
Abstract
The Neolithic revolution—the transition of our species from hunter and gatherer to cultivator—began approximately 14,000 years ago and is essentially complete for macroscopic food. Humans remain largely pre-Neolithic in our relationship with microbes but starting with the gut we continue our hundred-year project of approaching the ability to assess and cultivate benign microbiomes in our bodies. Buildings are analogous to the body and it is time to ask what it means to cultivate benign microbiomes in our built environment. A critical distinction is that we have not found, or invented, niches in buildings where healthful microbial metabolism occurs and/or could be cultivated. Key events affecting the health and healthfulness of buildings such as a hurricane leading to a flood or a burst pipe occur only rarely and unpredictably. The cause may be transient but the effects can be long lasting and, e.g., for moisture damage, cumulative. Non-invasive “building tomography” could find moisture and “sentinel microbes” could record the integral of transient growth. “Seed” microbes are metabolically inert cells able to grow when conditions allow. All microbes and their residue present actinic molecules including immunological epitopes (molecular shapes). The fascinating hygiene and microbial biodiversity hypotheses propose that a healthy immune system requires exposure to a set of microbial epitopes that is rich in diversity. A particular conjecture is that measures of the richness of diversity derived from microbiome next-generation sequencing (NGS) can be mechanistically coupled to—rather than merely correlated with some measures of—human health. These hypotheses and conjectures inspire workers and funders but an alternative is also consequent to the first Neolithic revolution: That the genetic uniformity of contemporary foods may also decrease human exposure to molecular biodiversity in a heath-relevant manner. Understanding the consequences—including the unintended consequences of the first Neolithic revolution—will inform and help us benignly implement the second—the microbial—Neolithic revolution.
Similar content being viewed by others


Background
This article is situated in the context of efforts to encourage creative interdisciplinary collaborations among architects, building engineers, chemists, immunologists, epidemiologists, physicians, and microbiologists. This review and commentary was stimulated by the author’s participation in the Sloan symposium: Healthy buildings 2015-Europe whose summary in this special issue of the journal microbiome contains the telling statement “There was general consensus that while the applied microbiology developments emerging in this research community—first and foremost, DNA recovery methodology and in particular, next-generation sequencing—have had notable impacts as judged by common academic metrics; however, these advances have not successfully translated into paths which are available for practitioners to apply such methods or interpret these results with confidence in the field.” [1]. Despite the courteous language of scholarship as well as grammatical imprecision, the message is clear: Attaining relevance for this incipient and promising field is not assured. By hypothesis, our highest probability path to attaining relevance requires identifying and occasionally suggesting extensions and alternatives to currently favored ideas and approaches. Contradictions in this article are not about matters of fact; they arise from considering alternative ideas for how microbiome research can contribute to understanding and enhancing the built environment’s effects on human health. Tables 1 and 2 tend toward pedagogy; they are given in the spirit of friendly interdisciplinary invitation. Microbiome, microbiotia, and related terms in this document refer to “little itty bitty living things,” i.e., inclusive of eubacteria, archea, fungi, protists, and cells of multicellular differentiated organisms (animals and plants in common usage) that are present in the environment detached from the main body; it is also inclusive of all phage and viruses.
Précis
-
1.
Humans have actively cultured plants and animals for over 10,000 years and have received benefits from such endeavors (Neolithic revolution). Important progress is currently being made in the century-long project of understanding and culturing benign metabolically active gut microbiomes.
-
2.
The analogy of buildings and the human body suggest a potential for benign microbiomes in buildings. Related ideas proposed here include (a) building tomography for the non-invasive detection of moisture, (b) sentinel microbes, (c) seed microbes on moisture-vulnerable internal surfaces, and (d) seed biofilms in outflow plumbing.
-
3.
Actionable wisdom for building practitioners (architects, building engineers, remediation specialists) has not followed from DNA recovery and next-generation sequencing (NGS) in contrast to the still-essential contributions of classical microbiology. One bottleneck in NGS relevance is that current applications do not differentiate among meanings of the term “microbiome.” These distinctions offer a route to relevance and appear technically challenging but within reach.
-
4.
The hygiene and microbial biodiversity hypotheses have merged with NGS to suggest that microbial sequence diversity provides a measure of health. This idea is both ingenious and inspiring, but it may be wrong. Aspects of hygiene-biodiversity hypotheses are examined: (a) A portion of unique sequence found in microbiome NGS studies may never have existed in living cells. (b) There may be not-yet-found keys to simplify today’s apparently irreducible complexity. (c) Non-microbial sources of epitope diversity may complement and perhaps supersede the relative contribution of changes in microbial diversity to human health.
-
5.
An unintended consequence of the first Neolithic revolution makes it likely that humans are exposed to less epitope variation in food. By hypothesis, uniformity of foodborne epitopes may also contribute to vulnerabilities in health. The quantitative and qualitative distinctions and interactions of food and microbial epitope exposure merit study.
The microbial Neolithic revolution
The Neolithic revolution—when our species transitioned from hunter and gatherer to domesticator and cultivator—is almost complete with respect to macroscopic food and can now become science-based stewardship [2]. In contrast, humans remain hunter-gatherers with respect to the microbial world in which our species is embedded. Our microbiomes have no doubt changed due to changes in civilization (e.g., agriculture and urbanization) but this has been unintentional and is in contrast to the deliberate ways that hunting and gathering have transformed into a deliberate and specific macroscopic agriculture of specified plants and animals. The beginnings of more deliberate microbial Neolithic transition are underway with respect to bodily, especially the gut, microbiomes. However, regarding the external and internal surfaces of our buildings and our clothes, humans remain hunter-gatherers and often attempted microbial genocidists [3]. The problem, and the opportunity, is that we do not live in a sterile world. Just as nature abhors a vacuum, the living world abhors sterility. To the extent we succeed in sterilizing them, surfaces and substances are uniquely available for opportunistic microbes. Biocidal agents themselves can be directly or indirectly hazardous to human health [4] sometimes in surprising ways such as promoting tolerance to and the evolution against clinical antibiotics [5]. As one alternative or complementary approach, we can search for and create opportunities for introducing and possibly even cultivating benign microbiota in our built environments. We must also be honest critics and skeptics about the nature of the unknowns and the possibility of unintended consequences. Our gut microflora has become the exemplar against which other hypotheses of health-promoting microbiomes may be compared. Prebiotics and specific inoculation to optimize the gut microflora are now clinically relevant though not yet widely practiced [6], an idea that has been around with varying degrees of acceptance and success for over a hundred years [7].
Analogies of microbiomes in buildings and bodies
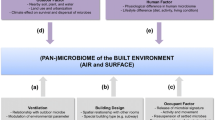
Buildings can be analogized to bodies and bodily microbiomes analogized to microbiomes in the built environment. Where do the ideas fit and where do they fail? Table 1 elaborates the analogy of a human body to a building [8] and candidate analogies of microbiomes in each. Future possibilities for deliberate inoculation in the built environment include surfaces such as the exterior and interior of walls, pipes, textiles such as carpets, furniture, and clothes. Moist and wet surfaces including pipes especially for outflow are candidates for deliberate cultivation of a benign and helpful microflora.
A weak point of this analogy is the apparently different role played by active microbial metabolism and growth. The fact of periodic defecation is proof that microbes in the gut actively metabolize and grow. Renewal proves growth and growth proves metabolism. There is an abundant, rapidly growing, and important literature (that will not be reviewed here) that strongly implies that “the right” gut microbiome contributes to healthful development and function of the organism. In contrast, there are no definitive health-positive examples of active metabolism and growth of microbes in buildings (other than that in or on the occupants).
Defining a microbiome
The word “microbiome” is routinely used to describe several distinct entities (Table 3). Conflation of microbiome types limits the value that can be gained from interpretation of sequence data. The microbiome research community is making an effort to standardize protocols for DNA extraction and purification as well as the NGS processing pipeline. Unfortunately, the currently recommended protocols [9] do not distinguish types of microbiomes (Table 3). Methods appear available (Table 3 legend) but are not widely validated or applied. Approaches that distinguish among the metabolically active, the potentially active, dead, and extracellular DNA would likely enhance the relevance of NGS to all aspects of microbial ecology and microbiome analysis. Until microbiome types are distinguished in NGS analyses, practical workers concerned with identifying sick buildings and performing biological remediation will probably remain wise to favor classical microbiological and microbial ecology approaches [10, 11]. The hygiene hypothesis is discussed in a subsequent section but a connection here is self evident to biologists but may not be to architects and building engineers: The metabolic state of microbes determines which of the microbial compounds and immunological epitopes encoded by the DNA and RNA sequences of bacteria, fungi, protists, or human cells are actually synthesized.
Growing health-promoting microbes in buildings
Health-positive roles for a metabolically active building microbiome may be awaiting our discovery or invention. Candidates worth investigating include the following: (a) Actinomycetes are ubiquitous in building walls [12]. With their complex and varied secondary metabolism, actinomycetes have long been the source of novel compounds including antibiotics and antifungals [13]. There is good evidence that some species of actinomycetes create toxic products that can interact synergistically with fungi to the detriment of occupant health [14, 15]. By hypothesis, there may also exist species and strains whose metabolic products inhibit fungi associated with building-related symptoms (BRS—more popularly known as sick building syndrome, SBS) and/or specific diseases that can be transmitted via the built environment. (b) Biofilms and planktonic growth often occur in plumbing and HVAC systems. These may provide an opportunity for intentional and knowledge-based cultivation that at least occupies the niche where random inoculation occasionally leads to niche occupation by microbial pathogens [16–18]. In infants, the first inoculated microbiome has long-term effects [19]. By hypothesis, the same will prove true for water outflow pipes. The relevant time scale of buildings can be short, e.g., mold can grow in a few days after a flood but the scale includes years, decades, and in some cases centuries. The longer time scales differ from laboratory experiments and the usual ecological contexts, e.g., soil and fecal, that because they are familiar, tend to provide the mental frame for thinking about microbiology in the built environment but in the context of geomicrobiology, doubling times of centuries or even millennia have been estimated [20].
Water is a fundamental limiting factor for microbial growth. The moisture content and distribution in buildings is complex in both space and time [21]. Non-invasive tomographic quantification of moisture in buildings could become a breakthrough technology in locating microbial growth. It is proposed here that the microwave absorption and reflection properties of water [22, 23] could be used to map moisture throughout buildings including spaces (e.g., inside walls) currently denied to non-invasive methods.
How robust do we want or need the built environment to be against the sequelae of rare but intense bursts of moisture? Occurrences of intense rainfall and flooding are expected to become more frequent consequent to climate change [24]. Pipes leak and burst unexpectedly. What can be done against rare but highly consequential events? Plumbing biofilms might be seeded with microbes anticipated to benignly bias microbial sequelae. Preparations of normally dry regions might include pre-treatment inoculation of buildings (including the interior of walls) with benign microbes in a quiescent state but able to germinate and take over a niche that when wetted would otherwise be vulnerable to the growth of noxious microbes. These are hopeful speculations but they suggest research to identify desiccation-tolerant and quickly revivable benign microbes. Microbial viability through multiple cycles of wetting and desiccation appears to be a marvelous topic at the fundamental level [25, 26] whose deeper understanding could also lead to practical consequences [27]. The interactions of microbes, moisture cycles, and the moisture retention properties of building materials could become a rich area for interdisciplinary study.
Sentinel microbes that are themselves innocuous but chosen or engineered to be easy to measure could be deliberately inoculated as monitors for microbial growth allowing quantification of accumulated stable isozymes of indicators such as β-galactosidase or GFP. In a similar way, sentinel microbes could be added to foods as a way to quantify if and how much the food experienced conditions allowing microbial growth. Extensions of NGS have the potential to identify metabolically active microbes (Table 3 legend). Sentinel microbes could be developed as another window to understanding the places and conditions in buildings that promote or permit microbial metabolism.
Hygiene and biodiversity hypotheses
The hygiene [28] and the related (microbial) biodiversity hypotheses [29] propose that diverse microbial exposure is key to the optimum development and function of the immune system (Table 2). A diversity of immune epitopes in the microbial environment is proposed to channel the immune system such that autoimmune reactivity becomes less likely. Despite—indeed because of—the attractiveness of these ideas, they should be critically examined and alternatives considered. We lack knowledge of which logical and operational definitions of biodiversity [30, 31] are most relevant to human health. Over 50 years ago, Dubos et al. demonstrated that a benign intestinal microflora protects against microbial pathogens [32]. However, proving a role for microbes is not the same as proving that microbial complexity or diversity are required—or even helpful—to do the job.
An astonishing finding contradicts a key predication of the biodiversity hypotheses regarding the role of a complex microbiome in normal intestinal development: Infection by a single norovirus strain corrects all defects associated with axenic intestinal development in the mouse [33]! There remains a great deal of suggestive and intriguing correlative but not definitive evidence in favor of the hygiene and biodiversity hypotheses [10, 34–41]. The propensity to develop asthma correlated with a low diversity of fungi in dust samples [42] and the intestinal microbiome may play a role immune conditioning regarding susceptibility to asthma [43, 44]. Gammaproteobacterial complexity on the forearm is associated with benign immune tolerance as shown by a decrease in atopy [45]. Hanski et al. favor a causal relationship in which the microbiota leads to immune tolerance while acknowledging that they cannot rule out the causally inverse interpretation that an intolerant immune reaction alters the microbiota. Subsequent studies show an immune-moderating role associated with endotoxin of one of the identified bacteria: Acinetobacter lwoffi in both human cells and a mouse sensitization protocol [46] but immune tolerance is a double-edged sword. Other experimental and clinical contexts show bacterial endotoxin-triggering immune responses that are in some experimental systems protective against infection but in others harmful to the organism [47]. Immunological tolerance can predispose to disease—susceptibility to infectious disease—as well as to health as in the diminishment of atopy. Tolerance toward environmental mycobacteria built-up as a consequence of birth and early life in environments rich in this biota may be part of the reason it is hard to make an effective vaccine against Mycobacterium tuberculosis and induced tolerance during infection may also be a part of the M. tuberculosis strategy for pathogenesis [48].
Is sequence complexity the medium or the message?
The hypothesis that microbiome diversity measured by NGS of genes encoding small subunit RNA (ssuRNA) is a positive driver of a building’s or an individual’s microbiome health [49] deserves consideration but, based on current evidence, not acceptance. Despite the cleverness and attractiveness of the idea, ecosystem and evolutionary complexity do not scale with measures of stability, “useful”, or “adaptive” [50, 51]. “Just so stories” in the popular press or TED talks may imply that differences in sequence distribution reflect microbial adaptation or benign “appropriateness” to a niche but correlation is not causation. Microbial distribution in what might have been thought to be reasonably uniformly mixed oceans appears largely consequent to the seeding of microorganisms that differ only in neutral mutations [52, 53]. The situation may be even more arbitrary and coincidental in the context of buildings. Sequences in a sample of dry dust from a building may sometimes (often?) represent nothing more than vagaries of air and human traffic circulation rather than microbial adaptation to the niche in which they are found. And yet, the health consequences of microbial epitope-human immune system interactions may be profound even if chance brought them together. Complexity in sequencing data as well as in the potential interactions of microbes with an already complicated indoor chemistry [54] are facts but complexity in interpretation can also be consequent to a deficit in theory [55]. Random sampling alone might explain why “complex” and “rich in diversity” microbiomes are more likely to contain rare but specific actinic compounds. In science, one should stay alert to the possibility that apparent complexity represents an intermediate state. Simplicity at the core of complexity remains a reasonable null hypothesis. On the other hand, the current state of knowledge cannot rule out complexity itself as a possible answer. Molecular mechanism(s) by which microbiome complexity itself might promote host health have been alluded to in an almost romantic fashion that is inspiring but not well articulated in forms that are testable and falsifiable experimentally. A meta-analysis finds that children raised on farms have approximately 25 % lower asthma prevalence [56] and one explanation is an ameliorative effect of exposure to a diverse microbiota [37]. Attempts are made to rule out confounding factors, but alternatives to the microbial biodiversity hypothesis include genetic predispositions, exercise, altered diet, environmental pollution either outdoors or indoors, changes in sleep patterns, and vitamin D insufficiency [57].
The food epitope hypothesis
Diet has been discussed as a way to alter immune function by several mechanisms including modification of the host commensal microbiome [58–61] but proposed here is an additional mechanism by which epitope diversity in the food itself influences the immune system: The Neolithic revolution brought about a decrease in the variety of foods consumed by our species [62]. Breeding practices in agriculture have changed over the last 50 years and it is a reasonable proposition—but remains to be proven—that consequently the allelic diversity of major foodstuffs destined for human consumption has been further reduced. Genetic uniformity of crops in contemporary agriculture is discussed in the context of food security and the susceptibility of plant and animal agriculture to infectious disease [63, 64]. The hypothesis proposed here is that there are also immunological, developmental, and neuro-psychiatric [65, 66] consequences of a punctate distribution of dietary food-derived near epitopes (Fig. 1 and for a definition of “near” epitopes, see the last paragraph of Table 2). The hypotheses for health benefits of exposure to a rich microbial diversity appear also applicable to the allelic diversity of food. The two views are not qualitatively at odds since immune function can be jointly modulated by diet and microbial exposure [67]. Quantitatively, daily food consumption with its associated epitopes exceeds exposure to microbial epitopes by orders of magnitude. Thus, we return at the end of this review and commentary to the point on which we began. The first Neolithic revolution has been our species’ most important innovation and has also led to large modifications of earth’s biosphere. Deepening our understanding of the first Neolithic revolutions consequences—including unintended consequences—will inform opportunities for further benign developments. Our species may not have another 10,000 years to get the second Neolithic revolution- the microbial Neolithic revolution- right.

The food epitope hypothesis. Epitope space [68] of food consumed by humans before (a), after (b) the Neolithic revolution, and the modern phase of agricultural genetics (c). All parameters of shape space are compressed into the X-axis. The Y-axis is a frequency distribution. a In the pre-Neolithic phase of our species, we were hunter-gatherers. The food that we ate was maximally (for our species) dispersed in the living world and we were exposed to more dietary epitopes. The variation in epitopes of each food source was based on the allelic variance within plant and animal populations [69]. Each food source was population based containing the genetic and allelic diversity that implies [70]. b The Neolithic revolution, i.e., the advent of agriculture and domestication of both plant and animal food sources. The epitope classes and their dietary distribution became relatively restricted. Most previous food sources were no longer consumed but a few made up the majority of the diet. Within these food sources, the amount of variation was also constrained because even in early agriculture, artificial selection limits the allelic and by implication the epitope diversity in food [71]. c The modern agricultural era of the last ca. 50 years has led to human food becoming more genetically uniform within each species of plants and livestock [72]. The associated distribution of food epitopes in shape space has become punctate
Conclusions
The microbial Neolithic revolution is underway with regard to the gut microbiome but its extension to buildings requires the clarification of key matters: (1) Active microbial metabolism in buildings is known to be associated and causative of SBS/BRS. Focused research will be required to learn if active microbial metabolism is in some cases, or could ever become, healthful in buildings. (2) The association of some measures of microbial diversity in buildings with some measures of human occupant health is intriguing but the responsible mechanism(s) remain unknown. Diversity may simply represent a bigger sampling more likely to contain a few (mostly unknown) key compounds or, alternatively, exposure to diversity itself may be healthful, again through mechanism(s) requiring clarification through focused research. (3) More research is needed to critically compare, contrast, hierarchically organize, quantify, and understand the health-relevant consequences of human exposure to diversity from multiple sources including microbes as well as the products of the first Neolithic revolution, food.
Abbreviations
- BRS:
-
building-related symptoms
- BRI:
-
building-related illness
- GFP:
-
green fluorescent protein
- HVAC:
-
heating ventilation and air-conditioning
- NGS:
-
next-generation sequencing
- SBS:
-
sick building syndrome
- ssuRNA:
-
small subunit RNA (inclusive of 16sRNA in prokaryotes and 18sRNA in eukaryotic cytoplasm)
References
Levin H, Taubel M, Hernandez M. Summary of Sloan symposium: healthy buildings 2015-Europe. Microbiome. 2015;3(1):68. doi:10.1186/s40168-015-0115-4.
Jordan N, Boody G, Broussard W, Glover JD, Keeney D, McCown BH, et al. Environment. Sustainable development of the agricultural bio-economy. Science. 2007;316(5831):1570–1.
Levin H. Building materials and indoor air quality. Occupational Medicine. 1989. p. 4.
Hrudey S, Charrois J, editors. Disinfection by-products: relevance to Human Health. London: IWA; 2012.
Kurenbach B, Marjoshi D, Amabile-Cuevas CF, Ferguson GC, Godsoe W, Gibson P, et al. Sublethal exposure to commercial formulations of the herbicides dicamba, 2,4-dichlorophenoxyacetic acid, and glyphosate cause changes in antibiotic susceptibility in Escherichia coli and Salmonella enterica serovar Typhimurium. mBio. 2015;6:2. doi:10.1128/mBio.00009-15.
Beale B. Probiotics: their tiny worlds are under scrutiny. The Scientist. 2002;16(15):1.
Mackowiak PA. Recycling Metchnikoff: probiotics, the intestinal microbiome and the quest for long life. Front Public Health. 2013;1:52. doi:10.3389/fpubh.2013.00052.
Levin H, Corsi R. Conceptual framework for building science in indoor microbiome studies. Indoor air Hong Kong. 2014.
Rubin BE, Sanders JG, Hampton-Marcell J, Owens SM, Gilbert JA, Moreau CS. DNA extraction protocols cause differences in 16S rRNA amplicon sequencing efficiency but not in community profile composition or structure. Microbiologyopen. 2014;3(6):910–21. doi:10.1002/mbo3.216.
Ege MJ, Mayer M, Normand AC, Genuneit J, Cookson WO, Braun-Fahrlander C, et al. Exposure to environmental microorganisms and childhood asthma. N Engl J Med. 2011;364(8):701–9. doi:10.1056/NEJMoa1007302.
Nunez M, Hammer H. Microbial specialists in below-grade foundation walls in Scandinavia. Indoor Air. 2014;24(5):543–51. doi:10.1111/ina.12095.
Rintala H. Actinobacteria in indoor environments: exposures and respiratory health effects. Front Biosci (Schol Ed). 2011;3:1273–84.
Sanglier JJ, Haag H, Huck TA, Fehr T. Novel bioactive compounds from Actinomycetes: a short review (1988–1992). Res Microbiol. 1993;144(8):633–42.
Penttinen P, Pelkonen J, Huttunen K, Hirvonen MR. Co-cultivation of Streptomyces californicus and Stachybotrys chartarum stimulates the production of cytostatic compound(s) with immunotoxic properties. Toxicol Appl Pharmacol. 2006;217(3):342–51. doi:10.1016/j.taap.2006.09.010.
Huttunen K, Pelkonen J, Nielsen KF, Nuutinen U, Jussila J, Hirvonen MR. Synergistic interaction in simultaneous exposure to Streptomyces californicus and Stachybotrys chartarum. Environ Health Perspect. 2004;112(6):659–65.
Feazel LM, Baumgartner LK, Peterson KL, Frank DN, Harris JK, Pace NR. Opportunistic pathogens enriched in showerhead biofilms. Proc Natl Acad Sci USA. 2009;106(38):16393–9.
Falkinham 3rd JO. Ecology of nontuberculous mycobacteria—where do human infections come from? Semin Respir Crit Care Med. 2013;34(1):95–102. doi:10.1055/s-0033-1333568.
Cao B, Liu X, Yu X, Chen M, Feng L, Wang L. A new oligonucleotide microarray for detection of pathogenic and non-pathogenic Legionella spp. PLoS One. 2014;9(12):e113863. doi:10.1371/journal.pone.0113863.
O’Doherty KC, Virani A, Wilcox ES. The human microbiome and public health: social and ethical considerations. Am J Public Health. 2016:e1-e7. doi:10.2105/AJPH.2015.302989.
Hoehler TM, Jorgensen BB. Microbial life under extreme energy limitation. Nat Rev Microbiol. 2013;11(2):83–94. doi:10.1038/nrmicro2939.
Sauni R, Verbeek JH, Uitti J, Jauhiainen M, Kreiss K, Sigsgaard T. Remediating buildings damaged by dampness and mould for preventing or reducing respiratory tract symptoms, infections and asthma. Cochrane Database Syst Rev. 2015;2:CD007897. doi:10.1002/14651858.CD007897.pub3.
Pelletier MG, Karthikeyan S, Green TR, Schwartz RC, Wanjura JD, Holt GA. Soil moisture sensing via swept frequency based microwave sensors. Sensors (Basel). 2012;12(1):753–67. doi:10.3390/s120100753.
Adib F, Kabelac Z, Katabi D. Multi-person localization via RF body reflections. Oakland, CA, USA: 12th USENIX Symposium on Networked Systems Design and Implementation (NSDI); 2015. p. 15.
Taylor J, Biddulph P, Davies M, Lai K. Predicting the microbial exposure risks in urban floods using GIS, building simulation, and microbial models. Environ Int. 2013;51:182–95. doi:10.1016/j.envint.2012.10.006.
Krisko A, Radman M. Biology of extreme radiation resistance: the way of Deinococcus radiodurans. Cold Spring Harb Perspect Biol. 2013;5:7. doi:10.1101/cshperspect.a012765.
Devigne A, Ithurbide S, Bouthier de la Tour C, Passot F, Mathieu M, Sommer S, et al. DdrO is an essential protein that regulates the radiation desiccation response and the apoptotic-like cell death in the radioresistant Deinococcus radiodurans bacterium. Mol Microbiol. 2015;96(5):1069–84. doi:10.1111/mmi.12991.
Taylor J, Davies M, Canales M, Lai KM. The persistence of flood-borne pathogens on building surfaces under drying conditions. Int J Hyg Environ Health. 2013;216(1):91–9. doi:10.1016/j.ijheh.2012.03.010.
D. P. S. Hay fever, hygiene and household size. BMJ. 1989;299:1259–60.
von Hertzen L, Beutler B, Bienenstock J, Blaser M, Cani PD, Eriksson J, et al. Helsinki alert of biodiversity and health. Ann Med. 2015;47(3):218–25. doi:10.3109/07853890.2015.1010226.
Tuomisto H. Defining, measuring, and partitioning species diversity. Encyclopedia of biodiversity. secondth ed. Waltham MA: Academic; 2013. p. 434–46.
Tuomisto H. A consistent terminology for quantifying species diversity? Yes, it does exist. Oecologia. 2010;164(4):853–60. doi:10.1007/s00442-010-1812-0.
Moberg CL. Rene Dubos: a harbinger of microbial resistance to antibiotics. Microb Drug Resist. 1996;2:287–97.
Kernbauer E, Ding Y, Cadwell K. An enteric virus can replace the beneficial function of commensal bacteria. Nature. 2014;516(7529):94–8. doi:10.1038/nature13960.
Braun-Fahrlander C, Riedler J, Herz U, Eder W, Waser M, Grize L, et al. Environmental exposure to endotoxin and its relation to asthma in school-age children. N Engl J Med. 2002;347(12):869–77. doi:10.1056/NEJMoa020057.
Wlasiuk G, Vercelli D. The farm effect, or: when, what and how a farming environment protects from asthma and allergic disease. Curr Opin Allergy Clin Immunol. 2012;12(5):461–6. doi:10.1097/ACI.0b013e328357a3bc.
von Mutius E, Vercelli D. Farm living: effects on childhood asthma and allergy. Nat Rev Immunol. 2010;10(12):861–8. doi:10.1038/nri2871.
Valkonen M, Wouters IM, Taubel M, Rintala H, Lenters V, Vasara R, et al. Bacterial exposures and associations with atopy and asthma in children. PLoS One. 2015;10(6):e0131594. doi:10.1371/journal.pone.0131594.
Heederik D, von Mutius E. Does diversity of environmental microbial exposure matter for the occurrence of allergy and asthma? J Allergy Clin Immunol. 2012;130(1):44–50. doi:10.1016/j.jaci.2012.01.067.
Weber J, Illi S, Nowak D, Schierl R, Holst O, von Mutius E, et al. Asthma and the hygiene hypothesis. Does cleanliness matter? Am J Respir Crit Care Med. 2015;191(5):522–9. doi:10.1164/rccm.201410-1899OC.
Mustonen K, Karvonen AM, Kirjavainen P, Roponen M, Schaub B, Hyvarinen A, et al. Moisture damage in home associates with systemic inflammation in children. Indoor Air. 2015. doi:10.1111/ina.12216.
Kirjavainen PV, Taubel M, Karvonen AM, Sulyok M, Tiittanen P, Krska R, et al. Microbial secondary metabolites in homes in association with moisture damage and asthma. Indoor Air. 2015. doi:10.1111/ina.12213.
Dannemiller KC, Mendell MJ, Macher JM, Kumagai K, Bradman A, Holland N, et al. Next-generation DNA sequencing reveals that low fungal diversity in house dust is associated with childhood asthma development. Indoor Air. 2014;24(3):236–47.
Earl CS, An SQ, Ryan RP. The changing face of asthma and its relation with microbes. Trends Microbiol. 2015;23(7):408–18. doi:10.1016/j.tim.2015.03.005.
Arrieta MC, Stiemsma LT, Dimitriu PA, Thorson L, Russell S, Yurist-Doutsch S, et al. Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci Transl Med. 2015;7(307):307ra152. doi:10.1126/scitranslmed.aab2271.
Hanski I, von Hertzen L, Fyhrquist N, Koskinen K, Torppa K, Laatikainen T, et al. Environmental biodiversity, human microbiota, and allergy are interrelated. Proc Natl Acad Sci USA. 2012;109(21):8334–9. doi:10.1073/pnas.1205624109.
Fyhrquist N, Ruokolainen L, Suomalainen A, Lehtimaki S, Veckman V, Vendelin J, et al. Acinetobacter species in the skin microbiota protect against allergic sensitization and inflammation. J Allergy Clin Immunol. 2014;134(6):1301–9. doi:10.1016/j.jaci.2014.07.059. e11.
Takeuchi O, Akira S. Pattern recognition receptors and inflammation. Cell. 2010;140(6):805–20. doi:10.1016/j.cell.2010.01.022.
Kim K, Perera R, Tan DB, Fernandez S, Seddiki N, Waring J, et al. Circulating mycobacterial-reactive CD4+ T cells with an immunosuppressive phenotype are higher in active tuberculosis than latent tuberculosis infection. Tuberculosis (Edinburgh, Scotland). 2014;94(5):494–501. doi:10.1016/j.tube.2014.07.002.
Lax S, Nagler CR, Gilbert JA. Our interface with the built environment: immunity and the indoor microbiota. Trends Immunol. 2015;36(3):121–3. doi:10.1016/j.it.2015.01.001.
Thompson J. Relentless evolution. University of Chicago Press. 2013.
Gould S. The structure of evolutionary theory. Belknap Press. 2002.
Giovannoni S, Nemergut D. Ecology. Microbes ride the current. Science. 2014;345(6202):1246–7. doi:10.1126/science.1259467.
Hellweger FL, van Sebille E, Fredrick ND. Biogeographic patterns in ocean microbes emerge in a neutral agent-based model. Science. 2014;345(6202):1346–9. doi:10.1126/science.1254421.
Weschler C. Changes in indoor pollutants since the 1950’s. Atmos Environ. 2009;43:153–69.
Brenner S. In theory. Curr Biol. 1997;7(3):202.
Genuneit J. Exposure to farming environments in childhood and asthma and wheeze in rural populations: a systematic review with meta-analysis. Pediatr Allergy Immunol. 2012;23(6):509–18. doi:10.1111/j.1399-3038.2012.01312.x.
Honeyman M, Harrison L. Alternative and additional mechanisms to the hygiene hypothesis. In: Rook GAW, editor. The Hygiene Hypothesis and Darwinian Medicine. Basel: Birkhäuser; 2009. p. 279.
Ponton F, Wilson K, Holmes AJ, Cotter SC, Raubenheimer D, Simpson SJ. Integrating nutrition and immunology: a new frontier. J Insect Physiol. 2013;59(2):130–7. doi:10.1016/j.jinsphys.2012.10.011.
Ponton F, Wilson K, Cotter SC, Raubenheimer D, Simpson SJ. Nutritional immunology: a multi-dimensional approach. PLoS Pathog. 2011;7(12):e1002223. doi:10.1371/journal.ppat.1002223.
Thorburn AN, Macia L, Mackay CR. Diet, metabolites, and “western-lifestyle” inflammatory diseases. Immunity. 2014;40(6):833–42. doi:10.1016/j.immuni.2014.05.014.
Rook GA, Lowry CA, Raison CL. Hygiene and other early childhood influences on the subsequent function of the immune system. Brain Res. 2015;1617:47–62. doi:10.1016/j.brainres.2014.04.004.
Armelagos GJ. Brain evolution, the determinates of food choice, and the omnivore’s dilemma. Crit Rev Food Sci Nutr. 2014;54(10):1330–41. doi:10.1080/10408398.2011.635817.
Lederberg JJBS. Haldane (1949) on infectious disease and evolution. Genetics. 1999;153(1):1–3.
Fu YB. Understanding crop genetic diversity under modern plant breeding. Theor Appl Genet. 2015;128(11):2131–42. doi:10.1007/s00122-015-2585-y.
Zuliani L, Graus F, Giometto B, Bien C, Vincent A. Central nervous system neuronal surface antibody associated syndromes: review and guidelines for recognition. J Neurol Neurosurg Psychiatry. 2012;83(6):638–45. doi:10.1136/jnnp-2011-301237.
Ransohoff RM, Schafer D, Vincent A, Blachere NE, Bar-Or A. Neuroinflammation: ways in which the immune system affects the brain. Neurotherapeutics. 2015. doi:10.1007/s13311-015-0385-3.
Brown EM, Wlodarska M, Willing BP, Vonaesch P, Han J, Reynolds LA, et al. Diet and specific microbial exposure trigger features of environmental enteropathy in a novel murine model. Nat Commun. 2015;6:7806. doi:10.1038/ncomms8806.
Povolotskaya IS, Kondrashov FA. Sequence space and the ongoing expansion of the protein universe. Nature. 2010;465(7300):922–6. doi:10.1038/nature09105.
Stoeckle MY, Thaler DS. DNA barcoding works in practice but not in (neutral) theory. PLoS One. 2014;9(7):e100755. doi:10.1371/journal.pone.0100755.
Fukunaga K, Hill J, Vigouroux Y, Matsuoka Y, Sanchez GJ, Liu K, et al. Genetic diversity and population structure of teosinte. Genetics. 2005;169(4):2241–54. doi:10.1534/genetics.104.031393.
Jaenicke-Despres V, Buckler ES, Smith BD, Gilbert MT, Cooper A, Doebley J, et al. Early allelic selection in maize as revealed by ancient DNA. Science. 2003;302(5648):1206–8. doi:10.1126/science.1089056.
Khoury CK, Bjorkman AD, Dempewolf H, Ramirez-Villegas J, Guarino L, Jarvis A, et al. Increasing homogeneity in global food supplies and the implications for food security. Proc Natl Acad Sci USA. 2014;111(11):4001–6. doi:10.1073/pnas.1313490111.
Shin H, Pei Z, Martinez 2nd KA, Rivera-Vinas JI, Mendez K, Cavallin H, et al. The first microbial environment of infants born by C-section: the operating room microbes. Microbiome. 2015;3(1):59. doi:10.1186/s40168-015-0126-1.
Marineli F, Tsoucalas G, Karamanou M, Androutsos G. Mary Mallon (1869–1938) and the history of typhoid fever. Ann Gastroenterol. 2013;26(2):132–4.
von Hippel PH. Remark while lecturing on DNA-protein interactions. 1983.
Pauling L, Delbruck M. The nature of the intermolecular forces operative in biological processes. Science. 1940;92(2378):77–9. doi:10.1126/science.92.2378.77.
Botstein D. Decoding the language of genetics. Cold Spring Harbor Laboratory Press. 2015.
Dylan B. Talkin New York Blues. Bob Dylan (first album). New York: Columbia Records; 1962.
Messmer BT, Sullivan JJ, Chiorazzi N, Rodman TC, Thaler DS. Two human neonatal IgM antibodies encoded by different variable-region genes bind the same linear peptide: evidence for a stereotyped repertoire of epitope recognition. J Immunol. 1999;162(4):2184–92.
Xu GJ, Kula T, Xu Q, Li MZ, Vernon SD, Ndung'u T, et al. Viral immunology. Comprehensive serological profiling of human populations using a synthetic human virome. Science. 2015;348(6239):aaa0698. doi:10.1126/science.aaa0698.
Attaf M, Huseby E, Sewell AK. alphabeta T cell receptors as predictors of health and disease. Cell Mol Immunol. 2015;12(4):391–9. doi:10.1038/cmi.2014.134.
Skums P, Bunimovich L, Khudyakov Y. Antigenic cooperation among intrahost HCV variants organized into a complex network of cross-immunoreactivity. Proc Natl Acad Sci U S A. 2015;112(21):6653–8. doi:10.1073/pnas.1422942112.
Toivola M, Alm S, Reponen T, Kolari S, Nevalainen A. Personal exposures and microenvironmental concentrations of particles and bioaerosols. J Environ Monit. 2002;4(1):166–74.
Wilton M, Charron-Mazenod L, Moore R, Lewenza S. Extracellular DNA acidifies biofilms and induces aminoglycoside resistance in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2015. doi:10.1128/AAC.01650-15.
Dlott G, Maul JE, Buyer J, Yarwood S. Microbial rRNA:rDNA gene ratios may be unexpectedly low due to extracellular DNA preservation in soils. J Microbiol Methods. 2015;115:112–20. doi:10.1016/j.mimet.2015.05.027.
Carell T. DNA repair. Angewandte Chemie (International ed). 2015. doi:10.1002/anie.201509770.
Cheng KC, Cahill DS, Kasai H, Nishimura S, Loeb LA. 8-Hydroxyguanine, an abundant form of oxidative DNA damage, causes G----T and A----C substitutions. The Journal of biological chemistry. 1992;267(1):166–72.
Bhagwat AS, Hao W, Townes JP, Lee H, Tang H, Foster PL. Strand-biased cytosine deamination at the replication fork causes cytosine to thymine mutations in Escherichia coli. Proc Natl Acad Sci U S A. 2016. doi:10.1073/pnas.1522325113.
Paabo S, Irwin DM, Wilson AC. DNA damage promotes jumping between templates during enzymatic amplification. J Biol Chem. 1990;265:4718–21.
Liu S, Thaler DS, Libchaber A. Signal and noise in bridging PCR. BMC Biotechnol. 2002;2(1):13.
Shade A, Gilbert JA. Temporal patterns of rarity provide a more complete view of microbial diversity. Trends Microbiol. 2015;23(6):335–40. doi:10.1016/j.tim.2015.01.007.
Sogin ML, Morrison HG, Huber JA, Welch DM, Huse SM, Neal PR, et al. Microbial diversity in the deep sea and the underexplored “rare biosphere”. Proc Natl Acad Sci U S A. 2006;103(32):12115–20.
Ley RE, Harris JK, Wilcox J, Spear JR, Miller SR, Bebout BM, et al. Unexpected diversity and complexity of the Guerrero Negro hypersaline microbial mat. Appl Environ Microbiol. 2006;72(5):3685–95.
Konstantinidis KT, Rossello-Mora R. Classifying the uncultivated microbial majority: a place for metagenomic data in the Candidatus proposal. Syst Appl Microbiol. 2015;38(4):223–30. doi:10.1016/j.syapm.2015.01.001.
Villarreal JV, Jungfer C, Obst U, Schwartz T. DNase I and proteinase K eliminate DNA from injured or dead bacteria but not from living bacteria in microbial reference systems and natural drinking water biofilms for subsequent molecular biology analyses. J Microbiol Methods. 2013;94(3):161–9. doi:10.1016/j.mimet.2013.06.009.
Okshevsky M, Regina VR, Meyer RL. Extracellular DNA as a target for biofilm control. Curr Opin Biotechnol. 2015;33:73–80. doi:10.1016/j.copbio.2014.12.002.
Paul BJ, Ross W, Gaal T, Gourse RL. rRNA transcription in Escherichia coli. Annu Rev Genet. 2004;38:749–70.
Kiviet DJ, Nghe P, Walker N, Boulineau S, Sunderlikova V, Tans SJ. Stochasticity of metabolism and growth at the single-cell level. Nature. 2014;514(7522):376–9. doi:10.1038/nature13582.
Flardh K, Cohen PS, Kjelleberg S. Ribosomes exist in large excess over the apparent demand for protein synthesis during carbon starvation in marine Vibrio sp. strain CCUG 15956. J Bacteriol. 1992;174(21):6780–8.
Vogel SJ, Tank M, Goodyear N. Variation in detection limits between bacterial growth phases and precision of an ATP bioluminescence system. Lett Appl Microbiol. 2014;58(4):370–5. doi:10.1111/lam.12199.
Shama G, Malik DJ. The uses and abuses of rapid bioluminescence-based ATP assays. Int J Hyg Environ Health. 2013;216(2):115–25. doi:10.1016/j.ijheh.2012.03.009.
Slade D, Radman M. Oxidative stress resistance in Deinococcus radiodurans. Microbiol Mol Biol Rev. 2011;75(1):133–91. doi:10.1128/MMBR.00015-10.
Hufnagel DA, Depas WH, Chapman MR. The biology of the Escherichia coli extracellular matrix. Microbiol Spectr. 2015;3:3. doi:10.1128/microbiolspec.MB-0014-2014.
Colwell RR. Viable but nonculturable bacteria: a survival strategy. J Infect Chemother. 2000;6(2):121–5. doi:10.1007/s101560000026.
Pietramellara G, Ascher J, Borgogn F, Ceccherini M, Guerri G, Nannipieri P. Extracellular DNA in soil and sediment: fate and ecological relevance. Biol Fertil Soils. 2008;45(3):219–35.
Redfield RJ. Evolution of bacterial transformation: is sex with dead cells ever better than no sex at all? Genetics. 1988;119(1):213–21.
Acknowledgements
Special thanks to Hal Levin and Martin Täubel for stimulating discussions, Maria Nunez, Anne Hyvärinen, Jordan Peccia, Martin Täubel, and Jack Gilbert for insight into building microbes, Bruce Levin, Rustom Antia, and George Armelagos for discussion and encouragement of the food epitope hypothesis, Marc Lipsitch for suggesting sentinel microbes, Jeffrey Siegel for inventing the phrase “building tomography,” Fiona Doetsch for formalizing Fig. 1, anonymous reviews for critique leading to major revisions (The précis is taken essentially verbatim from anonymous review), and the Sloan Foundation for support.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The author declares that he has no competing interests.
Authors’ contributions
The sole author is responsible for all aspects of this paper.
Ethics approval and consent to participate
This is a review and hypothesis paper. No additional consent for review of published work involving humans was required and none was sought. To the best of the author’s knowledge, all work reviewed herein was carried out with appropriate approvals and in compliance with the Helsinki Declaration.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Thaler, D.S. Toward a microbial Neolithic revolution in buildings. Microbiome 4, 14 (2016). https://doi.org/10.1186/s40168-016-0157-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40168-016-0157-2