
Abstract
Neoantigens derived from non-synonymous somatic mutations are restricted to malignant cells and are thus considered ideal targets for T cell receptor (TCR)-based immunotherapy. Adoptive transfer of T cells bearing neoantigen-specific TCRs exhibits the ability to preferentially target tumor cells while remaining harmless to normal cells. High-avidity TCRs specific for neoantigens expressed on AML cells have been identified in vitro and verified using xenograft mouse models. Preclinical studies of these neoantigen-specific TCR-T cells are underway and offer great promise as safe and effective therapies. Additionally, TCR-based immunotherapies targeting tumor-associated antigens are used in early-phase clinical trials for the treatment of AML and show encouraging anti-leukemic effects. These clinical experiences support the application of TCR-T cells that are specifically designed to recognize neoantigens. In this review, we will provide a detailed profile of verified neoantigens in AML, describe the strategies to identify neoantigen-specific TCRs, and discuss the potential of neoantigen-specific T-cell-based immunotherapy in AML.
Similar content being viewed by others

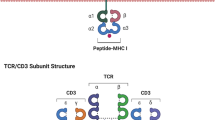
Introduction
Acute myeloid leukemia (AML) is the most common indication for hematopoietic stem cell transplantation (HSCT) [1, 2]. The curative potential of the T cell-mediated graft-versus-leukemia (GVL) effects after HSCT indicates that T cell-based immunotherapy may be a powerful strategy against AML cells [3]. The adoptive transfer of T cells, notably chimeric antigen receptor (CAR)- and T cell receptor (TCR)-engineered T cells, has demonstrated an amazing anti-leukemic potential in mice models and human research [4,5,6,7,8]. TCR-T cells kill leukemic cells through the interaction between their modified TCRs and antigens presented by human leukocyte antigen (HLA) molecules on the surfaces of leukemic cells. TCR-T cells targeting tumor-associated antigens (TAAs), such as WT1, PRAME, and HA-1, exert anti-leukemic effects in vitro and in clinical studies with high-risk AML patients post-HSCT [9,10,11,12]. However, owing to low antigen expression in non-malignant cells, antitumor immunity targeting TAAs, also known as “self-antigens”, might be accompanied by serious toxicities [13, 14]. Therefore, a major obstacle to TCR-T cell-based immunotherapy is the selection of appropriate targets. Unlike TAAs, neoantigens arise from somatic mutation and are restricted to malignant cells [15]. Immune escape by neoantigens is less plausible since tumor cells need to express these driver genes to retain their malignant phenotypes [16]. Neoantigens can be recognized as foreign antigens by autologous T cells and thus constitute ideal targets for immunotherapy [17]. T cells specific for neoantigens have been detected in AML patients and in donor T cells after HSCT [18, 19]. Furthermore, specific T-cell activities can also be induced in vitro by epitopes derived from neoantigens using peripheral blood mononuclear cells (PBMCs) from AML patients [20]. These results support the presumption that neoantigen-specific T cell-based immunotherapy represents an effective approach to treating AML patients without harming normal tissues. In this review, we provide an overview of known neoantigens in AML, describe TCR-based immunotherapies targeting certain neoantigens, and discuss the current efforts to translate these findings into the clinic.
Neoantigens in AML
Recent developments in sequencing technology have revealed that a stepwise accumulation of genetic abnormalities is a hallmark of cancer. Gene fusions, small insertions and deletions (indels), non-synonymous point mutations, and cancer-specific splicing variants can result in the production of neoantigens (Table 1) [16]. One high-quality neoantigen may be sufficient for disease control or perhaps a cure in the majority of instances, if it is derived from the mutation that plays a critical role in the malignant phenotype [21, 22]. AML individuals commonly harbor recurrent gene alterations, which may lead to shared neoantigens that trigger potent anti-leukemic responses [23, 24].
In general, the identification of neoantigens comprises the following procedures. The first step is usually to compile a list of genomic alterations (including somatic mutations, indels, and gene fusions) from next-generation sequencing data, including whole-genome sequencing, whole-exon sequencing, RNA-sequencing, and DNA panel data, and convert them into “neopeptides” with appropriate lengths. The second step is to predict the binding affinity between neopeptides and patient-specific HLA alleles. Many tools are available for this prediction, such as BIMAS [25] for HLA class I prediction, TEPITOPE [26], PickPocket [27], MULTIPRED2 [28], MultiRTA [29] for HLA class II prediction, and RANKPEP [30], SYFPEITHI [31], NetMHCpan [32,33,34,35], and IEDB [36] for both HLA class I and II prediction. The third step is the evaluation of the immunogenicity of the predicted neopeptides, which usually includes testing the ability of neoepitope to induce specific T cells, and validate the functions of specific T cells against malignant cells through cytokine-releasing methods and cytotoxic assays. In addition, the rapid progress in high-throughput mass-spectrometry approaches allows for direct identification of HLA-binding neopeptides [37]. Furthermore, to better apply neoantigen-based therapies for the treatment of cancer patients, potential immunogenic neopeptides should be rapidly and accurately screened. Numerous pipelines, including pTuneos [38], ScanNeo [39], INTEGRATE-Neo [40], CloudNeo [41], TIminer [42], TSNAD [43, 44], MuPeXI [45], pVAC-Seq [46], Neopepsee [47], and many others, have been established to implement mutation calling of multifarious sequencing data and affinity prediction of MHC molecular. Various neoantigens have been confirmed through the flexible use of the aforementioned methodologies.
Here we summarize hotspot neoantigens in AML.
Gene fusions
Gene fusions caused by chromosomal translocations play a significant role in AML, and they are implicated in approximately half of AML patients. Most gene fusions contribute to oncogenicity by altering hematopoietic-specific transcription factors, thus blocking myeloid differentiation at a specific stage. Due to the recurrent nature of fusion genes in AML, neoepitopes derived from these fusion genes may serve as efficient neoantigens for immunotherapy in AML patients.
CBFB-MHY11
The core binding factor (CBF) protein complex is the main target of translocations in AML [48]. The inv(16)(p13q22) and the less frequent t(16;16)(p13q22) result in the formation of a CBFB-MYH11 fusion gene that encodes the fusion protein CBFβ-SMMHC [49]. The resultant fusion protein disrupts CBF functions and leads to a block of myeloid differentiation and ultimately to leukemia [50,51,52]. A nonameric neopeptide (REEMEVHEL) derived from the CBFβ-SMMHC fusion protein has shown considerable immunogenicity in HLA-B*40:01 donors. CD8+ T cell clones induced by this neopeptide kill AML cell lines and primary human AML cells in vitro and in an in vivo patient-derived murine xenograft (PDX) model, in a CBFβ-SMMHC-specific and HLA-A*40:01-restricted manner [53].
PML-RARA
The PML-RARA fusion gene results from a chromosomal translocation t(15; 17)(q24.1;q12.1), which fuses the PML gene on chromosome 15 to the RARA gene on chromosome 17 [54]. The encoded PML-RARA protein serves as a driving event of acute promyelocytic leukemia (APL) [55]. The neoepitope BCR1/25 (NSNHVASGAGEAAIETQSSSSEEIV), encompassing the fusion region of the hybrid molecule PML-RARA, is recognized by CD4+ T cells in an HLA-DR11-restricted fashion. A T cell clone has been shown to lyse neoepitope-pulsed autologous cells or autologous cells transduced to express the PML-RARA fusion protein [56]. However, no PML-RARA-specific CD4+ T-cell responses were seen in four HLA-DR*11+ APL patients in a different study. Additionally, a DR11-restricted CD4+ T-cell clone obtained from a healthy donor unable to identify APL blasts from HLA-DR*11 patients since the APL cells lack the ability to present the peptide BCR1/25 [57]. As a result, PML-RARA is still regarded as a possible rather than a confirmed neoantigen.
Indels
Indels occurring in protein-coding genes frequently lead to frameshift peptides that are highly distinct from self and have reduced susceptibility to self-tolerance mechanisms. These frameshift peptides might be recognized by T-cells and induce cellular immune responses, thus indels might be an ideal source of tumor-derived neoantigens.
NPM1
In AML, nucleophosmin (NPM1) is among one of the most frequently mutated genes, being present in one-third of newly diagnosed cases [58]. Mutations in NPM1 are mostly restricted to exon 12 and cause a frameshift in the C-terminus, resulting in the replacement of the last 7 amino acids (WQWRKSL) with different residues [59]. Most NPM1 mutants carry a characteristic 4-bp frameshift insertion, with > 95% of mutations occurring between nucleotides 960 and 961. The substitution of different residues in the C-terminus of NPM1 destroys the nuclear export signal and causes the localization of the mutant protein (NPM1c) to the cytoplasm [59, 60]. Neoepitopes from NPM1c are capable of stimulating both CD4+ and CD8+ T-cell responses [19, 20]. Furthermore, NPM1c-specific T cells have been detected in a relapsed AML patient after receiving donor lymphocyte infusion [18]. The spontaneous appearance and persistence of NPM1c-specific T cells in AML patients may contribute to the maintenance of long-lasting remission [61]. Recently, a neoepitope (CLAVEEVSL) from NPM1c has been identified from HLA-A*02:01+ AML patients using tandem liquid chromatography-mass spectrometry (LC–MS/MS) technique, and then CD8+ T cells specific to this neoepitope have been isolated from healthy individuals. T cells transduced with the same TCR show the specific recognition and lysis of HLA-A*02:01-positive NPM1c AML cells [62]. As a result, the immunogenicity of NPM1c could be harnessed for immunotherapeutic approaches, including vaccine and adoptive T-cell therapy.
FLT3-ITD
Internal tandem duplication (ITD) mutations in the FMS-like tyrosine kinase 3 (FLT3) gene are present in about 30% of AML patients [63]. FLT3-ITD is a constitutively activated variant of the FLT3 tyrosine kinase receptor that enhances cellular proliferation and reduces apoptosis of hematopoietic blasts, leading to relatively poor clinical outcome and a higher risk of relapse in AML [64,65,66]. Duplication regions of FLT3-ITD most likely encode immunogenic neoepitopes that can be presented by individual HLA alleles. Neopeptides derived from FLT3-ITD of several AML cell lines or patients have been predicted with different affinity to HLA-A (A1, A2, A11, and A24) or HLA-B27, some of which showed intermediate or high affinity to HLA-A2 in a T2 cell-based assay [67]. Furthermore, CD8+ T cells from an FLT3-ITD-positive patient showed autologous anti-leukemic response to a neoepitope (YVDFREYEYY) encoded by the ITD protein region, in an HLA-A*01:01-restricted and FLT3-ITD-restricted manner [68]. Besides, due to the diversity of insert position of FLT3 and fragment length of ITD, a cohort of FLT3-ITD-positive patients is necessary to study the specific T-cell activities, including CD8+ and CD4+ T-cell responses to fully understand the potential of FLT3-ITD as an immunotherapeutic target.
Non-synonymous point mutations
Non-synonymous point mutations, which make up a significant portion of potential neoantigens for cancer immunotherapy, account for more than 90% of the driver mutations in malignancies, including AML. Some neoantigens generated from these mutations have high affinities to certain HLA alleles and are highly immunogenic when tested by vaccination. The mutated sites of these immunogenic peptides may be exposed to the TCR, leading to TCR rearrangement, or creating new anchor residues that increase binding affinities for HLA molecules.
IDH1
Missense mutations in isocitrate dehydrogenase 1 (IDH1) occur at specific arginine residues within the catalytic active sites of enzymes [69]. The hotspot mutation affects codon 132, resulting in a single amino acid substitution from arginine to histidine (R132H) [69]. IDH1R132H induces a leukemic DNA-methylation signature in a preclinical mouse model [69, 70], that is, similar to that observed in human IDH1-mutant AML [71]. The prognosis of AML patients harboring mutated IDH1 is generally poor [72], with an increased probability of relapse [73]. In syngeneic MHC-humanized mice, a peptide vaccine targeting IDH1R132H induces specific therapeutic MHC class II-restricted responses that are directed towards IDH1R132H-positive tumors [74]. The specific therapeutic T-helper cell responses to a corresponding peptide vaccine have also been observed in astrocytoma patients with IDH1R132H in a phase I clinical trial (NCT02454634) [75]. Although there is no reported evidence of immune responses against mutated IDH1 in AML, therapy targeting IDH1R132H should be equally applicable to IDH1R132H+ AML patients if the neoepitope is processed and presented appropriately, because the codon distribution of IDH1 mutations is not specific to the tissue origin of cancer.
KRAS
Mutations in the Kirsten rat sarcoma viral oncogene homolog (KRAS) oncogene have been considered as a driver of tumor initiation and maintenance [76]. The majority of mutant KRAS are recurrent “hot-spot” mutations that often occur at codon 12, 13, or 61, with codon 12 being the most frequent site of mutation [77, 78]. Substitution of the amino acid glycine (G) to aspartic acid (D) at this site, hereafter referred to as KRASG12D, is the most frequent KRAS mutant in human cancers. In AML, KrasG12D serves as a cooperating factor to promote myeloid leukemogenesis [79], and a contributor to clinical resistance to targeted inhibitors [80]. Vaccines that target KRASG12D have been utilized in clinical trials in patients with solid tumors, cooperates with chemotherapy and surgery or not [81]. Dendritic cells, DNA plasmids, viruses, yeast, or bacteria have also been used as delivery vectors in preclinical or clinical trials. Recently, a CD8+ T-cell reactivity against KRASG12D has been detected in tumor-infiltrating lymphocytes (TILs) from a patient with metastatic colon cancer, which could recognize and kill tumors in an HLA-C*08:02-restricted, KRASG12D-specific manner [82]. Subsequently, in a phase II clinical trial (NCT01174121), objective regression of metastases has been observed in a patient with metastatic colorectal cancer after infusion of HLA-C*08:02-restricted TILs composed of four different T-cell clonotypes that specifically targeted KRASG12D [21]. Furthermore, crystal structures of TCR-HLA-C complexes have revealed that TCRs of these reactive CD8+ T-cells can recognize KRASG12D nonamer with multiple conserved contacts through shared CDR2β and CDR3α, other than the corresponding wild-type peptide [83]. In addition, in a phase I/II clinical trial (NCT03480152), an mRNA vaccine containing KRASG12D has shown safety and efficacy in patients with metastatic gastrointestinal cancer. This vaccine can elicit KRASG12D-specific T-cell responses, and the specific TCRs have been verified and may be further used in adoptive T-cells therapy (ACT) in the future [84, 85]. As the ACT of T-cells genetically engineered to express tumor-reactive TCRs can elicit regression of widespread cancer in patients with metastatic disease, the TCRs isolated above may extend TCR gene therapy to HLA-C*08:02 patients whose tumor express KRASG12D, including AML patients.
TP53
TP53 mutated AML represents a distinct molecular subclass that has a poor response to chemotherapy and shows uniformly short overall survival (median survivals of 4–6 months) [86, 87]. The majority of TP53 variants are missense mutations evenly distributed throughout the coding sequence, particularly in the DNA-binding domain of exons 4 to 8 [88]. The p53 proteins encoded by mutant TP53 can be degraded by the proteasome, processed, and presented by HLA molecular to generate neoantigens recognizable by TCRs. TILs from patients with epithelial cancers have been shown to recognize autologous p53 neoantigens. These responses involve multiple TP53 mutations, including R175H, Y220C, G245S, R248Q, and R248W, and are restricted by a variety of HLA restriction elements [89, 90]. Interestingly, specific responses against mutated TP53, such as A161T, R267P, R175H, Y220C, and R248W, can also be induced in PBMCs from cancer patients with those mutations or from healthy donors without the matching expression [91,92,93]. Additionally, a peptide derived from p53R175H, HMTEVVRHC (mutant amino acid underlined), binds to HLA-A*02:01 with high affinity. Specific TCRs that recognize this peptide-HLA complex have been verified from a patient with metastatic colorectal cancer. These TCRs mediate the recognition of a variety of cancer cell lines endogenously expressing p53R175H, including ovarian cancer, uterine carcinoma, myeloma, and leukemia [94]. A TCR mimic antibody specific to this HLA-A*02:01-restricted p53R175H neoantigen has also been developed. This bispecific antibody can successfully activate T cells to lyse cancer cells that present the same neoantigen both in vitro and in mice [95]. Therefore, mutated TP53 is a promising target for many types of cancers due to its immunogenic and is a common and shared neoantigen.
Splicing variants
About 20% of AML patients harbor splicing variants, which represent a molecular feature of adverse AML with clinical implications. Additional messenger RNAs (mRNAs) are produced by splice isoforms, which are caused by retained introns, skipped exons, alternate 5′ or 3′ splice sites, or mutually exclusive exons. These mRNAs may be translated into a variety of proteins, which serves as an excellent source of neoantigens.
SF3B1
The splicing factor 3B subunit 1A (SF3B1) gene encodes a core component of the U2 nuclear ribonucleoprotein complex of the spliceosome [96]. Mutations of SF3B1 are found in about 5.6%–12.5% of de novo AML patients and are more frequent in secondary AML [23, 97, 98]. The common hotspot mutations include the consecutive HEAT domain of the C-terminal region, K700, or the conserved amino acids K666, H662, R625, and E622. Neoepitopes derived from SF3B1R625-related junctions have been found in tumor cells expressing mutated SF3B1, other than cells with wild-type SF3B1. Patients with SF3B1R625 mutations contain CD8+ T-cells specific for these neoepitopes, most of which display the effector/memory phenotype. These results indicate that SF3B1 variant-derived neoepitopes have been recognized by the immune system in patients harboring such mutations [99].
In addition to SF3B1, an HLA class I immunopeptidome has been established through the identification of peptides eluted from HLA—I molecules by means of LC–MS/MS. The results indicate that the proteasome-generated spliced peptide pool accounts for one-third of the entire HLA class I immunopeptidome in terms of diversity and one-fourth in terms of abundance [100], further support that spliced peptides represent a distinct pool of neoantigens with particular immunological characteristics.
Neoantigen-specific TCRs targeting AML cells
Experimental efforts to map TCR repertoires for a particular antigen have benefited from recent developments in immunology and high-throughput TCR sequencing techniques. The prerequisite is to accurately select the tumor-reactive and neoantigen-specific T cells. This section will discuss the sample sources and associated markers that are used to identify neoantigen-specific T cells, as well as the neoantigen-specific TCRs targeting AML cells.
Sources of neoantigen-specific T cells
Neoantigen-reactive T cells are commonly found in patients with solid or hematologic malignancies. Patient samples, including TILs and peripheral circulating lymphocytes, are the main sources of tumor-specific T cells [22, 68, 103,104,105]. Peripheral blood of an AML patient in complete remission has been found harboring specific T cells targeting an FLT3-ITD derived neoepitope, which secrets significant amounts of IFN-γ and Granzyme B after receiving the stimulation of this neoepitope [68]. Similarly, both CD4+ and CD8+ T cells against mutated NPM1 have been found in AML patients [20], and the majority of these cells are effector memory and central memory T cells, which may contribute to the maintenance of long-lasting remissions [61]. Additionally, tumor-specific T cells can also be detected in the body fluids of patients, such as urine, ascites, and pleural effusions [106, 107].
Peripheral blood from healthy donors is a second emerging source for neoantigen-specific T cells, which is less invasive in contrast to tumor biopsies and is thus readily available. It has been shown that healthy donors exhibit more frequent and efficient CALR-mutant-specific T-cell responses compared with CALR-mutant patients [108]. Neoepitopes derived from indels and fusion proteins of AML patients, such as mutated NPM1 and CBFB-MYH11, have also been found to be recognized by T cells from healthy donors in several studies [53, 62]. Therefore, induction of neoantigen-specific T cells from healthy donors may provide a means for adoptive cell therapy of hematologic malignancies.
Related markers to capture neoantigen-specific T cells
Neoantigen-specific T cells can be identified and isolated from bulk lymphocytes of patients or healthy donors using MHC multimers or tetramers carrying the same neoepitopes. However, it is estimated that just 0.002% of the entire lymphocyte population is made up of these T cells [105]. Several markers on T cell surface or T-cell secreted cytokines have been suggested for the further enrichment of neoantigen-reactive T cell clones.
PD-1, also known as CD279, is characterized as an inhibitory receptor on T cells. After in vitro expansion, tumor-reactive TILs have a greater percentage of PD-1-expressing CD8 + T cells, which produce much higher tumor-specific IFN-γ compared with the PD-1-negative counterparts [109]. Similar to this, the expression of PD-1 on peripheral blood lymphocytes has the potential to be used as a biomarker to detect T cells that are responsive to neoantigens [110]. In contrast to PD-1-negative cells, which are the limiting factor in the tumor specificity of cells produced from bulk CD8+ TILs, PD-1-positive T cell subsets have been shown to have advantages for adoptive T-cell treatment [111]. In another research, it was discovered that CD8+ TILs express PD-1, TIM-3 (a co-inhibitory receptor restricted to IFN-γ-producing T cells), and LAG-3 (an inhibitory receptor), which can be used to identify the autologous tumor-reactive repertoires, including mutant neoantigenic T cells [112].
4-1BB, also known as CD137 and TNFRSF9, is a costimulatory member of the TNF receptor family and has emerged as an important mediator of survival and proliferation [113, 114]. 4-1BB is essential for the survival of activated and memory CD8+ T cells. After TCR stimulation, 4-1BB is transiently upregulated from 12 h to up to 5 days [115]. Epitope discovery based on CD137+ selection has proven to be an efficient and sensitive in vitro technique for rapidly identifying and isolating antigen-specific CD8+ T cells that present at low frequencies and display heterogeneous functional profiles [115]. Through the enrichment of T cells from TILs by sorting for CD137+CD8+ markers, 27 TCRs that recognize 14 neoantigens produced by autologous tumor cells have been isolated from 6 patients [116]. Memory T cells targeting the mutated KRAS G12D and KRAS G12V have also been identified and isolated with 4-1BB as the T-cell activation marker in another study [117].
CD39, a transmembrane extracellular ATPase, is widely expressed by regulatory T cells, B cells, and some tumor cells. CD8+ T cells with high CD39 expression in TILs exhibit an exhausted phenotype with the reduced production of INF-γ, TNF, IL-2, and high expression of coinhibitory receptors [118]. Expression of genes related to cell proliferation and exhaustion which are characteristics of chronically stimulated T cells is enriched in CD39+CD8+ TILs [119]. A distinct population of CD8+ TILs that co-express CD39 and CD103 possesses an exhausted tissue-resident memory state, is capable of recognizing and eradicating autologous tumor cells in an MHC-class I-dependent manner [120]. Most antitumor neoantigen-reactive TILs are found in the differentiated CD39+ state [121]. However, in addition to nonresponders receiving adoptive T cell therapy, responsive patients have also been observed to harbor CD39− stem-like neoantigen-specific TILs [121].
Cytokines, such as IFN-γ and IL-2, can be used as markers for screening neoantigen-specific TCRs. A brand-new approach for identifying neoantigen-specific T cells has been developed recently [122]. It involves co-culturing TILs with antigen-presenting cells that have been pulsed with neoantigens and the single-cell RNA sequencing analysis of T cells. Neoantigen-reactive cells are those expressing high levels of IFN-γ and IL-2. Using this method, neoantigen-specific TCR sequences have been successfully identified from two melanoma and one colorectal cancer patient [123].
Taken together, the sources of neoantigen-specific T cells are diverse but limited in frequency. The status of CD8+ T cells is quite heterogeneous, both within and between patients [124]. There is currently no specific method or marker to exclusively recognize neoantigen-specific T cells, making the application of the aforementioned methodologies to identify and enrich such T cells challenging. To produce neoantigen-specific TCRs in particular, these methods may be helpful for enriching and expanding neoantigen-reactive T cells before going on to downstream analysis [125].
Identification of neoantigen-specific TCRs to AML
IN AML, direct identification of neoantigen-specific T cells from patients is hampered by lymphopenia and/or disease-related immunological dysfunction. T cells specific for certain neoantigens may not be present in adequate numbers in PBMCs isolated from AML patients in remission after chemotherapy. PBMCs from healthy donors have therefore been given priority. Using tetramer technology, a significant percentage of PBMCs from six HLA-A*02:01+ healthy individuals were directly identified as mutated NPM1-specific T cells. Some of these T cell clones have been effectively expanded and displayed specific cytotoxic activity against AML cells that express mutant NPM1. For example, T cells transferred with the modified TCR sequences (TRAV12-2 and TRBV5-1) expressed by the responder clone show the same specific activity against mutant NPM1 [62]. Similarly, high-avidity CD8+ CBFB-MYH11 epitope-specific T cell clones have been isolated by the neopeptide-tetramer from HLA-B*40:01+ healthy donors and subjected to TCR sequencing to identify the paired α and β chains. CD8+ T cells transduced with these high-avidity TCRs exhibit antileukemic activity in vitro [53].
These findings imply that specific anti-leukemic TCR rearrangements may be induced by neoantigens arising from genetic aberrant events of AML notably indel and fusion and that these changes can be accurately captured by specialized antibodies and TCR sequencing. An important source of neoantigen-specific TCRs for AML, is immune cells from healthy donor since chemotherapy-induced immunological dysfunction in AML patients makes it difficult for them to generate enough anti-leukemic immune responses. Recently, a time-efficient approach to identify pilot neoantigen-specific TCRs from HLA-matched healthy donors has been established, and two of the three chosen TCRs have demonstrated dose-dependent recognition of the corresponding neoantigens [126]. From the time that the neoantigen is stimulated until specific T cells are obtained, this protocol, which could also work for AML, only takes two weeks. Additionally, other than tetramer, immune checkpoints (such as PD-1, TIM3, and 4-1BB), the stem-cell-like marker CD39, and the cytokines mentioned above can also be employed to broaden the screening of neoantigen-specific T-cell for AML (Fig. 1).

Overview of the procedure for identifying neoantigen-specific TCRs from healthy donors
Clinical experience of TCR-based immunotherapy in AML
IT is estimated that about 16% of TCR-T cell-based clinical trials are designed for hematological malignancies, in particular AML (Table 2) [127]. The majority of clinical trials are based on TCR-engineered T cells against WT1, a classic TAA [128]. In patients with relapsed or high-risk leukemia, the administration of WT1-specific donor-derived CD8+ cytotoxic T cell clones is well tolerated, and the transferred cells exhibit antileukemic activities in two patients (NCT00052520) [129]. In a subsequent phase 1 clinical trial (NCT01640301), a high-affinity WT1-specific TCR was inserted into Epstein-Bar virus-specific donor CD8+ T cells and twelve AML patients with poor-risk prophylactically received these WT1-specific TCR-T cells. The treatments showed significant benefits related to relapse-free survival, overall survival, relapse, and non-relapse mortality [9]. In another first-in-human trial (UMIN000011519), eight patients with refractory AML or high-risk MDS were treated twice with autologous T cells engineered to express a TCR specific for the WT1234-243 peptide in an HLA-A*24:02-restricted manner, along with siRNAs to eliminate expression of endogenous TCR chains, followed by sequential WT1 peptide vaccines. The TCR-T cells persisted throughout the study period in five patients, four of whom survived more than 12 months [10, 130, 131]. An optimized TCR construct has been established recently, which contains a high-affinity HA-1 specific TCR, a CD8+ coreceptor, a safety switch (iCasp9), and a tracking marker. T cells that express this TCR construct are highly functional against HA-1+ leukemia cells in vitro, and they were rapidly and completely eliminated using a safety switch [132]. These engineered T cells are being evaluated on patients with hematological malignancies in an ongoing clinical trial (NCT03326921). In addition, clinical trials of TCR-T cell-based therapy that targets PRAME, a well-known cancer-testis antigen, have started in recent years. Two of these trials are NCT022743611 (BPX-701, Bellicum Pharmaceuticals) and NCT03503968 (MDG1011, Medigene AG).
Despite the lack of clinical studies for neoantigen-specific TCR-T cell-based therapy in AML, deep and durable tumor shrinkage has been observed in a heavily pretreated patient with metastatic pancreatic ductal adenocarcinoma who received an infusion of autologous T cells transduced with a TCR specific for mutant KRASG12D [85]. This finding strengthens the clinical application of neoantigen-specific TCR-T cells. Likewise, as more and more TCRs specific to neoantigens that arise from genetic aberrations of AML with different HLA-restriction are being identified in the future, patients who have this particular HLA allele and express the same neoantigens may potentially benefit from this TCR transduction therapy (Fig. 2).

Overview of the neoantigen-specific TCR-based ACT process
Comparison of TCR- and CAR-T cell therapy for AML
Another engineered T-cell-based therapy known as CAR-T is frequently employed in clinical trials to treat relapsed or refractory AML patients [133]. Both TCR- and CAR- therapies modify and expand T lymphocytes from patients or donors ex vivo before re-infusing them into the patient’s body to eradicate cancerous cells. Typically, viruses (such as lentiviral and retroviral) or CRISPR/Cas9 are used to introduce TCR or CAR structures into T cells. Both therapies have certain advantages and disadvantages, which have been well-reviewed elsewhere [134, 135]. Other than these, TCR- and CAR-modified T cells differ greatly, as described below.
Construct
The TCR complex is a heterodimer consisting of alpha and beta chains. Unlike TCR, CARs are synthetic receptors created from a combination of an antibody-derived single-chain variable fragment (scFV), a T-cell receptor-derived CD3ζ domain, a transmembrane domain, and one or more intracellular co-stimulatory domains (such as CD28, 4-1BB/CD137, ICOS, and/or OX40) that have evolved across five generations [136]. Other than that, safety switches have been introduced to TCR and CAR constructs [132, 137]. Modified T cells carrying this construct have been used in clinical trials for the treatment of AML patients to improve safety [132].
Targets and MHC involvement
The scFV domain of CAR-T cells binds to antigens expressed on the surface of cancer cells in an MHC-independent manner, therefore, targets for CAR-T therapy are all surface proteins, such as CD33 [138], CD123 [139, 140], NKG2D ligand [141], CD70 [142], CD13-TIM3 [4] for AML patients. In contrast to CAR-T, the heterodimer expressed in TCR-T cells recognizes peptides presented by MHC molecules. As a result, the spectrum of targets for TCR-T cells has expanded to include both intracellular and surface antigens. Currently, TCR-T cell therapy for AML patients concentrates on neoantigens, such as NPM1 and CBFB-MYH11, as well as TAAs like WT1, PRAME, and HA-1.
Toxicities
Modified T cells are potential for “On-target, off-tumor” toxicity in cases where the antigenic target is co-expressed on normal cells, such as CAR-T cells targeting leukemia-associated antigens and TCR-T cells targeting TAAs [136]. Neoantigens are only found in malignant cells, therefore employing them as targets might help with this problem. Other than this, CAR-related toxicities also include cytokine release syndrome and neurological toxicities, which are also referred to as “on/off tissue toxicity” effects and have varying degrees of severity but a similar clinical presentation [143].
Challenges for the application of TCR-T cells
Although neoantigen-specific T cell-based immunotherapy has shown promising potential in treating AML patients without harming normal tissues, there are still challenges ahead for the application of TCR-T cells. First, the accurate identification of neoantigens presented by HLA molecules on the cell surface is a major obstacle. LC coupled to MS/MS has proved to be a fast and less biased technology to advance the identification of endogenously processed and presented HLA-ligands, although sample handling methods, instrumentations, and data searching algorithms are still required to be improved. Currently, integrating MS HLA-ligand profiling with other “-omic” technologies, such as whole exon sequencing and RNA-sequencing, is indeed of help to identify tumor-specific antigens for AML immunotherapy [37]. Second, the induction of neoantigen-specific TCRs directly from AML patients may be restricted by illness-related immunodeficiency [53, 62]. PBMCs from healthy donors may provide a source to induce neoantigen-specific T cells. Third, the mismatch between exogenous and endogenous chains may hinder the correct expression of modified TCRs on the surface of T cells. Substitution the constant region of the TCR construct with a mouse version may reduce the frequency of alpha and beta chain mismatches [144]. With these improvements, the TCR-T cell application will become feasible on clinical timescales.
Conclusion
TCR-T cell-based immunotherapy for hematologic malignancies has made significant strides, especially in the treatment of high-risk AML. The major challenge is identifying appropriate targets. A variety of potential TAAs are being studied in preclinical or clinical settings as targets for TCR-T cell therapy, but special attention must be paid to the adverse effects caused by “off-targets”, because TAAs are also expressed in a small number of normal tissues. Neoantigens seem to be the best candidates for TCR-T targets in this regard since their expression is limited to AML cells. Encouragingly, high-avidity TCRs specific for mutant NPM1 and CBFB-MYH11 have been identified from the repertoires of healthy donors. T cells that have been genetically modified to express these TCRs shown specific killing abilities against AML cell lines as well as blast cells harboring mutated NPM1 and CBFB-MYH11, respectively. Given that the majority of AML patients exhibit these neoantigens, the results of future clinical trials are expected. Further research is needed into TCR-T cells that are directed against additional recurrent somatic mutations in AML, including IDH1/IDH2, FLT3-ITD, DNMT3A, and AML1-ETO.
In conclusion, neoantigen-specific TCR-T cell therapy has good potential to treat AML. Illustrating the interaction between TCRs and neoantigens that are responsible for T cell reactivity, anti-leukemic responses, and clinical effects is the cornerstone. The security and efficacy of genetically modified TCR-T cells are vital as well. The present article reviewed advances in neoantigens for AML and proximal new techniques which can identify neoantigen-specific TCRs and activate a T-cell response. There are several restrictions, nevertheless, that need to be addressed in further study.
Availability of data and materials
Not applicable.
Abbreviations
- ACT:
-
Adoptive T-cells therapy
- AML:
-
Acute myeloid leukemia
- APL:
-
Acute promyelocytic leukemia
- CAR:
-
Chimeric antigen receptor
- CBF:
-
Core binding factor
- GVL:
-
Graft-versus leukemia
- HLA:
-
Human leukocyte antigen
- HSCT:
-
Hematopoietic stem cell transplantation
- IDH1:
-
Isocitrate dehydrogenase 1
- Indel:
-
Insertion and deletion
- ITD:
-
Internal tandem duplication
- KRAS:
-
Kirsten rat sarcoma viral oncogene homolog
- LC–MS/MS:
-
Liquid chromatography–mass spectrometry
- PBMC:
-
Peripheral blood mononuclear cell
- PDX:
-
Patient-derived murine xenograft
- SF3B1:
-
Splicing factor 3B subunit 1A
- TAA:
-
Tumor-associated antigens
- TCR:
-
T cell receptor
- TIL:
-
Tumor-infiltrating lymphocytes
References
Passweg JR, Baldomero H, Chabannon C, Basak GW, de la Camara R, Corbacioglu S, Dolstra H, Duarte R, Glass B, Greco R, Lankester AC, Mohty M, Peffault DLR, Snowden JA, Yakoub-Agha I, Kroger N. Hematopoietic cell transplantation and cellular therapy survey of the EBMT: monitoring of activities and trends over 30 years. Bone Marrow Transplant. 2021;56:1651–64.
Xu LP, Wu DP, Han MZ, Huang H, Liu QF, Liu DH, Sun ZM, Xia LH, Chen J, Wang HX, Wang C, Li CF, Lai YR, Wang JM, Zhou DB, Chen H, Song YP, Liu T, Liu KY, Huang XJ. A review of hematopoietic cell transplantation in China: data and trends during 2008–2016. Bone Marrow Transplant. 2017;52:1512–8.
Sweeney C, Vyas P. The Graft-Versus-Leukemia Effect in AML. Front Oncol. 2019;9:1217.
He X, Feng Z, Ma J, Ling S, Cao Y, Gurung B, Wu Y, Katona BW, O’Dwyer KP, Siegel DL, June CH, Hua X. Bispecific and split CAR T cells targeting CD13 and TIM3 eradicate acute myeloid leukemia. Blood. 2020;135:713–23.
Myburgh R, Kiefer JD, Russkamp NF, Magnani CF, Nunez N, Simonis A, Pfister S, Wilk CM, McHugh D, Friemel J, Muller AM, Becher B, Munz C, van den Broek M, Neri D, Manz MG. Anti-human CD117 CAR T-cells efficiently eliminate healthy and malignant CD117-expressing hematopoietic cells. Leukemia. 2020;34:2688–703.
June CH, Sadelain M. Chimeric antigen receptor therapy. N Engl J Med. 2018;379:64–73.
Rath JA, Arber C. Engineering Strategies to Enhance TCR-Based Adoptive T Cell Therapy. Cells-Basel. 2020;9:45.
D’Ippolito E, Schober K, Nauerth M, Busch DH. T cell engineering for adoptive T cell therapy: safety and receptor avidity. Cancer Immunol Immunother. 2019;68:1701–12.
Chapuis AG, Egan DN, Bar M, Schmitt TM, McAfee MS, Paulson KG, Voillet V, Gottardo R, Ragnarsson GB, Bleakley M, Yeung CC, Muhlhauser P, Nguyen HN, Kropp LA, Castelli L, Wagener F, Hunter D, Lindberg M, Cohen K, Seese A, McElrath MJ, Duerkopp N, Gooley TA, Greenberg PD. T cell receptor gene therapy targeting WT1 prevents acute myeloid leukemia relapse post-transplant. Nat Med. 2019;25:1064–72.
Tawara I, Kageyama S, Miyahara Y, Fujiwara H, Nishida T, Akatsuka Y, Ikeda H, Tanimoto K, Terakura S, Murata M, Inaguma Y, Masuya M, Inoue N, Kidokoro T, Okamoto S, Tomura D, Chono H, Nukaya I, Mineno J, Naoe T, Emi N, Yasukawa M, Katayama N, Shiku H. Safety and persistence of WT1-specific T-cell receptor gene-transduced lymphocytes in patients with AML and MDS. Blood. 2017;130:1985–94.
Amir AL, van der Steen DM, van Loenen MM, Hagedoorn RS, de Boer R, Kester MD, de Ru AH, Lugthart GJ, van Kooten C, Hiemstra PS, Jedema I, Griffioen M, van Veelen PA, Falkenburg JH, Heemskerk MH. PRAME-specific Allo-HLA-restricted T cells with potent antitumor reactivity useful for therapeutic T-cell receptor gene transfer. Clin Cancer Res. 2011;17:5615–25.
Marijt WA, Heemskerk MH, Kloosterboer FM, Goulmy E, Kester MG, van der Hoorn MA, van Luxemburg-Heys SA, Hoogeboom M, Mutis T, Drijfhout JW, van Rood JJ, Willemze R, Falkenburg JH. Hematopoiesis-restricted minor histocompatibility antigens HA-1- or HA-2-specific T cells can induce complete remissions of relapsed leukemia. Proc Natl Acad Sci U S A. 2003;100:2742–7.
Coulie PG, Van den Eynde BJ, van der Bruggen P, Boon T. Tumour antigens recognized by T lymphocytes: at the core of cancer immunotherapy. Nat Rev Cancer. 2014;14:135–46.
Rosenberg SA, Restifo NP. Adoptive cell transfer as personalized immunotherapy for human cancer. Science. 2015;348:62–8.
Blass E, Ott PA. Advances in the development of personalized neoantigen-based therapeutic cancer vaccines. Nat Rev Clin Oncol. 2021;18:215–29.
Sahin U, Tureci O. Personalized vaccines for cancer immunotherapy. Science. 2018;359:1355–60.
Blankenstein T, Leisegang M, Uckert W, Schreiber H. Targeting cancer-specific mutations by T cell receptor gene therapy. Curr Opin Immunol. 2015;33:112–9.
Hofmann S, Gotz M, Schneider V, Guillaume P, Bunjes D, Dohner H, Wiesneth M, Greiner J. Donor lymphocyte infusion induces polyspecific CD8(+) T-cell responses with concurrent molecular remission in acute myeloid leukemia with NPM1 mutation. J Clin Oncol. 2013;31:e44–7.
Greiner J, Schneider V, Schmitt M, Gotz M, Dohner K, Wiesneth M, Dohner H, Hofmann S. Immune responses against the mutated region of cytoplasmatic NPM1 might contribute to the favorable clinical outcome of AML patients with NPM1 mutations (NPM1mut). Blood. 2013;122:1087–8.
Greiner J, Ono Y, Hofmann S, Schmitt A, Mehring E, Gotz M, Guillaume P, Dohner K, Mytilineos J, Dohner H, Schmitt M. Mutated regions of nucleophosmin 1 elicit both CD4(+) and CD8(+) T-cell responses in patients with acute myeloid leukemia. Blood. 2012;120:1282–9.
Tran E, Robbins PF, Lu YC, Prickett TD, Gartner JJ, Jia L, Pasetto A, Zheng Z, Ray S, Groh EM, Kriley IR, Rosenberg SA. T-Cell transfer therapy targeting mutant KRAS in cancer. N Engl J Med. 2016;375:2255–62.
Zacharakis N, Chinnasamy H, Black M, Xu H, Lu YC, Zheng Z, Pasetto A, Langhan M, Shelton T, Prickett T, Gartner J, Jia L, Trebska-McGowan K, Somerville RP, Robbins PF, Rosenberg SA, Goff SL, Feldman SA. Immune recognition of somatic mutations leading to complete durable regression in metastatic breast cancer. Nat Med. 2018;24:724–30.
Ley TJ, Miller C, Ding L, Raphael BJ, Mungall AJ, Robertson A, Hoadley K, Triche TJ, Laird PW, Baty JD, Fulton LL, Fulton R, Heath SE, Kalicki-Veizer J, Kandoth C, Klco JM, Koboldt DC, Kanchi KL, Kulkarni S, Lamprecht TL, Larson DE, Lin L, Lu C, McLellan MD, McMichael JF, Payton J, Schmidt H, Spencer DH, Tomasson MH, Wallis JW, Wartman LD, Watson MA, Welch J, Wendl MC, Ally A, Balasundaram M, Birol I, Butterfield Y, Chiu R, Chu A, Chuah E, Chun HJ, Corbett R, Dhalla N, Guin R, He A, Hirst C, Hirst M, Holt RA, Jones S, Karsan A, Lee D, Li HI, Marra MA, Mayo M, Moore RA, Mungall K, Parker J, Pleasance E, Plettner P, Schein J, Stoll D, Swanson L, Tam A, Thiessen N, Varhol R, Wye N, Zhao Y, Gabriel S, Getz G, Sougnez C, Zou L, Leiserson MD, Vandin F, Wu HT, Applebaum F, Baylin SB, Akbani R, Broom BM, Chen K, Motter TC, Nguyen K, Weinstein JN, Zhang N, Ferguson ML, Adams C, Black A, Bowen J, Gastier-Foster J, Grossman T, Lichtenberg T, Wise L, Davidsen T, Demchok JA, Shaw KR, Sheth M, Sofia HJ, Yang L, Downing JR, Eley G. Genomic and epigenomic landscapes of adult de novo acute myeloid leukemia. N Engl J Med. 2013;368:2059–74.
Papaemmanuil E, Gerstung M, Bullinger L, Gaidzik VI, Paschka P, Roberts ND, Potter NE, Heuser M, Thol F, Bolli N, Gundem G, Van Loo P, Martincorena I, Ganly P, Mudie L, McLaren S, O’Meara S, Raine K, Jones DR, Teague JW, Butler AP, Greaves MF, Ganser A, Dohner K, Schlenk RF, Dohner H, Campbell PJ. Genomic classification and prognosis in acute myeloid leukemia. N Engl J Med. 2016;374:2209–21.
Parker KC, Bednarek MA, Coligan JE. Scheme for ranking potential HLA-A2 binding peptides based on independent binding of individual peptide side-chains. J Immunol. 1994;152:163–75.
Zhang L, Chen Y, Wong HS, Zhou S, Mamitsuka H, Zhu S. TEPITOPEpan: extending TEPITOPE for peptide binding prediction covering over 700 HLA-DR molecules. PLoS ONE. 2012;7: e30483.
Zhang H, Lund O, Nielsen M. The PickPocket method for predicting binding specificities for receptors based on receptor pocket similarities: application to MHC-peptide binding. Bioinformatics. 2009;25:1293–9.
Zhang GL, DeLuca DS, Keskin DB, Chitkushev L, Zlateva T, Lund O, Reinherz EL, Brusic V. MULTIPRED2: a computational system for large-scale identification of peptides predicted to bind to HLA supertypes and alleles. J Immunol Methods. 2011;374:53–61.
Bordner AJ, Mittelmann HD. MultiRTA: a simple yet reliable method for predicting peptide binding affinities for multiple class II MHC allotypes. BMC Bioinformatics. 2010;11:482.
Reche PA, Reinherz EL. Prediction of peptide-MHC binding using profiles. Methods Mol Biol. 2007;409:185–200.
Rammensee H, Bachmann J, Emmerich NP, Bachor OA, Stevanovic S. SYFPEITHI: database for MHC ligands and peptide motifs. Immunogenetics. 1999;50:213–9.
Jurtz V, Paul S, Andreatta M, Marcatili P, Peters B, Nielsen M. NetMHCpan-4.0: Improved Peptide-MHC Class I interaction predictions integrating eluted ligand and peptide binding affinity data. J Immunol. 2017;199:3360–8.
Nielsen M, Andreatta M. NetMHCpan-30; improved prediction of binding to MHC class I molecules integrating information from multiple receptor and peptide length datasets. Genome Med. 2016;8:33.
Hoof I, Peters B, Sidney J, Pedersen LE, Sette A, Lund O, Buus S, Nielsen M. NetMHCpan, a method for MHC class I binding prediction beyond humans. Immunogenetics. 2009;61:1–13.
Nielsen M, Lundegaard C, Blicher T, Peters B, Sette A, Justesen S, Buus S, Lund O. Quantitative predictions of peptide binding to any HLA-DR molecule of known sequence: NetMHCIIpan. Plos Comput BioL. 2008;4: e1000107.
Vita R, Mahajan S, Overton JA, Dhanda SK, Martini S, Cantrell JR, Wheeler DK, Sette A, Peters B. The Immune Epitope Database (IEDB): 2018 update. Nucleic Acids Res. 2019;47:D339–43.
Ehx G, Larouche JD, Durette C, Laverdure JP, Hesnard L, Vincent K, Hardy MP, Theriault C, Rulleau C, Lanoix J, Bonneil E, Feghaly A, Apavaloaei A, Noronha N, Laumont CM, Delisle JS, Vago L, Hebert J, Sauvageau G, Lemieux S, Thibault P, Perreault C. Atypical acute myeloid leukemia-specific transcripts generate shared and immunogenic MHC class-I-associated epitopes. Immunity. 2021;54:737–52.
Zhou C, Wei Z, Zhang Z, Zhang B, Zhu C, Chen K, Chuai G, Qu S, Xie L, Gao Y, Liu Q. pTuneos: prioritizing tumor neoantigens from next-generation sequencing data. Genome Med. 2019;11:67.
Wang TY, Wang L, Alam SK, Hoeppner LH, Yang R. ScanNeo: identifying indel-derived neoantigens using RNA-Seq data. Bioinformatics. 2019;35:4159–61.
Zhang J, Mardis ER, Maher CA. INTEGRATE-neo: a pipeline for personalized gene fusion neoantigen discovery. Bioinformatics. 2017;33:555–7.
Bais P, Namburi S, Gatti DM, Zhang X, Chuang JH. CloudNeo: a cloud pipeline for identifying patient-specific tumor neoantigens. Bioinformatics. 2017;33:3110–2.
Tappeiner E, Finotello F, Charoentong P, Mayer C, Rieder D, Trajanoski Z. TIminer: NGS data mining pipeline for cancer immunology and immunotherapy. Bioinformatics. 2017;33:3140–1.
Zhou Z, Wu J, Ren J, Chen W, Zhao W, Gu X, Chi Y, He Q, Yang B, Wu J, Chen S. TSNAD v2.0: A one-stop software solution for tumor-specific neoantigen detection. Comput Struct Biotechnol J. 2021;19:4510–6.
Zhou Z, Lyu X, Wu J, Yang X, Wu S, Zhou J, Gu X, Su Z, Chen S. TSNAD: an integrated software for cancer somatic mutation and tumour-specific neoantigen detection. R Soc Open Sci. 2017;4: 170050.
Bjerregaard AM, Nielsen M, Hadrup SR, Szallasi Z, Eklund AC. MuPeXI: prediction of neo-epitopes from tumor sequencing data. Cancer Immunol Immunother. 2017;66:1123–30.
Hundal J, Carreno BM, Petti AA, Linette GP, Griffith OL, Mardis ER, Griffith M. pVAC-Seq: A genome-guided in silico approach to identifying tumor neoantigens. Genome Med. 2016;8:11.
Kim S, Kim HS, Kim E, Lee MG, Shin EC, Paik S, Kim S. Neopepsee: accurate genome-level prediction of neoantigens by harnessing sequence and amino acid immunogenicity information. Ann Oncol. 2018;29:1030–6.
Speck NA, Gilliland DG. Core-binding factors in haematopoiesis and leukaemia. Nat Rev Cancer. 2002;2:502–13.
Liu P, Tarle SA, Hajra A, Claxton DF, Marlton P, Freedman M, Siciliano MJ, Collins FS. Fusion between transcription factor CBF beta/PEBP2 beta and a myosin heavy chain in acute myeloid leukemia. Science. 1993;261:1041–4.
Castilla LH, Garrett L, Adya N, Orlic D, Dutra A, Anderson S, Owens J, Eckhaus M, Bodine D, Liu PP. The fusion gene Cbfb-MYH11 blocks myeloid differentiation and predisposes mice to acute myelomonocytic leukaemia. Nat Genet 1999;23:144–146.
Kuo YH, Landrette SF, Heilman SA, Perrat PN, Garrett L, Liu PP, Le Beau MM, Kogan SC, Castilla LH. Cbf beta-SMMHC induces distinct abnormal myeloid progenitors able to develop acute myeloid leukemia. Cancer Cell. 2006;9:57–68.
Zhao L, Melenhorst JJ, Alemu L, Kirby M, Anderson S, Kench M, Hoogstraten-Miller S, Brinster L, Kamikubo Y, Gilliland DG, Liu PP. KIT with D816 mutations cooperates with CBFB-MYH11 for leukemogenesis in mice. Blood 2012;119:1511–1521.
Biernacki MA, Foster KA, Woodward KB, Coon ME, Cummings C, Cunningham TM, Dossa RG, Brault M, Stokke J, Olsen TM, Gardner K, Estey E, Meshinchi S, Rongvaux A, Bleakley M. CBFB-MYH11 fusion neoantigen enables T cell recognition and killing of acute myeloid leukemia. J Clin Invest. 2020;130:5127–41.
de The H, Chen Z. Acute promyelocytic leukaemia: novel insights into the mechanisms of cure. Nat Rev Cancer. 2010;10:775–83.
Wang ZY, Chen Z. Acute promyelocytic leukemia: from highly fatal to highly curable. Blood. 2008;111:2505–15.
Gambacorti-Passerini C, Grignani F, Arienti F, Pandolfi PP, Pelicci PG, Parmiani G. Human CD4 lymphocytes specifically recognize a peptide representing the fusion region of the hybrid protein pml/RAR alpha present in acute promyelocytic leukemia cells. Blood. 1993;81:1369–75.
Dermime S, Bertazzoli C, Marchesi E, Ravagnani F, Blaser K, Corneo GM, Pogliani E, Parmiani G, Gambacorti-Passerini C. Lack of T-cell-mediated recognition of the fusion region of the pml/RAR-alpha hybrid protein by lymphocytes of acute promyelocytic leukemia patients. Clin Cancer Res. 1996;2:593–600.
Heath EM, Chan SM, Minden MD, Murphy T, Shlush LI, Schimmer AD. Biological and clinical consequences of NPM1 mutations in AML. Leukemia. 2017;31:798–807.
Falini B, Mecucci C, Tiacci E, Alcalay M, Rosati R, Pasqualucci L, La Starza R, Diverio D, Colombo E, Santucci A, Bigerna B, Pacini R, Pucciarini A, Liso A, Vignetti M, Fazi P, Meani N, Pettirossi V, Saglio G, Mandelli F, Lo-Coco F, Pelicci PG, Martelli MF. Cytoplasmic nucleophosmin in acute myelogenous leukemia with a normal karyotype. N Engl J Med. 2005;352:254–66.
Falini B, Brunetti L, Sportoletti P, Martelli MP. NPM1-mutated acute myeloid leukemia: from bench to bedside. Blood. 2020;136:1707–21.
Forghieri F, Riva G, Lagreca I, Barozzi P, Vallerini D, Morselli M, Paolini A, Bresciani P, Colaci E, Maccaferri M, Gilioli A, Nasillo V, Messerotti A, Pioli V, Arletti L, Giusti D, Bettelli F, Celli M, Donatelli F, Corradini G, Basso S, Gurrado A, Cellini M, Trenti T, Marasca R, Narni F, Martelli MP, Falini B, Potenza L, Luppi M, Comoli P. Characterization and dynamics of specific T cells against nucleophosmin-1 (NPM1)-mutated peptides in patients with NPM1-mutated acute myeloid leukemia. Oncotarget. 2019;10:869–82.
van der Lee DI, Reijmers RM, Honders MW, Hagedoorn RS, de Jong RC, Kester MG, van der Steen DM, de Ru AH, Kweekel C, Bijen HM, Jedema I, Veelken H, van Veelen PA, Heemskerk MH, Falkenburg J, Griffioen M. Mutated nucleophosmin 1 as immunotherapy target in acute myeloid leukemia. J Clin Invest. 2019;129:774–85.
Yokota S, Kiyoi H, Nakao M, Iwai T, Misawa S, Okuda T, Sonoda Y, Abe T, Kahsima K, Matsuo Y, Naoe T. Internal tandem duplication of the FLT3 gene is preferentially seen in acute myeloid leukemia and myelodysplastic syndrome among various hematological malignancies A study on a large series of patients and cell lines. Leukemia. 1997;11:1605–9.
Garg M, Nagata Y, Kanojia D, Mayakonda A, Yoshida K, Haridas KS, Zang ZJ, Okuno Y, Shiraishi Y, Chiba K, Tanaka H, Miyano S, Ding LW, Alpermann T, Sun QY, Lin DC, Chien W, Madan V, Liu LZ, Tan KT, Sampath A, Venkatesan S, Inokuchi K, Wakita S, Yamaguchi H, Chng WJ, Kham SK, Yeoh AE, Sanada M, Schiller J, Kreuzer KA, Kornblau SM, Kantarjian HM, Haferlach T, Lill M, Kuo MC, Shih LY, Blau IW, Blau O, Yang H, Ogawa S, Koeffler HP. Profiling of somatic mutations in acute myeloid leukemia with FLT3-ITD at diagnosis and relapse. Blood. 2015;126:2491–501.
Smith CC, Wang Q, Chin CS, Salerno S, Damon LE, Levis MJ, Perl AE, Travers KJ, Wang S, Hunt JP, Zarrinkar PP, Schadt EE, Kasarskis A, Kuriyan J, Shah NP. Validation of ITD mutations in FLT3 as a therapeutic target in human acute myeloid leukaemia. Nature. 2012;485:260–3.
Thiede C, Steudel C, Mohr B, Schaich M, Schakel U, Platzbecker U, Wermke M, Bornhauser M, Ritter M, Neubauer A, Ehninger G, Illmer T. Analysis of FLT3-activating mutations in 979 patients with acute myelogenous leukemia: association with FAB subtypes and identification of subgroups with poor prognosis. Blood. 2002;99:4326–35.
Scholl S, Salzmann S, Kaufmann AM, Hoffken K. Flt3-ITD mutations can generate leukaemia specific neoepitopes: potential role for immunotherapeutic approaches. Leuk Lymphoma. 2006;47:307–12.
Graf C, Heidel F, Tenzer S, Radsak MP, Solem FK, Britten CM, Huber C, Fischer T, Wolfel T. A neoepitope generated by an FLT3 internal tandem duplication (FLT3-ITD) is recognized by leukemia-reactive autologous CD8+ T cells. Blood. 2007;109:2985–8.
Kang MR, Kim MS, Oh JE, Kim YR, Song SY, Seo SI, Lee JY, Yoo NJ, Lee SH. Mutational analysis of IDH1 codon 132 in glioblastomas and other common cancers. Int J Cancer. 2009;125:353–5.
Sasaki M, Knobbe CB, Munger JC, Lind EF, Brenner D, Brustle A, Harris IS, Holmes R, Wakeham A, Haight J, You-Ten A, Li WY, Schalm S, Su SM, Virtanen C, Reifenberger G, Ohashi PS, Barber DL, Figueroa ME, Melnick A, Zuniga-Pflucker JC, Mak TW. IDH1(R132H) mutation increases murine haematopoietic progenitors and alters epigenetics. Nature. 2012;488:656–9.
Figueroa ME, Abdel-Wahab O, Lu C, Ward PS, Patel J, Shih A, Li Y, Bhagwat N, Vasanthakumar A, Fernandez HF, Tallman MS, Sun Z, Wolniak K, Peeters JK, Liu W, Choe SE, Fantin VR, Paietta E, Lowenberg B, Licht JD, Godley LA, Delwel R, Valk PJ, Thompson CB, Levine RL, Melnick A. Leukemic IDH1 and IDH2 mutations result in a hypermethylation phenotype, disrupt TET2 function, and impair hematopoietic differentiation. Cancer Cell. 2010;18:553–67.
Kavianpour M, Ahmadzadeh A, Shahrabi S, Saki N. Significance of oncogenes and tumor suppressor genes in AML prognosis. Tumour Biol. 2016;37:10041–52.
Ok CY, Loghavi S, Sui D, Wei P, Kanagal-Shamanna R, Yin CC, Zuo Z, Routbort MJ, Tang G, Tang Z, Jorgensen JL, Luthra R, Ravandi F, Kantarjian HM, DiNardo CD, Medeiros LJ, Wang SA, Patel KP. Persistent IDH1/2 mutations in remission can predict relapse in patients with acute myeloid leukemia. Haematologica. 2019;104:305–11.
Schumacher T, Bunse L, Pusch S, Sahm F, Wiestler B, Quandt J, Menn O, Osswald M, Oezen I, Ott M, Keil M, Balss J, Rauschenbach K, Grabowska AK, Vogler I, Diekmann J, Trautwein N, Eichmuller SB, Okun J, Stevanovic S, Riemer AB, Sahin U, Friese MA, Beckhove P, von Deimling A, Wick W, Platten M. A vaccine targeting mutant IDH1 induces antitumour immunity. Nature. 2014;512:324–7.
Platten M, Bunse L, Wick A, Bunse T, Le Cornet L, Harting I, Sahm F, Sanghvi K, Tan CL, Poschke I, Green E, Justesen S, Behrens GA, Breckwoldt MO, Freitag A, Rother LM, Schmitt A, Schnell O, Hense J, Misch M, Krex D, Stevanovic S, Tabatabai G, Steinbach JP, Bendszus M, von Deimling A, Schmitt M, Wick W. A vaccine targeting mutant IDH1 in newly diagnosed glioma. Nature. 2021;592:463–8.
Pylayeva-Gupta Y, Grabocka E, Bar-Sagi D. RAS oncogenes: weaving a tumorigenic web. Nat Rev Cancer. 2011;11:761–74.
Bamford S, Dawson E, Forbes S, Clements J, Pettett R, Dogan A, Flanagan A, Teague J, Futreal PA, Stratton MR, Wooster R. The COSMIC (Catalogue of Somatic Mutations in Cancer) database and website. Br J Cancer. 2004;91:355–8.
Stolze B, Reinhart S, Bulllinger L, Frohling S, Scholl C. Comparative analysis of KRAS codon 12, 13, 18, 61, and 117 mutations using human MCF10A isogenic cell lines. Sci Rep. 2015;5:8535.
Chang YI, You X, Kong G, Ranheim EA, Wang J, Du J, Liu Y, Zhou Y, Ryu MJ, Zhang J. Loss of Dnmt3a and endogenous Kras(G12D/+) cooperate to regulate hematopoietic stem and progenitor cell functions in leukemogenesis. Leukemia. 2015;29:1847–56.
McMahon CM, Ferng T, Canaani J, Wang ES, Morrissette J, Eastburn DJ, Pellegrino M, Durruthy-Durruthy R, Watt CD, Asthana S, Lasater EA, DeFilippis R, Peretz C, McGary L, Deihimi S, Logan AC, Luger SM, Shah NP, Carroll M, Smith CC, Perl AE. Clonal Selection with RAS Pathway Activation Mediates Secondary Clinical Resistance to Selective FLT3 Inhibition in Acute Myeloid Leukemia. Cancer Discov. 2019;9:1050–63.
Zhang Y, Ma JA, Zhang HX, Jiang YN, Luo WH. Cancer vaccines: Targeting KRAS-driven cancers. Expert Rev Vaccines. 2020;19:163–73.
Tran E, Ahmadzadeh M, Lu YC, Gros A, Turcotte S, Robbins PF, Gartner JJ, Zheng Z, Li YF, Ray S, Wunderlich JR, Somerville RP, Rosenberg SA. Immunogenicity of somatic mutations in human gastrointestinal cancers. Science. 2015;350:1387–90.
Sim M, Lu J, Spencer M, Hopkins F, Tran E, Rosenberg SA, Long EO, Sun PD. High-affinity oligoclonal TCRs define effective adoptive T cell therapy targeting mutant KRAS-G12D. Proc Natl Acad Sci U S A. 2020;117:12826–35.
Cafri G, Gartner JJ, Zaks T, Hopson K, Levin N, Paria BC, Parkhurst MR, Yossef R, Lowery FJ, Jafferji MS, Prickett TD, Goff SL, McGowan CT, Seitter S, Shindorf ML, Parikh A, Chatani PD, Robbins PF, Rosenberg SA. mRNA vaccine-induced neoantigen-specific T cell immunity in patients with gastrointestinal cancer. J Clin Invest. 2020;130:5976–88.
Leidner R, Sanjuan SN, Huang H, Sprott D, Zheng C, Shih YP, Leung A, Payne R, Sutcliffe K, Cramer J, Rosenberg SA, Fox BA, Urba WJ, Tran E. Neoantigen T-cell receptor gene therapy in pancreatic cancer. N Engl J Med. 2022;386:2112–9.
Rucker FG, Schlenk RF, Bullinger L, Kayser S, Teleanu V, Kett H, Habdank M, Kugler CM, Holzmann K, Gaidzik VI, Paschka P, Held G, von Lilienfeld-Toal M, Lubbert M, Frohling S, Zenz T, Krauter J, Schlegelberger B, Ganser A, Lichter P, Dohner K, Dohner H. TP53 alterations in acute myeloid leukemia with complex karyotype correlate with specific copy number alterations, monosomal karyotype, and dismal outcome. Blood. 2012;119:2114–21.
Hou HA, Chou WC, Kuo YY, Liu CY, Lin LI, Tseng MH, Chiang YC, Liu MC, Liu CW, Tang JL, Yao M, Li CC, Huang SY, Ko BS, Hsu SC, Chen CY, Lin CT, Wu SJ, Tsay W, Chen YC, Tien HF. TP53 mutations in de novo acute myeloid leukemia patients: longitudinal follow-ups show the mutation is stable during disease evolution. Blood Cancer J. 2015;5: e331.
Leroy B, Anderson M, Soussi T. TP53 mutations in human cancer: database reassessment and prospects for the next decade. Hum Mutat. 2014;35:672–88.
Malekzadeh P, Pasetto A, Robbins PF, Parkhurst MR, Paria BC, Jia L, Gartner JJ, Hill V, Yu Z, Restifo NP, Sachs A, Tran E, Lo W, Somerville RP, Rosenberg SA, Deniger DC. Neoantigen screening identifies broad TP53 mutant immunogenicity in patients with epithelial cancers. J Clin Invest. 2019;129:1109–14.
Deniger DC, Pasetto A, Robbins PF, Gartner JJ, Prickett TD, Paria BC, Malekzadeh P, Jia L, Yossef R, Langhan MM, Wunderlich JR, Danforth DN, Somerville R, Rosenberg SA. T-cell Responses to TP53 “Hotspot” mutations and unique neoantigens expressed by human ovarian cancers. Clin Cancer Res. 2018;24:5562–73.
Wang Z, Ran L, Chen C, Shi R, Dong Y, Li Y, Zhou X, Qi Y, Zhu P, Gao Y, Wu Y. Identification of HLA-A2-restricted mutant epitopes from neoantigens of esophageal squamous cell carcinoma. Vaccines (Basel). 2021;9:45.
Malekzadeh P, Yossef R, Cafri G, Paria BC, Lowery FJ, Jafferji M, Good ML, Sachs A, Copeland AR, Kim SP, Kivitz S, Parkhurst MR, Robbins PF, Ray S, Xi L, Raffeld M, Yu Z, Restifo NP, Somerville R, Rosenberg SA, Deniger DC. Antigen experienced T cells from peripheral blood recognize p53 Neoantigens. Clin Cancer Res. 2020;26:1267–76.
Wu D, Liu Y, Li X, Liu Y, Yang Q, Liu Y, Wu J, Tian C, Zeng Y, Zhao Z, Xiao Y, Gu F, Zhang K, Hu Y, Liu L. Identification of Clonal Neoantigens derived from driver mutations in an EGFR-mutated lung cancer patient benefitting from anti-PD-1. FRONT IMMUNOL. 2020;11:1366.
Lo W, Parkhurst M, Robbins PF, Tran E, Lu YC, Jia L, Gartner JJ, Pasetto A, Deniger D, Malekzadeh P, Shelton TE, Prickett T, Ray S, Kivitz S, Paria BC, Kriley I, Schrump DS, Rosenberg SA. Immunologic recognition of a shared p53 mutated neoantigen in a patient with metastatic colorectal cancer. Cancer Immunol Res. 2019;7:534–43.
Hsiue EH, Wright KM, Douglass J, Hwang MS, Mog BJ, Pearlman AH, Paul S, DiNapoli SR, Konig MF, Wang Q, Schaefer A, Miller MS, Skora AD, Azurmendi PA, Murphy MB, Liu Q, Watson E, Li Y, Pardoll DM, Bettegowda C, Papadopoulos N, Kinzler KW, Vogelstein B, Gabelli SB, Zhou S. Targeting a neoantigen derived from a common TP53 mutation. Science. 2021;371:45.
Alsafadi S, Houy A, Battistella A, Popova T, Wassef M, Henry E, Tirode F, Constantinou A, Piperno-Neumann S, Roman-Roman S, Dutertre M, Stern MH. Cancer-associated SF3B1 mutations affect alternative splicing by promoting alternative branchpoint usage. Nat Commun. 2016;7:10615.
Je EM, Yoo NJ, Kim YJ, Kim MS, Lee SH. Mutational analysis of splicing machinery genes SF3B1, U2AF1 and SRSF2 in myelodysplasia and other common tumors. Int J Cancer. 2013;133:260–5.
Hou HA, Liu CY, Kuo YY, Chou WC, Tsai CH, Lin CC, Lin LI, Tseng MH, Chiang YC, Liu MC, Liu CW, Tang JL, Yao M, Li CC, Huang SY, Ko BS, Hsu SC, Chen CY, Lin CT, Wu SJ, Tsay W, Tien HF. Splicing factor mutations predict poor prognosis in patients with de novo acute myeloid leukemia. Oncotarget. 2016;7:9084–101.
Bigot J, Lalanne AI, Lucibello F, Gueguen P, Houy A, Dayot S, Ganier O, Gilet J, Tosello J, Nemati F, Pierron G, Waterfall JJ, Barnhill R, Gardrat S, Piperno-Neumann S, Popova T, Masson V, Loew D, Mariani P, Cassoux N, Amigorena S, Rodrigues M, Alsafadi S, Stern MH, Lantz O. Splicing patterns in SF3B1-mutated uveal melanoma generate shared immunogenic tumor-specific neoepitopes. Cancer Discov. 2021;11:1938–51.
Liepe J, Marino F, Sidney J, Jeko A, Bunting DE, Sette A, Kloetzel PM, Stumpf MP, Heck AJ, Mishto M. A large fraction of HLA class I ligands are proteasome-generated spliced peptides. Science. 2016;354:354–8.
Makita M, Azuma T, Hamaguchi H, Niiya H, Kojima K, Fujita S, Tanimoto M, Harada M, Yasukawa M. Leukemia-associated fusion proteins, dek-can and bcr-abl, represent immunogenic HLA-DR-restricted epitopes recognized by fusion peptide-specific CD4+ T lymphocytes. Leukemia. 2002;16:2400–7.
Wang QJ, Yu Z, Griffith K, Hanada K, Restifo NP, Yang JC. Identification of T-cell Receptors Targeting KRAS-Mutated Human Tumors. Cancer Immunol Res. 2016;4:204–14.
Assadipour Y, Zacharakis N, Crystal JS, Prickett TD, Gartner JJ, Somerville R, Xu H, Black MA, Jia L, Chinnasamy H, Kriley I, Lu L, Wunderlich JR, Zheng Z, Lu YC, Robbins PF, Rosenberg SA, Goff SL, Feldman SA. Characterization of an immunogenic mutation in a patient with metastatic triple-negative breast cancer. Clin Cancer Res. 2017;23:4347–53.
Holmstrom MO, Martinenaite E, Ahmad SM, Met O, Friese C, Kjaer L, Riley CH, Thor SP, Svane IM, Hasselbalch HC, Andersen MH. The calreticulin (CALR) exon 9 mutations are promising targets for cancer immune therapy. Leukemia. 2018;32:429–37.
Cohen CJ, Gartner JJ, Horovitz-Fried M, Shamalov K, Trebska-McGowan K, Bliskovsky VV, Parkhurst MR, Ankri C, Prickett TD, Crystal JS, Li YF, El-Gamil M, Rosenberg SA, Robbins PF. Isolation of neoantigen-specific T cells from tumor and peripheral lymphocytes. J Clin Invest. 2015;125:3981–91.
Wong Y, Joshi K, Khetrapal P, Ismail M, Reading JL, Sunderland MW, Georgiou A, Furness A, Ben AA, Ghorani E, Oakes T, Uddin I, Tan WS, Feber A, McGovern U, Swanton C, Freeman A, Marafioti T, Briggs TP, Kelly JD, Powles T, Peggs KS, Chain BM, Linch MD, Quezada SA. Urine-derived lymphocytes as a non-invasive measure of the bladder tumor immune microenvironment. J Exp Med. 2018;215:2748–59.
Jang M, Yew PY, Hasegawa K, Ikeda Y, Fujiwara K, Fleming GF, Nakamura Y, Park JH. Characterization of T cell repertoire of blood, tumor, and ascites in ovarian cancer patients using next generation sequencing. Oncoimmunology. 2015;4: e1030561.
Holmstrom MO, Hasselbalch HC, Andersen MH. Neo-antigen specific memory T-cell responses in healthy individuals. Oncoimmunology. 2019;8:1599640.
Inozume T, Hanada K, Wang QJ, Ahmadzadeh M, Wunderlich JR, Rosenberg SA, Yang JC. Selection of CD8+PD-1+ lymphocytes in fresh human melanomas enriches for tumor-reactive T cells. J Immunother. 2010;33:956–64.
Gros A, Parkhurst MR, Tran E, Pasetto A, Robbins PF, Ilyas S, Prickett TD, Gartner JJ, Crystal JS, Roberts IM, Trebska-McGowan K, Wunderlich JR, Yang JC, Rosenberg SA. Prospective identification of neoantigen-specific lymphocytes in the peripheral blood of melanoma patients. Nat Med. 2016;22:433–8.
Fernandez-Poma SM, Salas-Benito D, Lozano T, Casares N, Riezu-Boj JI, Mancheno U, Elizalde E, Alignani D, Zubeldia N, Otano I, Conde E, Sarobe P, Lasarte JJ, Hervas-Stubbs S. Expansion of tumor-infiltrating CD8(+) T cells expressing PD-1 improves the efficacy of adoptive T-cell therapy. Cancer Res. 2017;77:3672–84.
Gros A, Robbins PF, Yao X, Li YF, Turcotte S, Tran E, Wunderlich JR, Mixon A, Farid S, Dudley ME, Hanada K, Almeida JR, Darko S, Douek DC, Yang JC, Rosenberg SA. PD-1 identifies the patient-specific CD8(+) tumor-reactive repertoire infiltrating human tumors. J Clin Invest. 2014;124:2246–59.
Wang C, Lin GH, McPherson AJ, Watts TH. Immune regulation by 4–1BB and 4–1BBL: complexities and challenges. Immunol Rev. 2009;229:192–215.
Croft M. The role of TNF superfamily members in T-cell function and diseases. Nat Rev Immunol. 2009;9:271–85.
Wolfl M, Kuball J, Ho WY, Nguyen H, Manley TJ, Bleakley M, Greenberg PD. Activation-induced expression of CD137 permits detection, isolation, and expansion of the full repertoire of CD8+ T cells responding to antigen without requiring knowledge of epitope specificities. Blood. 2007;110:201–10.
Parkhurst M, Gros A, Pasetto A, Prickett T, Crystal JS, Robbins P, Rosenberg SA. Isolation of T-cell receptors specifically reactive with mutated tumor-associated antigens from tumor-infiltrating lymphocytes based on CD137 expression. Clin Cancer Res. 2017;23:2491–505.
Cafri G, Yossef R, Pasetto A, Deniger DC, Lu YC, Parkhurst M, Gartner JJ, Jia L, Ray S, Ngo LT, Jafferji M, Sachs A, Prickett T, Robbins PF, Rosenberg SA. Memory T cells targeting oncogenic mutations detected in peripheral blood of epithelial cancer patients. Nat Commun. 2019;10:449.
Canale FP, Ramello MC, Nunez N, Araujo FC, Bossio SN, Gorosito SM, Tosello BJ, Del CA, Ledesma M, Sedlik C, Piaggio E, Gruppi A, Acosta RE, Montes CL. CD39 expression defines cell exhaustion in tumor-infiltrating CD8(+) T Cells. Cancer Res. 2018;78:115–28.
Simoni Y, Becht E, Fehlings M, Loh CY, Koo SL, Teng K, Yeong J, Nahar R, Zhang T, Kared H, Duan K, Ang N, Poidinger M, Lee YY, Larbi A, Khng AJ, Tan E, Fu C, Mathew R, Teo M, Lim WT, Toh CK, Ong BH, Koh T, Hillmer AM, Takano A, Lim T, Tan EH, Zhai W, Tan D, Tan IB, Newell EW. Bystander CD8(+) T cells are abundant and phenotypically distinct in human tumour infiltrates. Nature. 2018;557:575–9.
Duhen T, Duhen R, Montler R, Moses J, Moudgil T, de Miranda NF, Goodall CP, Blair TC, Fox BA, McDermott JE, Chang SC, Grunkemeier G, Leidner R, Bell RB, Weinberg AD. Co-expression of CD39 and CD103 identifies tumor-reactive CD8 T cells in human solid tumors. Nat Commun. 2018;9:2724.
Krishna S, Lowery FJ, Copeland AR, Bahadiroglu E, Mukherjee R, Jia L, Anibal JT, Sachs A, Adebola SO, Gurusamy D, Yu Z, Hill V, Gartner JJ, Li YF, Parkhurst M, Paria B, Kvistborg P, Kelly MC, Goff SL, Altan-Bonnet G, Robbins PF, Rosenberg SA. Stem-like CD8 T cells mediate response of adoptive cell immunotherapy against human cancer. Science. 2020;370:1328–34.
Lu YC, Zheng Z, Robbins PF, Tran E, Prickett TD, Gartner JJ, Li YF, Ray S, Franco Z, Bliskovsky V, Fitzgerald PC, Rosenberg SA. An efficient single-cell RNA-Seq approach to identify neoantigen-specific T cell receptors. Mol Ther. 2018;26:379–89.
Lu YC, Zheng Z, Lowery FJ, Gartner JJ, Prickett TD, Robbins PF, Rosenberg SA. Direct identification of neoantigen-specific TCRs from tumor specimens by high-throughput single-cell sequencing. J Immunother Cancer. 2021;9:456.
van der Leun AM, Thommen DS, Schumacher TN. CD8(+) T cell states in human cancer: insights from single-cell analysis. Nat Rev Cancer. 2020;20:218–32.
Scheper W, Kelderman S, Fanchi LF, Linnemann C, Bendle G, de Rooij M, Hirt C, Mezzadra R, Slagter M, Dijkstra K, Kluin R, Snaebjornsson P, Milne K, Nelson BH, Zijlmans H, Kenter G, Voest EE, Haanen J, Schumacher TN. Low and variable tumor reactivity of the intratumoral TCR repertoire in human cancers. Nat Med. 2019;25:89–94.
Matsuda T, Leisegang M, Park JH, Ren L, Kato T, Ikeda Y, Harada M, Kiyotani K, Lengyel E, Fleming GF, Nakamura Y. Induction of neoantigen-specific cytotoxic T Cells and Construction of T-cell receptor-engineered T Cells for Ovarian Cancer. Clin Cancer Res. 2018;24:5357–67.
Zhang J, Wang L. The emerging world of TCR-T cell trials against cancer: a systematic review. Technol Cancer Res Treat. 2019;18:1078098716.
Inoue K, Ogawa H, Sonoda Y, Kimura T, Sakabe H, Oka Y, Miyake S, Tamaki H, Oji Y, Yamagami T, Tatekawa T, Soma T, Kishimoto T, Sugiyama H. Aberrant overexpression of the Wilms tumor gene (WT1) in human leukemia. Blood. 1997;89:1405–12.
Chapuis AG, Ragnarsson GB, Nguyen HN, Chaney CN, Pufnock JS, Schmitt TM, Duerkopp N, Roberts IM, Pogosov GL, Ho WY, Ochsenreither S, Wolfl M, Bar M, Radich JP, Yee C, Greenberg PD. Transferred WT1-reactive CD8+ T cells can mediate antileukemic activity and persist in post-transplant patients. SCI TRANSL MED. 2013;5:127r–74r.
Ohminami H, Yasukawa M, Fujita S. HLA class I-restricted lysis of leukemia cells by a CD8(+) cytotoxic T-lymphocyte clone specific for WT1 peptide. Blood. 2000;95:286–93.
Ochi T, Fujiwara H, Okamoto S, An J, Nagai K, Shirakata T, Mineno J, Kuzushima K, Shiku H, Yasukawa M. Novel adoptive T-cell immunotherapy using a WT1-specific TCR vector encoding silencers for endogenous TCRs shows marked antileukemia reactivity and safety. Blood. 2011;118:1495–503.
Dossa RG, Cunningham T, Sommermeyer D, Medina-Rodriguez I, Biernacki MA, Foster K, Bleakley M. Development of T-cell immunotherapy for hematopoietic stem cell transplantation recipients at risk of leukemia relapse. Blood. 2018;131:108–20.
Marvin-Peek J, Savani BN, Olalekan OO, Dholaria B. Challenges and Advances in Chimeric Antigen Receptor Therapy for Acute Myeloid Leukemia. Cancers (Basel). 2022;14:34.
He Q, Jiang X, Zhou X, Weng J. Targeting cancers through TCR-peptide/MHC interactions. J Hematol Oncol. 2019;12:139.
Ye B, Stary CM, Gao Q, Wang Q, Zeng Z, Jian Z, Gu L, Xiong X. Genetically modified T-cell-based adoptive immunotherapy in hematological malignancies. J Immunol Res. 2017;2017:5210459.
Vishwasrao P, Li G, Boucher JC, Smith DL, Hui SK. Emerging CAR T Cell Strategies for the Treatment of AML. Cancers (Basel). 2022;14:98.
Petrov JC, Wada M, Pinz KG, Yan LE, Chen KH, Shuai X, Liu H, Chen X, Leung LH, Salman H, Hagag N, Liu F, Jiang X, Ma Y. Compound CAR T-cells as a double-pronged approach for treating acute myeloid leukemia. Leukemia. 2018;32:1317–26.
Kenderian SS, Ruella M, Shestova O, Klichinsky M, Aikawa V, Morrissette JJ, Scholler J, Song D, Porter DL, Carroll M, June CH, Gill S. CD33-specific chimeric antigen receptor T cells exhibit potent preclinical activity against human acute myeloid leukemia. Leukemia. 2015;29:1637–47.
Gill S, Tasian SK, Ruella M, Shestova O, Li Y, Porter DL, Carroll M, Danet-Desnoyers G, Scholler J, Grupp SA, June CH, Kalos M. Preclinical targeting of human acute myeloid leukemia and myeloablation using chimeric antigen receptor-modified T cells. Blood. 2014;123:2343–54.
Tasian SK, Kenderian SS, Shen F, Ruella M, Shestova O, Kozlowski M, Li Y, Schrank-Hacker A, Morrissette J, Carroll M, June CH, Grupp SA, Gill S. Optimized depletion of chimeric antigen receptor T cells in murine xenograft models of human acute myeloid leukemia. Blood. 2017;129:2395–407.
Baumeister SH, Murad J, Werner L, Daley H, Trebeden-Negre H, Gicobi JK, Schmucker A, Reder J, Sentman CL, Gilham DE, Lehmann FF, Galinsky I, DiPietro H, Cummings K, Munshi NC, Stone RM, Neuberg DS, Soiffer R, Dranoff G, Ritz J, Nikiforow S. Phase I Trial of Autologous CAR T Cells Targeting NKG2D Ligands in Patients with AML/MDS and Multiple Myeloma. Cancer Immunol Res. 2019;7:100–12.
Sauer T, Parikh K, Sharma S, Omer B, Sedloev D, Chen Q, Angenendt L, Schliemann C, Schmitt M, Muller-Tidow C, Gottschalk S, Rooney CM. CD70-specific CAR T cells have potent activity against acute myeloid leukemia without HSC toxicity. Blood. 2021;138:318–30.
Zhao L, Cao YJ. Engineered T Cell Therapy for Cancer in the Clinic. Front Immunol. 2019;10:2250.
Cohen CJ, Zhao Y, Zheng Z, Rosenberg SA, Morgan RA. Enhanced antitumor activity of murine-human hybrid T-cell receptor (TCR) in human lymphocytes is associated with improved pairing and TCR/CD3 stability. Cancer Res. 2006;66:8878–86.
Funding
This study is supported by the National Natural Science Foundation of China (81890994), the National Key R&D Program of China (2019YFA0905902), the Guangdong Basic and Applied Basic Research Foundation (2019A1515010299), the Guangzhou Science and Technology Plan Project (202102020727), the Innovative Research Team of High-level Local Universities in Shanghai, and the Dean' fund of Zhujiang Hospital, Southern Medical University (yzjj2019qn05).
Author information
Authors and Affiliations
Contributions
WZ and KW conceived, designed, and supervised the study. WZ and JY searched and reviewed the studies. WZ drew figures and tables, JY and YL helped with figures. WZ wrote the manuscript and KW revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
All other authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhou, W., Yu, J., Li, Y. et al. Neoantigen-specific TCR-T cell-based immunotherapy for acute myeloid leukemia. Exp Hematol Oncol 11, 100 (2022). https://doi.org/10.1186/s40164-022-00353-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40164-022-00353-3