
Abstract
Foodborne Shiga toxin-producing Escherichia coli (STEC) cause severe diarrhea and hemolytic uremic syndrome (HUS) in humans. However, traditional methods for STEC sterilization are difficult to apply to fresh food. To control the pathogen, phage infecting E. coli O157:H7 were isolated and characterized. The isolated phage vB_EcoM-ECP26 had an icosahedral head and a contractile tail, and was classified as belonging to the Myoviridae family. The phage showed a broad host range against STEC and exhibited a large burst size of 1914 PFU/cell. The phage was highly stable at high temperatures (65 °C) and wide ranges of pH (4–10). The genome of vB_EcoM-ECP26 consists of 136,993 nucleotides, 214 open reading frames, and does not contain lysogenicity-related genes. Phylogenetic analysis showed that vB_EcoM-ECP26 is a V5-like species. STEC O157 growth was inhibited by vB_EcoM-ECP26 for 8 h. Furthermore, this phage not only significantly decreased the STEC population (p < 0.05), but also persisted in fresh lettuce at 4 °C for 5 days. Therefore, these results reveal that the novel lytic phage vB_EcoM-ECP26 could be a useful agent for the control of foodborne STEC.
Similar content being viewed by others


Introduction
Escherichia coli is a Gram-negative, rod-shaped bacteria that is commonly found in the gut of mammals, including humans [44]. Most strains of E. coli are not only harmless but also play an important role in intestinal health [24]. However, some pathogenic E. coli strains can cause severe illness. Pathogenic E. coli can be classified based on their virulence factors; the most problematic type may be Shiga toxin-producing Escherichia coli (STEC) which cause severe diarrhea and hemolytic uremic syndrome (HUS) in humans and livestock [10, 35]. Most infections caused by STEC are foodborne and caught by consuming unheated food. Since STEC is destroyed at 70 °C, infections can be prevented by sufficient heating [45]. However, fresh fruits and leafy vegetables used in ready-to-eat (RTE) salads which are not heated or sterilized during the food preparation process often carry STEC [9]. Specifically, leafy vegetable-associated food poisoning by STEC is a problem worldwide. In the United States, the number of STEC related outbreaks are increasing and STEC were linked to 18% of leafy vegetable-associated outbreaks between 1973 and 2012 [26]. In 2011, 3950 people were infected by STEC in Germany, 855 of whom had HUS and 53 of whom died [8]. In 2016, 165 cases of STEC were reported in the UK, with 66 patients requiring hospitalization and 9 patients developing HUS [22].
For these reasons, several attempts have been made to use biological compounds such as antimicrobial peptides (AMPs) or essential oils to sterilize fresh food [23, 30]. However, due to their low efficiency and negative effects on food, commercial food applications have been difficult [40]. Bacteriophages, viruses that infect bacteria, could be an alternative strategy that to specifically target STEC in various foods [36, 47]. Several phage products have been approved and commercialized for food safety and protection such as LISTEXTM, EcoShieldTM and SalmoFreshTM [3, 51, 53].
However, there are still some challenges with phage application f½ or food safety. Temperate phages infects bacteria via both lysogenic and lytic cycles, and can be problematic because they do not lyse the bacterial cell in the prophage stage [54]. This could eventually lead to superinfection [14]. Moreover, the temperate phage can transfer toxin-associated genes like Shiga toxin and Botulinum toxin or antibiotic resistance genes through specialized transduction [13, 31]. Therefore, the use of temperate phage should be considered carefully. Lytic phages, on the other hand, only infect using the lytic cycle and are rarely capable of horizontal gene transfer; thus, lytic phages can be a reasonable vehicle for the biocontrol of STEC [18, 37].
As noted above, lytic phage could be a good candidate for foodborne STEC control. The aim of this study was to exploit more efficient lytic phage that targets foodborne STEC, we isolated and analyzed various characters of the putative lytic E. coli phage including environmental stability and growth inhibition activity. Bioinformatic studies were performed to identify the potential genetic risks of the phage genome. Furthermore, assessment of phage bactericidal activity on romaine lettuce was also investigated. These data will be useful in the study of lytic phages for the control of foodborne STEC.
Materials and methods
Phage host bacteria preparation and phage isolation
Escherichia coli NCCP 13,930 was used as the host bacteria for phage isolation. Host bacteria was grown at 37 °C on Luria–Bertani agar and broth (Difco, Detroit, MI, USA) supplemented with 10 mM CaCl2 (LBC). A sewage sample from Seongnam Water Reclamation Center (Gyeonggi, South Korea) was used as the source for the isolation of the phage. The sewage sample was centrifuged at 8000 rpm for 5 min and the supernatant was purified using a 0.22 μm filter system (Millipore, USA). 100 μL of the 24 h enrichment host culture and filtered samples were added to 5 mL of LBC soft agar (LBC broth with 0.8% agar) and overlaid on LBC agar. After incubation overnight at 37 °C, the number of plaques was counted and expressed as plaque-forming units per milliliter (PFU/mL), and a single plaque was selected and resuspended in 1 × SM buffer (100 mM NaCl, 10 mM MgSO4, 50 mM Tris–HCl, pH 7.5). Resuspended phage was filtered through a 0.22 μm filter to remove bacterial debris. Subsequently, isolated phage was stored at -80 °C in LBC broth containing 20% glycerol until further use [34, 11].
Morphological analysis of isolated phage
To visualize phage morphology, phage particles were concentrated in 20% polyethylene glycol (PEG) 8000 (Sigma, St Louis, MO, USA) and 2 M NaCl [4]. Concentrated phage particles (approximately 1010–1011 PFU/mL) were washed twice using 0.1 M ammonium acetate and suspended in 1x SM buffer. Partially purified phage was stained with 2% (w/v) uranyl acetate on a carbon-coated grid and then observed using a transmission electron microscopy (TEM) (H-7600, HITACHI, Tokyo, Japan) operated at 80 kV [4].
Host range determination
The lytic activity of the phage was tested against 35 E. coli strains by the spot method [33]. The E. coli strains were grown overnight in LBC broth and 100 μL of strain culture was added to 5 mL of LBC soft agar and overlaid on LBC agar. Then, 10 μL of diluted phage lysates (approximately 106 PFU/mL) were spotted onto the lawn of bacteria and incubated overnight at 37 °C. The lysis zone indicated the sensitivity of the bacteria to the phage.
One-step growth curve analysis
One-step growth curve analysis of the phage was performed with the host E. coli [34]. For adsorption, 1 mL of the diluted host and 1 mL of the diluted phage solution were inoculated in 8 mL of LBC broth and incubated at 37 °C for 10 min. Then, the suspension was centrifuged at 10,000×g for 10 min and infected cells were resuspended with fresh LBC broth. The samples were collected at intervals of 5 min while being incubated at 37 °C and phages were counted immediately by the plaque assay using a double layer agar method. The overlaid LBC agar plates were incubated at 37 °C overnight and the formed plaque was counted. Latent period and burst size were calculated from the one-step growth curve.
Phage genome sequencing and bioinformatic analysis
Genomic phage DNA extraction was carried out using a Phage DNA Isolation Kit (Norgen Biotek Corporation, Thorold, Ontario, Canada) as per the manufacturer’s instructions. The sequencing library was prepared by random fragmentation of the DNA sample, followed by 5′ and 3′ adapter ligation [25]. Sequencing was carried out at Macrogen Inc. (Seoul, Korea) using an Illumina Hiseq 2500 sequencer with paired-end reads. The reads were filtered using FastQC before assembly such that for a pair of PE reads, more than 90% of the bases had a base quality greater or equal to Q20 in each read (http://bioinformatics.babraham.ac.uk/projects/fastqc/). A single contig was assembled using the SOAPdenovo2 software Luo et al. [38]. The predictive open reading frames (ORFs) and possible tRNAs were predicted using Prokka [50]. NCBI BLAST was used for the identification of putative protein functions and homologous phage genomes based on the e-value (< 1E − 05) [5]. The genome sequence of vB_EcoM-ECP26 was deposited in the GenBank database under accession no. MK373780.1. Phylogenetic analysis of major capsid protein and DNA polymerase based on amino acid sequence between vB_EcoM-ECP26 and other Myoviridae phages was carried out using MEGA7 [32].
Thermal and pH stability of vB_EcoM-ECP26
The stability of the phage at high temperatures was analyzed by exposing the phage to heat for 30 min. The phage solution was exposed for 30 min to two different temperatures of 65 and 70 °C, and phage titer was determined using the spot assay on the lawn of LBC agar at 5 min intervals [12]. For the analysis of pH stability, the phage was incubated in 1x SM buffer adjusted to a pH of 2, 4, 7, or 10. The mixtures were incubated for 1 h at room temperature and the survival rate of each treated phage was determined using the spot assay on the lawn of LBC agar [29]. All overlaid plates were incubated at 37 °C overnight.
Phage control effect on E. coli O157:H7 in culture media
To evaluate the effect of the phage on the growth of E. coli O157:H7, we carried out a modified method of a previous report [46]. 10 μL of exponential host bacterial cultures of E. coli NCCP 13930 and 10 μL of vB_EcoM-ECP26 were inoculated in a 96-well plate with 180 μL of LBC broth. The mixtures were incubated at 37 °C for 10 h and optical density was measured at 600 nm using a spectrophotometer.
Evaluation of the antimicrobial effect of phage on fresh lettuce
Evaluation of the antimicrobial activity of the phage on fresh food was carried out using a modified method from previous reports [19, 48]. Romaine lettuce was purchased from a local grocery store and kept at 4 °C. The lettuce was rinsed with 70% ethanol and washed with distilled water twice, then the leaves were cut into pieces. The lettuce leaves were exposed to UV light in a laminar flow hood for 1 h. The lettuce leaves were then submerged into a 106 CFU/mL E. coli NCCP 13930 solution for 30 min. After 30 min, lettuce leaves were washed twice with distilled water to discard planktonic bacteria. The lettuce leaves were then submerged into a 106 PFU/mL vB_EcoM-ECP26 solution or in phosphate buffer solution (PBS) for 5 min. Phage solution or PBS was then washed off the lettuce and the leaves were dried for 30 min in a laminar flow hood. Lettuce leaves were stored in a plastic dish sealed with Petri film for up to 5 days at 4 °C. After the appropriate duration of time, 10 g of lettuce leaves were added to 90 mL of PBS in a sterile stomacher bag and processed using a stomacher for 5 min. The homogenate was centrifuged for 10 min at 8000 rpm, and cell pellets were resuspended in PBS. The bacteria were enumerated after serial dilutions and spread plated on SMAC agar (Oxoid, Basingstoke, Hampshire, UK) at 37 °C for 24 h. Colony counts were converted to log CFU/g. Supernatant was serially diluted and mixed with LBC soft agar with cultures of E. coli NCCP 13930. Soft agar was poured into LBC agar and plates were incubated at 37 °C for 24 h. Plaques resulting from phage lysis on the bacterial lawn were counted and titers were converted to log PFU/g.
Statistical analysis
The experiments were replicated three times and the experimental results are expressed as mean ± standard deviation (SD). Thermal and pH stability of phage were evaluated using Duncan’s multiple range test. The population of E. coli on the lettuce treated with the phage and the control were evaluated using a one-tailed t test. The data were analyzed using SPSS ver. 25 (SPSS Inc., Chicago, IL, USA).
Results and discussion
Isolation and morphological analysis of phage vB_EcoM-ECP26
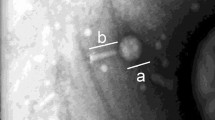
Escherichia coli specific phage vB_EcoM-ECP26 was isolated from a sewage sample. Phage vB_EcoM-ECP26 formed relatively clear and small (0.5–1 mm) plaques against E. coli O157:H7 NCCP 13930 (Fig. 1a). Transmission electron microscopy (TEM) analysis revealed that vB_EcoM-ECP26 belonged to the Myoviridae family with a contractile-tailed phage group (Fig. 1b). The phage head and tail size were approximately 72 nm and 104 nm, respectively. Plaque morphology could be a consequence of lytic activity [20]. Lytic phages typically produce clear plaques but temperate phages produce turbid plaques caused by their lysogenic ability [28]. In this light, phage vB_EcoM-ECP26 may be considered a lytic phage.

Transmission electron micrograph of plaque morphology (a) and Escherichia coli phage vB_EcoM-ECP26 (b)
Host range determination of vB_EcoM-ECP26
Since the host range of the phage may be a key factor for controlling pathogenic E. coli, we carried out a spot method assay with 35 E. coli strains. These E. coli strains were mostly STEC pathotypes (32/35). Phage vB_EcoM-ECP26 produced a lysis zone on 33 of the 35 (94.28%) strains tested (Table 1). Specifically, vB_EcoM-ECP26 formed lysis zone on all O157 strains. Bacteriophages might recognize host surface components using their receptor binding protein (RBP) [27]. Lipopolysaccharides (LPS) of the outer membrane in gram-negative bacteria is one of the well-known receptors of RBP. LPS can be classified into two types: the smooth type (S-type) which is the most common type and the rough type (R-type) with lipid A and a core, but no O-antigen [52]. Bacteriophages recognizing R-type LPS have a broad host range since the LPS core structure is conserved in various Gram-negative bacteria [49]. Moreover, many coliphages can recognize more than one surface receptor, such as OmpC [55]. Phage vB_EcoM-ECP26 was able to form lysis zones on E. coli NCCP 13893 (O antigen-negative) with a broad host spectrum. These results suggest that vB_EcoM-ECP26 may recognize R-type LPS and could be a potential biocontrol agent.
One-step growth curve analysis
The one-step growth curve analysis was used to assess vB_EcoM-ECP26 growth kinetics including the latent period and burst size. The calculated latent period was approximately 55 min and the burst size was 1914 PFU/cell (data not shown). Burst size is influenced by various factors such as host bacteria type, metabolism, and environmental conditions [2]. Burst size is an important factor of phage application since large burst size could contribute to the efficient propagation of the phage [6]. Therefore, the large burst size of the vB_EcoM-ECP26 could be an advantage when used as a biocontrol agent.
Bioinformatic analysis of vB_EcoM-ECP26 genome
The phage vB_EcoM-ECP26 genome consisted of 136,993 nucleotides with a mol % G + C content of 43.59. Analysis of the complete genome of phage vB_EcoM-ECP26 based on the query cover of NCBI BLASTn showed a high homology with V5-like phages. A maximum similarity of 92.68% was seen with Escherichia phage vB_EcoM_HdK5 (Genbank accession MK373780.1).
Phage vB_EcoM-ECP26 encoded 214 putative ORFs and five tRNAs. Among the predicted 214 ORFs, 59 (27.5%) ORFs revealed protein functions. The 59 ORFs were classified into four groups based on their functions: (i) structure (tail fiber, tail sheath, baseplate, tape measure, head, and structural proteins), (ii) replication and regulation (polymerase, helicase, anti-sigma factor, terminase, transcriptional regulator, nuclease, kinase, ligase), (iii) host lysis (cell wall hydrolase, spanins, and lysozyme), and (iv) additional functions (nucleotide metabolism, host recognition associated proteins). Functional ORFs, hypothetical ORF positions, homology, and accession number information are summarized in Additional file 1: Table S1. Notably, like other V5 like phages, none of the ORFs of vB_EcoM-ECP26 showed homology to the holin involved in permeabilizing the host cytoplasmic membrane [49]. However, a hypothetical gene (nt 88483-88689) which was located next to the endolysin gene might be a putative holin because of its high transmembrane probability and small size. In addition, genes associated with lysogeny such as integrase or anti-repressor and virulence-associated genes were not found. These results suggest a high possibility of applying vB_EcoM-ECP26 as a tool for controlling foodborne pathogenic E. coli, because the possibility of phage-mediated gene transfer was excluded.
Phylogenetic analysis of essential and conserved protein sequences has been used for demonstrating the evolutionary relationships between phages [21, 49]. To deduce the homology and evolutionary relationship between vB_EcoM-ECP26 and other Myoviridae phages, we analyzed their DNA polymerase and major capsid protein (MCP) amino acid sequences. According to the phylogenetic analysis, vB_EcoM-ECP26 and V5-like phages were contained in the same group (Fig. 2). These results suggest that vB_EcoM-ECP26 belongs to the group of V5-like species. In addition, V5-like species might be distinct from T4-like phages which are known as the representative E. coli infecting lytic phages.

Neighbor-joining phylogenetic tree analysis. Analysis was based on the amino acid sequence alignment of DNA polymerase (a) and major capsid protein (b) from Escherichia coli phage vB_EcoM-ECP26, and other Myoviridae phages using MEGA7 and ClustalW. Bootstrap analysis was performed with 1000 repetitions. The branch mean bootstrap value and the length of the branch is associated with the relative protein difference among phages
Thermal and pH stability of vB_EcoM-ECP26
To characterize phage vB_EcoM-ECP26, thermal and pH stability tests were performed. Heat treatment at 65 °C for 30 min did not significantly reduce the titer of the phage (p > 0.05). However at 70 °C, the phage showing reduction only 0.8 log PFU/mL after 5 min (p > 0.05) and lost activity after 10 min (p < 0.05) (Fig. 3a). Exposure of vB_EcoM-ECP26 to pH 4, 7, and 10 had no significant effect on titers, showing low reduction from 0.32 log PFU/mL to maximum log 0.6 PFU/mL (p > 0.05), but exposure to a pH 2 solution induced loss of phage activity (p < 0.05) (Fig. 3b). Kim et al. [29] reported that broad host range bacteriophages are relatively stable at pH 3–10 but unstable at 70 °C and Park et al. [43] reported that phage SFP10 was highly stable at pH 4–10, but completely inactivated under pH 2. These data reveal that vB_EcoM-ECP26 has a moderate heat and pH resistance. Therefore, the high tolerance to environmental stress of vB_EcoM-ECP26 would be advantageous for the application of biocontrol agents.

Thermal and pH stability test results. Thermal stability (a) and pH stability (b) of phage vB_EcoM-ECP26. Thermal stability tests were performed in 1x SM buffer. pH stability tests were performed for 1 h at room temperature (20 °C). The data are presented as mean ± SD (n = 3). The asterisk indicates significant differences (**p > 0.05; *p < 0.05). N/D not detected
Growth inhibition assay
To evaluate the ability of vB_EcoM-ECP26 to lyse host bacteria in culture media, we carried out a host growth inhibition assay. After infection of E. coli NCCP 13930 with vB_EcoM-ECP26 (MOI of 1), the host bacterial growth was inhibited after 1 h and this inhibition lasted for 8 h. The inhibition activity was also similar in the MOI 0.1 treatment group (Fig. 4a). These results show that phage infection with a MOI of 0.1 can effectively control E. coli O157:H7. However, after 10 h of infection, resistance was observed despite vB_EcoM-ECP26 being a lytic phage. This phenomenon might be due to the emergence of bacteriophage insensitive mutants (BIMs) by methods such as the restriction-modification system or CRISPR-Cas immunity [15, 39]. BIM also can induced by altering the LPS or cell envelope structures [41]. To overcome BIM emergence, phage cocktail techniques or a combination with other natural antibacterial agents could be used [16]. Endersen et al. [17] reported that a combined-phage preparation possessed a strong bactericidal effect on Cronobacter sakazakii. Therefore, for more efficient control of pathogenic E. coli, lytic phages with a broad host spectrum should be screened and used as a cocktail with vB_EcoM-ECP26.

Effect of vB_EcoM-ECP26 on growth of E. coli O157:H7. Growth curve of E. coli O157:H7 NCCP 13930 co-cultivated with phage vB_EcoM-ECP26 (a). Reduction of E. coli O157:H7 NCCP 13930 counts on romaine lettuce treated with vB_EcoM-ECP26 for 5 days (applied 106 PFU/mL) (b). P.C: 1x SM buffer treatment; T: Phage treatment; P: Phage titer in romaine lettuce. The data are presented as mean ± SD (n = 3). The asterisk indicates significant differences (*p < 0.05). N/D not detected
Evaluation of the phage antimicrobial effect on romaine lettuce
The effectiveness of the phage vB_EcoM-ECP26 against STEC was measured on romaine lettuce for 5 days. When bacteria inoculated lettuce leaves were exposed to the phage solution for 5 min, the reduction activity was 0.9 log CFU/g compared to the PBS (p < 0.05) (Fig. 4b). Storage of the phage-treated romaine leaves at 4 °C revealed a time-dependent E. coli O157:H7 decrease (Fig. 4b). On day 1, the E. coli O157:H7 population showed a non-significant decrease of 0.3 log CFU/g compared to day 0 (p > 0.05). However, on day 3, E. coli O157:H7 populations showed a significant decrease of 1.2 log CFU/g compared to day 0 (p < 0.05). On day 5, E. coli O157:H7 populations were reduced to undetectable levels in phage-treated lettuce. Moreover, we observed phage persistence on lettuce coupons at 4 °C without phage treatment. Phage populations were maintained in the lettuce and were in the range of 4.2–4.8 log PFU/g for 5 days (p > 0.05) (Fig. 4b). These data showed that phage vB_EcoM-ECP26 not only could control the STEC present in fresh foods but also could maintain its activity at low temperatures. Generally, E. coli could survive at low temperatures but could not grow, so its phage could not propagate at low temperatures. Nevertheless, the phage could lyse bacteria because of its lysis from without (LO) mechanism [1]. LO has been most studied in T-even phages where LO is mediated by the tail-associated lysozyme of the phage [7]. Ferguson et al. [19] reported that the LO of the E. coli phage cocktail could control the E. coli O157:H7 contaminated fresh-cut lettuce at low temperatures. Oliveira et al. [42] also revealed that fresh foods contaminated with Listeria monocytogenes could be controlled at low temperatures through a phage cocktail, and that phage persistence was maintained at low temperatures. The most crucial factors in the application of bacteriophage in fresh food are its stability and lytic ability in food. Therefore, it could be concluded that the lytic phage vB_EcoM-ECP26 is a novel phage which remains stable and does not lose its lytic activity on fresh food at low temperatures.
Availability of data and materials
The dataset supporting the conclusion of the research are included within this manuscript.
References
Abedon ST (2011) Lysis from without. Bacteriophage 1:46–49
Abedon ST, Herschler TD, Stopar D (2001) bacteriophage latent-period evolution as a response to resource availability. Appl Environ Microbiol 67:4233–4241
Abuladze T, Li M, Menetrez MY, Dean T, Senecal A, Sulakvelidze A (2008) Bacteriophages reduce experimental contamination of hard surfaces, tomato, spinach, broccoli, and ground beef by Escherichia coli O157:H7. Appl Environ Microbiol 74:6230–6238
Ackermann H-W (2009) Basic phage electron microscopy. In: Clokie MRJ, Kropinski AM (eds) Bacteriophages: Methods and Protocols, vol 1. Isolation, Characterization, and Interactions. Humana Press, Totowa, pp 113–126
Altschul SF et al (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res 25:3389–3402
Amarillas L, Rubi-Rangel L, Chaidez C, Gonzalez-Robles A, Lightbourn-Rojas L, Leon-Felix J (2017) Isolation and characterization of phiLLS, a novel phage with potential biocontrol agent against multidrug-resistant Escherichia coli. Front Microbiol 8:1355
Arisaka F, Kanamaru S, Leiman P, Rossmann MG (2003) The tail lysozyme complex of bacteriophage T4. Int J Biochem Cell Biol 35:16–21
Askar M et al (2011) Update on the ongoing outbreak of haemolytic uraemic syndrome due to Shiga toxin-producing Escherichia coli (STEC) serotype O104, Germany, May 2011. Eurosurveillance 16:19883
Bardasi L, Taddei R, Nocera L, Ricchi M, Merialdi G (2015) Shiga toxin-producing Escherichia coli in meat and vegetable products in Emilia Romagna Region, Years 2012–2013. Ital J Food Saf 4:4511
Bolocan AS, Callanan J, Forde A, Ross P, Hill C (2016) Phage therapy targeting Escherichia coli—a story with no end? FEMS Microbiol Lett. https://doi.org/10.1093/femsle/fnw256
Bonilla N, Rojas MI, Netto Flores Cruz G, Hung SH, Rohwer F, Barr JJ (2016) Phage on tap-a quick and efficient protocol for the preparation of bacteriophage laboratory stocks. PeerJ 4:e2261
Capra ML, Binetti AG, Mercanti DJ, Quiberoni A, Reinheimer JA (2009) Diversity among Lactobacillus paracasei phages isolated from a probiotic dairy product plant. J Appl Microbiol 107:1350–1357
Colomer-Lluch M, Jofre J, Muniesa M (2011) Antibiotic resistance genes in the bacteriophage DNA fraction of environmental samples. PLoS ONE 6:e17549
Dedrick RM et al (2017) Prophage-mediated defence against viral attack and viral counter-defence. Nat Microbiol 2:16251
Deng K, Fang W, Zheng B, Miao S, Huo G (2018) Phenotypic, fermentation characterization, and resistance mechanism analysis of bacteriophage-resistant mutants of Lactobacillus delbrueckii ssp. bulgaricus isolated from traditional Chinese dairy products. J Dairy Sci 101:1901–1914
Drulis-Kawa Z, Majkowska-Skrobek G, Maciejewska B, Delattre A-S, Lavigne R (2012) Learning from bacteriophages–advantages and limitations of phage and phage-encoded protein applications. Curr Protein Pept Sci 13:699–722
Endersen L et al (2017) Investigating the biocontrol and anti-biofilm potential of a three phage cocktail against Cronobacter sakazakii in different brands of infant formula. Int J Food Microbiol 253:1–11
Endersen L, O’Mahony J, Hill C, Ross RP, McAuliffe O, Coffey A (2014) Phage therapy in the food industry. Annu Rev Food Sci Technol 5:327–349
Ferguson S, Roberts C, Handy E, Sharma M (2013) Lytic bacteriophages reduce Escherichia coli O157. Bacteriophage 3:e24323
Gallet R, Kannoly S, Wang I-N (2011) Effects of bacteriophage traits on plaque formation. BMC Microbiol 11:181
Glazko G, Makarenkov V, Liu J, Mushegian A (2007) Evolutionary history of bacteriophages with double-stranded DNA genomes. Biolo Direct 2:36
Gobin M et al. (2018) National outbreak of Shiga toxin-producing Escherichia coli O157:H7 linked to mixed salad leaves, United Kingdom, 2016. Euro Surveill 23
Guilhelmelli F, Vilela N, Albuquerque P, Derengowski L, Silva-Pereira I, Kyaw C (2013) Antibiotic development challenges: the various mechanisms of action of antimicrobial peptides and of bacterial resistance. Front Microbiol 4:353
He L et al (2019) Intestinal probiotics E. coli Nissle 1917 as a targeted vehicle for delivery of p53 and Tum-5 to solid tumors for cancer therapy. J Biol Eng 13:58
Head SR et al (2014) Library construction for next-generation sequencing: overviews and challenges. Biotechniques 56:61–64
Herman KM, Hall AJ, Gould LH (2015) Outbreaks attributed to fresh leafy vegetables, United States, 1973–2012. Epidemiol Infect 143:3011–3021
Javed MA, Poshtiban S, Arutyunov D, Evoy S, Szymanski CM (2013) Bacteriophage receptor binding protein based assays for the simultaneous detection of Campylobacter jejuni and Campylobacter coli. PLoS ONE 8:e69770
Jurczak-Kurek A et al (2016) Biodiversity of bacteriophages: morphological and biological properties of a large group of phages isolated from urban sewage. Sci Reports 6:34338
Kim EJ, Chang HJ, Kwak S, Park JH (2016) Virulence factors and stability of coliphages specific to Escherichia coli O157:H7 and to various E. coli infection. J Microbiol Biotechnol 26:2060–2065
Kim YG, Lee JH, Kim SI, Baek KH, Lee J (2015) Cinnamon bark oil and its components inhibit biofilm formation and toxin production. Int J Food Microbiol 195:30–39
Krüger A, Lucchesi PMA (2015) Shiga toxins and stx phages: highly diverse entities. Microbiology 161:451–462
Kumar S, Stecher G, Tamura K (2016) MEGA7: molecular evolutionary genetics analysis Version 7.0 for bigger datasets. Mol Biol Evol 33:1870–1874
Kutter E (2009) Phage host range and efficiency of plating. In: Clokie MRJ, Kropinski AM (eds) Bacteriophages: Methods and Protocols, vol 1. Isolation, Characterization, and Interactions. Humana Press, Totowa, pp 141–149
Lee YD, Kim JY, Park JH (2013) Characteristics of coliphage ECP4 and potential use as a sanitizing agent for biocontrol of Escherichia coli O157:H7. Food Control 34:255–260
Liao YT, Miller MF, Loneragan GH, Brooks JC, Echeverry A, Brashears MM (2014) Non-O157 shiga toxin-producing Escherichia coli in US. retail ground beef. J Food Prot 77:1188–1192
Lim TH et al (2012) Use of bacteriophage for biological control of Salmonella Enteritidis infection in chicken. Res Vet Sci 93:1173–1178
Lin DM, Koskella B, Lin HC (2017) Phage therapy: an alternative to antibiotics in the age of multi-drug resistance. World J Gastrointest Pharmacol Ther 8:162–173
Luo R et al. (2012) SOAPdenovo2: an empirically improved memory-efficient short-read de novo assembler. GigaScience 1::2047
Marraffini LA, Sontheimer EJ (2010) Self versus non-self discrimination during CRISPR RNA-directed immunity. Nature 463:568–571
Meng X et al (2011) Application of a bacteriophage lysin to disrupt biofilms formed by the animal pathogen Streptococcus suis. Appl Environ Microbiol 77:8272–8279
Mizoguchi K, Morita M, Fischer CR, Yoichi M, Tanji Y, Unno H (2003) Coevolution of bacteriophage PP01 and Escherichia coli O157:H7 in continuous culture. Appl Environ Microbiol 69:170–176
Oliveira M, Vinas I, Colas P, Anguera M, Usall J, Abadias M (2014) Effectiveness of a bacteriophage in reducing Listeria monocytogenes on fresh-cut fruits and fruit juices. Food Microbiol 38:137–142
Park M et al (2012) Characterization and comparative genomic analysis of a novel bacteriophage, SFP10, simultaneously inhibiting both Salmonella enterica and Escherichia coli O157:H7. Appl Environ Microbiol 78:58–69
Percival SL, Williams DW (2014) Escherichia coli. In: Microbiology of Waterborne Diseases, pp 89-117
Persad AK, LeJeune JT (2014) Animal Reservoirs of Shiga Toxin-Producing Escherichia coli. Microbiology Spectrum 2
Raya RR et al (2006) Isolation and characterization of a new T-Even bacteriophage, CEV1, and determination of its potential to reduce Escherichia coli O157:H7 levels in sheep. Appl Environ Microbiol 72:6405–6410
Sadekuzzaman M, Mizan MFR, Yang S, Kim HS, Ha SD (2018) Application of bacteriophages for the inactivation of Salmonella spp. in biofilms. Food Sci Technol Int 24:424–433
Sadekuzzaman M, Yang S, Mizan MFR, Ha SD (2017) Reduction of Escherichia coli O157:H7 in biofilms using bacteriophage BPECO 19. J Food Sci 82:1433–1442
Santos SB et al (2011) Genomic and proteomic characterization of the broad-host-range Salmonella phage PVP-SE1: creation of a new phage genus. J Virol 85:11265–11273
Seemann T (2014) Prokka: rapid prokaryotic genome annotation. Bioinformatics 30:2068–2069
Soni KA, Nannapaneni R, Hagens S (2010) Reduction of Listeria monocytogenes on the surface of fresh channel catfish fillets by bacteriophage listex P100. Foodborne Pathog Dis 7:427–434
Steimle A, Autenrieth IB, Frick J-S (2016) Structure and function: lipid a modifications in commensals and pathogens. Int J Med Microbiol 306:290–301
Sukumaran AT, Nannapaneni R, Kiess A, Sharma CS (2015) Reduction of Salmonella on chicken breast fillets stored under aerobic or modified atmosphere packaging by the application of lytic bacteriophage preparation SalmoFreshTM1,2. Poult Sci 95:668–675
Touchon M, Bernheim A, Rocha EPC (2016) Genetic and life-history traits associated with the distribution of prophages in bacteria. ISME J 10:2744–2754
Washizaki A, Yonesaki T, Otsuka Y (2016) Characterization of the interactions between Escherichia coli receptors, LPS and OmpC, and bacteriophage T4 long tail fibers. Microbiol Open 5:1003–1015
Acknowledgements
This research was supported by the Korea Institute of Planning and Evolution for Biotechnology of Food, Agriculture, Forestry and Fisheries funded by the Korean Government (117060033HD020).
Funding
Korea Institute of Planning and Evolution for Biotechnology of Food, Agriculture, Forestry and Fisheries funded by the Korean Government (117060033HD020).
Author information
Authors and Affiliations
Contributions
JHP designed the experiments, reviewed the results, and supervised the whole project. DWP and DYL conducted the experiments and processed the data. DWP and YDL wrote the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Park, DW., Lim, Gy., Lee, Yd. et al. Characteristics of lytic phage vB_EcoM-ECP26 and reduction of shiga-toxin producing Escherichia coli on produce romaine. Appl Biol Chem 63, 19 (2020). https://doi.org/10.1186/s13765-020-00502-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13765-020-00502-4