoretz J, Fox E, Merrifield M, et al. Science-based and stakeholder-driven marine protected area network planning: a successful case study from north central California. Ocean Coast Manag. 2010;53(2):52–68.
Maxwell SM, Hazen EL, Lewison RL, Dunn DC, Bailey H, Bograd SJ, et al. Dynamic ocean management: defining and conceptualizing real-time management of the ocean. Mar Policy. 2015;58:42–50.
Peel D, Lloyd MG. The social reconstruction of the marine environment: towards marine spatial planning? Town Plan Rev. 2004;75(3):359–78.
Gell FR, Roberts CM. Benefits beyond boundaries: the fishery effects of marine reserves. Trends Ecol Evol. 2003;18(9):448–55.
Lubchenco J, Palumbi SR, Gaines SD, Andelman S. Plugging a hole in the ocean: the emerging science of marine reserves 1. Ecol Appl. 2003;13(sp1):3–7.
Lubchenco J, Grorud-Colvert K. OCEAN. Making waves: the science and politics of ocean protection. Science. 2015;350(6259):382–3.
Spalding MD, Fish L, Wood LJ. Toward representative protection of the world’s coasts and oceans—progress, gaps, and opportunities. Conserv Lett. 2008;1(5):217–26.
Edgar GJ, Stuart-Smith RD, Willis TJ, Kininmonth S, Baker SC, Banks S, et al. Global conservation outcomes depend on marine protected areas with five key features. Nature. 2014;506(7487):216.
Agardy T, di Sciara GN, Christie P. Mind the gap: addressing the shortcomings of marine protected areas through large scale marine spatial planning. Mar Policy. 2011;35(2):226–32.
Bennett NJ, Dearden P. From measuring outcomes to providing inputs: governance, management, and local development for more effective marine protected areas. Mar Policy. 2014;50:96–110.
Gill DA, Mascia MB, Ahmadia GN, Glew L, Lester SE, Barnes M, et al. Capacity shortfalls hinder the performance of marine protected areas globally. Nature. 2017;543(7647):665.
Gallagher AJ, Amon DJ, Bervoets T, Shipley ON, Hammerschlag N, Sims DW. The Caribbean needs big marine protected areas. Science. 2020;367(6479):749.
Cooke SJ. Biotelemetry and biologging in endangered species research and animal conservation: relevance to regional, national, and IUCN Red List threat assessments. Endanger Species Res. 2008;4(1–2):165–85.
Hays GC, Ferreira LC, Sequeira AMM, Meekan MG, Duarte CM, Bailey H, et al. Key questions in marine megafauna movement ecology. Trends Ecol Evol. 2016;31(6):463–75.
Knip DM, Heupel MR, Simpfendorfer CA. Evaluating marine protected areas for the conservation of tropical coastal sharks. Biol Conserv. 2012;148(1):200–9.
Allen AM, Singh NJ. Linking movement ecology with wildlife management and conservation. Front Ecol Evol. 2016;3:1–13.
Lea JSE, Humphries NE, von Brandis RG, Clarke CR, Sims DW. Acoustic telemetry and network analysis reveal the space use of multiple reef predators and enhance marine protected area design. Proc R Soc B Biol Sci. 1834;2016(283):20160717.
Heppell SS, Snover ML, Crowder LB. Sea Turtle Population Ecology. In: Lutz P, Musick J, Wyneken J, editors. Vol. II, The biology of sea turtles. Boc Raton, FL: CRC Press; 2003. p. 275.
Bolten AB. Variation in sea turtle life history patterns: neritic vs. oceanic developmental stages. In: Lutz PL, Musick JA, Wyneken J, editors. The biology of sea turtles, Chap. 9, Vol. 2. The biology of sea turtles. CRC Press; 2003. P. 243–257.
Jones TT, Seminoff JA. Feeding biology: advances from field-based observations, physiological studies, and molecular techniques. In: The biology of sea turtles. Boca Raton: CRC Press; 2013. p. 211–47. (The Biology of Sea Turtles, Volume III; vol. 3).
Hays GC. Good news for sea turtles. Trends Ecol Evol. 2004;19(7):349–51.
Mazaris AD, Schofield G, Gkazinou C, Almpanidou V, Hays GC. Global sea turtle conservation successes. Sci Adv. 2017;3(9):e1600730.
Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, D’Agrosa C, et al. A global map of human impact on marine ecosystems. Science. 2008;319(5865):948–52.
Fuentes MMPB, Gillis AJ, Ceriani SA, Guttridge TL, Bergmann MPMVZ, Smukall M, et al. Informing marine protected areas in Bimini, Bahamas by considering hotspots for green turtles (Chelonia mydas). Biodivers Conserv. 2019;28(1):197–211.
Hamann M, Godfrey MH, Seminoff JA, Arthur K, Barata PCR, Bjorndal KA, et al. Global research priorities for sea turtles: informing management and conservation in the 21st century. Endanger Species Res. 2010;11(3):245–69.
Hussey NE, Kessel ST, Aarestrup K, Cooke SJ, Cowley PD, Fisk AT, et al. ECOLOGY. Aquatic animal telemetry: a panoramic window into the underwater world. Science. 2015;348(6240):1255642.
Scott R, Hodgson DJ, Witt MJ, Coyne MS, Adnyana W, Blumenthal JM, et al. Global analysis of satellite tracking data shows that adult green turtles are significantly aggregated in Marine Protected Areas. Glob Ecol Biogeogr. 2012;21(11):1053–61.
Hart KM, Zawada DG, Fujisaki I, Lidz BH. Habitat use of breeding green turtles Chelonia mydas tagged in Dry Tortugas National Park: making use of local and regional MPAs. Biol Conserv. 2013;161:142–54.
Brownscombe JW, Lédée EJI, Raby GD, Struthers DP, Gutowsky LFG, Nguyen VM, et al. Conducting and interpreting fish telemetry studies: considerations for researchers and resource managers. Rev Fish Biol Fish. 2019;29:369–400.
Donaldson MR, Hinch SG, Suski CD, Fisk AT, Heupel MR, Cooke SJ. Making connections in aquatic ecosystems with acoustic telemetry monitoring. Front Ecol Environ. 2014;12(10):565–73.
Crossin GT, Heupel MR, Holbrook CM, Hussey NE, Lowerre-Barbieri SK, Nguyen VM, et al. Acoustic telemetry and fisheries management. Ecol Appl. 2017;27(4):1031–49.
Griffin LP, Brownscombe JW, Adams AJ, Boucek RE, Finn JT, Heithaus MR, et al. Keeping up with the Silver King: using cooperative acoustic telemetry networks to quantify the movements of Atlantic tarpon (Megalops atlanticus) in the coastal waters of the southeastern United States. Fish Res. 2018;205:65–76.
Ellis RD, Flaherty-Walia KE, Collins AB, Bickford JW, Boucek R, Burnsed SLW, et al. Acoustic telemetry array evolution: from species-and project-specific designs to large-scale, multispecies, cooperative networks. Fish Res. 2019;209:186–95.
Cooke SJ, Iverson SJ, Stokesbury MJW, Hinch SG, Fisk AT, VanderZwaag DL, et al. Ocean Tracking Network Canada: a network approach to addressing critical issues in fisheries and resource management with implications for ocean governance. Fisheries. 2011;36(12):583–92.
Makowski C, Seminoff JA, Salmon M. Home range and habitat use of juvenile Atlantic green turtles (Chelonia mydas L.) on shallow reef habitats in Palm Beach, Florida, USA. Mar Biol. 2006;148(5):1167–79.
Scales KL, Lewis JAPA, Lewis JAPA, Castellanos D, Godley BJ, Graham RT. Insights into habitat utilisation of the hawksbill turtle, Eretmochelys imbricata (Linnaeus, 1766), using acoustic telemetry. J Exp Mar Biol Ecol. 2011;407(1):122–9.
Hazel J, Hamann M, Lawler IR. Home range of immature green turtles tracked at an offshore tropical reef using automated passive acoustic technology. Mar Biol. 2013;160(3):617–27.
Fujisaki I, Hart KM, Sartain-Iverson AR. Habitat selection by green turtles in a spatially heterogeneous benthic landscape in Dry Tortugas National Park, Florida. Aquat Biol. 2016;24(3):185–99.
Chevis MG, Godley BJ, Lewis JP, Lewis JJ, Scales KL, Graham RT. Movement patterns of juvenile hawksbill turtles Eretmochelys imbricata at a Caribbean coral atoll: long-term tracking using passive acoustic telemetry. Endanger Species Res. 2017;32:309–19.
Griffin L, Finn J, Diez C, Danylchuk A. Movements, connectivity, and space use of immature green turtles within coastal habitats of the Culebra Archipelago, Puerto Rico: implications for conservation. Endanger Species Res. 2019;40:75–90.
Smith BJ, Selby TH, Cherkiss MS, Crowder AG, Hillis-Starr Z, Pollock CG, et al. Acoustic tag retention rate varies between juvenile green and hawksbill sea turtles. Anim Biotelemetry. 2019;7(1):1–8. https://doi.org/10.1186/s40317-019-0177-3.
Heithaus MR, Frid A, Wirsing AJ, Dill LM, Fourqurean JW, Burkholder D, et al. State-dependent risk-taking by green sea turtles mediates top-down effects of tiger shark intimidation in a marine ecosystem. J Anim Ecol. 2007;76(5):837–44.
Ogden JC, Robinson L, Whitlock K, Daganhardt H, Cebula R, Odgen JC, et al. Diel foraging patterns in juvenile green turtles (Chelonia mydas L.) in St. Croix United States Virgin Islands. J Exp Mar Biol Ecol. 1983;66(3):199–205.
Taquet C, Taquet M, Dempster T, Soria M, Ciccione S, Roos D, et al. Foraging of the green sea turtle Chelonia mydas on seagrass beds at Mayotte Island (Indian Ocean), determined by acoustic transmitters. Mar Ecol Prog Ser. 2006;306:295–302.
Blumenthal JM, Austin TJ, Bothwell JB, Broderick AC, Ebanks-Petrie G, Olynik JR, et al. Life in (and out of) the lagoon: fine-scale movements of green turtles tracked using time-depth recorders. Aquat Biol. 2010;9(2):113–21.
Casselberry GA, Danylchuk AJ, Finn JT, Deangelis BM, Jordaan A, Pollock CG, et al. Network analysis reveals multispecies spatial associations in the shark community of a Caribbean marine protected area. Mar Ecol Prog Ser. 2020;633:105–26.
Brill RW, Balazs GH, Holland KN, Chang RKC, Sullivan S, George JC. Daily movements, habitat use, and submergence intervals of normal and tumor-bearing juvenile green turtles (Chelonia mydas L.) within a foraging area in the Hawaiian-Islands. J Exp Mar Biol Ecol. 1995;185(2):203–18.
Chambault P, Dalleau M, Nicet J-B, Mouquet P, Ballorain K, Jean C, et al. Contrasted habitats and individual plasticity drive the fine scale movements of juvenile green turtles in coastal ecosystems. Mov Ecol. 2020;8(1):1–15.
Seminoff JA, Resendiz A, Nichols WJ. Home range of green turtles Chelonia mydas at a coastal foraging area in the Gulf of California, Mexico. Mar Ecol Prog Ser. 2002;242:253–65.
McLoughlin PD, Morris DW, Fortin D, vander Wal E, Contasti AL. Considering ecological dynamics in resource selection functions. J Anim Ecol. 2010;79(1):4–12.
Selby T, Hart K, Smith B, Pollock C, Hillis-Starr Z, Oli M. Juvenile hawksbill residency and habitat use within a Caribbean marine protected area. Endanger Species Res. 2019;40:53–64.
Hays GC, Bailey H, Bograd SJ, Bowen WD, Campagna C, Carmichael RH, et al. Translating marine animal tracking data into conservation policy and management. Trends Ecol Evol. 2019.
Costa DP, Breed GA, Robinson PW. New insights into pelagic migrations: implications for ecology and conservation. Annu Rev Ecol Evol Syst. 2012;43:73–96.
Novak AJ, Becker SL, Finn JT, Danylchuk AJ, Pollock CG, Hillis-Starr Z, et al. Inferring residency and movement patterns of horse-eye jack Caranx latus in relation to a Caribbean marine protected area acoustic telemetry array. Anim Biotelemetry. 2020;8:1–13.
Guarderas AP, Hacker SD, Lubchenco J. Current status of marine protected areas in Latin America and the Caribbean. Conserv Biol. 2008;22(6):1630–40.
Hart KM, Iverson AR, Benscoter AM, Fujisaki I, Cherkiss MS, Pollock C, et al. Satellite tracking of hawksbill turtles nesting at Buck Island Reef National Monument, US Virgin Islands: inter-nesting and foraging period movements and migrations. Biol Conserv. 2019;229:1–13.
Costa BM, Tormey S, Battista TA. Benthic habitats of Buck Island Reef National Monument. NOAA Technical Memorandum NOS NCCOS, 142. Silver Spring, MD: NOAA/National Centers for Coastal Ocean Science; 2012.
Bryan DR, Feeley MW, Nemeth RS, Pollock C, Ault JS. Home range and spawning migration patterns of queen triggerfish Balistes vetula in St. Croix, US Virgin Islands. Mar Ecol Prog Ser. 2019;616:123–39.
Becker SL, Finn JT, Novak AJ, Danylchuk AJ, Pollock CG, Hillis-Starr Z, et al. Coarse-and fine-scale acoustic telemetry elucidates movement patterns and temporal variability in individual territories for a key coastal mesopredator. Environ Biol Fishes. 2020;103(1):13–29.
Becker SL, Finn JT, Danylchuk AJ, Pollock CG, Hillis-Starr Z, Lundgren I, et al. Influence of detection history and analytic tools on quantifying spatial ecology of a predatory fish in a marine protected area. Mar Ecol Prog Ser. 2016;562:147–61.
Selby TH, Hart KM, Smith BJ, Pollock CG, Hillis-Starr Z, Oli MK. Juvenile hawksbill residency and habitat use within a Caribbean marine protected area. Endanger Species Rese. 2019;40:53–64.
Service NMF, Fish US, Service W. Recovery plan for US Pacific populations of the East Pacific green turtle (Chelonia mydas). 1998.
Simpfendorfer CA, Heupel MR, Hueter RE. Estimation of short-term centers of activity from an array of omnidirectional hydrophones and its use in studying animal movements. Can J Fish Aquat Sci. 2002;59(1):23–32.
Campbell HA, Watts ME, Dwyer RG, Franklin CE. V-Track: software for analysing and visualising animal movement from acoustic telemetry detections. Mar Freshw Res. 2012;63(9):815–20.
Worton BJ. Kernel methods for estimating the utilization distribution in home-range studies. Ecology. 1989;70(1):164–8.
Lichti NI, Swihart RK. Estimating utilization distributions with kernel versus local convex hull methods. J Wildl Manag. 2011;75(2):413–22.
Calenge C. The package “adehabitat” for the R software: a tool for the analysis of space and habitat use by animals. Ecol Model. 2006;197(3–4):516–9.
Kernohan BJ, Gitzen RA, Millspaugh JJ. Analysis of animal space use and movements. Radio tracking and animal populations. Amsterdam: Elsevier; 2001. p. 125–66.
Manly BFL, McDonald L, Thomas DL, McDonald TL, Erickson WP. Resource selection by animals: statistical design and analysis for field studies. Berlin: Springer Science & Business Media; 2007.
Boyce MS, McDonald LL. Relating populations to habitats using resource selection functions. Trends Ecol Evol. 1999;14(7):268–72.
Nielsen SE, Johnson CJ, Heard DC, Boyce MS. Can models of presence-absence be used to scale abundance? Two case studies considering extremes in life history. Ecography. 2005;28(2):197–208.
Johnson CJ, Seip DR, Boyce MS. A quantitative approach to conservation planning: using resource selection functions to map the distribution of mountain caribou at multiple spatial scales. J Appl Ecol. 2004;41(2):238–51.
Thurfjell H, Ciuti S, Boyce MS. Applications of step-selection functions in ecology and conservation. Mov Ecol. 2014;2(1):4.
Chetkiewicz CB, Boyce MS. Use of resource selection functions to identify conservation corridors. J Appl Ecol. 2009;46(5):1036–47.
Selby TH, Hart KM, Fujisaki I, Smith BJ, Pollock CJ, Hillis-Starr Z, et al. Can you hear me now? Range-testing a submerged passive acoustic receiver array in a Caribbean coral reef habitat. Ecol Evol. 2016;6(14):4823–35.
Hijmans RJ, van Etten J, Cheng J, Mattiuzzi M, Sumner M, Greenberg JA, et al. Package ‘raster.’ R package. 2015.
Pebesma E, Bivand R, Pebesma ME, RColorBrewer S, Collate AAA. Package ‘sp.’ The Comprehensive R Archive Network. 2012.
Bivand R, Lewin-Koh N, Pebesma E, Archer E, Baddeley A, Bearman N, et al. Package ‘maptools.’ 2019.
Magnusson A, Skaug HJ, Nielsen A, Berg C, Kristensen K, Maechler M, et al. glmmTMB: generalized linear mixed models using template model builder. R package version 0.1. 2017.
Barton K, Barton MK. Package. MuMIn. R package version. 2019;1(6).
Lüdecke D. sjPlot: data visualization for statistics in social science. R package version. 2018;2(1).
Acknowledgements
We acknowledge the USGS Natural Resource Protection Program (NRPP) for funding. A. Danylchuk is supported by the National Institute of Food & Agriculture, U.S. Department of Agriculture, the Massachusetts Agricultural Experiment Station, and Department of Environmental Conservation. We also thank Margaret Lamont at USGS for review of an earlier draft of the manuscript along with the valuable input of the anonymous reviewers in earlier drafts. Any use of trade, product, or firm names is for descriptive purposes only and does not imply endorsement by the U.S. Government.
Funding
We acknowledge the USGS Natural Resource Protection Program (NRPP) for funding.
Author information
Authors and Affiliations
Contributions
KH secured funding for and designed the study. LG and BS analyzed the data. LG, KH, and BS interpreted the data and wrote the manuscript. KH and BS provided analytical, interpretation, writing, and editorial oversight. KH, CP, ZHS, MC, AC performed the field work, designed the study, and provided editorial input, and AD, MC, and AC provided writing and editorial input on the manuscript. All authors were major contributors and contributed to writing and editing the final manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Fieldwork was permitted under NMFS permits 16146 and 20315, issued to K. Hart, National Park IACUC USGS-SESC2014-02 and USGS IACUC WARC\GNV 2017-04. Additional permits issued to K. Hart BUIS-2011-SCI-0012; BUIS-2014-SCI-0009; BUIS-2016-SCI-0009.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1.
Additional figures and tables.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Griffin, L.P., Smith, B.J., Cherkiss, M.S. et al. Space use and relative habitat selection for immature green turtles within a Caribbean marine protected area. Anim Biotelemetry 8, 22 (2020). https://doi.org/10.1186/s40317-020-00209-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40317-020-00209-9