
Abstract
Cullin 4B (CUL4B) is a scaffold of the Cullin4B-Ring E3 ligase complex (CRL4B) that plays an important role in proteolysis and is implicated in tumorigenesis. Aberrant expression of CUL4B has been reported in various types of human diseases. Recently, studies have shown that CUL4B was overexpressed in a multitude of solid neoplasms and affect the expression of several tumor suppressor genes. In this review, we aim to summarize the biological function of CUL4B in order to better understand its pathogenesis in human cancers.
Similar content being viewed by others

Background
Cullins are evolutionarily highly conserved protein molecule family in most mammals. There are eight subtypes cullins in human genome, including cullin1, cullin2, cullin3, cullin4A, cullin4B, cullin5, cullin7 and PARC, also known cullin9 [1]. The cullin protein functions as a scaffold in the largest class of really interesting new gene (RING) E3 ligases, by binding to RING box proteins through a highly conserved homology domain. Cullins play a key role in selective degradation of various proteins which are involved in versatile cell biology behaviors, including cell cycle and signaling regulation. More recently, studies have focused on their connections with human diseases, including cancer [2,3,4].
There are two members in cullin4 (CUL4) family, CUL4A and CUL4B. CUL4B is highly sequence homology with CUL4A, sharing 83% sequence identity [5]. Scientists used to think that CUL4A and CUL4B involved several cell life activities in an almost similar way. Nevertheless, latest publications indicate that there are big differences in functions and specific mechanisms between CUL4A and CUL4B. Difference from CUL4A and other cullins that it carries their nuclear localization signal (NLS) in their C termini, whereas NLS in CUL4B is located in its N terminus [5]. CUL4A and CUL4B can interact with DDB21, a substrate adaptor, and they may target the same substrates and function redundantly in some cellular functions, such as genome integrity maintenance, DNA replication and cell cycle regulation [6,7,8]. However, CUL4B has been demonstrated to target substrates, such as WD repeat containing protein5 (WDR5) and peroxiredoxin III (PrxIII), that are not targeted by CUL4A [9,10,11]. CUL4B knockout mice were embryonic lethal [12, 13]. In contrast, except for failure in spermatogenesis, CUL4B knockout mice have no remarkable abnormalities [14, 15]. CUL4A is highly expressed in the pancreas, testes and in T cells [16], while CUL4B is most highly expressed in pancreatic tissue, endocrine glands, the cerebellum, the lower GI tract, bone marrow and the testes [17].
Recently, the function of CUL4B in solid tumors has been gradually uncovered and attracts a lot of interests. Plenty of studies have demonstrated that the expression of CUL4B is abnormal in a wide variety of diseases and physiological process (Table 1) [3, 18,19,20,21]. Through upregulating or decreasing the expression of CUL4B in cancer cells, these studies have demonstrated clearly that CUL4B acts a pivotal part in cell proliferation, DNA damage and repair, cell cycle progression, metastasis, invasion, DNA methylation and histone acetylation modification, as well as signaling pathways. Furthermore, CUL4B may present potential diagnostic value for human diseases. This review intends to summarize recent insights into CUL4B functions (Fig. 1). In addition, we revealed the findings demonstrating CUL4B’s association with oncogenesis and its important function as bio-marker for tumor diagnosis.

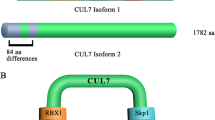
The schematic representation of CUL4B and associated factors
CUL4B expression in central nervous system
In 2007, Tarpey et al. [22] and Zou et al. [23] noticed that mutations in CUL4B caused an x-linked mental retardation syndrome (XLMR). Individuals with XLMR show obvious clinical features, including growth retardation, mental retardation, relative macrocephaly, truncal obesity and hypogonadism [22, 23]. XLMR-linked CUL4B mutations induced the accumulation of WDR5 and the activation of neuronal genes which could promote neurite extension [24]. Moreover, one of the CUL4B specific substrate, PrxIII, has been reported to affect neural development via regulating the level of reactive oxygen species (ROS) [10]. When CUL4B was silenced, the accumulation of PrxIII inhibited the generation of ROS and enhanced resistance to hypoxia and H2O2-induced apoptosis, and then interfered with cell proliferation and normal cellular functions [10].
Vulto-van Silfhout et al. [25] also discovered that CUL4B was necessary for brain development. They provided first evidence that there was a firm association between CUL4B variants and cerebral malformations. The interaction between CUL4B and WDR62 might contribute to the development of cerebral malformations in patients with CUL4B variants [25]. Furthermore, observation of abnormal neuronal organization in the hippocampus in CUL4B knock-out mice testified that CUL4B played an important role in brain development [7]. In the CUL4B knock-out mice, exons 4-5 of CUL4B in the X chromosome were excised by Sox2-Cre [7]. CUL4B knock-out mice presented hippocampus-related spatial learning and memory deficiencies, while presenting normal appearances [7].
Dong et al. [26] demonstrated that CUL4B might function as an oncogene in malignant glioma tumorigenesis. This paper showed that knockdown CUL4B significantly decreased cell proliferation via inducing G1 phase cell cycle arrest and inhibited the tumor growth in xenografts in nude mice, which revealed that CUL4B knockdown might alleviated glioma tumorigenesis [26].
CUL4B expression in human cancers
Immunohistochemical staining in esophageal, lung, gastric, colon, pancreatic, cervical, renal, liver, and bladder tumor and matched tissues, showed a significant overexpression of CUL4B in tumor tissue [3, 18,19,20]. Higher expression of CUL4B in esophageal cancerous tissues has a closely association with lower histological grades [3]. Yang et al. [20] reported that CUL4B was frequently increased in 64 human cervical carcinoma samples compared to 30 adjacent non-tumor cervical tissues. They also found a positive correlation between CUL4B expression and cervical carcinoma histological grades. Meanwhile, CUL4B mRNA level was also higher in 12 of 15 selected paired samples of each grade cancers [20].
High expression of CUL4B expression was reported to be related to tumor histological differentiation, invasion, distant metastasis, tumor size, lymph node metastasis, overall and recurrence rate, as well as disease-free survival in breast, colorectal, cervical, oesophageal and lung cancers [3, 18, 20, 27]. Study has shown that knockdown CUL4B could suppress the epithelial–mesenchymal transition (EMT) progress [28]. However, the mechanism is still unclear. Based on the above facts, CUL4B is generally overexpressed in the cancer tissues and is associated with unfavorable prognosis, which may be a novel tumor marker for tumor diagnosis and prognosis. Further studies are needed to examine the relationship between CUL4B and other non-solid malignancies.
CUL4B and DNA damage and repair
It has been identified that the CRLs formed by several members of cullin family are involved in DNA damage and repair procedures. Hereinto, CUL4A participated in nucleotide excision repair by CUL4A-DDBl-CSA complex or CUL4A-DDBl-DDB2 complex [29]. Homology of CUL4B and CUL4A is high and only one kind of CUL4 is expressed in primary eukaryotes [5]. Compared with CUL4A, CUL4B possess specific amino acid end consisting of 149 amino acids [30]. Study has certified that CUL4 was engaged in DNA homogenous recombination repair in a way of DDBl-CUL4CDT2 complex [31]. Guerrero-Santoro et al. [32] manifested that both DDBl-Cul4ADDB2 and DDBl-CUL4BDDB2 recognized UV-damaged DNA in the same DDB2-dependent manner. While, the latter is more efficient than the former in monoubiquitinating histone H2A. CUL4B promotes the transfer of DDB1 into the nucleus independently of DDB2 after UV irradiation [32]. CUL4B can regulate the DNA damage induced by UV irradiation.
Further, DNA double strands break increased after treatment of camptothecin (CPT) in lymphoblastic cells conveying CUL4B mutation [31]. Cells from patients with mutant CUL4B were more sensitive to camptothecin (CPT), which indicated that CUL4B may impair CPT-induced topoisomerase I (Topo I) degradation and ubiquitination [31]. Consistent with this, these cells exhibited increased levels of CPT-induced DNA breaks. It has been reported that downregulation of HUWE1 in response to DNA damage is accompanied by the activation of CUL4B and CRL4B was required for proteasomal degradation of HUWE1 [33]. These evidence manifests that CUL4B is associated with DNA damage repair.
CUL4B and cell cycle progression
Cell cycle is important to maintain tumor or normal cells growth. Lots of studies have shown that deregulation of cell cycle progression could make cells eventually develop as tumor. Cell cycle progression is strictly controlled by multiple mechanisms to ensure its strict and orderly progress. Cyclin-dependent kinases (CDKs), CDK inhibitors (CKIs) and Cyclins are main regulators in cell cycle [34, 35]. Accordingly, maintaining the balance of the above proteins is very necessary.
Cyclin E is important in cell cycle which can bind to CDK2 to regulate G1-S transition and guarantee DNA replication accuracy [36]. Cyclin E alteration has been reported in many cancers and is associated with pathogenesis of malignancies [37,38,39]. Several papers have shown that cyclin E is regulated by ubiquitin-mediated proteolysis system [40,41,42]. Knockdown CUL4B induces significant accumulation of cyclin E [5, 42] and a prolonged S phase, resulting in an inhibition of cell proliferation [5]. In conclusion, these studies are suggestive of CUL4B probably targeting cyclin E for degradation to inhibit cell proliferation [5].
P21, one kind of CKIs, can regulate cell cycle by inhibiting the kinase activity of CDK2. Many studies have manifested that p21 accumulates during G1 phase but degraded during S phase [43]. CUL4B negatively regulates the function of p21 by transcriptional repression (repressing the transcription of CDKN1A which encodes p21), but not by proteolysis [44]. CUL4B promotes cell proliferation at least partially through repressing the transcription of Cdkn1a which encodes p21. Silencing of CUL4B expression caused accumulation of p21 protein in extra-embryonic cells, HEK293 and HeLa cells [13, 44].
CUL4B can also upregulate DNA replication by positively regulating CDC6 [8]. Study proves that CDC6 is essential for the formation of pre-replication complexes (pre-RC) [31, 32]. CDK2 is responsible for the stabilization and phosphorylation of CDC6. CULB upregulates the CDK2 expression by negatively transcription in miR-372/373, resulting in the positive regulation of CDC6.
CUL4B in post-transcriptional modification
Epigenetics is the study of gene post-transcriptional modification. Epigenetic modifications are important for the development and progression of tumors [45]. As research continues, epigenetics has evoked widespread attention to their functions in the regulation of gene expression, including promoter–enhancer interactions, histone modifications, DNA methylation, and noncoding RNA-mediated regulation. The coordinated operation of aspects above is involved in cell cycle regulation, proliferation, apoptosis and so on, and relatives to the ultimate responses in human health and disease [46]. Recently, DNA methylation and histone modifications have been reported in various cancers, and some of which s are found to be associated with poor prognosis.
DNA methylation is one of the most common epigenetic events in eukaryotic cell. It has been revealed that DNA methylation plays a key regulatory role in normal development. Studies have revealed that CUL4B could regulate gene expression by DNA methylation. Study in XLMR shows that eliminated CUL4B expression in nervous cell resulted in the accumulation of WDR5 functioning as a critical substrate of CUL4B and increased the level of H3K4me3 on the neuronal gene promoters [11]. Therefore, CUL4B could degrade via ubiquitination of H3K4 methyltransferase component WDR5 to participate in the transcriptional regulation of epigenetic.
Hu et al. [3] identified CUL4B could play a role of epigenetic transcriptional regulation in gene expression. In order to clarify the mechanistic role of CUL4B, the proteins combined with CUL4B were identified by using affinity purification and mass spectrometry [3]. They demonstrated that CUL4B promoted PRC2 to bind target gene promoters and promoted PRC2 catalyzing the methylation of H3K27. Knockdown CUL4B resulted in loss of H2AK119 monoubiquitination and H3K27 trimethylation, leading to many genes abnormal epigenetic transcription including some suppressor genes such as p16, PTEN [3].
CRL4B plays a crucial role in the recruitment and stabilization of SUV39H1/HP1/DNMT3A onto IGFBP3 promoters [20]. This revealed that the abundance of CpG islands located within the IGFBP3 promoter region were significantly hypomethylated in CUL4B-depleted cells. All of these provide a powerful evidence of that CUL4B is required for the maintenance of epigenetic silencing of target genes [20].
The histone acetyltransferases (HAT) and histone deacetylases (HDAC) are two kinds of enzymes that regulate acetylation and deacetylation [47, 48]. HAT and HDAC are two kinds of important mechanisms in gene transcription regulation. HAT is associated with the activation of gene transcription, while HDAC is associated with the gene transcriptional suppression. The latter contributes to the tumor genesis and progression by inhibiting the transcription of tumor suppressor gene, which has become a popular target in the research of antitumor drug. Now, HDAC inhibitors are effective antineoplastic agents. It is widely used in clinical treatment in various malignancies [49].
CUL4B has been reported to regulate gene expression by transcriptional repression [3, 44]. HDAC1 and HDAC2 are involved in CUL4B-mediated transcriptional repression. Interference HDAC1/2 expression and using of TSA (a kind of HDAC inhibitor) could decrease the transcriptional repressive activity of CUL4B [44]. Meanwhile, up-regulation of CUL4B increased DDB1, HDAC1/2 and SIN3A which were binding to the gene promoter, accompany with catalyzing H2AK119 monoubiquitylation and decrease of the levels of histone acetylation [44]. CRL4B and SIN3A-HDAC complexes interact with each other and co-combine with the CDKN1A and CDKN1C promoters [44]. They caused H2AK119 monoubiquitylation and histone deacetylation and repressed target gene expression at the transcription level, and then regulated cell cycle as described above [44]. DNA methylation and histone ubiquitination/methylation in transcription repression prove that CUL4B may function as an oncogene in tumorigenesis. Although studies have shown that the roles of CUL4B in post-transcriptional modification, there are limited studies about epigenetic related roles so far. Further studies are needed to explore other epigenetic related roles of CUL4B.
CUL4B and apoptosis
Apoptosis is a complementary of cell proliferation in cell life activity [50, 51]. Many studies have found that tumor cell apoptosis is key to tumor therapy [52, 53]. Down-regulation of CUL4B can inhibit cell proliferation, lead to cell cycle S phase arrest and DNA replication abnormalities, which will promote tumor cell apoptosis. Study has shown that silencing of CUL4B inhibited colorectal cancer cells proliferation by inhibiting Wnt/β-catenin signaling pathway and promoted apoptosis in vitro and in vivo experiments [54]. Chen et al. [21] discovered that interference CUL4B gene expression could effectively inhibit the proliferation of osteosarcoma cells and induce apoptosis. Based on these studies, CUL4B may be a potential target for abnormal regulation associated with malignancy. However, it remains to carry out more studies to explore the relationship between CUL4B and cell apoptosis.
CUL4B related signaling pathways
As we all know, tumorigenesis has closely connections with abnormal signaling pathways. Recently, many studies reported that aberrant CUL4B expression is related to the activation of several signal transduction pathways. Wnt/β-catenin signaling pathway has been demonstrated abnormal activated in many kinds of cancer [55,56,57]. At present, Yuan et al. [19] found CUL4B functioned as a positively regulator in Wnt/β-catenin signaling in HCC. CUL4B and β-catenin are over-expressed in HCC tissues and have positive correlation. They manifested that β-catenin was downregulated when they knockdown CUL4B in HCC cell lines. Further, they discovered that CUL4B could protect β-catenin from GSK3-mediated degradation and prevent β-catenin degradation by repressing Wnt antagonists. In addition, CRL4B/PRC2 complexes was proved to activate Wnt signaling pathway through promoting H2AK119me1 and H3K27me3 to repress expression of Wnt inhibitors [19].
The PI3K/Akt/mTOR pathway is one of the most recurrently changed signaling pathway in cancers. Its abnormal activation plays a central role in promoting cancer genesis. PTEN, an inhibitor of PI3K signal pathway, is frequently lost in a multitude of cancers. Absent expression of PTEN leads to accumulation of PIP3 and then increases activity of AKT. As known, mTOR is abnormally activated because of PTEN loss, dysregulation of mTOR regulators and aberrant upstream signaling such as PI3K/AKT activation.
CRL4 might influenced mTOR activity potentially by interacting with DDB1, mTOR complex components mLST8 and Raptor [58]. Silence of CUL4B greatly decrease mTOR-mediated S6K1 phosphorylation [58]. Lately, Hu et al. proved that CUL4B knockdown in several different cancer cell lines led to increased expression of p16 and PTEN at the transcriptional level. Taken together, CUL4B may upregulate PI3K/Akt/mTOR pathway in oncogenesis to promote cellular survival, cell cycle progression and growth. However, details still need more investigations.
In order to discover and enhance cancer immunotherapy, abundant studies have focused on the novel examination about harnessing the treatment of cell signaling pathways mediating the tumor-associated immune suppression. Cancer occurrence and progression require a favorable microenvironment. AKT/β-catenin signaling cascades in malignancies are often regarded as therapeutic targets. Recently, Qian et al. [59] demonstrated that accumulation of myeloid-derived suppressor cells (MDSCs) whose presence acts as an important characteristic of tumor-permissive microenvironment, was mediated by downregulation of AKT/β-catenin pathway in CUL4B knock-out mice. CUL4B has previously been reported to be up-expressed in many malignancies and appears to be positively correlated with tumor progression [3, 19,20,21, 27]. on the contrary, Qian et al. [59] proved that CUL4B acted as a suppressor of tumorigenesis via limiting the partial functions of MDSCs to maintaining the immunosuppressive tumor microenvironment.
CUL4B and hematopoiesis
Hematopoietic stem cells (HSCs) have the ability to self-renew and produce identical, multipotent daughter HSCs or undergo differentiation to form any and every cell type in the blood system. Hematopoiesis is a tightly regulated, context-dependent process that relies on several intrinsic and extrinsic factors. Many HOX genes, highly conserved regulators, were expressed during the early stages of hematopoiesis within progenitor cells and underwent downregulation as cells differentiate. Paradoxically, some studies announced that constitutive CUL4A- and CUL4B-knockout mice exhibited no defects in hematopoiesis [13]. However, another showed that the deletion of either of the CUL4A nor CUL4B led to increased accumulation of HOXB4, indicating a redundant role for CUL4A and CUL4B in hematopoiesis [16]. It has been indicated that CRL4s played a significant role in regulating the self-renewal and differentiation of hematopoietic stem cells by targeting HOXA9 and HOXB4 for ubiquitination and degradation [16]. Future work is needed to identify the exact and different roles of CUL4A and CUL4B in hematopoiesis.
Conclusions and future perspectives
In conclusion, aberrant expression of CUL4B was found in several malignant diseases. CUL4B works as an oncogene in most of cancers. CUL4B influents human diseases development and progression by regulating a wide range of cellular processes, such as cell cycle, promoter methylation and histone deacetylation, DNA damage and repair, etc. Downregulation of CUL4B results in inhibited cancer cell proliferation, and it may act as a potential index of diagnose and therapeutic strategy in cancers. On the contrary, CUL4B acts as a tumor suppressor gene in the immunosuppressive tumor microenvironment. Upregulation of CUL4B is closely associated with the activation of many cancer-associated signaling pathways. Since CUL4B can influence human diseases development and progression by regulating a wide range of cellular processes, CUL4B will provide new insights into cancer diagnosis and treatment. However, further investigations are needed to clarify concrete mechanisms of CUL4B in cancers. Consequently, works on CUL4B in human diseases, especially cancers, deserve more concerns in the future.
Abbreviations
- CUL4B:
-
cullin 4B
- RING:
-
really interesting new gene
- CRL:
-
Cullin -Ring E3 ligase complex
- NLS:
-
nuclear localization signal
- XLMR:
-
x-linked mental retardation syndrome
- CDKs:
-
cyclin-dependent kinases
- CKIs:
-
CDK inhibitors
- WDR5:
-
WD repeat containing protein 5
- H3K4me3:
-
trimethylated H3K4
- HAT:
-
histone acetyltransferases
- HDAC:
-
histone deacetylases
- HCC:
-
hepatocellular carcinoma
- MDSCs:
-
myeloid-derived suppressor cells
- PrxIII:
-
peroxiredoxin III
- ROS:
-
reactive oxygen species
- HSCs:
-
hematopoietic stem cells
References
Sarikas A, Hartmann T, Pan ZQ. The cullin protein family. Genome Biol. 2011;12(4):220.
Lee J, Zhou P. Cullins and cancer. Genes Cancer. 2010;1(7):690–9.
Hu H, Yang Y, Ji Q, Zhao W, Jiang B, Liu R, Yuan J, Liu Q, Li X, Zou Y, et al. CRL4B catalyzes H2AK119 monoubiquitination and coordinates with PRC2 to promote tumorigenesis. Cancer Cell. 2012;22(6):781–95.
He F, Lu D, Jiang B, Wang Y, Liu Q, Liu Q, Shao C, Li X, Gong Y. X-linked intellectual disability gene CUL4B targets Jab1/CSN5 for degradation and regulates bone morphogenetic protein signaling. Biochim Biophys Acta. 2013;1832(5):595–605.
Zou Y, Mi J, Cui J, Lu D, Zhang X, Guo C, Gao G, Liu Q, Chen B, Shao C, et al. Characterization of nuclear localization signal in the N terminus of CUL4B and its essential role in cyclin E degradation and cell cycle progression. J Biol Chem. 2009;284(48):33320–32.
Jackson S, Xiong Y. CRL4s: the CUL4-RING E3 ubiquitin ligases. Trends Biochem Sci. 2009;34(11):562–70.
Chen CY, Tsai MS, Lin CY, Yu IS, Chen YT, Lin SR, Juan LW, Chen YT, Hsu HM, Lee LJ, et al. Rescue of the genetically engineered Cul4b mutant mouse as a potential model for human X-linked mental retardation. Hum Mol Genet. 2012;21(19):4270–85.
Zou Y, Mi J, Wang W, Lu J, Zhao W, Liu Z, Hu H, Yang Y, Gao X, Jiang B, et al. CUL4B promotes replication licensing by up-regulating the CDK2-CDC6 cascade. J Cell Biol. 2013;200(6):743–56.
Ohtake F, Baba A, Takada I, Okada M, Iwasaki K, Miki H, Takahashi S, Kouzmenko A, Nohara K, Chiba T, et al. Dioxin receptor is a ligand-dependent E3 ubiquitin ligase. Nature. 2007;446(7135):562–6.
Li X, Lu D, He F, Zhou H, Liu Q, Wang Y, Shao C, Gong Y. Cullin 4B protein ubiquitin ligase targets peroxiredoxin III for degradation. J Biol Chem. 2011;286(37):32344–54.
Nakagawa T, Xiong Y. X-linked mental retardation gene CUL4B targets ubiquitylation of H3K4 methyltransferase component WDR5 and regulates neuronal gene expression. Mol Cell. 2011;43(3):381–91.
Jiang B, Zhao W, Yuan J, Qian Y, Sun W, Zou Y, Guo C, Chen B, Shao C, Gong Y. Lack of Cul4b, an E3 ubiquitin ligase component, leads to embryonic lethality and abnormal placental development. PLoS ONE. 2012;7(5):e37070.
Liu L, Yin Y, Li Y, Prevedel L, Lacy EH, Ma L, Zhou P. Essential role of the CUL4B ubiquitin ligase in extra-embryonic tissue development during mouse embryogenesis. Cell Res. 2012;22(8):1258–69.
Liu L, Lee S, Zhang J, Peters SB, Hannah J, Zhang Y, Yin Y, Koff A, Ma L, Zhou P. CUL4A abrogation augments DNA damage response and protection against skin carcinogenesis. Mol Cell. 2009;34(4):451–60.
Yin Y, Lin C, Kim ST, Roig I, Chen H, Liu L, Veith GM, Jin RU, Keeney S, Jasin M, et al. The E3 ubiquitin ligase Cullin 4A regulates meiotic progression in mouse spermatogenesis. Dev Biol. 2011;356(1):51–62.
Hannah J, Zhou P. Distinct and overlapping functions of the cullin E3 ligase scaffolding proteins CUL4A and CUL4B. Gene. 2015;573(1):33–45.
Uhlen M, Fagerberg L, Hallstrom BM, Lindskog C, Oksvold P, Mardinoglu A, Sivertsson A, Kampf C, Sjostedt E, Asplund A, et al. Proteomics. Tissue-based map of the human proteome. Science. 2015;347(6220):1260419.
Mi J, Zou Y, Lin X, Lu J, Liu X, Zhao H, Ye X, Hu H, Jiang B, Han B, et al. Dysregulation of the miR-194-CUL4B negative feedback loop drives tumorigenesis in non-small-cell lung carcinoma. Mol Oncol. 2017;11:305–19.
Yuan J, Han B, Hu H, Qian Y, Liu Z, Wei Z, Liang X, Jiang B, Shao C, Gong Y. CUL4B activates Wnt/beta-catenin signalling in hepatocellular carcinoma by repressing Wnt antagonists. J Pathol. 2015;235(5):784–95.
Yang Y, Liu R, Qiu R, Zheng Y, Huang W, Hu H, Ji Q, He H, Shang Y, Gong Y, et al. CRL4B promotes tumorigenesis by coordinating with SUV39H1/HP1/DNMT3A in DNA methylation-based epigenetic silencing. Oncogene. 2015;34(1):104–18.
Chen Z, Shen BL, Fu QG, Wang F, Tang YX, Hou CL, Chen L. CUL4B promotes proliferation and inhibits apoptosis of human osteosarcoma cells. Oncol Rep. 2014;32(5):2047–53.
Tarpey PS, Raymond FL, O’Meara S, Edkins S, Teague J, Butler A, Dicks E, Stevens C, Tofts C, Avis T, et al. Mutations in CUL4B, which encodes a ubiquitin E3 ligase subunit, cause an X-linked mental retardation syndrome associated with aggressive outbursts, seizures, relative macrocephaly, central obesity, hypogonadism, pes cavus, and tremor. Am J Hum Genet. 2007;80(2):345–52.
Zou Y, Liu Q, Chen B, Zhang X, Guo C, Zhou H, Li J, Gao G, Guo Y, Yan C, et al. Mutation in CUL4B, which encodes a member of cullin-RING ubiquitin ligase complex, causes X-linked mental retardation. Am J Hum Genet. 2007;80(3):561–6.
Lee J, Zhou P. Pathogenic role of the CRL4 ubiquitin ligase in human disease. Front Oncol. 2012;2:21.
Vulto-van Silfhout AT, Nakagawa T, Bahi-Buisson N, Haas SA, Hu H, Bienek M, Vissers LE, Gilissen C, Tzschach A, Busche A, et al. Variants in CUL4B are associated with cerebral malformations. Hum Mutat. 2015;36(1):106–17.
Dong J, Wang XQ, Yao JJ, Li G, Li XG. Decreased CUL4B expression inhibits malignant proliferation of glioma in vitro and in vivo. Eur Rev Med Pharmacol Sci. 2015;19(6):1013–21.
Jiang T, Tang HM, Wu ZH, Chen J, Lu S, Zhou CZ, Yan DW, Peng ZH. Cullin 4B is a novel prognostic marker that correlates with colon cancer progression and pathogenesis. Med Oncol. 2013;30(2):534.
Wang X, Chen Z. Knockdown of CUL4B Suppresses the proliferation and invasion in non-small cell lung cancer cells. Oncol Res. 2016;24(4):271–7.
Hannah J, Zhou P. Regulation of DNA damage response pathways by the cullin-RING ubiquitin ligases. DNA Repair (Amst). 2009;8(4):536–43.
Moss J, Tinline-Purvis H, Walker CA, Folkes LK, Stratford MR, Hayles J, Hoe KL, Kim DU, Park HO, Kearsey SE, et al. Break-induced ATR and Ddb1-Cul4(Cdt)(2) ubiquitin ligase-dependent nucleotide synthesis promotes homologous recombination repair in fission yeast. Genes Dev. 2010;24(23):2705–16.
Kerzendorfer C, Whibley A, Carpenter G, Outwin E, Chiang SC, Turner G, Schwartz C, El-Khamisy S, Raymond FL, O’Driscoll M. Mutations in Cullin 4B result in a human syndrome associated with increased camptothecin-induced topoisomerase I-dependent DNA breaks. Hum Mol Genet. 2010;19(7):1324–34.
Guerrero-Santoro J, Kapetanaki MG, Hsieh CL, Gorbachinsky I, Levine AS, Rapic-Otrin V. The cullin 4B-based UV-damaged DNA-binding protein ligase binds to UV-damaged chromatin and ubiquitinates histone H2A. Cancer Res. 2008;68(13):5014–22.
Yi J, Lu G, Li L, Wang X, Cao L, Lin M, Zhang S, Shao G. DNA damage-induced activation of CUL4B targets HUWE1 for proteasomal degradation. Nucleic Acids Res. 2015;43(9):4579–90.
Lim S, Kaldis P. Cdks, cyclins and CKIs: roles beyond cell cycle regulation. Development. 2013;140(15):3079–93.
Gallastegui E, Bachs O. Expression and purification of recombinant cyclins and CDKs for activity evaluation. Methods Mol Biol. 2016;1336:9–12.
Stein GS, van Wijnen AJ, Stein JL, Lian JB, Montecino M, Zaidi SK, Braastad C. An architectural perspective of cell-cycle control at the G1/S phase cell-cycle transition. J Cell Physiol. 2006;209(3):706–10.
Alexander A, Karakas C, Chen X, Carey JP, Yi M, Bondy M, Thompson P, Cheung KL, Ellis IO, Gong Y, et al. Cyclin E overexpression as a biomarker for combination treatment strategies in inflammatory breast cancer. Oncotarget. 2017;8:14897.
Boueroy P, Hahnvajanawong C, Boonmars T, Saensa-Ard S, Anantachoke N, Vaeteewoottacharn K, Reutrakul V. Antitumor effect of forbesione isolated from Garcinia hanburyi on cholangiocarcinoma in vitro and in vivo. Oncol Lett. 2016;12(6):4685–98.
Li W, Zhang G, Wang HL, Wang L. Analysis of expression of cyclin E, p27kip1 and Ki67 protein in colorectal cancer tissues and its value for diagnosis, treatment and prognosis of disease. Eur Rev Med Pharmacol Sci. 2016;20(23):4874–9.
Ikeuchi K, Marusawa H, Fujiwara M, Matsumoto Y, Endo Y, Watanabe T, Iwai A, Sakai Y, Takahashi R, Chiba T. Attenuation of proteolysis-mediated cyclin E regulation by alternatively spliced Parkin in human colorectal cancers. Int J Cancer. 2009;125(9):2029–35.
Siu KT, Xu Y, Swartz KL, Bhattacharyya M, Gurbuxani S, Hua Y, Minella AC. Chromosome instability underlies hematopoietic stem cell dysfunction and lymphoid neoplasia associated with impaired Fbw7-mediated cyclin E regulation. Mol Cell Biol. 2014;34(17):3244–58.
Higa LA, Yang X, Zheng J, Banks D, Wu M, Ghosh P, Sun H, Zhang H. Involvement of CUL4 ubiquitin E3 ligases in regulating CDK inhibitors Dacapo/p27Kip1 and cyclin E degradation. Cell Cycle. 2006;5(1):71–7.
Brugarolas J, Chandrasekaran C, Gordon JI, Beach D, Jacks T, Hannon GJ. Radiation-induced cell cycle arrest compromised by p21 deficiency. Nature. 1995;377(6549):552–7.
Ji Q, Hu H, Yang F, Yuan J, Yang Y, Jiang L, Qian Y, Jiang B, Zou Y, Wang Y, et al. CRL4B interacts with and coordinates the SIN3A-HDAC complex to repress CDKN1A and drive cell cycle progression. J Cell Sci. 2014;127(Pt 21):4679–91.
Dawson MA, Kouzarides T. Cancer epigenetics: from mechanism to therapy. Cell. 2012;150(1):12–27.
Chen Z, Li S, Subramaniam S, Shyy JY, Chien S. Epigenetic regulation: a new frontier for biomedical engineers. Annu Rev Biomed Eng. 2017.
Fu M, Rao M, Wu K, Wang C, Zhang X, Hessien M, Yeung YG, Gioeli D, Weber MJ, Pestell RG. The androgen receptor acetylation site regulates cAMP and AKT but not ERK-induced activity. J Biol Chem. 2004;279(28):29436–49.
Timmermann S, Lehrmann H, Polesskaya A, Harel-Bellan A. Histone acetylation and disease. Cell Mol Life Sci. 2001;58(5–6):728–36.
Perri F, Longo F, Giuliano M, Sabbatino F, Favia G, Ionna F, Addeo R, Della Vittoria Scarpati G, Di Lorenzo G, Pisconti S. Epigenetic control of gene expression: potential implications for cancer treatment. Crit Rev Oncol Hematol. 2017;111:166–72.
Basu A, Sridharan S. Regulation of anti-apoptotic Bcl-2 family protein Mcl-1 by S6 kinase 2. PLoS ONE. 2017;12(3):e0173854.
Bai D, Yu S, Zhong S, Zhao B, Qiu S, Chen J, Lunagariya J, Liao X, Xu S. d-amino acid position influences the anticancer activity of galaxamide analogs: an apoptotic mechanism study. Int J Mol Sci. 2017;18(3):544.
Chen L, Fletcher S. Mcl-1 inhibitors: a patent review. Expert Opin Ther Pat. 2017;27(2):163–78.
Indovina P, Pentimalli F, Casini N, Vocca I, Giordano A. RB1 dual role in proliferation and apoptosis: cell fate control and implications for cancer therapy. Oncotarget. 2015;6(20):17873–90.
Song B, Zhan H, Bian Q, Li J. Knockdown of CUL4B inhibits proliferation and promotes apoptosis of colorectal cancer cells through suppressing the Wnt/beta-catenin signaling pathway. Int J Clin Exp Pathol. 2015;8(9):10394–402.
Shan Y, Ying R, Jia Z, Kong W, Wu Y, Zheng S, Jin H. LINC00052 promotes gastric cancer cell proliferation and metastasis via activating Wnt/beta-catenin signaling pathway. Oncol Res. 2017.
Hoffmeyer K, Raggioli A, Rudloff S, Anton R, Hierholzer A, Del Valle I, Hein K, Vogt R, Kemler R. Wnt/beta-catenin signaling regulates telomerase in stem cells and cancer cells. Science. 2012;336(6088):1549–54.
Moon RT, Bowerman B, Boutros M, Perrimon N. The promise and perils of Wnt signaling through beta-catenin. Science. 2002;296(5573):1644–6.
Ghosh P, Wu M, Zhang H, Sun H. mTORC1 signaling requires proteasomal function and the involvement of CUL4-DDB1 ubiquitin E3 ligase. Cell Cycle. 2008;7(3):373–81.
Qian Y, Yuan J, Hu H, Yang Q, Li J, Zhang S, Jiang B, Shao C, Gong Y. The CUL4B/AKT/beta-catenin axis restricts the accumulation of myeloid-derived suppressor cells to prohibit the establishment of a tumor-permissive microenvironment. Cancer Res. 2015;75(23):5070–83.
Badura-Stronka M, Jamsheer A, Materna-Kiryluk A, Sowińska A, Kiryluk K, Budny B, Latos‐Bieleńska A. A novel nonsense mutation inCUL4Bgene in three brothers with X-linked mental retardation syndrome. Clin Genet. 2010;77(2):141–4.
Wang HL, Chang NC, Weng YH, Yeh TH. XLID CUL4B mutants are defective in promoting TSC2 degradation and positively regulating mTOR signaling in neocortical neurons. Biochim Biophys Acta. 2013;1832(4):585–93.
Yin Y, Liu L, Yang C, Lin C, Veith GM, Wang C, Sutovsky P, Zhou P, Ma L. Cell autonomous and nonautonomous function of CUL4B in mouse spermatogenesis. J Biol Chem. 2016;291(13):6923–35.
Kerzendorfer C, Hart L, Colnaghi R, Carpenter G, Alcantara D, Outwin E, Carr AM, O’Driscoll M. CUL4B-deficiency in humans: understanding the clinical consequences of impaired Cullin 4-RING E3 ubiquitin ligase function. Mech Ageing Dev. 2011;132(8–9):366–73.
Cang Y, Zhang J, Nicholas SA, Bastien J, Li B, Zhou P, Goff SP. Deletion of DDB1 in mouse brain and lens leads to p53-dependent elimination of proliferating cells. Cell. 2006;127(5):929–40.
Pan WW, Zhou JJ, Yu C, Xu Y, Guo LJ, Zhang HY, Zhou D, Song FZ, Fan HY. Ubiquitin E3 ligase CRL4(CDT2/DCAF2) as a potential chemotherapeutic target for ovarian surface epithelial cancer. J Biol Chem. 2013;288(41):29680–91.
Yuan J, Jiang B, Zhang A, Qian Y, Tan H, Gao J, Shao C, Gong Y. Accelerated hepatocellular carcinoma development in CUL4B transgenic mice. Oncotarget. 2015;6(17):15209–21.
Mok MT, Cheng AS. CUL4B: a novel epigenetic driver in Wnt/beta-catenin-dependent hepatocarcinogenesis. J Pathol. 2015;236(1):1–4.
Qu Z, Li D, Xu H, Zhang R, Li B, Sun C, Dong W, Zhang Y. CUL4B, NEDD4, and UGT1As involve in the TGF-beta signalling in hepatocellular carcinoma. Ann Hepatol. 2016;15(4):568–76.
Jia L, Yan F, Cao W, Chen Z, Zheng H, Li H, Pan Y, Narula N, Ren X, Li H, et al. Dysregulation of CUL4A and CUL4B ubiquitin ligases in lung cancer. J Biol Chem. 2017;292(7):2966–78.
Authors’ contributions
All authors have contributed to data preparation, drafting and revising the manuscripts. Both authors read and approved the final manuscript.
Acknowledgements
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
Funding
This study was partly supported by: National Natural Science Foundation (No. 81270598, and No. 81473486); Taishan Scholar Foundation of Shandong Province.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Li, Y., Wang, X. The role of cullin4B in human cancers. Exp Hematol Oncol 6, 17 (2017). https://doi.org/10.1186/s40164-017-0077-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40164-017-0077-2