
Abstract
There is growing interest among nutritionists in feeding reduced protein diets to broiler chickens. Although nearly a century of research has been conducted providing biochemical insights on the impact of reduced protein diets for broilers, practical limitation still exists. The present review was written to provide insights on further reducing dietary protein in broilers. To construct this review, eighty-nine peer reviewed manuscripts in the area of amino acid nutrition in poultry were critiqued. Hence, nutritional research areas of low protein diets, threonine, glycine, valine, isoleucine, leucine, phenylalanine, histidine, and glutamine have been assessed and combined in this text, thus providing concepts into reduced protein diets for broilers. In addition, linkages between the cited work and least cost formation ingredient and nutrient matrix considerations are provided. In conclusion, practical applications in feeding reduced protein diets to broilers are advancing, but more work is warranted.
Similar content being viewed by others
Background
Investigations in reducing dietary crude protein (CP) in chickens began in the early 1940’s, which commenced nearly a century of exploration in this area of poultry nutrition research. Advances have been made, but many of the hurdles identified in the 1940’s vex poultry nutrition researchers today. Most notably, in the early 1940’s research was carried out in chickens fed various diets substituted with ingredients differing in protein and energy content, and it was established that diets with increased protein resulted in chickens having over two fold lower body fat content [1]. In addition to the former effect on carcass composition, low CP-amino acid fortified diet research in the early 1940’s failed to produce birds with equal performance to higher CP fed control birds that consumed less feed grade amino acids [2]. Decades thereafter ensuing industrial growth of broiler production, low CP-amino acid research continued, but it was widely accepted in practice that the dietary inclusion of amino acids beyond the sulfur containing amino acids and lysine would not economically sustain production in practice [3,4,5]. Towards the end of the twentieth century, adoption of the third feed grade amino acid, i.e., L-threonine (Thr), was aided by advances in ideal protein formulation tools and the expression of amino acids in dietary formulation on a digestible basis [6]. At present, L-valine (Val), the fourth limiting amino in most broiler diets devoid of meat meals, is being used in practice. Further, the approach of U.S. nutritionists on the use of feed grade amino acids in least cost poultry diets from 1950 to present has been reviewed [7] and is summarized in Table 1. Indeed, formulation adoption of a synthetic methionine source with crystalline lysine, threonine, valine, isoleucine, and arginine can allow for over a 50% reduction in soybean meal and over a 20% reduction in crude protein in a broiler grower diet based on corn, soybean meal, meat and bone meal, and distillers grains.
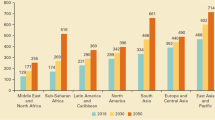
Production of poultry (e.g., primarily broilers) from the Food and Agriculture Organization of the United Nations 2005/2007 estimate to the 2050 projection will more than double from 82 to 181 million tons of poultry products [8]. In order to provide to the future need for poultry consumption, the bulk of the relative increase in poultry production will occur in developing countries with specific reference to those in Sub-Saharan Africa, North Africa/Near East, South Asia, and East Asia [8]. The former regions are more self-sufficient in cereals than that of oilseeds, deeming the need to broaden practical knowledge for feeding poultry low CP-feed grade amino acid fortified diets to satisfy future food security.
Utilization of the ideal protein concept allows for precision feeding of poultry, but a keen understanding of the birds’ dietary lysine need over time is warranted. However, additional amino acids must be continuously assessed as their minimums can vary based on dietary protein status, bird strain, bird environment, and company production objectives to satisfy economics [9]. As such, this review will provide an assessment of key amino acid considerations in the low CP puzzle and future research considerations which have been summarized in Table 2.
Recent low CP research
The increasing interest to lower dietary CP has led to more research defining interrelationships of nutrient digestion and absorption in reduced CP fed broilers. Following the ideal protein concept, a moderate reduction in CP, 20–30 g/kg, maintained performance and processing yields [10, 11]. Lowering CP more than 30 g/kg has been shown to inhibit performance and increase adipose fat deposition [10,11,12]. Reducing CP in a standard corn/soybean meal diet will lower soybean meal while increasing the proportion of corn, or other cereals, while decreasing feed-grade oils. This results in less intact protein-bound amino acids, increased inclusion of crystalline unbound amino acids and increased starch in the diet.
It was previously determined that intestinal uptakes of glucose and amino acids interrelationships are considered pivotal to broiler performance [13], and the process of starch degradation into glucose in broilers has been described [14]. Further, extent, rate, and site of nutrient digestion should be considered jointly to appropriately balance amino acids and glucose at sites of protein synthesis [15]. Starch and protein digestive dynamics (Table 2) are described as three-tiered process: digestion of starch and protein, absorption of glucose and amino acids from gut lumen, and the transition of glucose and amino acids across enterocytes to enter portal circulation [16]. Starch is more rapidly taken up than that of protein in broilers fed a sorghum-based diet [17]. Starch source plays an important role in its digestive rate and one important focus of digestive dynamics is that starch is digested too rapidly and protein too slowly [18]. Diets with decreased CP and increased starch flood the small intestine with glucose and compete with amino acids for absorption through their respective sodium dependent pathways [19]. Further, starch has a higher ileal digestibility coefficient than protein and amino acids [17, 20], pointing to the need for further research on digestive dynamics in low protein-crystalline amino acid-based diets [21].
Amino acid digestibility in reduced CP diets has been a recent focus. A reduction of 45 g/kg (210 and 165 g/kg) CP effects on broilers raised under tropical conditions significantly increased (0.790 vs. 0.744) average ileal amino acid digestibility coefficients by 6.18% [22]. Reducing CP by 45 g/kg and 30 g/kg increased ileal digestibilities of 16 amino acids by 5.82% and 9.10%, respectively [11, 23]. These fluctuations of ileal digestibility coefficients are credited to a reduction of endogenous amino acid flows in each respective diet. Pepsin, trypsin and remaining endogenous proteolytic enzymes, thus mucin as a large source of endogenous amino acids, are attenuated in reduced CP diets with less intact proteins [24]. The reduced endogenous amino acid flow, and the view that supplemental amino acids are 100% bioavailable and more rapidly absorbed in the proximal small intestine than protein bound amino acids [25], leads to the assumption that less amino acids are being supplied for processes that support gut health. Certain health promoting bacterial groups, that depend on amino acids, are known to stimulate production of mucin and catalyze the proliferation of epithelial cells [26, 27]. Maintaining the optimum function of the epithelial wall in the small intestine is important in controlling permeability, thus reducing incidences of leaky gut. Epithelial cells covering the intestinal villi are linked together through complex proteins; adherens junctions, desmosomes and tight junctions. Tight junctions facilitate paracellular permeability which has a role in absorbing nutrients and preventing entry of microbes and toxins into the body [28]. Increased intestinal permeability can have negative effects on health, bacterial and toxin translocation, lameness and economic losses from impaired performance [29]. Recent research has shown a significant increase in intestinal permeability in birds fed a low protein diet (170/150 g/kg of CP) fortified with all essential amino acids compared to a high protein diet (220/210 g/kg of CP) that exceeded recommendations by 10% [28]. There were no differences of intestinal permeability between a standard protein diet (202/190 g/kg) CP and the reduced protein diet. Reducing CP without the addition of essential amino acids exacerbates the effect of aflatoxicosis on intestinal permeability, which was improved with a 10% increase in CP [30]. These trials display that elevated gut permeability can be observed in birds fed inadequate levels of bound or purified amino acids, and that further research is needed to determine the role of individual amino acid concentrations on leaky gut. The former points to the need to feed the bird for gut integrity when low CP diets are offered, which is primarily highlighted in a subsequent section of this review on glutamine (Gln) (Table 2).
Excessive amino acids (nitrogen) undergo catabolism, presumably from the degradation of imbalanced amino acids, and can accumulate ammonia [31]. Reducing CP 30 g/kg (220 g/kg to 190 g/kg) increased plasma ammonia by 59.4% (7.27 vs. 4.56 mg/dL) and negatively impacted performance [32]. Ammonia detoxification occurs through its reaction with glutamate to form Gln. Glutamine enters the Krebs cycle which produces uric acid and nitrogen waste [33]. Glycine (Gly), serine (Ser), and glutamic acid are also needed for uric acid production, and the former can be limiting in low CP diets. It has been suggested that a minimum Gly + Ser value should be set between 2% to 2.5% [7]. An imbalance of amino acids resulting in inadequate ammonia detoxification could result in ammonia toxicity and reduced performance [31]. Moreover, bird performance could be further reduced due to increased litter nitrogen resulting in footpad lesions. For example, Ross 308 broilers fed reduced CP at 22 g/kg to 23 g/kg had reduced litter nitrogen and footpad lesions, with no adverse effects on performance or yields [34].
Threonine and glycine linkages
With specific reference to feed, consumer preferences that have been implemented into poultry practice at a rapid rate represent the production of broilers on feed with no antibiotics and no ingredients of animal origin. With reference to sustainable feed solutions, broiler diets will continue to be reduced in CP with improved nitrogen and water balance. Hence, lower CP diets contain less oilseed meals resulting in reduced water intake in broilers. Amino acid nutrition is central in the former preferences and feed solutions, and more specifically, Thr [6] (Table 2).
An assessment of Thr and Gly biochemistry must first consider Gly precursors. Hence, conclusions from early chick trials with Gly considered it to be essential for growth [2]. Serine is a Gly precursor and its dietary abundance was credited for the nonessentiality of Gly in a chick trial [35]. In two trials where low-CP diets were fed to chicks (minimal N-crystalline amino acid diets), Gly and Ser were required for good chick growth, but Ser completely replaced the Gly value on a mole to mole basis [36]. In addition to Ser sparing Gly, Baker and Sugahara [37], conducted two chick trials and demonstrated that choline can partially replace Gly through the pathway of choline to betaine to dimethylglycine to sarcosine to Gly. Hence, variability of research results on Gly needs and Gly by Thr interactions clearly stems from diet variability of Gly precursors.
Similar to that of Ser, Gly precursor knowledge is essential in understanding Gly needs and Thr is the principal Gly precursor in chicks. Thr metabolism with reference to Gly needs has been reviewed [38]. The glucogenic catabolism of Thr occurs through three main pathways through the de novo action of Thr aldolase, Thr dehydrogenase, and Thr dehydratase [38]. Further, both Thr aldolase and Thr dehydrogenase yield Gly to Ser to pyruvate. It must be pointed out that the reverse pathway from Ser to Gly exists. Hence, one-carbon units from formic acid allows for the reversible interconversion of Ser to Gly [39]. In order to determine if the reverse pathway from Gly to Thr existed, Baker et al. [40] conducted a chick trial with end products from Thr aldolase. Chicks fed a Gly rich diet deficient in Thr failed to sustain growth when the diet was fortified with acetaldehyde, indicating the reverse pathway of Gly to Thr is of no biological value in chicks [40]. Therefore, the catabolism of Thr yields pyruvate for energy or Gly to support metabolic processes through the conversion of protein, creatine, Ser, uric acid, guanidino acetic acid, glutathionine, and bile salts [38]. Further, research in chickens has shown that Thr dehydrogenase and Thr aldolase are the most active Thr-degrading enzymes [41].
The previously mentioned commercial trends in poultry production to move towards low or no antimicrobial-feed additive use and all vegetable-based feed ingredients have resulted in an increased awareness of gut inflammation and techniques to mitigate sub-clinical and clinical gut infections. Absorption of nutrients by the upper villus is dependent on mucin, which is partially dependent upon amino acids for integrity. Of the most limiting amino acids for mucin are methionine (Met) and Thr [42]. Methionine for the biosynthesis of cysteine, and Thr for its villi essentiality and the biosynthesis of Gly and Ser [42]. Furthermore, the presence of sub-clinical intestinal insults in poultry can increase the Thr value needed in least cost formulation [43] (Table 2). Similarly, it has been shown that finishing broilers reared on used pine shavings have increased Thr needs compared to those reared on fresh pine shavings [44, 45]. Thus, the Thr value in least cost formulation is dependent upon bird intestinal health and the dietary Gly level. Indeed, the commercial acceptance and use of dietary betaine (Table 2) for osmoregulatory and methyl donation may be interconnected with Thr metabolism. Further, research that assesses Thr and Gly interrelationships in broilers that have mild to moderate enteric stress is warranted, especially in situations where CP varies. Kidd et al. [46] assess CP by Thr interactions in chicks and failed to produce protein efficiency or nitrogen excretion interactions, feed efficiency was improved by CP and Thr interactions (CP linear × Thr linear). Moreover, the former research demonstrated that for each 1% decrease in dietary CP that nitrogen excretion by the bird is decreased approximately 8% [46].
The Gly need in chicks has been shown to be essential early in life and nonessential later in life [47]. Recent work by Kriseldi et al. [48] with starter chicks demonstrated success with low CP diets (2.6% reductions in CP) when the total Gly + Ser to digestible Lys ratio was 190. Similarly, Schutte et al. [49] conducted two trials with Ross chicks (1–14 and 1–21 d) fed corn or wheat and soybean meal-based diets with graduations of Gly and Glu and found responses to Gly, but not Glu. From their work it was concluded that the total Gly + Ser requirement for chicks was between 1.8% and 1.9% [49]. Because total Lys varied from 1.23% to 1.33% of diet, recent work of Kriseldi et al. [48] agrees with Schutte et al. [49]. Work by Heger and Pack [50] indicated that the Gly + Ser need is dependent upon CP. They demonstrated a Gly + Ser chick requirement range of 1.5% to 1.6% at 17% CP and 1.7% to 1.8% at 23% CP, presumably to aid in uric acid synthesis [50]. However, Lys was also increased in the high CP experiment suggesting relevance to a Gly + Ser to Lys ratio [50]. Future work with Gly + Ser, with relevance to Thr needs, should take biosynthetic precursors and gut health into consideration to validate least cost minimum levels and CP reduction potential.
Branch-chained amino acid interplay
As previously mentioned, L-Val is currently being integrated into broiler diets and L-isoleucine (Ile) is in the early phases of commercialization, and although diet costs are decreasing potential branched-chain amino acid (BCAA) antagonisms may be increasing. The interconnected nature of the BCAA has been a known phenomenon in poultry since the late 1960's [51], but this interplay has been largely ignored due to a series of articles published in the 1990’s and early 2000’s that indicated that antagonisms among BCAA were of little concern in practical broiler diets [52,53,54]. Thirty years prior to these experiments, Bray [55] observed similar responses in the practical diets of pullets but postulated that with increases in amino acid production, and their subsequent integration into poultry diets, that these antagonisms could be of particular concern when formulating poultry diets as BCAA excess was reduced.
Overwhelming data suggests that BCAA antagonism is primarily leucine (Leu) mediated, but the exact mechanism involved remains unknown [56,57,58]. In addition, the antagonism among the BCAA is not limited to Leu excess and can be induced when any of the three are provided at superoptimal levels [56]. These findings have shown that when considering the BCAA, balance among the BCAA is just as important as balancing them in the overall amino acid profile, if not more so. Ousterhout [59] found that feeding diets devoid in Ile and Val resulted in death after 18 and 19 days, respectively, but that when all three of the BCAA were removed in concert, mean survival times were 34 days, effectively doubling their survival time. On the other hand, D’Mello and Lewis [56] found that reductions in body weight gain caused by excess dietary Leu could be corrected by feeding increased levels of Ile and Val. Similarly, Tuttle and Balloun [60] observed that Leu induced reductions in body weight gain in poults and this response could be corrected through additional supplementation of Ile and Val in concert, but not individually.
These observations have led some researchers to theorize that currently implemented BCAA dietary requirements are influenced by inherent BCAA diet levels, leading to an over inflation to counteract the effects of antagonism [61, 62]. Therefore, determining the ratios of BCAA to each other and if these ratios are altered by diet amino acid supply, will be key to formulating to optimal BCAA in reduced CP diets (Table 2). This understanding may not only allow for use of L-Val and L-Ile in diet formulation, but their overall needed nutrient levels may lower, thus further reducing diet costs.
Phenylalanine + tyrosine considerations
Sequel to use of synthetic Met, and crystalline Lys, Thr, and Val in broiler diets, several amino acids, formerly thought to be of little practical concern, are poised to impact broiler performance and formulation strategies. Phenylalanine (Phe) has long been taken for granted as it has proven to be one of the least limiting amino acids in broiler diets in the previous decades, but its dietary levels have been observed to be marginally adequate or deficient in some practical diets [63, 64]. Maynard et al. [65] conducted a deletion experiment evaluating the 4th limiting amino acid in corn-soybean meal poultry diets and observed a negative effect from Phe deletion on carcass parts weights but could not reproduce effects in a subsequent experiment attempting to induce responses in live performance. It was theorized that the lack of repeated response may have been the result of not accounting for tyrosine (Tyr) when formulating the Phe test diets [65]. Similarly, other researchers have focused on Phe despite the fact that the interconnected relationship of Phe and Tyr has been known in poultry since the 1940’s [2, 66,67,68]. Sasse and Baker [69] determined that the Tyr requirement makes up 42.5% of the Phe plus Tyr requirement, which has been reported to be 112% of digestible lysine [70]. Unfortunately the studies evaluating the Phe plus Tyr requirements have focused on the first 2 weeks of the chick’s life and therefore have left a void in literature, leading to the generation of suggested amino acid requirements that remain constant for the entire growout period [71]. As poultry diets are continually refined and amino acid excesses are minimized, the levels of dietary Phe and Tyr may influence diet formulation not only due to concerns of reduced growth performance (Table 2) but also in broiler health as Phe has been found to reduce mortality in the face of mycotoxin poisoning [72].
Histidine and the need for further research
As diets continue to be lowered in CP, research on histidine (His) responses and its minimum in least cost formulation warrant attention (Table 2). In 1926, Cox and Rose [73] created a purified diet, virtually devoid of His by precipitation of hydrolyzed casein, and fed the diet to rats with and without purine supplementation in a long-term study (100 plus days). Dietary addition of adenine, guanine, creatinine, creatine, and their combinations failed to improve performance in the His deficient diet indicting the essentiality of His via irreversible purine synthesis [73]. However, in 1959 Leveille and Fisher [74] assessed His needs for maintenance using a His-free diet in the adult rooster in a short-term study (5-day feeding period) and determined it to be non-essential as measured by nitrogen balance. Subsequent work utilized the former His-free diet in the same laboratory in 1960 [75], but fed the His-free diet to adult roosters for 14 days, and found decreased breast muscle anserine and carnosine levels, indicating the essentiality of His. Further, Fisher et al. [76] fed adult cockerels protein depleted diets for 7 weeks and found that wet breast muscle had decreased carnosine and increased His, from which they concluded that the high breast His arose from carnosine breakdown or the decrease in its synthesis.
Robbins et al. [77] fed chicks His graduations and assessed growth performance, plasma free His, and breast muscle carnosine. It was determined that maximized stores of muscle carnosine and adequate growth based on His adequacy were required before plasma free His levels accumulated. It was concluded that His evaluation through reduced or His-free diets should include a pre-experimental adjustment period in order to reduce both muscle carnosine and plasma His so as not to underestimate the His response [77].
Kai et al. [78] fed diets with deficient (67% of requirement), adequate (100% of requirement), and excess (200% of requirement) His to broilers from days 15 to 24 and found that His deficiency impaired growth and breast meat. These reductions in growth and breast meat yield were also accompanied by a complete depletion in breast muscle carnosine [78]. Moreover, recent research in our laboratory [79] demonstrated that when diets containing approximately 89% of the His requirement were fed to broilers from 15 to 35 days of age, no negative effects were observed on body weight gain, but breast meat and right thigh yield were significantly reduced. These data suggest that broilers will mobilize bound carnosine to maintain growth, sacrificing carcass part yields in the process. Thus, low CP diets marginal in His in grower and finisher diet phases may result in unexpected yield losses at processing. Furthermore, assessing His needs in broilers warrants depletion diets, similar to those implemented in mineral studies, as suggested by Robbins et al. [77].
Glutamine and intestinal recovery from low protein diets
A better understanding of the bird’s intestinal homeostasis for amino acid needs and dietary form of amino acids in low CP diets necessitates research. Feeding reduced CP diets to broilers has shown a marked increase in diet AME values and amino acid digestibility coefficients, but upon further investigation these changes appear to be the result of diet composition, and not a result of the birds’ metabolism [11, 22, 80]. With feed-grade amino acids making up a larger portion of broiler diets, amino acids are able to bypass the gut and enter the blood stream without the need of digestion [25]. Unfortunately, this beneficial attribute of reduced CP diets may not be as beneficial as it seems when the status of the gut is examined. Various researchers have reported negative effects on live performance and intestinal morphology when feeding reduced CP diets [81,82,83]. These negative responses may be the result of how the gastrointestinal tract feeds through first pass metabolism. Increased feed grade amino acid usage is accompanied by a decrease in non-essential amino acids, adhering to the ideal protein concept, which deprives the gut mucosa of its primary energy source. When considering nonessential amino acids, the gut mucosa consumes and utilizes 66%, 98%, and 99% of Gln, glutamate, and aspartate, respectively, from intraluminal sources before they reach the blood supply [84]. Of these three, only Glu can be measured in arterial blood supply post absorption [84]. Non-essential amino acid reports in the literature are predominated by Gly, which, unlike Gln, is not used to the extent in the bird’s lumen [84]. As CP continues to be reduced, therefore, non-essential amino acid supply for intestinal integrity (e.g., Gln) warrants attention (Table 2).
Bortoluzzi et al. [85] reviewed Gln and indicated that its supplementation may reduce intestinal atrophy and aid in mucosal repair following trauma. The former review [85] focused on pathogen challenges, particularly coccidiosis, and hypothesized that amino acid supplementation could aid in supporting the bird’s intestinal integrity in the absence of subtherapeutic antibiotic use in diets. Under reduced CP conditions the trauma in which the gut is subjected to could be a form of starvation due to the reduced amount of available nonessential amino acids. The intestinal changes observed under reduced CP conditions include reduced villi height and absorptive surface area, sharing some commonalities with a coccidia infection [82, 83, 86]. Further, Gln supplementation has been shown to increase intestinal weight and villi height [87, 88], in addition to providing intraluminal and arterial energy [84]. Nontraditional levels of amino acids may be needed in reduced CP diets to offset the reduction in nonessential amino acid levels in order to maintain and stimulate gut development (Table 2). Further, nitrogen supply in the lumen and its effect on pathogenic bacteria is not entirely understood.
Conclusions for linear programming
Realized feeding of reduced CP diets for broilers requires constant attention to the linear programming nutrient matrix. For example, formulators are considering establishing true protein values for ingredients to better reflect cereal and oilseed nitrogen levels [89] (Table 2). Chronological events of linear programing in the U.S. from the 1950’s to present has been reviewed [7], and include: removal of CP nutrient minimums, removal of feed grade amino acid ingredient maximum, addition of all amino acids on a digestible basis in the ingredient and nutrient matrix, and expression of digestible amino acids to Lys ratios in the formula matrix (Table 1). The former events became realized as feed grade amino acid prices decreased and resulted in lower diet costs. Although feed cost is the highest contributor to broiler production costs, today’s nutritionists are evaluating the reduction in nitrogen output while considering unique, and somewhat revolutionary, criteria (e.g., bird water intake, nutrient intake as a function of life-cycle analysis, welfare, and soil and air quality). For example, Table 2 is provided to summarize amino acid responses while considering both the ingredient and nutrient matrix in least cost formulation to provide commercial nutritionists information on reducing nitrogen excesses and researchers with concepts for improving nitrogen efficiency.
Availability of data and materials
Data may be provided following request to the corresponding author.
Abbreviations
- BCAA:
-
Branched-chain amino acids
- CP:
-
Crude protein
- Gln:
-
Glutamine
- Gly:
-
Glycine
- His:
-
Histidine
- Ile:
-
Isoleucine
- Leu:
-
Leucine
- Lys:
-
Lysine
- Met:
-
Methionine
- Phe:
-
Phenylalanine
- Ser:
-
Serine
- Thr:
-
Threonine
- Tyr:
-
Tyrosine
- Val:
-
Valine
References
Fraps GS. Relation of protein, fat, and energy of the ration to the composition of chickens. Poult Sci. 1943;22:421–4.
Almquist HJ, Grau CR. The amino acid requirements of the chick. J Nutr. 1944;28:325–31.
Almquist HJ. Proteins and amino acids in animal nutrition. 5th ed. New York: S. B. Penick and Co; 1972.
Lipstein B, Bornstein S. The replacement of some of the soybean meal by the first limiting amino acids in practical broiler diets. 2. Special additions of methionine and lysine as partial substitutes for protein in finisher diets. Br Poult Sci. 1975;16:189–200.
Waldroup PW, Mitchell RJ, Payne JR, Hazen KR. Performance of chicks fed diets formulated to minimize excess levels of essential amino acids. Poult Sci. 1976;55:243–53.
Kidd MT. Nutritional considerations concerning threonine in broilers. World’s Poult Sci J. 2000;56:139–51.
Kidd MT, Tillman PB, Waldroup PW, Holder W. Feed-grade amino acid use in the United States: the synergetic inclusion history with linear programming. J Applied Poult Res. 2013;22:583–90.
Alexandratos N, Bruinsma J. World agriculture towards 2030/2050: the 2012 revision. ESA working paper no. 12-03. Agriculture development economics division. Food and agriculture Organization of the United Nations. 2012. Rome, Italy.
Kidd MT, Tillman PB. Key principles concerning dietary amino acid responses in broilers. An. Feed Sci. Tech. 2016;221:314–22.
Van Harn J, Dijkslag MA, van Krimpen M. Effect of low dietary protein levels on performance, litter quality and footpad lesions in broilers. Poult Sci. 2019;98:4868–77.
Chrystal PV, Moss AF, Khoddami A, Naranjo VD, Selle PH, Liu SY. Impacts of reduced-crude protein diets on key parameters in male broiler chickens offered maize-based diets. Poult Sci. 2020;99:505–16.
Fancher BI, Jensen LS. Male broiler performance during the starting and growing periods as affected by dietary protein, essential amino acids and potassium levels. Poult Sci. 1989;68:1385–95.
Cant JP, McBride BW, Croom WJ. The regulation of intestinal metabolism and its impact on whole animal energetics. Anim Sci. 1996;74:2541–53.
Moran ET. Starch digestion in fowl. Poult Sci. 1982;61:1257–67.
Liu SY, Selle PH. A consideration of starch and protein digestive dynamics in chicken-meat production. World’s Poult Sci. 2015;71:297–310.
Liu SY, Selle PH. Starch and protein digestive dynamics in low-protein diets supplemented with crystalline amino acids. Anim Prod Sci. 2017;57:2250–6.
Liu SY, Selle PH, Cowieson AJ. The kinetics of starch and nitrogen digestion regulate growth performance and nutrient utilisation of broilers fed coarsely ground, sorghum-based diets. Anim Prod Sci. 2013;53:1033–40.
Selle PH, Liu SY. The relevance of starch and protein digestive dynamics in poultry. J Appl Poult Res. 2019;28:531–45.
Moss AF, Sydenham CJ, Khoddami A, Naranjo VD, Liu SY, Selle PH. Dietary starch influences growth performance, nutrient utilisation and digestive dynamics of protein and amino acids in broiler chickens offered low-protein diets. Anim Feed Sci Tech. 2018;237:55–67.
Selle PH, Liu SY, Cai J, Cowieson AJ. Steam-pelleting temperatures, grain variety, feed form and protease supplementation of mediumly ground, sorghum-based broiler diets: influences on growth performance, relative gizzard weights, nutrient utilisation, starch and nitrogen digestibility. Anim Prod Sci. 2013;53:378–87.
Selle PH, Truong HH, Liu SY. On free amino acids: their role in starch and protein digestive dynamics. Proc. Aust. Poult. Sci. Symp. 2015;26:97–105.
Awad EA, Zulkifli I, Farjam AS, Chwen LT, Hossain MA, Alijoubori A. Effect of low-protein diet, gender and age on the apparent ileal amino acid digestibilities in broiler chickens raised under hot-humid tropical condition. Indian J Anim Sci. 2016;86:696–701.
Hilliar M, Hargreave G, Girish CK, Barekatain R, Wu S-B, Swick RA. Using crystalline amino acids to supplement broiler chicken requirements in reduced protein diets. Poult Sci. 2020;99:1551–63.
Lien KA, Sauer WC, He JM. Dietary influences on the secretion into and degradation of mucin in the digestive tract of monogasteric animals and humans. J Anim Feed Sci. 2001;10:223–45.
Wu GY. Amino acids: metabolism, functions, and nutrition. Amino Acids. 2009;37:1–17.
Deplancke B, Gaskins HR. Microbial modulation of innate defense: goblet cells and the intestinal mucus layer. Am J Clin Nutr. 2001;73:1131–41.
Hörmann N, Brandão I, Jäckel S, Ens N, Lillich M, Walter U, Reinharft C. Gut microbial colonization orchestrates TLR2 expression, signaling and epithelial proliferation in the small intestinal mucosa. PLoS One. 2014;9(11):e113080.
Barekatain R, Nattrass G, Tilbrook AJ, Chousalkar K, Gilani S. Reduced protein diet and amino acid concentration alter intestinal barrier function and performance of broiler chickens with or without synthetic glucocorticoid. Poult Sci. 2019;98:3662–75.
Gilani S, Howarth GS, Kitessa SM, Tran CD, Forder REA, Hughes RJ. New biomarkers for increased intestinal permeability induced by dextran sodium sulphate and fasting in chickens. Ani Prod Sci. 2016;56:1984–97.
Chen X, Naehrer K, Applegate T. Interactive effects of dietary protein concentration and aflatoxin B1 on performance, nutrient digestibility, and gut health in broiler chicks. Poult Sci. 2016;95:1312–25.
Stern RA, Mozdziak PK. Differential ammonia metabolism and toxicity between avian and mammalian species, and effect of ammonia on skeletal muscle: a comparative review. J Anim Physiol Anim Nutr. 2019;103:774–85.
Ospina-Rojas IC, Murakami AE, Duarte CRA, Eyng C, Oliveira CAL, Janeiro V. Valine, isoleucine, arginine and glycine supplementation of low-protein diets for broiler chickens during the starter and grower phases. Br Poult Sci. 2014;55:766–73.
Salway JG. The Krebs uric acid cycle: a forgotten Krebs cycle. Trends Biochem Sci. 2018;43:847–9.
van Harn J, Dijkslag MA, van Krimpen MM. Effect of low protein diets supplemented with free amino acids on growth performance, slaughter yield, litter quality, and footpad lesions of male broilers. Poult Sci. 2019;98:4868–77.
Sugahara M, Ariyoshi S. The nonessentiality of glycine and the essentiality of L-proline in chick nutrition. Agr Biol Chem. 1967;31:106–10.
Baker DH, Sugahara M, Scott HM. The glycine-serine interrelationship in chick nutrition. Poult Sci. 1968;47:1376–7.
Baker DH, Sugahara M. Nutritional investigation of the metabolism of glycine and its precursors by chicks fed a crystalline amino acid diet. Poult Sci. 1970;49:756–60.
Kidd MT, Kerr BJ. L-Threonine for poultry: A review. J. Appl. Poult Sci. 1996;5:538–367.
Meister A. Biochemistry of the amino acids. Vol. II. New York: Academic press; 1965. p. 636–72.
Baker DH, Hill TM, Kleiss AJ. Nutritional evidence concerning formation of glycine from threonine in the chick. J Anim Sci. 1972;34:582–6.
Davis AJ, Austic RE. Dietary threonine imbalance alters threonine dehydrogenase activity in isolated hepatic mitochondria of chicks and rats. J Nutr. 1994;124:1667–77.
Moran ET Jr. Gastric digestion of protein through pancreatic action optimizes intestinal forms for absorption, mucin formation and villus integrity. An. Feed Sci. Tech. 2016;221:284–303.
Star L, Rovers M, Corrernt E, van der Klis DJ. Threonine requirement of broiler chickens during subclinical intestinal Clostridium infection. Poult Sci. 2012;91:643–52.
Kidd MT, Barber SJ, Virden WS, Dozier WA Jr, Chamblee DW, Wiernusz C. Threonine responses of cobb male finishing broilers in differing environmental conditions. J Appl Poult Res. 2003;12:115–23.
Corzo A, Kidd MT, Dozier WA, Pharr GT, Koutsos EA. Dietary threonine needs for growth and immunity of broilers raised under different litter conditions. J Appl Poult Res. 2007;16:574–82.
Kidd MT, Gerard PD, Heger J, Kerr BJ, Rowe D, Sistani K, et al. Threonine and crude protein responses in broiler chicks. An Feed Sci Tech. 2001;94:57–64.
Coon CN, Grossie VB Jr, Couch JR. Glycine-serine requirement for chicks. Poult Sci. 1974;53:1709–13.
Kriseldi R, Tillman PB, Jiang Z, Dozier WA III. Effects of feeding broilers reduced crude protein diets on growth performance, nitrogen excretion, and plasma uric acid concentration of broiler chicks during the starter period. Poult Sci. 2018;97:1614–26.
Schutte JB, Smink W, Pack M. Requirement of young broiler chicks for glycine + serine. Arch Geflugelk. 1997;61:43–7.
Heger J, Pack M. Effects of dietary glycine and serine on starting broiler chick performance as influenced by dietary crude protein levels. Agribiol Res. 1996;49:257–65.
Mathieu D, Scott HM. Growth depressing effect of excess leucine in relation to the amino acid composition of the diet. Poult. Sci. 1968;47:1694 (Abstr.).
Barbour G, Latshaw LD. Isoleucine requirement of broiler chicks as affected by the the concentrations of leucine and valine in practical diets. Br Poult Sci. 1992;33:561–8.
Burnham D, Emmans GC, Gous RM. Isoleucine requirements of the chicken: the effect of excess leucine and valine on the response to isoleucine. Br Poult Sci. 1992;33:71–87.
Waldroup PW, Kersey JH, Fritts CA. Influence of branched-chain amino acid balance in broiler diets. Int J Poult Sci. 2002;1:136–44.
Bray DJ. The isoleucine and valine nutrition of young laying pullets as influenced by excessive dietary leucine. Poult Sci. 1970;49:1334–41.
D’Mello JPF, Lewis D. Amino acid interactions in chick nutrition. 2. Interrelationships between leucine, isoleucine and valine. Br. Poult. Sci. 1970;11:313–23.
Boldizsár HK, Boorman KN, Buttery PJ. The effect of excess leucine on valine catabolism in the chick. Br J Nutr. 1973;30:501–10.
Smith TK, Austic RE. The branched-chain amino acid antagonism in chicks. J Nutr. 1978;108:1180–91.
Ousterhout L. Survival time and biochemical changes in chicks fed diets lacking different essential amio acids. J Nutr. 1960;2:226–34.
Tuttle WL, Balloun SL. Leucine, isoleucine and valine interactions in Turkey poults. Poult Sci. 1976;55:1737–43.
Farran MT. Branched-chain amino acids: requirements and antagonism in the male broiler chick. PhD. Dissertation. College Park: University of Maryland; 1987.
Maynard CW, Liu SY, Kidd MT. Branched-chain amino acids: potential antagonisms in practical formulation. Proc Aust Poult Sci Symp. 2020;31:67–70.
Austic RE. Phiddling with phenylalanine: influence of dietary amino acids on enzymes of phenylalanine metabolism in the chick. Cornell Nutrition Conference For Feed Manufacturers. 2009:122–32.
Chrystal PV. Personal communication; 2019.
Maynard CW, Liu SY, Lee JT, Caldas J, Diehl E, Rochell SJ, et al. Determining the 4th limiting amino acid in low crude protein diets for male and female cobb mv × 500 broilers. Br Poult Sci. 2020. https://doi.org/10.1080/00071668.2020.1782348.
Grau CR. Interrelations of phenylalanine and tyrosine in the chick. J Bio Chem. 1947;170:661–9.
Lartey FM, Austic RE. Phenylalanine requirement, imbalance, and dietary excess in one-week-old chicks: growth and phenylalanine hydroxylase activity. Poult Sci. 2008;87:291–7.
Sakomura NK, Ekmay RD, Mei SJ, Coon CN. Lysine, methionine, arginine, valine, isoleucine, leucine, and threonine maintenance requirements of broiler breeders. Poult Sci. 2015;94:2715–21.
Sasse CE, Baker DH. The phenylalanine and tyrosine requirements and their interrelationship for the young chick. Poult Sci. 1972;51:1531–6.
Franco SM, Tavernari FC, Maia RC, Barros VRSM, Albino LFT. Rostagno, et al. estimation of optimal ratios of digestible phenylalanine plus tyrosine, histidine, and leucine to digestible lysine for performance and breast yield in broilers. Poult. Sci. 2017;96:829–37.
Wu G. Dietary requiremnts of synthesizable amino acids by animals: a paradigm shift in protein nutrition. J Anim Sci. 2014;5:34–45.
Gibson RM, Bailey CA, Kubena LF, Huff WE, Harvey RB. Impact of l-phenylalanine supplementation on the performance of three-week-old broilers fed diets containing ochratoxin a. 1. Effects on body weight, feed conversion, relative organ weight, and mortality. Poult. Sci. 1990;69:414–9.
Cox GJ, Rose WC. Can purines, creatinine, or creatine replace histidine in the diet for purposes of growth? J Biol Chem. 1926;68:769–80.
Leveille GA, Fisher H. Amino acid requirements for maintenance in the adult rooster. II. The requirements for glutamic acid, histidine, lysine and arginine. J Nutr. 1959;69:289–94.
Leveille GA, Shapiro R, Fisher H. Amino acid requirements for maintenance in the adult rooster. IV. The requirements for methionine, cystine, phenylalanine, tyrosine and tryptophan; The adequacy of the determined requirements. J Nutr. 1960;72:8–15.
Fisher H, Konlande J, Strumeyer D. Levels of histidine and histidine derivatives in breast muscle of protein-depleted and repleted adult cockerels. Nutr Metabol. 1975;18:120–6.
Robbins KR, Baker DH, Norton HW. Histidine status in the chick as measured by growth rate, plasma free histidine and breast muscle carnosine. J Nutr. 1977;107:2055–61.
Kai S, Watanabe G, Kubota M, Kadowaki M, Fujimura S. Effect of dietary histidine on contents of carnosine and anserine in muscles of broilers. Anim Sci J. 2015;86:541–6.
Maynard CW, Kidd MT, Chrystal PV, Selle PH, Liu SY. Determination of the 4th limiting amino acid for Ross 308 male broilers from 15 to 35 day in low crude protein wheat-based diets. Poult. Sci. 2020;98(E-suppl. 1):68.
Maynard CW, Ghane AE, Chrystal PV, Selle PH, Liu SY. Low crude protein diets: Does the modern broiler adapt to diet composition through manipulation of nutrient metabolism or are macro nutrient utilization values fiat data points? Poult. Sci. 2020;98(E-suppl. 1):64.
Abbasi MA, Mahdavi AH, Samie AH, Jahanian R. Effects of different levels of dietary crude protein and threonine on performance, humoral immune responses and intestinal morphology of broiler chicks. Braz J Poult Sci. 2014;16:35–44.
Ding XM, Li DD, Li ZR, Wang JP, Zeng QF, Bai SP, et al. Effects of dietary crude protein levels and exogenous protease on performance, nutrient digestibility, trypsin activity and intestinal morphology in broilers. Livest Sci. 2016;193:26–31.
Law FL, Zulkifli I, Soleimani AF, Liang JB, Awad EA. The effects of low-protein diets and protease supplementation on broiler chickens in a hot and humid tropical environment. Asian-Australss.J. Anim. Sci. 2018;31:1291–300.
Wu G. Intestinal mucosa amino acid catabolism. J Nutr. 1998;128:1249–52.
Bortoluzzi C, Rochell SJ, Applegate TJ. Threonine, arginine, and glutamine: influences on intestinal physiology, immunology, and microbiology in broilers. Poult Sci. 2018;97:937–45.
Fernando MA, McGraw BM. Mucosal morphology and cellular renewal in the intestine of chickens following a single infection of eimeria acervulina. J Paristol. 1973;59:493–501.
Bartell SM, Batal AB. The effect of supplemental glutamine on growth performance, development of the gastrointestinal tract, and humoral immune response of broilers. Poult Sci. 2007;86:1940–7.
Oxford JH, Selvaraj RK. Effects of glutamine supplementation on broiler performance and intestinal immune parameters during an experimental coccidiosis infection. J Appl Poult Res. 2019;28:1279–87.
Mariott F, Tomé D, Mirand PP. Converting nitrogen into protein-beyond 6.25 and Jones’ factors. Crit. Rev. Food Sci Nutr. 2008;48:117–84.
Acknowledgements
The authors would like to thank Professor Jianmin Yuan Ph.D. of China Agriculture University for the invitation and encouragement for us to prepare this updated typescript on amino acid nutrition.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
The authors’ contributions are as follows: all authors contributed equally on manuscript preparation, writing, and finalizing. All authors read and approved the final version of the manuscript and approved publication.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Kidd, M.T., Maynard, C.W. & Mullenix, G.J. Progress of amino acid nutrition for diet protein reduction in poultry. J Animal Sci Biotechnol 12, 45 (2021). https://doi.org/10.1186/s40104-021-00568-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40104-021-00568-0