
Abstract
The microbiota is indispensable for human health and the regulation of various body functions, including energy metabolism. The harmonic crosstalk between the microbiota and the intestinal epithelial barrier determines gut homeostasis and health status in the healthy subject. Obesity and type 2 diabetes risk are, to some extent, explained by alterations in the microbiota. Since recent data indicate that the population of gut microorganisms can influence nutrient absorption and energy storage thus prevalence on obesity and metabolic disorders. Moreover, metabolic disease conditions, such as obesity, may be stimulated by genetic, environmental factors and by pathways that link metabolism with the immune system. On the basis of the above considerations, this review compiles the current results obtained in recent studies indicating the gut microbiota contribution to obesity development.
Similar content being viewed by others
Introduction
The prevalence of obesity and diabetes in developed societies remains as one of the main public health problems worldwide and its incidence is extensively increasing. Obesity results from the accumulation of excess adipose tissue; however, its etiology is complex, and symptoms are heterogeneous due to high comorbidity with metabolic diseases, including type 2 diabetes, which is a concomitant pathology. It has been suggested that diet is important in energy balance, and also plays a fundamental role in maintaining the diversity and proper functioning of our gut microbiota. Thus appropriate dietary intervention, such as high-fiber diet may improve health status via regulation of microbiota in humans [1]. In this context, microbiota manipulation by prebiotics becomes a possible modifier of the microbial profile and can improve host health, by triggering beneficial systemic responses and reducing adiposity [2].
Microbiota composition in the human gut
The microorganism’s community inhabited in human body are known as the microbiota. The microbial cell number concentration in the lumen is ten times greater than the host eukaryotic cells, representing 1–2 kg of body weight [3]. The gut microbiota has been classified into approximately 1000 different species [4]. Three distinctive dominant phylum has been identified in the human intestine: Firmicutes, Bacteroidetes and Actinobacteria [5], which account for approximately 90% of total bacteria in the gut, mainly in its terminal part, the colon. The diverse biochemical activity of these bacteria have vital metabolic contributions for the human body, in which the body could not perform independently [6].
Recent gut microbiota studies have indicated a greater Firmicutes to Bacteroidetes (F/B) ratio in obese individuals and obese (ob/ob) mice compared to those of lean counterparts [7,8,9,10,11]. For instance, the study of Ley et al., was the first to exhibit powerful correlation between gut microbiota and obesity in leptin-deficient ob/ob mice, which is a mouse model for obesity. Results showed different gut microbiota composition in ob/ob mice compared to those of the lean wild-type mice (+/+), reporting a higher ratio of Firmicutes instead of Bacteroidetes [8]. Although the bacterial composition of the microbiota varies from one healthy subject to another, the functional gene profile associated with each microbiota tends to be similar [11]. Consequently, the microbiota strongly affects the intestinal genes expression involving energy balance [12], the regulation of intestinal barrier function [13], intestinal satietogenic hormones release stimulation [14], the bile acids metabolic activity modulation [15], the digestion/absorption of nutrients by intestinal mucosa of the host [16], and the generation of short-chain fatty acids (SCFAs) via non-digestible carbohydrates bacterial fermentation, in which these SCFAs along with plant polysaccharides absorbed by the host are transported to the liver, where they are converted to more complex lipids [15,16,17,18], contributing significantly to human physiology and metabolism.
Likewise, the microbiota participates in the homeostasis of individuals, providing a series of key functions such as the dietary non-digestible polysaccharides degradation, regulation of energy storage obtained, synthesis of vitamins, modulation of the immune system and also has a protective effect of the intestinal barrier [13]. Hence, any factor or events that affect one of these functions has repercussions on others and negatively impacts the entire organism. Taken together, this article aims to compile scientific literature on the relevance of microbiota to obesity and the possible mechanisms involved preventing or treating the obesity pandemic.
Altered gut microbiota profile in obesity
Dysbiosis, alterations in gut microbiota composition, may lead to the obesity development, thus, emphasizing that gut-nutrients-targeting linked to obesity amelioration should be recognized [19]. Obesity and diabetes, are consequences of intricate interaction between several genes and environmental factors, involving energy imbalance due to a sedentary lifestyle, excessive energy consumption, or both [20, 21]. The recent rising-interest in gut microbiota role as a potential target for the rapid escalation of obesity incidence globally [22, 23]. Several studies have reported that the intestinal microbiota differs among individuals, depending on adiposity and body mass index (BMI). Microbial composition changes in diabetic and metabolic syndrome animal models were observed [11]. Reinforcing the possible association of microbial composition variation to metabolic disorders. In addition, several human studies have also indicated the correlation of microbiota composition changes in the obese population. A schematic summary of gut microbiota population, altered in obesity and lean individuals, is shown in Fig. 1. Alike, consistent with results from animal studies, common gut microbiota changes in obese subjects seems to be linked to an increased and decreased abundance of Firmicutes and Bacteroidetes, respectively [24]. Table 1 summarizes key human studies on gut microbiota role in obesity development.

Summary of gut microbiota composition increase in obese and lean subjects
Evidence suggests that dietary intervention could ameliorate obesity by altering microbiota profile. Reportedly, green tea and its processed products such as black along with oolong tea, promotes beneficial lipid metabolism and obesity effects [25]. In line with these findings, Seo et al. showed exhibitory anti-obesity effects of fermented green tea extracts (FGT; 500 mg/kg; dissolved in 0.1% methylcellulose) in mice, in which high-fat diet (HFD) fed obese mice were given a daily oral administration of dried green tea leaves, fermented by Bacillus subtilis, while the control groups were administered 0.1% methylcellulose as vehicle. Results indicated 8-week of FGT administration dramatically decreased the Firmicutes/Bacteroidetes ratio while lowering plasma glucose and lipid levels. Moreover, the dietary intervention reduced lipogenic and pro-inflammatory gene expression, thereby preventing hyperlipidemia [26, 27]. Taken together, it is suggested that gut microbiota modification through dietary intervention might be a promising obesity therapeutic adjuvant.
Microbiota, obesity, and prebiotics correlation
Studies in animal models of genetic or diet-induced obesity showed that prebiotics can enhance intestinal barrier function, glucose tolerance, and lipid metabolism modulation while reducing weight gain, fat mass, and inflammatory status. Likewise, it can regulate the production of intestinal hormones by the trophic effect that promote the mucosa [21, 28,29,30]. Table 2 shows the summary of mice and human key prebiotic studies.
Furthermore, studies in mice have shown the relationship between energy balance, diet, and gut microbiota composition. Intestinal microbiota has been suggested to be affected by nutrient acquisition, energy storage, and a host of metabolic pathways [5]. The microbiota has dramatic biological effects in the human organism; in terms of nutrient acquisition, the microbiota metabolizes non-digestible food residues, such as dietary polysaccharides (e.g., oligosaccharides, resistant starch and fructose). Consequently, intestinal microbiota metabolic activities differences may lead to variations in ingested calories from dietary substances, storage of calories in adipose tissue, and the availability of energy for microbial proliferation. Such intestinal microbiota differences are also linked to variation in energy uptake in humans, explaining aspects of obesity which may be responsible for an individual’s predisposition to metabolic disorders [24].
Most microbiota composition alterations are reversible suggesting microbiota of an individual is an innate characteristic [20]. However, these alterations in obese subjects (genetic or diet induced) can be reversed by oral transfer of the lean mice intestinal microbiota [9, 10] or by dietary intervention of prebiotics administration [31]. Inflammation, diabetes, obesity, and insulin resistance in mice can be improved by fecal transplantation [21].
Only in animals, modulation of microbiota compositions may improve body weight. Obese animals to germ-free mice (GFM) and lean animals microbiota transplantation results in obesity, while the contrary is observed by transfer microbiota from lean to obese animals [11, 32, 33]. For instance, Turnbaugh et al. demonstrated that fecal transplantation of ob/ob into GFM led to increased body weight over a 2-week period [10]. Likewise, Goodman et al. carried out a study using mice that were transplanted with human microbiota from lean individuals and fed a high-fat-high-sugar diet, resulting in a Firmicutes increase and a reduction in Bacteroidetes group attributable to bacterial fermentation [34]. In line with these findings, Bäckhed et al. showed that GFM C57BL/6 following a colonization by conventional mice distal microbiota, increases 60% BMI along with insulin resistance after 14 days while reducing food consumption, revealed that gut microbiota promotes absorption of monosaccharides, resulting in de novo hepatic lipogenesis induction [20].
Prebiotics are ingredients selectively fermented to promote beneficial modification in gastrointestinal microbiota’s composition and/or activity and thus are capable of conferring health benefits to the individuals [28, 35, 36]. They are usually “non-digestible” dietary elements, but are fermented by gut microbiota, serving as energy source [37, 38]. The effects of prebiotics on energy homeostasis and satiety regulation are linked to a decrease in metabolic disorders and obesity incidence [39, 40] (Fig. 2). Although exact gut microbiota and obesity correlations are not fully known, diet-based intestinal microbiota manipulations, particularly via prebiotics and dietary fibers, can be a potential approach for reversing or obesity prevention.

Prebiotic supplementation correlation with gut microbiota and its effects on obesity and metabolic syndrome
Short-chain fatty acids production via microbiota
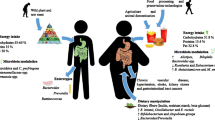
Recently, dietary fibers have gain interest due to their exert beneficial metabolic functions including gut microbiota SCFAs production. SCFAs affects the growth and differentiation of enterocytes and colonocytes. The energy extracted from dietary fibers by microbiota, become available to the body and avoids their loss in stools. The fermentation of dietary fibers by the microbiota releases SCFAs such as acetate, propionate and butyrate, whose total concentrations can reach 130 mM in the colon [41, 42]. While butyrate is metabolized principally by colonocytes; acetate and propionate are absorbed, reaching concentrations of 300 to 450 μM in portal blood and 50 and 100 μM in peripheral blood [16, 20, 43]. Acetate is the dominant type of SCFAs in humans and these SCFAs seems to play an interesting role in modulation of protein kinase activity (PKA) activated by 5′-AMP-activated protein kinase (AMPK) along with macrophage infiltration into adipose tissue [12].
In contrast, the de novo synthesis of lipids or glucose is triggered by propionate, acting as an energy source for the host. Moreover, butyrate and propionate initiate intestinal gluconeogenesis via a gut-brain axis, promoting metabolic advantages on glucose and body weight regulation [43]. SCFAs can function as signals derived from microbes that influence carbohydrate metabolism and intestinal physiology by stimulating the secretion of hormones such as ghrelin, and the greater peptide tyrosine tyrosine (PYY) and glucagon-like peptide-1 (GLP-1) release, while serving as intestinal epithelial cells energy source (Fig. 3). Acetate, in particular, represents a preferred substrate for gluconeogenesis and the synthesis of cholesterol and triglycerides [28, 41, 44].

Interaction between SCFAs, GPR43 and GPR41 receptors. (1) Bacteria in the gut interact with complex dietary carbohydrates produced by SCFAs through hydrolysis and fermentation; (2) The binding between SCFAs and GPR43 receptor in enterocytes promotes the release of GLP-1, increasing insulin sensitivity; (3) GPR43 activation via SCFAs interactions in the adipocyte suppresses insulin release, inhibiting fat stores in adipose tissue; (4) SCFAs-GPR41 receptor binding increases the release of PYY, reducing the development of obesity
In addition, from their role in energy recovery, the SCFAs are ligand compounds for G protein coupled free fatty acid receptor 2 (FFAR2/GPR43) and 3 (FFAR3/GPR41). Acetate selectively activates GPR43 in vitro; propionate exerts a similar activation on GPR43 and GPR41; however, butyrate has an affinity-activation over GPR41 [45]. These receptors are implicated in appetite and energy metabolism regulation that affect insulin resistance and adiposity in individuals. According to Gao et al. butyrate supplementation to DIO mice raised intestinal and circulating SCFAs resulting in weight gain suppression, AMPK activation and increase mitochondrial function. These results indicated that feeding a HFD along with butyrate supplementation to diet-induced obese mice, reverses and prevents insulin resistance [46].
Alternatively, GPR43 stimulation promotes energy storage by increasing adipogenesis, inhibiting lipolysis in adipocytes, and decreasing energy expenditure [44]. Propionate, modulates energy homeostasis by promoting the activation of sympathetic neurons mediated GPR41, in contrast to ketone bodies [44]. In addition, compared to wild types, knockout animals for GPR41 or GPR43 have less hepatic triglycerides and plasma cholesterol, greater insulin sensitivity and a lower weight gain when fed on a HFD [47]. Thus, gut epithelium or liver greatly uses SCFAs as an energy source to confer metabolic benefits to the host, such as suppression of food intake and stimulation of gut hormone secretion while protecting against glucose intolerance and high-fat-induced weight gain.
In the intestinal lumen, SCFAs binds to GPR41, increasing key insulin signaling molecules, such as PYY, which delays gut motility, thus increasing nutrient absorption [48] and increase GLP-1 levels, regulating satiety [18, 43, 49]. Similarly, GPR43 role in regulating inflammatory responses by microbiota modulation have been studied [5, 16]. These results indicate that GPR41 and GPR43 are important for gut immunity. Furthermore, based on studies SCFAs are precursors of hepatic cholesterol biosynthesis and fatty acids. For instance, according to Canfora et al. propionate acts as de novo gluconeogenesis precursor and attenuates lipogenesis in the liver through the suppression of fatty acid synthesis (FAS) while butyrate and acetate directly activates hepatic AMPK-phosphorylation via peroxisome proliferator-activated receptor (PPAR-α) target genes upregulation. Thus, elevating fatty acid oxidation (FAO) and glycogen storage, presumably to be triggered by GPR41/GPR43-dependent mechanisms [50]. Consistently, according to Brown et al., the GPR43 highest concentrations were found in immune cells while GPR41 was highly expressed in adipose tissue [45]. GPR43 stimulation of has been shown to promote energy storage by increasing adipogenesis, inhibiting lipolysis in adipocytes and decreasing energy expenditure hereby regulating energy metabolism [51]. Hence GPR43 function as a sensor for excessive dietary energy in adipose tissue by modulating metabolic homeostasis [52]. Taken together, SCFAs-GPRs signaling pathways is involved in lipid, glucose along with cholesterol metabolism regulation.
Prebiotics effects on metabolic syndrome and obesity
The consumption of oligofructose has been correlated with blood glucose and fasting insulin improvement in diabetic rats and appetite inhibition in HFD-fed animals [53]. Clinical trials of healthy subjects have confirmed the findings in animals, showing that the consumption of oligofructose modulates ghrelin, GLP-1 and PYY plasma concentrations in humans, decreasing the postprandial blood glucose changes and the sensation of hunger in these subjects [18, 54]. As reported by Cani et al. the oligofructose supplementation effect on human satiety hormones was assessed; 10 adults (5 men and 5 women) randomly placed into groups receiving either 16 g oligofructose/day or 16 g dextrin-maltose/day for 2 weeks. The results showed that the incorporation of prebiotics into the diet may be an interesting strategy to control appetite by modulating the microbiota, since GLP-1 was remarkably higher after prebiotic treatment compared to the controls. In prebiotic-treated subjects, PYY levels were notably elevated after 10 min [18]. Similarly, Delzenne et al. showed that the administration of a diet supplemented with 10% dietary inulin-type fructans extracted from chicory root to male Wistar rats for 3 weeks, decreases the food consumption and the epididymal fat mass of animals due to the decrease in the release of ghrelin, and the greater release of GLP-1 and PYY via enteroendocrine cells of ileal and colonic epithelium [55].
Furthermore, it has been suggested that prebiotics could exert anti-obesity effects through the reduction of adipogenesis. Dewulf et al. observed that 4-week dietary supplementation of obese mice with inulin-type fructan (0.2 g/day/mouse) reduced GPR43 mRNA expression in subcutaneous adipose tissue, decreased induced-HFD fat mass development while it increased lipolysis, improving tissue insulin response. Subsequently, modulated peroxisome proliferator-activated receptor (PPAR-γ) activation and reduction in the expression of CCAAT-enhancer-binding protein (C/EBPα), lipoprotein lipase (LPL), and fatty acid binding protein (FABP/ap2). These proteins are involved in the processes of lipid accumulation and adipocyte differentiation in adipogenesis and, therefore, presented a positive correlation with the reduction of adipocytes size and fat mass. Hence, the administration of inulin-type fructan to HFD-fed mice led to an important increase of Bifidobacteria levels and counteracted all the HFD-induced alterations, including a restoration of PPAR-γ activity and GPR43 expression. However, the specific modulation of the gut microbiota in which inulin counteract HFD-induced PPAR-γ remains elusive [51].
Neyrinck et al., also demonstrated that mice under a 4-week HFD supplementation with prebiotic arabinoxylan (10% w/w), showed lower FAS uptake due to decreased fatty acid synthase and LPL activity. Also, reduced body weight gain reduction, serum and hepatic cholesterol accumulation and insulin resistance index, while restoring the number of bacteria (Bacteroides-Prevotella spp. and Roseburia spp.) and prompting bifidobacteria that were decreased upon HFD-fed [19]. Arabinoxylan supplementation leads to modifications of fatty acid pattern in the adipose tissue by increasing the rumenic acid amount, a linoleic acid metabolite which belong to the conjugated linoleic acid (CLA) family. CLA isomers have been shown to exert a variety of biological effects, including anti-obesity effects [56]. Arabinoxylan hypocholesterolemic effects were previously demonstrated in rats, by decreasing dietary cholesterol absorption and an increasing fecal excretion of cholesterol and bile acids, while increasing the 3-hydroxy-3-methyl-glutaryl-coenzyme A reductase (HMG-CoA reductase) expression [19, 57]. In addition, Abrams et al. carried out a 1-year study in 97 adolescents that were supplemented with 8 g/day inulin-type fructan and noted that subjects receiving the prebiotic had smaller BMI increase and fat mass index (FMI) compared to the controls (8 g/day maltodextrin) [17].
In spite of these evidences, Neyrinck et al. showed that the 4-week administration of 10% chitin-β-glucan has no effect on GLP-1 plasma levels and proglucagon expression in obese mice, suggesting that in contrast to oligofructose, chitin-β-glucan effect on obesity is independent of GLP-1 production and Bifidobacteria changes. It was postulated that chitin-β-glucan affect host lipid metabolism via gut microbiota modulation. Regardless of those findings, there was a significant body weight and fat mass reduction, while having metabolic profile improvements such as decreasing fasting hyperglycemia, hepatic triglyceride accumulation and hypercholesterolemia [58]. Alternatively, galacto-oligosaccharide supplementation (5.5 g/day for 12 weeks) did not affect the anthropometric parameters of individuals with metabolic syndrome and overweight, however, it did alter some markers related to this disorder such as a reduction in plasma total cholesterol, triglycerides, insulin, and C-reactive protein concentrations. It improved intestinal immune function assessed by an increase in IgA secretion and decreased calprotectin (intestinal inflammatory marker) in feces. Suggesting that galacto-oligosaccharide positively influences the immune response by shifting from less beneficial bacteria to more beneficial bifidobacteria numbers in the fecal microbiota [59].
Other positive results of prebiotic ingestion in obesity in humans have been reported by Genta et al. in which overweight and mild dyslipidemia women underwent a weight loss program for 120 days (hypocaloric diet + physical activity); those who included yacon syrup, rich in fructo-oligosaccharides (FOS) to their diet (enough to provide 0.14 g of FOS/kg of body weight/day) had a greater reduction in body weight, BMI and waist circumference. Regarding the biochemical variables, this group of women presented a reduction in the values of HOMA-IR, fasting insulin and low-density lipoprotein cholesterol. The authors emphasize that these results cannot be attributed exclusively to FOS supplementation, however, the beneficial effect was advised [60]. Despite prebiotics appear to be promising tools in the nutritional strategy for reducing the risk of obesity or as an adjuvant in the treatment of this clinical condition through their capacity to promote secretion of endogenous gastrointestinal hormones involved in appetite regulation. Further human studies are required to confirm the dietary fibers effects in reducing adiposity as well as obesity associated comorbidities.
Availability of data and materials
Not applicable.
Abbreviations
- AMPK:
-
5′-adenosine monophosphate-activated protein kinase
- BMI:
-
body mass index
- C/EBPα:
-
CCAAT/enhancer-binding protein alpha
- CLA:
-
conjugated linoleic acid
- FABP/ap2:
-
fatty acid binding protein/adipocyte protein 2
- FAO:
-
fatty acid oxidation
- FFAR2/GPR43:
-
free fatty acid receptor 2
- FFAR3/GPR41:
-
free fatty acid receptor 3
- FGT:
-
fermented green tea extracts
- FMI:
-
fat mass index
- FOS:
-
fructo-oligosaccharides
- GFM:
-
germ-free mice
- GLP-1:
-
glucagon-like peptide-1
- HMG-CoA reductase:
-
3-hydroxy-3-methyl-glutaryl-coenzyme A reductase
- HOMA-IR:
-
homeostatic model assessment of insulin resistance
- LDL-C:
-
low-density lipoprotein cholesterol
- LPL:
-
lipoprotein lipase
- PKA:
-
protein kinase activity
- PPAR-α:
-
peroxisome proliferator-activated receptor alpha
- PPAR-γ:
-
peroxisome proliferator-activated receptor
- PYY:
-
peptide tyrosine tyrosine
- SCFAs:
-
short-chain fatty acids
- TC:
-
total cholesterol
References
Frazier TH, DiBaise JK, McClain CJ (2011) Gut microbiota, intestinal permeability, obesity-induced inflammation, and liver injury. JPEN J Parenter Enteral Nutr 35(5suppl):14S–20S
Qin J, Li R, Raes J, Arumugam M, Solvsten Burgdorf K, Manichanh C et al (2010) A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464:59
Shanahan F (2011) Translating the microbiota to medicine. Nat Rev Gastroenterol Hepatol 9:72–74
Geurts L, Lazarevic V, Derrien M, Everard A, Van Roye M, Knauf et al (2011) Altered gut microbiota and endocannabinoid system tone in obese and diabetic leptin-resistant mice: impact on apelin regulation in adipose tissue. Front Microbiol 2:149
Nicholson JK, Holmes E, Kinross J, Burcelin R, Gibson G, Jia W et al (2012) Host-gut microbiota metabolic interactions. Science 336(6086):1262–1267
Neish AS (2009) Microbes in gastrointestinal health and disease. Gastroenterology 136(1):65–80
Benjamin S, Spener F (2009) Conjugated linoleic acids as functional food: an insight into their health benefits. Nutr Metab 6:36
Bell DS (2015) Changes seen in gut bacteria content and distribution with obesity: causation or association? Postgrad Med J 127(8):863–868
Turnbaugh PJ, Backhed F, Fulton L, Gordon JI (2008) Marked alterations in the distal gut microbiome linked to diet-induced obesity. Cell Host Microbe 3(4):213–223
Turnbaugh PJ, Ley RE, Mahowald MA, Magrini V, Mardis ER et al (2006) An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 444:1027–1031
Vijay-Kumar M, Aitken JD, Carvalho FA, Cullender TC, Mwangi S, Srinivasan S et al (2010) Metabolic syndrome and altered gut microbiota in mice lacking Toll-like receptor 5. Science 328(5975):228–231
Carvalho BM, Guadagnini D, Tsukumo DM, Schenka AA, Latuf-Filho P, Vassallo J et al (2012) Modulation of gut microbiota by antibiotics improves insulin signalling in high-fat fed mice. Diabetologia 55(10):2823–2834
Cho I, Blaser MJ (2012) The Human Microbiome: at the interface of health and disease. Nat Rev Genet 13(4):260–270
Lim SM, Jeong JJ, Woo KH, Han MJ, Kim DH (2016) Lactobacillus sakei OK67 ameliorates high-fat diet–induced blood glucose intolerance and obesity in mice by inhibiting gut microbiota lipopolysaccharide production and inducing colon tight junction protein expression. Nutr Res 36(4):337–348
Everard A, Lazarevic V, Gaïa N, Johansson M, Ståhlman M, Backhed F et al (2014) Microbiome of prebiotic-treated mice reveals novel targets involved in host response during obesity. ISME J 8:2116
Galvez J, Rodríguez-Cabezas ME, Zarzuelo A (2005) Effects of dietary fiber on inflammatory bowel disease. Mol Nutr Food Res 49(6):601–608
Abrams SA, Griffin IJ, Hawthorne KM, Ellis KJ (2007) Effect of prebiotic supplementation and calcium intake on body mass index. J Pediatr 151:293–298
Cani PD, Lecourt E, Dewulf EM, Sohet FM, Pachikian BD, Naslain D et al (2009) Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am J Clin Nutr 90(5):1236–1243
Neyrinck AM, Possemiers S, Druart C, Van de Wiele T, De Backer F, Cani PD et al (2011) Prebiotic effects of wheat arabinoxylan related to the increase in bifidobacteria, Roseburia and Bacteroides/Prevotella in diet-induced obese mice. PLoS ONE 6(6):e20944
Bäckhed F, Ding H, Wang T, Hooper LV, Koh GY, Nagy A et al (2004) The gut microbiota as an environmental factor that regulates fat storage. Proc Natl Acad Sci USA 101(44):15718–15723
Burcelin R, Serino M, Chabo C, Blasco-Baque V, Amar J (2011) Gut microbiota and diabetes: from pathogenesis to therapeutic perspective. Acta Diabetol 48(4):257–273
Harris K, Kassis A, Major G (2012) Chou CJ (2012) Is the gut microbiota a new factor contributing to obesity and its metabolic disorders? J Obes 879151:1–14
Tehrani AB, Nezami BG, Gewirtz A, Srinivasan S (2012) Obesity and its associated disease: a role for microbiota? J Neurogastroenterol Motil 24(4):305–311
Kallus SJ, Brandt LJ (2012) The intestinal microbiota and obesity. J Clin Gastroenterol 46(1):16–24
Kim JH, Jia Y, Lee JG, Nam B, Lee JH, Shin KS et al (2014) Hypolipidemic and antiinflammation activities of fermented soybean fibers from meju in C57BL/6J mice. Phytother Res 28(9):1335–1341
Carmody RN, Gerber GK, Luevano JM Jr, Gatti DM, Somes L, Svenson KL et al (2015) Diet dominates host genotype in shaping the murine gut microbiota. Cell Host Microbe 17(1):72–84
Seo DB, Jeong HW, Cho D, Lee BJ, Lee JH, Choi JY et al (2015) Fermented green tea extract alleviates obesity and related complications and alters gut microbiota composition in diet-induced obese mice. J Med Food 18(5):549–556
Festi D, Schiumerini R, Eusebi LH, Marasco G, Taddia M, Colecchia A (2014) Gut microbiota and metabolic syndrome. World J. Gastroentero 20(43):16079–16094
Lesniewska V, Rowland I, Cani PD, Neyrinck AM, Delzenne NM, Naughton PJ (2006) Effect on components of the intestinal microflora and plasma neuropeptide levels of feeding Lactobacillus delbrueckii, Bifidobacterium lactis, and inulin to adult and elderly rats. Appl Environ Microbiol 72(10):6533–6538
Macpherson AJ, Hunziker L, McCoy K, Lamarre A (2001) IgA responses in the intestinal mucosa against pathogenic and non-pathogenic microorganisms. Microb Infect 3(12):1021–1035
Cani PD, Neyrinck AM, Fava F, Knauf C, Burcelin RG, Tuohy KM et al (2007) Selective increases of bifidobacteria in gut microflora improve high-fat-diet-induced diabetes in mice through a mechanism associated with endotoxaemia. Diabetologia 1 50(11):2374–2383
Kulecka M, Paziewska A, Zeber-Lubecka N, Ambrozkiewicz F, Kopczynski M, Kuklinska U et al (2016) Prolonged transfer of feces from the lean mice modulates gut microbiota in obese mice. Nutr Metab 13(1):57
da Silva ST, dos Santos CA, Bressan J (2013) Intestinal microbiota; relevance to obesity and modulation by prebiotics and probiotics. Nutr Hosp 28(4):1039–1048
Goodman AL, Kallstrom G, Faith JJ, Reyes A, Moore A, Dantas G et al (2011) Extensive personal human gut microbiota culture collections characterized and manipulated in gnotobiotic mice. Proc Natl Acad Sci USA 108(15):6252–6257
Clarke SF, Murphy EF, Nilaweera K, Ross PR, Shanahan F, O’Toole PW et al (2012) The gut microbiota and its relationship to diet and obesity: new insights. Gut Microbes 3(3):186–202
Gibson GR, Scott KP, Rastall RA, Tuohy KM, Hotchkiss A, Dubert-Ferrandon A et al (2010) Dietary prebiotics: current status and new definition. Food Sci Technol Bull Funct Foods 7(1):1–9
Quigley EM (2010) Prebiotics and probiotics; modifying and mining the microbiota. Pharmacol Res 61(3):213–218
Slavin J (2013) Fiber and prebiotics: mechanisms and health benefits. Nutrients 5(4):1417–1435
Roberfroid M, Gibson GR, Hoyles L, McCartney AL, Rastall R, Rowland I et al (2010) Prebiotic effects: metabolic and health benefits. Br J Nutr 104(S2):S1–S63
Ley RE, Bäckhed F, Turnbaugh P, Lozupone CA, Knight RD, Gordon JI (2006) Obesity alters gut microbial ecology. Proc Natl Acad Sci USA 102(31):11070–11075
Cummings J, Pomare EW, Branch WJ, Naylor CP, Macfarlane GT (1987) Short chain fatty acids in human large intestine, portal, hepatic and venous blood. Gut 28(10):1221–1227
Wong JM, De Souza R, Kendall CW, Emam A, Jenkins DJ (2006) Colonic health: fermentation and short chain fatty acids. J Clin Gastroenterol 40(3):235–243
Kasubuchi M, Hasegawa S, Hiramatsu T, Ichimura A, Kimura I (2015) Dietary gut microbial metabolites, short-chain fatty acids, and host metabolic regulation. Nutrients 7(4):2839–2849
Kimura I, Inoue D, Maeda T, Hara T, Ichimura A, Miyauchi S et al (2011) Short-chain fatty acids and ketones directly regulate sympathetic nervous system via G protein-coupled receptor 41 (GPR41). Proc Natl Acad Sci USA 108(19):8030–8035
Brown AJ, Goldsworthy SM, Barnes AA, Eilert MM, Tcheang L, Daniels D et al (2003) The Orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J Biol Chem 278(13):11312–11319
Gao Z, Yin J, Zhang J, Ward RE, Martin RJ, Lefevre M et al (2009) Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 58(7):1509–1517
Samuel BS, Shaito A, Motoike T, Rey FE, Backhed F, Manchester JK et al (2008) Effects of the gut microbiota on host adiposity are modulated by the short-chain fatty-acid binding G protein-coupled receptor, Gpr41. Proc Natl Acad Sci USA 105(43):16767–16772
Baothman OA, Zamzami MA, Taher I, Abubaker J, Abu-Farha M (2016) The role of gut microbiota in the development of obesity and diabetes. Lipids Health Dis 15(1):108
Conterno L, Fava F, Viola R, Tuohy KM (2011) Obesity and the gut microbiota: does up-regulating colonic fermentation protect against obesity and metabolic disease? Genes Nutr 6(3):241–260
Canfora EE, Jocken JW, Blaak EE (2015) Short-chain fatty acids in control of body weight and insulin sensitivity. Nat Rev Endocrinol 10:577
Dewulf EM, Cani PD, Neyrinck AM, Possemiers S, Van Holle A, Muccioli GG et al (2011) Inulin-type fructans with prebiotic properties counteract GPR43 overexpression and PPARγ-related adipogenesis in the white adipose tissue of high-fat diet-fed mice. J Nutr Biochem 22(8):712–722
Kimura I, Ozawa K, Inoue D, Imamura T, Kimura K, Maeda T et al (2013) The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat Commun 7(4):1829
Cani PD, Neyrinck AM, Maton N, Delzenne NM (2005) Oligofructose promotes satiety in rats fed a high-fat diet: involvement of glucagon-like peptide-1. Obes Res 13(6):1000–1007
Parnell JA, Reimer RA (2009) Weight loss during oligofructose supplementation is associated with decreased ghrelin and increased peptide YY in overweight and obese adults. Am J Clin Nutr 89(6):1751–1759
Delzenne NM, Cani PD, Daubioul C, Neyrinck AM (2007) Impact of inulin and oligofructose on gastrointestinal peptides. Br J Nutr 93(S1):S157–S161
Santacruz A, Collado MC, Garcia-Valdes L, Segura MT, Martin-Lagos JA, Anjos T et al (2010) Gut microbiota composition is associated with body weight, weight gain and biochemical parameters in pregnant women. Br J Nutr 104(1):83–92
Vrieze A, Van Nood E, Holleman F, Salojärvi J, Kootte RS, Bartelsman JF et al (2012) Transfer of intestinal microbiota from lean donors increases insulin sensitivity in individuals with metabolic syndrome. Gastroenterology 143(4):913–916
Neyrinck AM, Possemiers S, Verstraete W, De Backer F, Cani PD, Delzenne NM (2012) Dietary modulation of clostridial cluster XIVa gut bacteria (Roseburia spp.) by chitin–glucan fiber improves host metabolic alterations induced by high-fat diet in mice. J Nutr Biochem 23(1):51–59
Vulevic J, Juric A, Tzortzis G, Gibson GR (2013) A mixture of trans-galactooligosaccharides reduces markers of metabolic syndrome and modulates the fecal microbiota and immune function of overweight adults. J Nutr 143(3):324–331
Genta S, Cabrera W, Habib N, Pons J, Carillo IM, Grau A et al (2009) Yacon syrup: beneficial effects on obesity and insulin resistance in humans. Clin Nutr 28(2):182–187
Cotillard A, Kennedy SP, Kong LC, Prifti E, Pons N, Le Chatelier E et al (2013) Dietary intervention impact on gut microbial gene richness. Nature 500(7464):585
Cox LM, Cho I, Young SA, Anderson WK, Waters BJ, Hung SC et al (2013) The nonfermentable dietary fiber hydroxypropyl methylcellulose modulates intestinal microbiota. FASEB J 27(2):692–702
Damms-Machado A, Mitra S, Schollenberger AE, Kramer KM, Meile T, Königsrainer A et al (2015) Effects of surgical and dietary weight loss therapy for obesity on gut microbiota composition and nutrient absorption. Biomed Res Int 2015:806248
Eslinger AJ, Eller LK, Reimer RA (2014) Yellow pea fiber improves glycemia and reduces Clostridium leptum in diet-induced obese rats. Nutr Res 34(8):714–722
Hamilton MK, Ronveaux CC, Rust BM, Newman JW, Hawley M, Barile D et al (2017) Prebiotic milk oligosaccharides prevent development of obese phenotype, impairment of gut permeability, and microbial dysbiosis in high fat-fed mice. Am J Physiol Gastrointest Liver Physiol 312(5):G474–G487
Hoang MH, Houng SJ, Jun HJ, Lee JH, Choi JW, Kim SH et al (2011) Barley intake induces bile acid excretion by reduced expression of intestinal ASBT and NPC1L1 in C57BL/6J mice. J Agric Food Chem 59(12):6798–6805
Hou YP, He QQ, Ouyang HM, Peng HS, Wang Q, Li J et al (2017) Human gut microbiota associated with obesity in Chinese children and adolescents. Biomed Res Int 2017:7585989
Hume MP, Nicolucci AC, Reimer RA (2017) Prebiotic supplementation improves appetite control in children with overweight and obesity: a randomized controlled trial. Am J Clin Nutr 105(4):790–799
Krupa-Kozak U, Markiewicz L, Lamparski G, Juśkiewicz J (2017) Administration of inulin-supplemented gluten-free diet modified calcium absorption and caecal microbiota in rats in a calcium-dependent manner. Nutr J 9(7):702
Reimer R, Maathuis A, Venema K, Lyon M, Gahler R, Wood S (2014) Effect of the novel polysaccharide PolyGlycopleX® on short-chain fatty acid production in a computer-controlled in vitro model of the human large intestine. Nutr J 6(3):1115–1127
Xiao S, Fei N, Pang X, Shen J, Wang L, Zhang B et al (2014) A gut microbiota-targeted dietary intervention for amelioration of chronic inflammation underlying metabolic syndrome. FEMS Microbiol Ecol 87(2):357–367
Zhang C, Yin A, Li H, Wang R, Wu G, Shen J et al (2015) Dietary modulation of gut microbiota contributes to alleviation of both genetic and simple obesity in children. EBioMedicine 2(8):968–984
Acknowledgements
Not applicable.
Funding
This work was carried out with the support of the ‘Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ011253042018)’, Rural Development Administration (RDA), Republic of Korea and by the National Research Foundation of Korea (NRF) grant funded by the Korea government (MSIP) (NRF-2018R1A4A1022589).
Author information
Authors and Affiliations
Contributions
ARP and SJL wrote the manuscript. Both authors read and approved the manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Rivera-Piza, A., Lee, SJ. Effects of dietary fibers and prebiotics in adiposity regulation via modulation of gut microbiota. Appl Biol Chem 63, 2 (2020). https://doi.org/10.1186/s13765-019-0482-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13765-019-0482-9