
Abstract
Pop-up satellite archival transmitting (PSAT) tags are capable of storing high-resolution behavioral and environmental information for extended periods of time (approximately 1 year), rendering them especially valuable for studying highly mobile species. In this review, we synthesize published PSAT data to understand the biophysical drivers that influence movements of billfishes (families Xiphiidae and Istiophoridae). To date, over 1,080 PSATs have been deployed on billfishes, with individuals demonstrating both trans-equatorial and trans-basin movements. Using this dataset, we identify four main physical variables that drive billfish behavior: temperature, light, oxygen, and complex water mixing (e.g. fronts and eddies). Of the seven species that have been studied with PSAT technology, all exhibited a strong thermal preference for water >22°C, though vertically migrating swordfish additionally occupied waters <10°C while at depth. Ambient light levels influence vertical movements, especially those associated with foraging, as billfish possess large eyes and thermoregulatory abilities that facilitate feeding behaviors below warm surface layers. Mounting evidence suggests that some billfishes actively avoid regions with low dissolved oxygen (<3.5 mL L−1). Human-induced climate change is expected to increase the horizontal and vertical extent of hypoxic water and may further compress habitat and concentrate fishing pressure on pelagic fishes. Finally, complex submeso- and mesoscale processes provide critical habitat for spawning, larval feeding, and retention, but our understanding of these and other behavioral aspects of billfish biology remains limited. Future research efforts should leverage technical advancements while integrating existing and future tag data with chemical and physical oceanographic datasets to gain a better understanding of the relevant biophysical interactions for billfishes, thereby enhancing management capabilities for this ecologically and economically important group of fishes.
Similar content being viewed by others


Background
Tag technologies
Historically, insights into the behavior and ecology of large pelagic fishes in their natural environment were limited to those obtained visually and at the ocean’s surface. These constraints allow access to only a small portion of a species’ behavior, and it has long been recognized that novel approaches are needed to better understand pelagic fishes. Mark-recapture techniques employing external tags have been used extensively since the early twentieth century, but this approach only provides deployment and retrieval locations with no data on a fish’s behavior in the interim [1].
The development of electronic tags enabled researchers to gather a more holistic view of the behaviors of large pelagic organisms. By the 1960s, acoustic technologies were used to actively follow a single-tagged individual for short periods (up to 2 weeks). Longer term datasets became available in the early 1980s with the development of passive acoustic techniques that were well suited to long-term monitoring of high activity regions such as seasonal aggregation sites [2].
Satellite-linked archival tags are the latest development in telemetry research and have demonstrated considerable promise. Since their inception, these tags have been increasingly deployed to understand horizontal and vertical movements [3-6], residency [7], mortality [8], aggregative and feeding behaviors [9], and other important aspects of the biology and ecology of marine organisms. This technology, coupled with large-scale deployment efforts that focus on different species and geographical regions, facilitates extensive data collection. These datasets are amenable to insightful literature reviews and syntheses that can provide valuable information on biophysical drivers and species ecology. While such a review has been performed for sharks [10], a synthesis of the biophysical interactions evident from the numerous satellite tagging studies on billfishes has not yet been undertaken.
Stock status
Herein, we use “billfishes” to apply to the Istiophoridae and the Xiphiidae, two families of large, predatory fishes that inhabit the pelagic realm [11]. Billfishes (suborder Xiphioidei) are distinct from the scombroids [12] and comprise the sailfish and marlins (family Istiophoridae) and swordfish (of the monotypic family Xiphiidae). Ten billfish species have been identified [12]. Three species occur worldwide while three are restricted to the Atlantic, three to the Indo-Pacific, and one to the Mediterranean (Table 1).
Swordfish and several tuna species are major targets of large- and small-scale, directed commercial fisheries around the world. Istiophorids are largely targeted by lucrative directed recreational fisheries, but bycatch from tuna and swordfish pelagic longline (PLL) fisheries is the source of most fishing-induced mortality. Artisanal handline fisheries also yield significant pressure on some species [13]. These compound to result in overfishing of many billfish stocks [14], but stock status varies among and within species and ocean basins.
Historically, commercially valuable fishes, like swordfish, have been overexploited. Yet, despite their economic value, swordfish now appear to be adequately managed throughout the Atlantic and Pacific [15], with North Atlantic stocks exhibiting recovery after years of overfishing[16,17]. However, Mediterranean swordfish stocks are currently both overfished and data deficient [18].
Assessments of blue (Makaira nigricans) and white marlin (Kajikia albida) by the International Commission for the Conservation of Atlantic Tunas (ICCAT) indicate a significant decrease in landings of both species since the 1950s [19]. Catch data and modeling efforts indicate that both populations are below biomass levels for maximum sustainable yield (MSY) and are poorly managed despite robust ICCAT management criteria [20]. Blue marlin have historically been heavily overexploited, largely as bycatch from the PLL fleet [21], and have demonstrated up to a 64% population decline over three generations [15]. Pacific populations of blue marlin are also poorly managed and likely declining [15]. White marlin biomass reached historic lows at the turn of the twenty-first century [22] and was estimated at 15% of maximum sustainable yield (MSY) [23]. This has fostered multiple, though unsuccessful, attempts to list white marlin as threatened or endangered under the U.S. Endangered Species Act.
Sailfish (Istiophorus platypterus) are currently managed as two separate (east and west) stocks in the Atlantic, and both are considered overharvested [24]. Pacific populations are also declining but assessments are data deficient [15]. Due to similar data deficiencies, both black (Istiompax indica) and striped marlin (Kajikia audax) are lacking full stock assessments. Both species are caught as bycatch in other fisheries and are likely declining; striped marlin have declined by >20% globally in three generations [15].
Little biological, ecological, or population-status information is available for the spearfishes: shortbill spearfish (Tetrapturus angustirostris), longbill spearfish (Tetrapturus pfluegeri), roundscale spearfish (Tetrapturus georgii), and Mediterranean spearfish (Tetrapturus belone). Shortbill spearfish are often included in aggregate catch statistics and are thus considered data deficient. Longbill spearfish are not a target of any large-scale fisheries, whereas roundscale spearfish are almost completely unstudied [25]. Mediterranean spearfish are in decline as both a target (e.g. Strait of Messina) [26,27] and non-target species (Aegean Sea swordfish driftnet fishery) [28]. Given that the data-deficient spearfishes are often caught on the same gear as targeted species, they may be experiencing similar declines [29].
One particularly confounding effect of our understanding of white marlin and roundscale spearfish populations in the Atlantic is the recent evidence that some roundscale spearfish have long been misidentified as white marlin [30-32]. The consequences of this misidentification include overestimation of white marlin populations and misunderstanding of both species’ biology, ecology, and harvest.
The paucity of ecological information, including population sizes, movements, and habitat use, for several billfish species is concerning since commercial and recreational fishing pressure remain intense. Improved ecological understanding from data gathered by satellite tags can be used to inform management strategies. Pop-up satellite archival transmitting (PSAT) tag studies studies have provided novel and crucial insights into the ecology of many marine organisms, especially large pelagic fishes (e.g. [3]). In the face of extensive declines of poorly understood, economically important billfish species, a thorough review is warranted to integrate the current literature on satellite tagging. Here, we conduct such a review to synthesize current knowledge on: (1) temperature, (2) light, (3) oxygen, and (4) complex water mixing as biophysical drivers of billfish behavior. In doing so, this analysis highlights the value of PSAT technology and the urgent need for further study on data-deficient species and better spatial coverage for currently studied species. We also contrast the effect of biophysical drivers on swordfish behavior with the istiophorid billfishes. Finally, we provide suggestions for future reporting of PSAT data in the literature to maximize the benefits of inter- and intra-specific data syntheses and comparisons.
Review
Approach and knowledge base
We performed Web of Knowledge and Google Scholar searches in September 2014 using topic and title searches for relevant keywords and common and scientific names for each species [e.g. topic = swordfish AND (“pop-up” OR “pop up”)]. Our search yielded 45 studies since 2001 that have employed PSATs to study billfish (Table 2), peaking at 7 per year in 2011. For those species studied extensively with PSATs, tagging efforts represented a comprehensive size range. Striped marlin, swordfish, and blue marlin dominate the focus of this body of work (cumulatively make up 70% of tag deployments), (Figure 1) while there were no published reports of PSAT studies on roundscale, Mediterranean, or shortbill spearfish. In addition, because roundscale spearfish were described only recently and came after the majority of studies occurred, the possibility exists that published white marlin data include those of roundscale spearfish. For simplicity and practicality, we refer to each species as it was originally identified and reported.

Chronology of billfish pop-up satellite tagging publications. Cumulative frequency distribution of billfish publications over time for the seven species studied with this technology. Species abbreviations are listed in Table 1. Three species (MSP, RSP, SSP) have not been studied with PSAT tags.
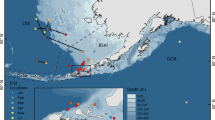
In order to synthesize and quantify overall billfish behavior, we first compiled a database of all published billfish PSAT data. We eliminated redundancy where several publications use the same individual datasets for different purposes (similar to the approach of [74]). We identified 1,082 unique satellite tag deployments yielding 824 unique tag datasets (Table 2). Individual datasets were only plotted in Figure 2 if tag and pop-up locations were given or enough spatial information could be found in the text or a published map to estimate individual displacement based on general tagging location (e.g. eastern Florida coast), overall heading, and mean straight-line distance (MSLD). With these criteria, we plotted 486 individual tag datasets (Figure 2) along with species-specific distribution data [15]. To date, tagging efforts have been concentrated in nearshore regions of Australia, New Zealand, and much of North America. These studies demonstrate displacement up to 5,970 km (mean 586 km, median 329 km) and deployments as long as 411 days (mean 62 days, median 41 days). The wide range in many of these study metrics is a product of the diverse objectives apparent in the reviewed body of literature. Many of the istiophorid studies are characterized by short-term tag deployments to assess post-release mortality, effects of hook type, and other fisheries-specific hypotheses (reviewed in [14,75]), whereas others seek insight into larger scale migratory behaviors.

Movements of PSAT-tagged billfishes. Deployment (green triangles) and pop-up locations (red circles) of 486 billfishes in all pop-up satellite archival transmitting (PSAT) tag studies published between 2001 and 2014. Panels include locations for (a) 108 sailfish, (b) 119 blue marlin, (c) 184 swordfish, (d) 48 white marlin, (e) 2 longbill spearfish, (f) 20 striped marlin, and (g) 5 black marlin. Background polygons show species distribution, and color corresponds to species color in Figure 1. Panels h–j show distributions of shortbill, roundscale, and Mediterranean spearfish (respectively) that have yet to be studied with PSAT technology. Dashed polygons in panels f–g refer to spatial extent of 100+ and 42 tag datasets of striped (f) and black (g) marlin for which individual tracks could not be plotted (see note ‘p’ in Table 2).
Biology
Movements and behavior are inherently coupled to species biology and ecology (e.g. trophic and environmental niches) and are directed by the physiological maintenance of homeostasis in the face of environmental variation [76]. As such, we first provide a brief overview of some of the few known biological aspects of billfishes in order to place our review of PSAT tagging data into the greater context of ecological and biophysical interactions.
In general, billfishes are obligate ram ventilators [77]; they must swim constantly to move oxygenated water over their gills, which have structural modifications that enable respiratory and swimming efficiency benefits. In addition, all billfishes maintain elevated eye and brain temperatures that facilitate cold water foraging without sacrificing visual and cognitive function [78,79]. Common food items among all species include small pelagic fishes and cephalopods [80]. Thorough larval collections in the Straits of Florida [81] indicate that spawning activity for the dominant istiophorids in the Atlantic (sailfish and blue and white marlin) generally occurs from April to October, while swordfish larvae have been collected year-round [81,82]. Other collections have identified billfish larvae from Hawaii, Gulf of California, and the Great Barrier Reef [61,83-85], most of which encompassed several months of the year.
Swordfish
Xiphias gladius have the broadest geographical distribution of all billfishes (Figure 2c) and are found worldwide from 45°S to 45°N [86]. Based on surveys of stomach contents, small pelagic fishes and squid are important prey items [87,88], and swordfish exploit their ability to maintain brain temperatures above ambient water to capture prey in cold, deep waters [89]. Swordfish in the northwest Atlantic appear to spawn primarily south of the Sargasso Sea and east of the Antillean Arc [90]. This may increase advection of larvae and early juveniles toward foraging grounds in the northwest Atlantic by the Gulf Stream [91]. Less consensus has developed in the Pacific, as studies suggest distinct north–south stocks [92], east–west stocks [71], and some stock structure within the western Pacific [93,94].
Blue Marlin
M. nigricans occupy tropical and temperate waters (45°S to 45°N) of the Indo-Pacific and Atlantic Oceans (Figure 2b). Like swordfish, blue marlin are able to forage at depth [23] largely on squid and small pelagic fishes [80]. They are believed to reach maturity at 2 years of age [95] and live up to 20 years [96]. Larvae have been identified near the Dominican Republic [38], the Bahamas [97], and the Florida Straits [46,81], co-occurring in these areas, respectively, with larvae of white marlin, sailfish, and both these species plus swordfish, suggesting that these regions are essential habitat for both spawning adults and developing larvae. Larvae and eggs have also been identified near Hawaii [83].
White Marlin
K. albida occupy temperate and tropical waters of the Atlantic Ocean and adjacent seas and are often found in areas of high food availability including fronts with and without flotsam and marked bathymetric features (e.g. banks, shoals, canyons) (Figure 2d) [98]. They are generally piscivorous, with prey specifically including dolphinfish, herring, and scombrids [80]. Seasonal movements between the northern coast of South America and the Mid-Atlantic Bight are common, and both trans-Atlantic and trans-equatorial crossings have been documented [99]. Spawning is thought to occur in at least five areas in the western North Atlantic, including the Gulf of Mexico, the Mona Passage (Dominican Republic), southwest of Bermuda, Little Bahama Bank near the Abaco Islands, and northwest of Grand Bahama Island [98]. Larvae have been identified in the Mona Passage [38], Florida Straits [81,100], and Gulf of Mexico [98].
Black marlin
I. indica primarily inhabits tropical and subtropical waters of the Indo-Pacific (Figure 2g) where it feeds on cephalopods, small tunas, and other fishes. Like other billfishes, black marlin make extensive seasonal migrations to high latitudes in summer months, where they occasionally occupy temperate waters, and return to low latitudes during winter in both hemispheres. However, only conventional tag data indicates extensive trans-equatorial and trans-Pacific movements [101]; thus, this behavior is still poorly understood. Along these routes, there is evidence of spawning in the South China Sea and Coral Sea, offshore of Taiwan, and near Cairns, Australia [11,61].
Striped marlin
K. audax is a highly migratory Indo-Pacific species found throughout tropical and temperate waters (Figure 2f). It is believed to have a unique distribution among billfishes in the Pacific that stretches from the northwest Pacific to the eastern and southwest Pacific in a horseshoe shape [11]. Shimose et al. [102] suggest that striped marlin migrate between open-ocean feeding habitats and nearshore spawning areas in Hawaii [84] and in the Gulf of California [85]. Limited PSAT data for this species support this migration hypothesis and suggest regional site fidelity in some areas [7]. K. audax is mainly piscivorous and has been shown to feed on a variety of taxa including Molidae, Ostraciidae, Scombridae, and others [102].
Sailfish
I. platypterus is found worldwide in tropical and subtropical waters and is unique amongst the billfishes in its association with near-coastal waters over shallow continental shelves, although catch records suggest a nominal presence in some offshore regions of the tropical Atlantic (Figure 2a) [103]. Larval densities suggest spawning at front edges in the Florida Current [104], and Simms et al. [105] found the highest larval densities at convergent fronts in the northern Gulf of Mexico created by freshwater input and current convergence.
Longbill Spearfish
T. pfluegeri is a rare, relatively small istiophorid with a primarily offshore distribution. It occurs in the Atlantic Ocean and adjacent seas from Georges Bank to South Africa (Figure 2e) [50]. These fish are visual predators that likely feed primarily during the day [106,107] on epipelagic prey [80]; however, little else is known about the biology of this species.
Roundscale Spearfish
T. georgii is widely distributed (Figure 2i) and was initially thought to largely inhabit the western North Atlantic [25,108]. While it was recently found in the central North and South Atlantic [30], no PSAT studies have been conducted to confirm movements within or among these areas. Though somewhat cryptic, the validity of the species among the spearfishes (Genus: Tetrapturus) has been confirmed [12,32]. However, the recent realization that roundscale spearfish have often been characterized as white marlin has led to uncertainty in the collective knowledge of both white marlin and roundscale spearfish—especially since roundscale spearfish may be more abundant than white marlin at particular times in some areas of the North Atlantic [25,31]. In general, however, aside from morphological information from confirmed roundscale spearfish specimens [25], there is, as of yet, little biological or ecological information available for the species.
Mediterranean spearfish
T. belone has the most limited distribution of all spearfishes as it is largely restricted to the Mediterranean Sea (Figure 2j), but occasional catches in the Atlantic have been reported [26]. Although not a target species for most fisheries, it is caught by harpoon in the Straits of Messina, where it is also known to reproduce [26]. Analysis of stomach contents suggests that pelagic fish and cephalopods comprise the diet of this species [109,110]. However, no reports of satellite-tagged Mediterranean spearfish were found, and there is little other information available about the movements and habitat preferences of this species.
Shortbill spearfish
T. angustirostris is a widespread (Figure 2h) but rare, data-deficient species that inhabits tropical to temperate waters of the Indo-Pacific. This species is generally found above the thermocline and well offshore, rarely entering coastal waters (Figure 2h). They are mainly piscivorous and primarily feed on Molidae [102], crustaceans, and cephalopods [11]. Shortbill spearfish eggs have been identified in the equatorial western Indian Ocean [111] and near Kona, Hawaii [83].
Biophysical drivers
Temperature
Temperature controls biochemical reactions and metabolic rates of fishes [112] and is arguably the best understood and most influential environmental variable driving billfish behavior and distribution. PSAT studies for all billfish species report high occupation of narrow thermal regimes with some inter-specific variation in magnitude and range. Most billfishes appear to be largely confined to the warmer side of the 22°C–24°C isotherm [21,34,38,47,50]. In one study, the majority of blue marlin tagged near Bermuda spent >75% of their time in the upper 10 m of the water column where they experienced water temperatures >26°C [21]. Near the Dominican Republic, individuals spent at least 50% of their time in the top 25 m and up to 75% in temperatures 28°C–30°C [38]. Blue marlin tagged in the Caribbean spent more time in cooler waters during the day than at night, with some fish briefly entering waters cooler than 10°C [41]. Similar depth occupation is reported for the slightly smaller white marlin along the United States. Atlantic coast south to Venezuela [38,58]. Sailfish are among the most surface oriented of the billfishes, spending over 80% of their time at surface temperatures during the day and 93% at night in the Atlantic, which is consistent with the behavior reported for sailfish in the Pacific [63,72,73] and for the only two longbill spearfish that have been tagged [50]. Black marlin in the Coral Sea were found to occupy shallow waters warmer than 24°C [60], and their horizontal movements correlated with the spread of 26°C–27°C sea surface temperatures (SST) [61]. Striped marlin are also largely surface oriented, spending 80% of their time in the mixed layer and 72% in the top 5 m [66]. Collectively, heavy use of the surface and mixed layer suggests that a subsurface temperature barrier plays a key role in determining billfish vertical behavior [21,34,58].
Swordfish are the most vertically active billfishes. They typically follow the >22°C isotherm trend during nocturnal occupation but deviate considerably during daytime hours, when they inhabit the upper mesopelagic region (8°C–16°C) before returning to the surface again at sunset, e.g. [55]. This use of a cooler, deep layer may contribute to the occasional and brief daytime basking behavior commonly observed in swordfish tagging studies [54,70].
Diel depth and, consequently, temperature differences are driven by a combination of factors including thermal inertia relative to an individual’s body mass, cardiac limitations, neural warming, foraging opportunities at depth, and light availability required by these visual predators [41,113,114]. Data are currently unavailable on the diel variability of prey consumption for most istiophorids and specifically whether a nocturnal behavioral change is related to feeding; however, variation in swordfish diets has demonstrated predation on deep scattering layer (DSL) organisms [115]. Some studies show epipelagic tuna and billfish species are limited to temperatures within approximately 8°C of SST (e.g. [48,63]) because of the effects of temperature on cardiac function; deeper dwelling tunas and swordfish compensate for their activity in cooler, low-oxygen, subthermocline waters via specific physiological adaptations in their cardiorespiratory system [116-118]. In epipelagic tunas, poor cardiac calcium cycling causes excitation-contraction coupling to break down in cardiac myocytes when instantaneous temperature changes exceed a roughly 8°C instantaneous range [118,119]. Similar processes are presumed to occur in istiophorid hearts [114,116]. Accordingly, forays to depth must be brief and cardiac muscle must thereafter be warmed, resulting in a vertical movement pattern reminiscent of an air-breathing vertebrate in which vertical excursions are followed by surface “recovery” periods [58]. Body size and consequent thermal inertia (i.e. how long the fish retains the heat gained from occupying the mixed layer) influence the variation in dive depths and ambient temperatures that is often observed among individuals and between species [41,48,120]. In contrast, deeper dwelling pelagic fishes, such as swordfish and some tunas, forage extensively below the mixed layer by maintaining cardiac function via greater capacity for cardiac calcium cycling at reduced temperatures [118,121]. Selective heating of the neural infrastructure of the brain and eyes in billfishes and tunas also contribute to their ability to conduct sustained forays into cold water [78,89,122] and provide superior vision for deep foraging [79]. Additionally, surface basking may be a behavioral mechanism for thermoregulation [54,70].
Light
As visual predators, istiophorids and swordfish require adequate ambient light for image formation and foraging. Disproportionately large eyes are characteristic of all billfishes and, combined with selective heating of neural infrastructure as mentioned above, increase the ability to form and neurally process images of moving prey [79]. These adaptations are thus thought to increase prey capture success in cold, dark waters during the day and in the moonlit surface layer at night [68,123]. Consequently, large vertical excursions often occur during daylight periods when illumination at depth is greater.
Swordfish vertical movement is strongly related to light penetration in the Atlantic [52,54,124], Pacific [54,68,70], and Mediterranean Sea [125]. Specifically, maximum daytime depth is significantly correlated with light penetration, and nighttime depth has been shown to increase during the full moon, suggesting that swordfish are feeding on deep scattering layers at depth during the day and vertical migrators in the upper water column at night [52,54,55]. Their large eyes likely provide significant benefits for prey capture in low light conditions [79], and, as such, this species is generally exposed to very limited solar illumination except when basking during the day [54,70].
Although istiophorid billfish also have large eyes that are presumably similarly advantageous in low light conditions, most evidence indicates they are largely surface oriented, especially during daylight hours. Blue marlin spend the majority of time at the surface [23] while making occasional foraging trips below 50 m and give little indication of diel differences in behavior [37,39]. Sailfish exhibit marginal diel changes in vertical movement behavior [48,49,73], with some reports of increased vertical activity during the day than at night [64]. No diel changes are apparent for black or white marlin, aside from occasional qualitative observations of the latter that only persisted for short periods of the tag records [58,60]. While striped marlin in the southwest Pacific largely occupied the top 5 m, they spent significantly more time below 100 m during the day than at night [66]. Thus, although equipped with seemingly adequate vision for deep daytime foraging as observed in swordfish, istiophorids exhibit limited deep foraging behavior presumably due to a subsurface temperature barrier and physiological limitations of functioning in cold water (see section “Temperature” above) .
Oxygen
Dissolved oxygen (DO) also has a substantial influence on billfish behavior and distributions because at low levels it constrains maximum metabolic rates and, thus, metabolic scope [112]. Regions of low dissolved oxygen occupy nearly all of the equatorial Atlantic and eastern tropical Pacific [43,126], and the volume, extent, and severity of these zones are expected to increase with climate change [127]. Although little direct support is available from satellite tags due to their inability (until recently) to measure DO in the water column, compelling evidence exists for some tagged individuals. For example, two longbill spearfish tagged in the South Atlantic appear to have been deterred from areas with DO <3.5 mL L−1 [50]. Furthermore, the mean daily maximum depth of two blue marlin doubled when they left the oxygen minimum layer (OML) region of the eastern tropical Atlantic [43]. Both blue marlin and sailfish occupied the surface layer more extensively in the eastern tropical Atlantic than in the northwestern Atlantic where they traveled deeper and utilized greater depths more often. This behavior correlated with the presence and extent of OMLs that presumably constrained vertical habitat in the eastern tropical Atlantic [43]. As obligate ram ventilators with high metabolic demand and, by extension, high oxygen consumption [128], it is unlikely that billfish can remain in hypoxic waters for an extended period before they incur oxygen debt and require greater DO levels. This may drive avoidance of waters with low DO [129]; however, Dewar et al. [54] report that all 31 Pacific swordfish in their study experienced oxygen levels below 2.15 mL L−1, and Abecassis et al. [70] demonstrate that swordfish can endure severe hypoxia (<0.2 mL L−1). The apparent ability of swordfish to tolerate low DO compared to istiophorids may be at least partially due to differences in gill surface area. Branching of distal gill filaments resulting in an increased number of secondary lamellae is characteristic of swordfish but not found in istiophorids [130]. Blood oxygen binding may also play a role in DO tolerance in billfishes, but its role is thus far unknown.
Current climate change predictions suggest that eutrophication-derived coastal hypoxia and large-scale declines in oxygen levels of tropical waters will result in as much as a 7% decrease in global ocean oxygen content [131,132]. As ocean oxygen content declines and OMLs continue to grow [126], there will be increased habitat compression in both the vertical and horizontal dimensions [120,127]. Stramma et al. [127] suggest that between 1960 and 2010 around 15% of billfish habitat in the upper ocean layer of the tropical northeast Atlantic has been lost to OML expansion. As this expansion continues, restriction of vertical habitat will concentrate billfishes in the surface layers of the Atlantic. At the same time, prey concentrations subject to the same conditions also gather at the surface, which could allow increased billfish growth as a result of increased feeding opportunities and caloric intake. Indeed, Prince and Goodyear [120] hypothesize that sailfish are larger in the eastern Atlantic than in the western Atlantic as a result of habitat compression and increased feeding opportunities. However, with loss of vertical habitat, these fish will be more vulnerable to exploitation by fishing gear [43,120]. Thus, an already overexploited group may experience heightened pressure in the future as available habitat with sufficient oxygen continues to decline, concentrating both prey and fishing effort.
Complex water mixing
Submeso- and mesoscale features are also hypothesized to be a main driver of billfish distributions and behaviors [91]. For example, satellite tracking of swordfish off the southeastern coast of the U.S. showed that these fish were attracted to areas with high-relief bottom structure and the complex thermal structure of fronts that emerge when warm Gulf Stream water meets colder shelf and slope waters near the Charleston Bump [51]. Four of 29 tagged fish stayed in the vicinity of the Bump for extended periods of time where numerous thermal fronts are generated by Gulf Stream deflection and upwelling. Those fish that moved away occupied regions above submarine canyons and seamounts, which also exhibit complex oceanographic processes. Supporting these results, catches of swordfish are high along frontal regions in the Mid-Atlantic Bight off southern New England where Labrador Current waters interact with the Gulf Stream [133], and PSAT data indicate swordfish frequently occupy and traverse this region (Figure 2c). High riverine nutrient inputs also lead to enhanced production along fronts and is often coupled with enhanced frontal upwelling in regions like the Mississippi [134,135] and Amazon river plumes [136], both of which are places of concentrated blue marlin catch [44] and high-density PSAT data (Figure 2b) in the Atlantic.
Larval billfish may have an even stronger relationship with frontal structures as they are often found in the vicinity of major frontal zones and other oceanographic features [137]. Fast growing billfish larvae, which are found in low-latitude, oligotrophic regions with high sea surface temperatures, need concentrated food to fuel their high metabolic demands, and spawning at fronts would serve as a strategy to reduce (even if slightly; cf. [138]) the high-mortality larval phase and increase spawning success. Several regions (mostly in the western North Atlantic) have evidence of relatively high larval densities [38,81,139]. Larval age estimates indicate that adult sailfish spawn during eddy formation, presumably to ensure larvae are retained within a relatively food-rich environment while they develop [104]. An additional, but related, mechanism was proposed by Mourato et al. [140]. The authors describe a dramatic shift in wind direction off the southeastern Brazilian coast during summer that drives strong coastal upwelling and a large intrusion of nutrient rich water onto the continental shelf. This shift often coincides with high adult sailfish densities in the region (although thus far unsupported by PSAT data; Figure 2a) and may constitute an important temporary food source for sailfish larvae. Thus, these areas may provide critical spawning habitat for some large pelagic species [137].
Although there is evidence for billfish association with complex physical features, the technological limitations inherent in PSAT technology (namely, large uncertainty in position estimates) preclude robust, quantitative associations between large pelagic fishes and meso- and submesoscale features. PSAT tags are capable of providing purely objective measurements of biophysical relationships, but several studies demonstrate that conventional light-based geolocation techniques are inadequate due to large spatial error [141,142]. Higher resolution methods are now becoming available [69,143] and should facilitate rapid improvements in this field.
Conclusions
Biophysical interactions
To date, over 1,080 tags have been deployed on billfishes worldwide. Although much of this effort assessed post-release survival (notably the effect of hook type and bycatch mortality) and vulnerability to various fishing gear types with short tag deployments, these studies collectively represent a substantial dataset for drawing inferences about billfish biology and ecology. In this review, we synthesized available information to explain the main biophysical factors influencing billfish distribution and behavior. Many tracked individual billfishes spent significant time in the warm surface layer suggesting a strong thermal utilization of waters >22°C. However, swordfish in particular demonstrate repeated deep vertical movements that suggest foraging at depth where their greater thermal inertia, large eyes, and ability to selectively warm their eye and brain tissues may allow them to maximize capture efficiency and make these movements energetically favorable. Some evidence suggests waters with low dissolved oxygen can influence billfish distribution and movements; however, swordfish appear uniquely capable of associating with low dissolved oxygen concentrations. Tagged billfish also demonstrate considerable occupation of frontal regions that may provide increased adult and larval feeding opportunities and could elicit decreased larval mortality. Current climate change predictions indicate that low oxygen zones will increase in size, which may lead to further habitat compression and thus increased vulnerability of these species. Additionally, species ranges may shift as a result of predicted ocean warming [144].
Future directions and recommendations
Future research efforts and funding paradigms should focus on designing and supporting studies with explicit ecological hypotheses that link PSATs to integrated and real-time oceanographic technologies. This is a grand challenge as PSATs are expensive and studies using them have often been funded to address specific applied management concerns. Specifically, studies should focus on hypotheses related to changing distribution patterns as a result of environmental shifts. For example, understanding whether billfish range expansions are occurring as a result of changing temperature and prey distributions is a key ecological question that PSAT studies could address. Additional PSAT tag deployments will be similarly critical for quantifying the effects of low DO on billfish biology and the implications of future climate scenarios for these species. To this end, new PSAT tag technologies will facilitate both high-resolution locations to understand fine-scale biophysical interactions (e.g. front association) and in situ DO measurements on tagged individuals.
Integration of large volumes of oceanographic data (e.g. ARGO float program) with animal-borne tag records will enable a better understanding of both billfish ecology and oceanographic patterns, especially when one considers that tag-bearing fish are essentially autonomous samplers of the water column. The collective synthesis afforded by such an approach is critical for designing effective management strategies and sustainable harvest. In addition, efforts should be made to expand PSAT deployment to some of the little-studied billfish species (e.g. longbill, roundscale, shortbill, and Mediterranean spearfishes) and those with poor spatial coverage (Figure 2). Improved metrics for species identification should facilitate disambiguating white marlin and roundscale spearfish and allow directed study on each species.
Finally, after extensive review of the PSAT literature, we propose three recommendations from which future comparisons and syntheses could benefit. Specifically, we suggest: (1) if a tag dataset is analyzed or reported on in publications subsequent to its initial publication (e.g. addressing different questions), this should be stated clearly to avoid confusion and facilitate synthesis across studies; (2) reporting a thorough description of tag data and metadata for each tag deployed (when possible) that includes tag identification number (serial, platform terminal transmission, etc.), tag type, deployment and pop-up locations and dates, fish size and sex, days at liberty, depth range, temperature range, and bins used for depth and temperature summary data; and (3) publishing depth and temperature bin data (e.g. time spent in programmed depth intervals) to facilitate comparison among studies and taxa. Most journals allow online supplemental material at no additional cost. This is the perfect venue for publication of additional tag metadata that would further enable synthesis across studies.
References
Kohler NE, Turner PA. Shark tagging: a review of conventional methods and studies. Environ Biol Fish. 2001;60:191–223.
Berumen ML, Braun CD, Cochran JE, Skomal GB, Thorrold SR. Movement patterns of juvenile whale sharks tagged at an aggregation site in the Red Sea. PLoS ONE. 2014;9:e103536.
Thorrold SR, Afonso P, Fontes J, Braun CD, Skomal GB, Berumen ML. Extreme diving behavior in devil rays link surface waters and the deep ocean. Nat Commun. 2014;5:7.
Braun CD, Skomal GB, Thorrold SR, Berumen ML. Diving behavior of the reef manta ray links coral reefs with adjacent deep pelagic habitats. PLoS ONE. 2014;9:e88170.
Werry JM, Planes S, Berumen ML, Lee KA, Braun CD, Clua E. Reef-fidelity and migration of Tiger Sharks, Galeocerdo cuvier, across the Coral Sea. PLoS ONE. 2014;9:e83249.
Skomal GB, Zeeman SI, Chisholm JH, Summers EL, Walsh HJ, McMahon KW, et al. Transequatorial migrations by basking sharks in the western Atlantic Ocean. Curr Biol. 2009;19:1019–22.
Domeier ML. An analysis of Pacific striped marlin (Tetrapturus audax) horizontal movement patterns using pop-up satellite archival tags. Bull Mar Sci. 2006;79:811–25.
Musyl MK, Brill RW, Curran DS, Fragoso NM, McNaughton LM, Nielsen A, et al. Postrelease survival, vertical and horizontal movements, and thermal habitats of five species of pelagic sharks in the central Pacific Ocean. Fish Bull. 2011;109:341–68.
Jorgensen SJ, Arnoldi NS, Estess EE, Chapple TK, Rückert M, Anderson SD, et al. Eating or meeting? Cluster analysis reveals intricacies of White Shark (Carcharodon carcharias) migration and offshore behavior. PLoS ONE. 2012;7:e47819.
Hammerschlag N, Gallagher A, Lazarre D. A review of shark satellite tagging studies. J Exp Mar Biol Ecol. 2011;398:1–8.
Nakamura I. Billfishes of the world: an annotated and illustrated catalogue of marlins, sailfishes, spearfishes and swordfishes known to date. In: Book Billfishes of the world: an annotated and illustrated catalogue of marlins, sailfishes, spearfishes and swordfishes known to date (Editor ed.^eds.), vol. 5. City: FAO; 1985.
Collette BB, McDowell JR, Graves JE. Phylogeny of recent billfishes (Xiphioidei). Bull Mar Sci. 2006;79:455–68.
Arocha F, Ortiz M. Standardized catch rates for blue marlin (Makaira nigricans) and white marlin (Tetrapturus albidus) from the Venezuelan pelagic longline fishery off the Caribbean Sea and the western Central Atlantic: period 1991–2004. ICCAT Coll Vol Sci Pap. 2006;59:315–22.
Graves JE, Horodysky AZ. Asymmetric conservation benefits of circle hooks in multispecies billfish recreational fisheries: a synthesis of hook performance and analysis of blue marlin (Makaira nigricans) postrelease survival. Fish Bull. 2010;108:433–41.
Billfish species status reviews http://www.iucnredlist.org.
Neilson J, Arocha F, Cass-Calay S, Mejuto J, Ortiz M, Scott G, et al. The recovery of Atlantic swordfish: the comparative roles of the regional fisheries management organization and species biology. Rev Fish Sci. 2013;21:59–97.
Collette B, Carpenter K, Polidoro B, Juan-Jordá M, Boustany A, Die D, et al. High value and long life—double jeopardy for tunas and billfishes. Science. 2011;333:291–2.
ICCAT: Report of the. ICCAT Mediterranean swordfish stock assessment meeting. Madrid. 2010;2010:40.
ICCAT. Report of the fourth ICCAT billfish workshop. ICCAT Coll Vol Sci Pap. 2001;53:1–22.
ICCAT. Report of the 2011 blue marlin stock assessment and white marlin data preparatory meeting. Madrid. 2011. p. 71.
Graves JE, Luckhurst BE, Prince ED. An evaluation of pop-up satellite tags for estimating postrelease survival of blue marlin (Makaira nigricans) from a recreational fishery. Fish Bull. 2002;100:134–42.
ICCAT. Report of the 2002 white marlin stock assessment meeting. ICCAT Coll Vol Sci Pap. 2003;55:350–452.
Graves JE, Kerstetter DW, Luckhurst BE, Prince ED. Habitat preferences of istiophorid billfishes in the western North Atlantic: applicability of archival tag data to habitat-based stock assessment methodologies. Collect Vol Sci Pap ICCAT. 2003;55:594–602.
ICCAT. Report of the 2009 sailfish assessment (June 1-5, 2009). Collect Vol Sci Pap ICCAT. 2009;65:1507–632.
Beerkircher L, Lee D, Hinteregger G. Roundscale spearfish Tetrapturus georgii: morphology, distribution, and relative abundance in the western North Atlantic. Bull Mar Sci. 2008;82:155–70.
Di Natale A, Mangano A, Celona A, Valastro M. Size frequency composition of the Mediterranean spearfish (Tetrapturus belone, Rafinesque) catches in the Tyrrhenian Sea and in the Strait of Messina in 2003. ICCAT Coll Vol Sci Pap. 2003;58:589–95.
Di Natale A, Celona A, Mangano A. A series of catch records by the harpoon fishery in the Strait of Messina from 1976 to 2003. Collect Vol Scientif Pap ICCAT. 2005;58:1348–59.
Akyol O, Erdem M, Unal V, Ceyhan T. Investigations on drift-net fishery for swordfish (Xiphias gladius L.) in the Aegean Sea. Turk J Vet Anim Sci. 2005;29:1225–31.
Arocha F, Bárrios A, Lee DW. Spatial-temporal distribution, sex ratio at size and gonad index of white marlin (Tetrapturus albidus) and longbill spearfish (Tetrapturus pfluegeri) in the western Central Atlantic during the period of 2002-2005. Collect Vol Sci Pap ICCAT. 2007;60:1746–56.
Bernard AM, Shivji MS, Domingues RR, Hazin FHV, de Amorim AF, Domingo A, et al. Broad geographic distribution of roundscale spearfish (Tetrapturus georgii) (Teleostei, Istiophoridae) in the Atlantic revealed by DNA analysis: implications for white marlin and roundscale spearfish management. Fish Res. 2013;139:93–7.
Beerkircher L, Arocha F, Barse A, Prince E, Restrepo V, Serafy J, et al. Effects of species misidentification on population assessment of overfished white marlin Tetrapturus albidus and roundscale spearfish T. georgii. Endanger Species Res. 2009;9:81–9.
Shivji MS, Magnussen JE, Beerkircher L, Hinteregger G, Lee DW, Serafy J, et al. Validity, identification, and distribution of the roundscale spearfish, Tetrapturus georgii (Teleoistei: Istiophoridae): morphological and molecular evidence. Bull Mar Sci. 2006;79:483–91.
Matsumoto T, Miyabe N, Saito H, Okazaki M, Chow S. Report of 2000-2001 research cruise by R/V Shoyo-Maru conducted under the ICCAT's BETYP. Col Vol Sci Pap ICCAT. 2002;54:68–90.
Kerstetter DW, Luckhurst BE, Prince ED, Graves JE. Use of pop-up satellite archival tags to demonstrate survival of blue marlin (Makaira nigricans) released from pelagic longline gear. Fish Bull. 2003;101:939–48.
Matsumoto T, Saito H, Miyabe N. Report of observer program for Japanese Tuna Longline Fishery in the Atlantic Ocean from September 2001 to March 2002. Col Vol Sci Pap ICCAT. 2003;55:1679–718.
Matsumoto T, Saito H, Miyabe N. Report of the observer program for the Japanese Tuna Longline Fishery in the Atlantic Ocean from September 2002 to January 2003. Col Vol Sci Pap ICCAT. 2004;56:254–81.
Saito H, Takeuchi Y, Yokawa K. Vertical distribution of Atlantic blue marlin obtained from pop-up archival tags in the tropical Atlantic Ocean. Col Vol Sci Pap ICCAT. 2004;56:201–11.
Prince ED, Cowen RK, Orbesen ES, Luthy SA, Llopiz JK, Richardson DE, et al. Movements and spawning of white marlin (Tetrapturus albidus) and blue marlin (Makaira nigricans) off Punta Cana, Dominican Republic. Fish Bull. 2005;103:659–69.
Saito H, Yokawa K. Use of pop-up tags to estimate vertical distribution of Atlantic blue marlin (Makaira nigricans) released from the commercial and research longline cruise during 2002 and 2003. Col Vol Sci Pap ICCAT. 2006;59:252–64.
Kraus R, Rooker J. Patterns of vertical habitat use by Atlantic blue marlin (Makaira nigricans) in the Gulf of Mexico. Gulf Caribbean Res. 2007;19:89.
Goodyear CP, Luo J, Prince ED, Hoolihan JP, Snodgrass D, Orbesen ES, et al. Vertical habitat use of Atlantic blue marlin Makaira nigricans: interaction with pelagic longline gear. Mar Ecol Prog Ser. 2008;365:233–45.
Hoolihan JP, Luo J, Richardson DE, Snodgrass D, Orbesen ES, Prince ED. Vertical movement rate estimates for Atlantic istiophorid billfishes derived from high-resolution pop-up satellite archival data. Bull Mar Sci. 2009;84:257–64.
Prince ED, Luo J, Phillip Goodyear C, Hoolihan JP, Snodgrass D, Orbesen ES, et al. Ocean scale hypoxia‐based habitat compression of Atlantic istiophorid billfishes. Fish Oceanogr. 2010;19:448–62.
Kraus R, Wells RJD, Rooker J. Horizontal movements of Atlantic blue marlin (Makaira nigricans) in the Gulf of Mexico. Mar Biol. 2011;158:699–713.
Kerstetter D, Graves J. Postrelease survival of sailfish caught by commercial pelagic longline gear in the southern Gulf of Mexico. N Am J Fish Manag. 2008;28:1578–86.
Richardson DE, Cowen RK, Prince ED, Sponaugle S. Importance of the Straits of Florida spawning ground to Atlantic sailfish (Istiophorus platypterus) and blue marlin (Makaira nigricans). Fish Oceanogr. 2009;18:402–18.
Kerstetter D, Bayse S, Graves J. Sailfish (Istiophorus platypterus) habitat utilization in the southern Gulf of Mexico and Florida Straits with implications on vulnerability to shallow-set pelagic longline gear. Collect Vol Sci Pap ICCAT. 2010;65:1701–12.
Hoolihan JP, Luo J, Goodyear C, Orbesen ES, Prince ED. Vertical habitat use of sailfish (Istiophorus platypterus) in the Atlantic and eastern Pacific, derived from pop‐up satellite archival tag data. Fish Oceanogr. 2011;20:192–205.
Kerstetter DW, Bayse SM, Fenton JL, Graves JE. Sailfish habitat utilization and vertical movements in the southern Gulf of Mexico and Florida Straits. Marine Coastal Fish. 2011;3:353–65.
Kerstetter DW, Orbesen ES, Snodgrass D, Prince ED. Movements and habitat utilization of two longbill spearfish Tetrapturus pfluegeri in the eastern tropical South Atlantic Ocean. Bull Mar Sci. 2009;85:173–82.
Sedberry G, Loefer J. Satellite telemetry tracking of swordfish, Xiphias gladius, off the eastern United States. Mar Biol. 2001;139:355–60.
Loefer JK, Sedberry GR, McGovern JC. Nocturnal depth distribution of western north Atlantic swordfish (Xiphias gladius, Linnaeus, 1758) in relation to lunar illumination. Gulf Caribbean Res. 2007;19:83–8.
Neilson JD, Smith S, Royer F, Paul SD, Porter JM, Lutcavage M. Investigations of horizontal movements of Atlantic swordfish using pop-up satellite archival tags. In: Nielsen JL, editor. Tagging and tracking of marine animals with electronic devices. New York: Springer; 2009. p. 145–59.
Dewar H, Prince E, Musyl M, Brill R, Sepulveda C, Luo J, et al. Movements and behaviors of swordfish in the Atlantic and Pacific Oceans examined using pop‐up satellite archival tags. Fish Oceanogr. 2011;20:219–41.
Lerner JD, Kerstetter DW, Prince ED, Talaue-McManus L, Orbesen ES, Mariano A, et al. Swordfish vertical distribution and habitat use in relation to diel and lunar cycles in the western North Atlantic. Trans Am Fish Soc. 2013;142:95–104.
Horodysky AZ, Graves JE. Application of pop-up satellite archival tag technology to estimate postrelease survival of white marlin (Tetrapturus albidus) caught on circle and straight-shank (“J”) hooks in the western North Atlantic recreational fis. Fish Bull. 2005;103:84–96.
Kerstetter DW, Graves JE. Survival of white marlin (Tetrapturus albidus) released from commercial pelagic longline gear in the western North Atlantic. Fish Bull. 2006;104:434–44.
Horodysky AZ, Kerstetter DW, Latour RJ, Graves JE. Habitat utilization and vertical movements of white marlin (Tetrapturus albidus) released from commercial and recreational fishing gears in the western North Atlantic Ocean: inferences from short duration pop-up archival satellite tags. Fish Oceanogr. 2007;16:240–56.
Graves JE, Horodysky AZ. Does hook choice matter? Effects of three circle hook models on postrelease survival of white marlin. N Am J Fish Manag. 2008;28:471–80.
Gunn JS, Patterson TA, Pepperell JG. Short-term movement and behaviour of black marlin Makaira indica in the Coral Sea as determined through a pop-up satellite archival tagging experiment. Mar Freshwater Res. 2003;54:515–25.
Domeier ML, Speare P. Dispersal of adult black marlin (Istiompax indica) from a Great Barrier Reef spawning aggregation. PLoS ONE. 2012;7:e31629.
Prince E, Holts DB, Snodgrass D, Orbesen ES, Luo J, Domeier M, et al. Transboundary movement of sailfish, Istiophorus platypterus, off the Pacific coast of Central America. Bull Mar Sci. 2006;79:827–38.
Chiang W-C, Musyl MK, Sun C-L, Chen S-Y, Chen W-Y, Liu D-C, et al. Vertical and horizontal movements of sailfish (Istiophorus platypterus) near Taiwan determined using pop-up satellite tags. J Exp Mar Biol Ecol. 2011;397:129–35.
Chiang WC, Kawabe R, Musyl M, Sun C, Hung HM, Lin HC, et al. Diel oscillations in sailfish vertical movement behavior in the East China Sea. J Mar Sci Technol. 2013;21:267–73.
Domeier ML, Dewar H, Nasby-Lucas N. Mortality rate of striped marlin (Tetrapturus audax) caught with recreational tackle. Mar Freshwater Res. 2003;54:435–45.
Sippel TJ, Davie PS, Holdsworth JC, Block BA. Striped marlin (Tetrapturus audax) movements and habitat utilization during a summer and autumn in the Southwest Pacific Ocean. Fish Oceanogr. 2007;16:459–72.
Sippel T, Holdsworth J, Dennis T, Montgomery J. Investigating behaviour and population dynamics of striped Marlin (<italic > Kajikia audax</italic>) from the Southwest Pacific Ocean with satellite tags. PLoS ONE. 2011;6:e21087.
Abascal FJ, Mejuto J, Quintans M, Ramos-Cartelle A. Horizontal and vertical movements of swordfish in the Southeast Pacific. Ices J Mar Sci. 2010;67:466–74.
Evans K, Baer H, Bryant E, Holland M, Rupley T, Wilcox C. Resolving estimation of movement in a vertically migrating pelagic fish: Does GPS provide a solution? J Exp Mar Biol Ecol. 2011;398:9–17.
Abecassis M, Dewar H, Hawn D, Polovina J. Modeling swordfish daytime vertical habitat in the North Pacific Ocean from pop-up archival tags. Mar Ecol Prog Ser. 2012;452:219–36.
Evans K, Abascal F, Kolody D, Sippel T, Holdsworth J, Maru P. The horizontal and vertical dynamics of swordfish in the South Pacific Ocean. J Exp Mar Biol Ecol. 2014;450:55–67.
Hoolihan J. Horizontal and vertical movements of sailfish (Istiophorus platypterus) in the Arabian Gulf, determined by ultrasonic and pop-up satellite tagging. Mar Biol. 2005;146:1015–29.
Hoolihan JP, Luo J. Determining summer residence status and vertical habitat use of sailfish (Istiophorus platypterus) in the Arabian Gulf. ICES J Mar Sci: J du Conseil. 2007;64:1791–9.
Musyl M, Domeier M, Nasby-Lucas N, Brill RW, McNaughton LM, Swimmer J, et al. Performance of pop-up satellite archival tags. Mar Ecol Prog Ser. 2011;433:1–28.
Graves JE, Horodysky AZ. Challenges of estimating post-release mortality of istiophorid billfishes caught in the recreational fishery: a review. Fish Res. In press.
Claireaux G, Lefrancoise C. Linking environmental variability and fish performance: integration through the concept of scope for activity. Philos Trans R Soc B. 2007;362:2031–41.
Wegner NC, Sepulveda CA, Aalbers SA, Graham JB. Structural adaptations for ram ventilation: gill fusions in scombrids and billfishes. J Morphol. 2013;274:108–20.
Block BA, Finnerty JR, Stewart AF, Kidd J. Evolution of endothermy in fish: mapping physiological traits on a molecular phylogeny. Science. 1993;260:210.
Fritsches KA, Brill RW, Warrant EJ. Warm eyes provide superior vision in swordfishes. Curr Biol. 2005;15:55–8.
Júnior TV, Vooren CM, Lessa RP. Feeding habits of four species of Istiophoridae (Pisces: Perciformes) from northeastern Brazil. Environ Biol Fish. 2004;70:293–304.
Richardson DE, Llopiz JK, Guigand CM, Cowen RK. Larval assemblages of large and medium-sized pelagic species in the Straits of Florida. Prog Oceanogr. 2010;86:8–20.
Govoni JJ, Stender BW, Pashuk O. Distribution of larval swordfish, Xiphias gladius, and probable spawning off the southeastern United States. Fish Bull. 2000;98:64–74.
Hyde J, Lynn E, Humphreys Jr R, Musyl M, West A, Vetter R. Shipboard identification of fish eggs and larvae by multiplex PCR, and description of fertilized eggs of blue marlin, shortbill spearfish, and wahoo. Mar Ecol Prog Ser. 2005;286:269–77.
Hyde JR, Humphreys R, Musyl M, Lynn E, Vetter R. A central North Pacific spawning ground for striped marlin, Tetrapturus audax. Bull Mar Sci. 2006;79:683–90.
Gonzalez-Armas R, Klett-Traulsen A, Hernandez-Herrera A. Evidence of billfish reproduction in the southern Gulf of California, Mexico. Bull Mar Sci. 2006;79:705–17.
Palko BJ, Beardsley GL, Richards WJ. Synopsis of the biology of the swordfish, Xiphias gladius Linnaeus. NOAA Technical Report 1981:28.
Scott W, Tibbo S. Food and feeding habits of swordfish, Xiphias gladius, in the western North Atlantic. J Fish Board Canada. 1968;25:903–19.
Stillwell C, Kohler N. Food and feeding ecology of the swordfish Xiphias gladius in the western North Atlantic Ocean with estimates of daily ration. Mar Ecol Progress Ser Oldendorf. 1985;22:239–47.
Carey F. A brain heater in the swordfish. Science. 1982;216:1327–9.
Arocha F, Lee D. Maturity at size, reproductive seasonality, spawning frequency, fecundity and sex ratio in swordfish from the Northwest Atlantic. Collect Vol Sci Pap ICCAT. 1995;45:350–7.
Olson DB, Hitchcock GL, Mariano AJ, Ashjian CJ, Peng G, Nero RW, et al. Life on the edge: marine life and fronts. Oceanography. 1994;7:52–60.
Sakagawa GT, Bell RR. Swordfish, Xiphias gladius. In: Shomura RS, editor. Summary report of the billfish stock assessment workshop; pacific resources: Honolulu Laboratory, Southwest Fisheries Center, Honolulu, Hawaii, 5-14 December 1977. Seattle, Washington: US Department of Commerce; 1980. p. 43–55.
Grall C, De Sylva DP, Houde ED. Distribution, relative abundance, and seasonality of swordfish larvae. Trans Am Fish Soc. 1983;112:235–46.
Nishikawa Y, Honma M, Ueyanagi S, Kikawa S. Average distribution of larvae of oceanic species of scombrid fishes, 1956-1981. Far Seas Fish Res Lab S Ser. 1985;12:1–99.
Prince E, Lee D, Zweifel J, Brothers E. Estimating age and growth of young Atlantic blue marlin Makaira nigricans from otolith microstructure. Fish Bull. 1991;89:441–59.
Wilson CA, Dean JM, Prince ED, Lee DW. An examination of sexual dimorphism in Atlantic and Pacific blue marlin using body weight, sagittae weight, and age estimates. J Exp Mar Biol Ecol. 1991;151:209–25.
Serafy JE, Cowen RK, Paris CB, Capo TR, Luthy SA. Evidence of blue marlin, Makaira nigricans, spawning in the vicinity of Exuma Sound, Bahamas. Mar Freshwater Res. 2003;54:299–306.
White Marlin Biological Review Team. Atlantic white marlin status review. Report to National Marine Fisheries Service, Southeast Regional Office, December 10, 2007. p. 88.
Orbesen ES, Hoolihan JP, Serafy JE, Snodgrass D, Peel E, Prince E. Transboundary movement of Atlantic istiophorid billfishes among international and US domestic management areas inferred from mark-recapture studies. Mar Fish Rev. 2008;70:14–23.
Luthy SA, Cowen RK, Serafy JE, McDowell JR. Toward identification of larval sailfish (Istiophorus platypterus), white marlin (Tetrapturus albidus), and blue marlin (Makaira nigricans) in the western North Atlantic Ocean. Fish Bull. 2005;103:588–600.
Ortiz M, Prince E, Serafy J, Holts DB, Davy KB, Pepperell JG, et al. Global overview of the major constituent-based billfish tagging programs and their results since 1954. Mar Freshwater Res. 2003;54:489–507.
Shimose T, Yokawa K, Saito H. Habitat and food partitioning of billfishes (Xiphioidei). J Fish Biol. 2010;76:2418–33.
Uozumi Y, Nakano H. A historical review of Japanese longline fishery and billfish catches in the Atlantic Ocean. Col Vol Sci Pap ICCAT. 1994;41:233–43.
Richardson DE, Llopiz JK, Leaman KD, Vertes PS, Muller-Karger FE, Cowen RK. Sailfish (Istiophorus platypterus) spawning and larval environment in a Florida Current frontal eddy. Prog Oceanogr. 2009;82:252–64.
Simms JR, Rooker JR, Holt SA, Holt GJ, Bangma J. Distribution, growth, and mortality of sailfish (Istiophorus platypterus) larvae in the northern Gulf of Mexico. Fish Bull. 2010;108:478–90.
Witzell W. Longbill spearfish, Tetrapturus pfleugeri, incidentally caught by recreational billfishermen in the western North Atlantic Ocean, 1974-1986. Fish Bull. 1989;87:982–4.
Yokawa K, Saito H. Results of comparison of catch ratio between shallow and deep setting obtained from 2002 Shoyo-maru survey in the tropical Atlantic. Col Vol Sci Pap ICCAT. 2004;56:195–200.
Garcia S. World review of highly migratory species and straddling stocks. Rome: FAO; 1994.
Castriota L, Finoia MG, Campagnuolo S, Romeo T, Potoschi A, Andaloro F. Diet of Tetrapturus belone (Istiophoridae) in the central Mediterranean Sea. J Mar Biol Assoc UK. 2008;88:183–7.
Romeo T, Consoli P, Castriota L, Andaloro F. An evaluation of resource partitioning between two billfish, Tetrapturus belone and Xiphias gladius, in the central Mediterranean Sea. J Mar Biol Assoc U K. 2009;89:849–57.
Collette BB, Cole K. Reproduction and development in epipelagic fishes. In: Reproduction and sexuality in marine fishes: patterns and processes. Berkeley: University of California Press; 2010. p. 21–63.
Fry FEJ. Effects of environment on animal activity. Univ Toronto Biol Ser. 1947;55:1–62.
Brill RW, Block BA, Boggs CH, Bigelow KA, Freund EV, Marcinek DJ. Horizontal movements and depth distribution of large adult yellowfin tuna (Thunnus albacares) near the Hawaiian Islands, recorded using ultrasonic telemetry: implications for the physiological ecology of pelagic fishes. Mar Biol. 1999;133:395–408.
Brill RW, Lutcavage M. Understanding environmental influences on movements and depth distributions of tunas and billfishes can significantly improve population assessments. Am Fish Sci Symp. 2001;25:179–98.
Varghese S, Somvanshi VS, Dalvi R. Diet composition, feeding niche partitioning and trophic organisation of large pelagic predatory fishes in the eastern Arabian Sea. Hydrobiologia. 2014;736:99–114.
Bernal D, Sepulveda C, Musyl M, Brill R. The eco-physiology of swimming and movement patterns of tunas, billfishes, and large pelagic sharks. In: Domenici P, Kapurr D, editors. Fish Locomotion - an etho-ecological perspective. Enfield, New Hampshire: Scientific Publishers; 2009.
Brill RW, Lowe TE, Cousins KL. How water temperature really limits the vertical movements of tunas and billfishes–it's the heart stupid, International Congress on Biology of Fish. Towson University: American Fisheries Society; 1998. p. 4.
Galli GL, Shiels HA, Brill RW. Temperature sensitivity of cardiac function in pelagic fishes with different vertical mobilities: yellowfin tuna (Thunnus albacares), bigeye tuna (Thunnus obesus), mahimahi (Coryphaena hippurus), and swordfish (Xiphias gladius). Physiol Biochem Zool. 2009;82:280–90.
Shiels HA, Di Maio A, Thompson S, Block BA. Warm fish with cold hearts: thermal plasticity of excitation-contraction coupling in bluefin tuna. Proc Biol Sci. 2011;278:18–27.
Prince ED, Goodyear CP. Hypoxia‐based habitat compression of tropical pelagic fishes. Fish Oceanogr. 2006;15:451–64.
Galli GL, Lipnick MS, Shiels HA, Block BA. Temperature effects on Ca2+ cycling in scombrid cardiomyocytes: a phylogenetic comparison. J Exp Biol. 2011;214:1068–76.
Block BA. Structure of the brain and eye heater tissue in marlins, sailfish, and spearfishes. J Morphol. 1986;190:169–89.
Hays G. Large-scale patterns of diel vertical migration in the North Atlantic. Deep-Sea Res I Oceanogr Res Pap. 1996;43:1601–15.
Carey FG, Robison BH. Daily patterns in the activities of swordfish, Xiphias gladius, observed by acoustic telemetry. Fish Bull. 1981;79:277–92.
Canese S, Garibaldi F, Orsi Relini L, Greco S. Swordfish tagging with pop-up satellite tags in the Mediterranean Sea. Collect Vol Sci Pap ICCAT. 2007;62:1052–7.
Stramma L, Johnson GC, Sprintall J, Mohrholz V. Expanding oxygen-minimum zones in the tropical oceans. Science. 2008;320:655–8.
Stramma L, Prince ED, Schmidtko S, Luo J, Hoolihan JP, Visbeck M, et al. Expansion of oxygen minimum zones may reduce available habitat for tropical pelagic fishes. Nat Clim Change. 2012;2:33–7.
Brill RW. Selective advantages conferred by the high performance physiology of tunas, billfishes, and dolphin fish. Comp Biochem Physiol. 1996;113:3–15.
Randall D. Gas exchange in fishes. In: Hoar W, Randall D, editors. Fish physiology, vol. 4. New York: Academic; 1970. p. 253–92.
Wegner NC, Sepulveda CA, Bull KB, Graham JB. Gill morphometrics in relation to gas transfer and ram ventilation in high‐energy demand teleosts: Scombrids and billfishes. J Morphol. 2010;271:36–49.
Bijma J, Portner HO, Yesson C, Rogers AD. Climate change and the oceans–what does the future hold? Mar Pollut Bull. 2013;74:495–505.
Keeling RF, Kortzinger A, Gruber N. Ocean deoxygenation in a warming world. Ann Rev Mar Sci. 2010;2:199–229.
Podestá GP, Browder JA, Hoey JJ. Exploring the association between swordfish catch rates and thermal fronts on US longline grounds in the western North Atlantic. Cont Shelf Res. 1993;13:253–77.
Müller-Karger FE, Walsh JJ, Evans RH, Meyers MB. On the seasonal phytoplankton concentration and sea surface temperature cycles of the Gulf of Mexico as determined by satellites. J Geophys Res Oceans. 1991;96:12645–65.
Walsh JJ, Dieterle DA, Meyers MB, Müller-Karger FE. Nitrogen exchange at the continental margin: a numerical study of the Gulf of Mexico. Prog Oceanogr. 1989;23:245–301.
Dagg M, Benner R, Lohrenz S, Lawrence D. Transformation of dissolved and particulate materials on continental shelves influenced by large rivers: plume processes. Cont Shelf Res. 2004;24:833–58.
Bakun A. Fronts and eddies as key structures in the habitat of marine fish larvae: opportunity, adaptive response and competitive advantage. Sci Mar. 2006;70:105–22.
Houde ED. Fish early life dynamics and recruitment variability. Am Fish Soc Symp Ser. 1987;2:17–29.
Llopiz JK, Cowen RK. Precocious, selective and successful feeding of larval billfishes in the oceanic Straits of Florida. Mar Ecol Prog Ser. 2008;358:231.
Mourato BL, Hazin F, Bigelow K, Musyl M, Carvalho F, Hazin H. Spatio-temporal trends of sailfish, Istiophorus platypterus, catch rates in relation to spawning ground and environmental factors in the equatorial and southwestern Atlantic Ocean. Fish Oceanogr. 2014;23:32–44.
Lam CH, Nielsen A, Sibert JR. Improving light and temperature based geolocation by unscented Kalman filtering. Fish Res. 2008;91:15–25.
Sibert JR, Musyl MK, Brill RW. Horizontal movements of bigeye tuna (Thunnus obesus) near Hawaii determined by Kalman filter analysis of archival tagging data. Fish Oceanogr. 2003;12:141–51.
Holdsworth J, Sippel T, Block B. Near real time satellite tracking of striped marlin (Kajikia audax) movements in the Pacific Ocean. Mar Biol. 2009;156:505–14.
Robinson LM, Hobday AJ, Possingham HP, Richardson AJ. Trailing edges projected to move faster than leading edges for large pelagic fish habitats under climate change, Deep Sea Research Part II: Topical Studies in Oceanography. 2014.
Acknowledgements
AZH was supported by the NOAA Living Marine Resources Cooperative Science Center (#NA11SEC4810002) and the National Science Foundation Educational Partnership in Climate Change and Sustainability (NSF-HRD 1137465). JKL was supported by the Woods Hole Oceanographic Institution’s Ocean Life Institute, Academic Programs Office, and the Penzance Endowed Fund in Support of Assistant Scientists.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
CDB and MBK performed the literature review. CDB, MBK, AZH, and JKL wrote the manuscript. CDB created the figures and conducted analyses. All authors read and approved the final manuscript.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Braun, C.D., Kaplan, M.B., Horodysky, A.Z. et al. Satellite telemetry reveals physical processes driving billfish behavior. Anim Biotelemetry 3, 2 (2015). https://doi.org/10.1186/s40317-014-0020-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40317-014-0020-9