
Abstract
Introduction
The brown planthopper (BPH), Nilaparvata lugens (Stål) (Hemiptera: Delphacidae), is a notorious insect pest of rice. Nutrient supplementation can alter plant biochemical compositions which may indirectly affect on ecological fitness parameters of its insect pest. However, few findings are available on nutrient-induced ecological fitness of BPH and relationship of BPH fitness parameters with its host rice plant biochemical contents.
Methods
We studied the main and interaction effects of nitrogen (N), phosphorus (P), and potassium (K) inputs on BPH fitness, as measured by the following variables: nymph survival, nymphal duration, adult body weight, and its survival. Brown planthopper fitness parameters were regressed as function of rice plant (Oryza sativa) biochemical composition. A completely randomized design with four replications in a factorial scheme was used considering N, P, and K levels as factors.
Results
Nitrogen application to the host rice plants provided greater survival and enhanced body weight of BPH. The nymphal duration was found to decrease with N subsidy that caused shorter generation time of BPH. Nitrogenous compounds N and soluble protein (SP), total free sugar (TFS), and silicon (Si) content in rice plant tissue correlated strongly with all fitness traits of BPH. Nitrogen, SP, and TFS associated positively with BPH survival and body weight, while negatively with nymphal duration. In contrast, Si had negative correlation with BPH survival and body weight, but positive with nymphal duration. Phosphorus supplementation to host plant contributed to increase BPH body weight. Relationships of P with all parameters of BPH were weak except body weight which was highly significant and positive. Interaction between N and P inputs demonstrated significant effect on BPH body weight. Application of K had no significant effect on BPH survival and development. Regression analysis did not detect significant relationship of BPH fitness parameters with plant tissue content of K.
Conclusions
The results suggest that BPH ecological fitness characters were improved after N and P fertilization to rice plants which are associated with biochemical content of rice plant. Therefore, N and P inputs should be used judiciously in rice cultivation to keep BPH ecological fitness potential at minimum level.
Similar content being viewed by others


Explore related subjects
Find the latest articles, discoveries, and news in related topics.Introduction
The brown planthopper (BPH), Nilaparvata lugens (Stål) (Hemiptera: Delphacidae), was a relatively minor pest of rice (Oryza sativa) prior to the advent of high-input rice farming. However, it dramatically came out as a major insect pest of high-yielding varieties (HYVs) of rice (Bottrell and Schoenly 2012). Planthopper outbreaks in rice fields have intensified in the last decade resulting in heavy rice yield losses (Chakravarthy et al. 2013; Sogawa 2015).
Population abundance and dynamics of an insect pest is dependent on its important ecological fitness parameters such as higher survival rate, faster development, and increased body size. Ecological fitness is the measure of how well an individual or a population adapts to a specific niche. Ecological fitness of an insect may vary depending on how it interacts individually or as a community with the surrounding environment or ecosystem (Lu and Heong 2009). Host plant quality plays a major role in the population development of planthopper by influencing performance and fitness (Cook and Denno 1994). Factors such as habitats, morphological, and chemical components (sugar, nitrogen, amino acids, and semiochemicals) in host plants can alter fitness parameters of insects (Sogawa 1982; Fischer and Fiedler 2000). Organisms are likely to be more fit when the elemental composition of available food is in balance with their requirement (Boersma and Elser 2006). Nutrient supplementation to rice plants enhanced BPH fitness and increased starvation tolerance through the improvement of its biological and ecological parameters (Lu and Heong 2009).
Nitrogen (N) is regarded as an indicator of plant quality (Lu et al. 2007), and its limitation is widely studied for different insects. The herbivores on N-enriched plants showed higher survival rate (Prestidge 1982; Wier and Boethel 1995; Huberty and Denno 2006; Lu and Heong 2009) with bigger body sizes (Kaneshiro and Johnson 1996; Jauset et al. 2000; Huberty and Denno 2006), shorter developmental time (Fischer and Fiedler 2000; Huberty and Denno 2006), fewer instars (Wier and Boethel 1995), and higher fecundity (Rashid et al. 2016b). Nitrogen supplementation causes vigorous plant growth (Rashid et al. 2016a) that may enhance ecological fitness potential of BPH through improvement of insect microecology.
Phosphorus (P) limitation is poorly studied for herbivores. In studies that have investigated the effect of P on insect, there is evidence showing that it is an important determinant of survivorship and body size of invertebrates (Huberty and Denno 2006). Phosphorus may be a much more important component for phytophagous insects because it is required for RNA synthesis and can impose severe consequences for the growth rate of consumers when it is limiting (Sterner and Elser 2002). Phosphorus limitation has been documented widely in many species of aquatic invertebrates, particularly in Daphnia (Urabe et al. 1997; Elser et al. 2001). Aquatic and terrestrial insects face similar stoichiometric mismatches for both N and P content in their body compared to their food resources (Elser et al. 2000). Therefore, P availability may be an important factor for growth and development as well as ecological fitness parameters of terrestrial insect like BPH through improvement of nutritional ecology.
Potassium (K) supplementation alters host plant quality for insects (Armengaud et al. 2009). It is an essential nutrient that affects most of the biochemical and physiological processes. It influences plant growth and metabolism (Wang et al. 2013). High levels of K in plants are associated with reduced carbohydrate accumulation and elimination of amino acids (Baskaran et al. 1985), higher silica content, and increase in the sclerenchymous layer (Dale 1988). It regulates the metabolite pattern of higher plants and changes metabolite concentrations in plant tissues (Marschner 2012). The primary metabolites such as soluble sugars, particularly reducing sugars, organic acids, amino acids, and amides tend to increase in K-deficient plants (Amtmann et al. 2008). These nutrient sources may affect on ecological fitness parameters of BPH.
Several studies pointed out that high doses of N use might be an important cause of N. lugens outbreak as secondary pest (Lu and Heong 2009; Bottrell and Schoenly 2012). In the earlier studies, the nutrient variable used in most experiments was the amount of nutrients applied rather than tissue nutrient content. In this paper, we investigated the main and interaction effects of major plant nutrient—N, P, and K inputs on BPH survival, nymphal duration, and body weight, and regressed those BPH fitness parameters as a function of plant tissue nutrients. We tested the hypothesis that BPH survival, developmental duration, and adult body weight differ with nutrient supplementation to rice plants which might contribute to their ecological fitness potentials.
Methods
Culture of brown planthopper
Brown planthopper population was mass cultured according to Rashid et al. (2016a) with the susceptible variety BR3. Forty five to 60-day-old potted rice plants were cleaned and outer leaf sheaths were removed. Gravid BPH females were released in a cage with those rice plants for oviposition and removed 24 h later with the help of an aspirator. The plants with BPH eggs were shifted every day to a series of rearing cages for egg hatching. Then, host plants in the cage were changed at 3–4 days interval to provide sufficient food for the development of the nymphs to adulthood. This cycle was maintained to obtain required number of BPH as and when necessary for the experiments.
Experimental design and nutrient treatments
Fifteen-day-old rice seedlings of variety BR3 were transplanted (three hills/pot and two seedlings/hill) in each pot and exposed to three levels of each nutrient: nitrogen (0, 100, 200 kg N ha−1), phosphorous (0, 20, 40 kg P2O5 ha−1) potassium (0, 60, 120 kg K2O ha−1), and combinations of all the levels. Thus, 27 different nutrient treatments were used to determine their effect on survival and development of BPH. Experiments were set up as a completely randomized design in a factorial scheme (N, P, and K rates as factors) and replicated four times considering each pot as a replication. Therefore, 108 pots (16 cm high × 14 cm diameter) were used for each experiment. Two kilograms of dry soil was taken in each pot. The soil consisted of clay loam, with pH = 5.3, organic C = 0.72%, total N = 0.11%, available P = 6.60 ppm, exchangeable K = 0.09 meq 100 g−1 soil, and available Si = 0.21%. The exact amount of fertilizer inputs to be applied to each pot was calculated based on the amount of soil in each pot to retain fixed content in each treatment according to Asher et al. (2002) considering 1 ha cultivated field contains 2 × 106 kg soil in its root zone. Half of N fertilizer (in the form of urea), whole of P (in the form of triple super phosphate), and K (in the form of muriate of potash) were mixed in the soil. The remaining half of N was applied when the plants were 30 days old. The macronutrient treatments are here after referred to as N0, N100, and N200 for nitrogen, P0, P20, and P40 for phosphorus, and K0, K60, and K120 for potassium. Plants with different nutrient levels were labeled and placed under natural condition. Regular watering and weeding were done in the pots to allow sufficient growth of plants.
Determination of chemical composition of rice plant
Effect of N, P, and K inputs on biochemical content of rice plant was determined from four randomly chosen pots per treatment which were never exposed to BPH. For nutrient determination plants were harvested and oven dried at 70 °C ± 5 °C for 72 h. Plant samples were flushed with tap water to get rid of soil and were oven dried at 70 °C ± 5 °C for 72 h. Dried plant samples were ground in a Wiley mill. The resulting sample powder was used in determination of N, P, K, Si, and total free sugars (TFS) content in rice plant by the volumetric micro Kjeldahl distillation, colorimetric ternary mixture digestion, flame photometric HCl extraction, gravimetric dry ashing, and anthrone method, respectively (Yoshida et al. 1976). Soluble protein (SP) content in rice plants was analyzed according to Lowry’s method (Lowry et al. 1951).
Effect of nutrient levels on fitness of brown planthopper
Forty five-day-old rice plants (var. BR3) grown in pots with different nutrient combinations were trimmed to six tillers and were covered with transparent Mylar’s cage. Effects of nutrient inputs on BPH nymph survival and development, and adult survival rate were studied separately.
Brown planthopper nymph survival and development
Nymph survival rate, time required to complete nymphal period, and body weight of freshly developed adult on each nutrient level were investigated. Twenty newly hatched first instar BPH nymphs collected from the culture of each nutrient treatment were introduced into each Mylar’s cage with 45-day-old rice plants of same nutrient level. Observations on surviving nymphs and adults were recorded daily after release. Nymph survival rate was calculated using nymphs that reached adult stage and total number of nymphs. Nymph developmental duration was computed by the days from newly hatched nymphs to production of newly molted adults. Fresh body weight of ten adult females raised on rice plants with each nutrient treatment was measured using an electric balance.
Brown planthopper adult survival
Five pairs of newly molted adults from each BPH culture reared under each nutrient level were introduced into each cage with 45-day-old rice plants grown with same level of nutrient. The numbers of surviving adults were recorded on 5 and 7 days after release.
Statistical analyses
All data were tested for normality and homogeneity of variance, and appropriate transformations (arcsine or square root) were applied where these conditions were not met before analysis. However, untransformed data are presented in the figures. We performed three-way ANOVA, followed by Tukey’s multiple range test (p = 0.05) for mean comparisons in order to determine the effects of N, P, and K inputs on BPH nymph survival, nymphal duration, body weight of adult, and its survival (SPSS 2007). We also focused on how the biochemical contents (N, P, K, Si, TFS, and SP) in rice plant tissues changed after application of N, P, and K inputs to reflect more accurately what the nutrient levels were in the plants after uptake. Linear regression analysis was conducted to examine the effect of independent variables of N, P, K, Si, TFS, and SP content in rice plant tissue on the dependent variables BPH nymph survival, nymphal duration, adult body weight, and its survivability (SPSS 2007).
Results
Brown planthopper nymph survival and development
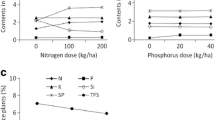
Brown planthopper nymph survival rate increased significantly with the increase in N application to rice plants (F 2, 81 = 437.38, p < 0.001, Fig. 1a). Significant difference was found in nymph survival of BPH reared on N100 and N200 plants (Fig. 1a). There was only marginal increase in survival rate of BPH due to P application, but the difference was insignificant (p > 0.05, Table 1, Fig. 1a). The nymph survival rate of BPH was statistically similar among different K treatment levels (F 2, 81 = 1.21, p = 0.30, Fig. 1a). No interactions of N and P, P and K, N and K, and all three nutrients (N, P, and K) on BPH nymph survival were detected (p > 0.05, Table 1). Brown planthopper nymph developmental duration decreased significantly with the N application to rice plants (F 2, 81 = 106.56, p < 0.001, Fig. 1b). Brown planthopper nymph feeding on N-enriched rice plant reduced its developmental time by about 2–3 days compared to those reared on N0 plants (Fig. 1b). Its nymphal period did not differ significantly due to P and K supplementation (p > 0.05, Table 1, Fig. 1b). Similarly, no significant difference was detected for BPH nymphal duration when the interactions of different nutrients were considered (p > 0.05, Table 1).

Ecological fitness parameter of Nilaparvata lugens reared on rice plants exposed to three levels of N (N0, N100, and N200), P (P0, P20, and P40), and K (K0, K60, and K120) fertilization in nethouse. a Nymph survival rate (%), b nymphal duration (days), c adult body weight (mg), d adult survival rate at 5 days after infestation (DAI), and e adult survival rate at 7 DAI
Fresh weight of N. lugens adult female increased markedly with the increase of N application to rice plants (F 2, 81 = 501.56, p < 0.001, Fig. 1c). There was significant difference in fresh body weight of BPH reared on N200 and N100 plants (Fig. 1c). Phosphorus fertilization to rice plants also significantly increased fresh body weight of N. lugens (F 2, 81 = 49.83, p < 0.001, Fig. 1c). However, there was no significant difference in BPH body weight between P20 and P40 treatment levels (Fig. 1c). Brown planthopper body weights did not differ with K fertilization (F 2, 81 = 0.21, p = 0.81, Fig. 1c). The interaction of N and P (F 2, 81 = 2.60, p < 0.049) was significant on body weight of adult BPH, whereas no interactions of N and K, P and K, and all three nutrients (N, P, and K) were detected (p > 0.05, Table 1).
Brown planthopper adult survival
Survival of adult brown planthopper BPH followed same trend both at 5th and 7th days after infestation. It increased significantly with the increase of N fertilization to its host rice plants (p < 0.001, Fig. 1d, e). There was significant difference in survival of BPH reared on N200 and N100 plants in both the occasions (Fig. 1d, e). Adult survival rate did not differ due to P and K fertilization to rice plants (p > 0.05, Fig. 1d, e). In similar way, no interactions of N and P, P and K, N and K, and all three nutrients (N, P, and K) on adult survival were detected (p > 0.05, Table 1).
Relationships between fitness parameters of BPH and biochemical contents in rice plant
Relationships of BPH survival rate, nymphal duration, and fresh body weight with plant tissue nutrients—N, total free sugars, soluble proteins, and Si were fitted to the linear models (Figs. 2, 3, 4, and 5). Highly significant and positive correlations were found to exists for BPH nymph survival rate with plant tissue nutrient concentrations of N (r 2 = 0.712; F 1,25 = 62.014; p < 0.001), total free sugars (r 2 = 0.591; F 1,25 = 36.218; p < 0.001), and soluble proteins (r 2 = 0.731; F 1,25 = 68.313; p < 0.001) (Fig. 2). Likewise, adult survival rate at 5 days after infestation (DAI) had strong positive correlation with N (r 2 = 0.59; F 1,25 = 36.011; p < 0.001), total free sugars (r 2 = 0.407; F 1,25 = 12.207; p < 0.001), and soluble proteins (r 2 = 0.676; F 1,25 = 52.368; p < 0.001) (Fig. 3). Similarly, adult survival at 7 DAI had also positive association with N (r 2 = 0.68; F 1,25 = 53.114; p < 0.001), total free sugars (r 2 = 0.301; F 1,25 = 10.79; p = 0.003), and soluble proteins (r 2 = 0.723; F 1,25 = 65.334; p < 0.001) (Fig. 3). Moreover, fresh body weight of BPH adult was correlated positively with plant tissue nutrients of N (r 2 = 0.783; F 1,25 = 90.211; p < 0.001), total free sugars (r 2 = 0.527; F 1,25 = 27.918; p < 0.001), and soluble proteins (r 2 = 0.784; F 1,25 = 90.885; p < 0.001) (Fig. 5). Host plant tissue P content had positive but weak (p > 0.05) relationship with both nymph and adult survival (Figs. 2a and 3b), while it was strongly associated with BPH body weight (r 2 = 0.311; F 1,25 = 11.396; p = 0.002) (Fig. 5). However, BPH survival rate and its fresh body weight were not associated significantly with the content of K (p > 0.05, Figs. 2a, 3c, and 5a). On the other hand, Si had highly significant but negative correlations with survival rate of nymph (r 2 = 0.645; F 1,25 = 45.602; p < 0.001, Fig. 2a) and adult (Fig. 5d) both at 5 DAI (r 2 = 0.51; F 1,25 = 26.068; p < 0.001) and 7 DAI (r 2 = 0.712; F 1,25 = 61.859; p < 0.001). Likewise, negative correlation was found for body weight of adult BPH (r 2 = 0.527; F 1,25 = 27.85; p < 0.001) with plant tissue content of Si (Fig. 4a). Brown planthopper nymphal duration had strong but negative relationship with N (r 2 = 0.649; F 1,25 = 46.246; p < 0.001), total free sugars (r 2 = 0.528; F 1,25 = 28.127; p < 0.001), and soluble proteins (r 2 = 0.753; F 1,25 = 76.57; p < 0.001) but not with K (r 2 = 0.026; F 1,25 = 0.67; p = 0.421) (Fig. 3). On the contrary, it had positive association with Si (r 2 = 0.607; F 1,25 = 38.637; p < 0.001) (Fig. 3). Relationship between nymphal duration and P was negative but weak (r 2 = 0.131; F 1,25 = 3.781; p = 0.063).

Relationship of Nilaparvata lugens nymph survival rate with plant tissue nutrient concentration. a nitrogen (N), phosphorus (P), potassium (K), and silicon (Si), b soluble protein (SP), and total free sugar (TFS) content in rice plant (n = 27)

Relationship of Nilaparvata lugens adult survival rate at 5 days after infestation (DAI) and 7 DAI with plant tissue nutrient concentration. a nitrogen (N), b phosphorus (P), c potassium (K), d silicon (Si), e total free sugar (TFS), and f soluble protein content in rice plant (n = 27)

Relationship of Nilaparvata lugens nymphal duration with plant tissue nutrient concentration. a nitrogen (N), phosphorus (P), potassium (K), and silicon (Si), b soluble protein (SP), and total free sugar (TFS) content in rice plant (n = 27)

Relationship of Nilaparvata lugens adult body weight with plant tissue nutrient concentration. a nitrogen (N), phosphorus (P), potassium (K), and silicon (Si), b soluble protein (SP), and total free sugar (TFS) content in rice plant (n = 27)
Discussion
Brown planthopper is the most prominent insect pest of rice. Its outbreaks in rice fields have intensified in the last decade, due to disruption of ecological balance (Sogawa 2015). Nutrition management is one of the most important practices for high production system of rice and consequently affects host-insect interaction by altering their ecology (Lu and Heong 2009). Population development of an insect pest in a particular agroecology largely depends on ecological fitness potential of the insect (Lu and Heong 2009). The characters used in the present study for measurement of ecological fitness of BPH are survival rates, development duration, and body weight under different nutrient regimes with rice variety BR3. In this section, we discussed how different nutrient treatment combinations influenced plant biochemical compositions and in turn affected the ecological fitness traits of BPH. Nutrient application to any plant results in higher nutrient content in their tissues by enhancing nutrient uptake through their roots (Rashid et al. 2016a).
Nutrient application changes the host-plant morphology, biochemistry, and physiology, which improves nutritional conditions for herbivores (Bernays 1990), thus playing a key role on insects. Nitrogen (N) content is one of the most important performance limiting factors of herbivores (De Bruyn et al. 2002; Facknath and Lalljee 2005; Kerpel et al. 2006; Lu et al. 2007; Lu and Heong 2009; Han et al. 2014; Oliveira et al. 2014; Rashid et al. 2016a, b, 2017). In the present study, survival rate of both nymph and adult BPHs was markedly higher on N-enriched plant than N-deficient one, and it was almost double and triple for nymph and adult respectively. This study confirms that BPH survival rate is enhanced with N supplementation to rice plants as reported earlier (Preap et al. 2001; Lu et al. 2004; Lu and Heong 2009). Sucker insect derives soluble protein, i.e., amino acids from their host plants through sucking for body maintenance (Slansky and Scriber 1985). Application of N to the rice plant results in significant increase in N and soluble protein (SP) content in plant tissues (Rashid et al. 2016a), from which BPH could ingest more amino acids through feeding compared to N-limited plants. Moreover, concentration of non-nitrogenous compounds, such as silicon (Si) content in plant tissues could also be reduced by N supplementation (Rashid et al. 2016a) or N-enriched plants might have lower accumulation of plant allelochemicals which can be less toxic to herbivorous insects. All these contributed to increased survival of BPH when reared on N-enriched host plant. Some reports showed that N is essential for insects mainly for its role in amino acid production and protein synthesis (Sterner and Elser 2002; Schoonhoven et al. 2005). On the other hand, many studies have shown that Si could enhance plant resistance to herbivores (Kvedaras and Keeping 2007; Reynolds et al. 2009; Kvedaras et al. 2010; Han et al. 2015; Nikpay et al. 2015). Furthermore, regression analysis in the present study clearly indicated that survival of BPH was correlated positively with plant tissue content of N, total free sugar (TFS), and SP but negatively with Si. Brown planthopper survival was not affected by potassium (K) supplementation to its host plants. This is as because both feeding stimulants (SP and TFS) and feeding deterrent-Si were reduced simultaneously in the K-applied host plant tissue. A report showed that K supplementation increases the concentration of K, but it reduces SP, TFS, and Si (Rashid et al. 2016a). Moreover, regression analysis clearly indicated that survival of BPH was not correlated with plant tissue content of K. However, K subsidy may affect indirectly by limiting other nutrients and also affect on other species. Several reports showed that K had negative effect on other insects (Salim 2002; Denke et al. 2000; Sarwar et al. 2011; El-Zahi et al. 2012; Sarwar 2012). In the present study, a marginal increase of BPH survival was recorded with phosphorus (P) supplementation to host plant although it was statistically insignificant. Rashid et al. (2016a) reported that application of P to the rice plants increased the concentration of P. Regression analysis in the present study indicated that P content in plant tissue had positive but weak relationship with survival of BPH. Similarly, positive effect of P on survivorship of planthopper Prokelisia marginata has been reported by Huberty and Denno (2006).
Generation time of insect mainly depends on development period from immature to adult stage. It is an important factor of BPH population development in rice field. Brown planthopper has to complete at least three generation cycles (life cycle) in a same rice field for producing sufficient population that can cause hopper burn. Generally, BPH has little scope to complete these three cycles in a same rice crop season, but there is a great chance if BPH can reduce its generation time. In our study, N inputs had remarkable effect on growth and development of BPH. Brown planthopper nymph raised on N-enriched plants developed more rapidly than that of N-deficient plants. Its development period from first instar nymph to adult was reduced by about 3 days when reared on N-enriched compared to N-deficient rice plant. The result of the present study is in conformity with the report of Lu et al. (2004). The phloem amino acids such as aspartic and glutamic acids remain in host phloem as soluble protein which are feeding stimulants of BPH (Sogawa 1982). On the other hand, Si-enriched plants are more tolerant as it is less palatable to insects (Rashid et al. 2016a). Rice plants supplemented with higher N fertilizer has higher soluble protein concentrations (Rashid et al. 2016a), which might provide a more favorable condition and better nutrition for BPH that caused its faster development. Furthermore, regression analysis in this study clearly indicated that BPH nymphal duration was negatively associated with plant tissue content of N, SP, and TFS, but positively with Si. Some reports showed that N is essential for growth and development of different insects (Facknath and Lalljee 2005; Huberty and Denno 2006; Lu et al. 2007). On the other hand, P and K inputs had no effect on nymphal duration of BPH. However, regression analysis detected weak relationship between plant tissue P content and BPH nymphal duration. Huberty and Denno (2006) reported that P fertilization reduced development time of planthopper P. marginata.
The N fertilization to the rice plants strongly influenced weights of N. lugens. In the present study, BPH grew to larger size and gained more body weight when it was reared on N-enriched plants. Its body weight was increased by about 30 to 40% when raised on the N-enriched plants. Enhanced N and SP with a smaller amount of Si in N-applied rice plant tissues might contribute to increased body weight of BPH. A good number of authors reported that the BPH had higher body weight on N-enriched plants (Lu et al. 2004; Prasad et al. 2005). Likewise, BPH showed a significant increase in its adult body weight on P-enriched plants compared to the P-poor plants. Some reports showed that P is an important determinant of body size of invertebrates (Busch and Phelan 1999; Huberty and Denno 2006). Body weight increase was only around 10% for P subsidy. Effect of P on BPH body weight was far less in compared to N, as because the relative mismatch in N content between the insect and their host rice plant was greater than that for P (Rashid et al. 2016a). Huberty and Denno (2006) reported that P limitation and associated fitness penalties on performance and population dynamics of P. marginata were not as severe as N limitation. However, K fertilization to rice plant had no significant effect on body weight of BPH.
Conclusions
In conclusion, N inputs improved all ecological fitness traits of BPH providing greater survival, shorter generation time, and larger body size. BPH outbreak in rice field could be explained by the association between its ecological fitness parameters and plant tissue content of N, P, Si, soluble protein, and soluble sugar. All fitness traits of BPH were found to be correlated strongly with the rice plant (host) tissue concentration of N, Si, soluble protein, and total free sugar. Phosphorus application markedly increased body weight of BPH. Furthermore, P content had positive linear relationship with BPH body weight, but its relationships with all other parameters of the insect were weak. Potassium supplementation had no effect on BPH fitness traits. Nitrogen and P inputs are to be used judiciously; otherwise, overuse of these two inputs may lead to BPH outbreak through enhancement of its ecological fitness potential. These findings can lead to better predictive capabilities in both nutritional ecology and pest management of rice.
References
Amtmann A, Troufflard S, Armengaud P (2008) The effect of potassium nutrition on pest and disease resistance in plants. Physiol Plant 133:682–691
Armengaud P, Sulpice R, Miller AJ, Stitt M, Amtmann A, Gibon Y (2009) Multilevel analysis of primary metabolism provides new insights into the role of potassium nutrition for glycolysis and nitrogen assimilation in Arabidopsis roots. Plant Physiol 150:772–785
Asher C, Grundon N, Menzies NN, 2002. How to unravel and solve soil fertility problems. ACIAR Monograph, No 83, Canberra, p 139
Baskaran P, Narayanasamy P, Pari A (1985) The role of potassium in incidence of insect pests among crop plants, with particular reference to rice. Role of Potassium in Crop Resistance to Insect Pests. Research Series no. 3, Potash Research Institute of India, Guragaon, Haryana, India, pp 63–68
Bernays EA (1990) Insect-plant interactions. vol 3, CRC Press, Boca Raton, p 176
Boersma M, Elser JJ (2006) Too much of a good thing: on stoichiometrically balanced diets and maximal growth. Ecology 87:1325–1330
Bottrell DG, Schoenly KG (2012) Resurrecting the ghost of green revolutions past: The brown planthopper as a recurring threat to high-yielding rice production in tropical Asia. J Asia-Pacific Entomol 15:122–140
Busch JW, Phelan L (1999) Mixture models of soybean growth and herbivore performance in response to nitrogen-sulphur-phosphorus nutrient interactions. Ecol Entomol 24:132–145
Chakravarthy AK, Doddabasappa B, Shashank PR (2013) The potential impacts of climate change on insect pests in cultivated ecosystems: an Indian perspective. In: Nautiyal S et al (eds) Knowledge systems of societies for adaptation and mitigation of impacts of climate change: Environmental science and engineering. Springer, Berlin Heidelberg, pp 143–162
Cook AG, Denno RF (1994) Planthopper/plant interactions: feeding behavior, plant nutrition, plant defense and host plant specialization. In: Denno RF, Perfect TJ (eds) Planthoppers: Their ecology and management. Chapman and Hall, London, pp 114–139
Dale D (1988) Plant mediated effects of soil mineral stress on insects. In: Heinrich EA (ed) Plant stress insect interactions. John Wiley and Sons, New York, pp 35–110
De Bruyn L, Scheirs J, Verhagen R (2002) Nutrient stress, host plant quality and herbivore performance of a leaf-mining fly on grass. Oecologia 130:594–599
Denke D, Schulthess F, Bonato O, Gounou S, Smith H (2000) Effect of the application of potassium on the development, survival and fecundity of Sesamia calamistis Hampson and of Eldana saccharina Walker on maize. Insect Sci Appl 20:151–156
Elser JJ, Fagan WF, Denno RF, Dobberfuhl DR, Folarin A, Huberty A, Interlandi S, Kilham SS, McCauley E, Schulz KL, Siemann EH, Sterner RW (2000) Nutritional constraints in terrestrial and freshwater food webs. Nature 408:578–580
Elser JJ, Hayakawa K, Urabe J (2001) Nutrient limitation reduces food quality for zooplankton: Daphnia response to seston phosphorus enrichment. Ecology 82:898–903
El-Zahi ES, Arif SA, Jehan BA, El-Naggar MEH, El-Dewy (2012) Inorganic fertilization of cotton field-plants in relation to sucking insects and yield production components of cotton plants. J Am Sci 8:509–517
Facknath S, Lalljee B (2005) Effect of soil applied complex fertilizer on an insect host plant relationship: Liriomyza trifolii on Solanum tuberosum. Entomol Exp Appl 115:67–77
Fischer K, Fiedler K (2000) Response of the copper butterfly Lycaena tityus to increased leaf nitrogen in natural food plants: evidence against the nitrogen limitation hypothesis. Oecologia 124:235–241
Han P, Lavoir AV, Le Bot J, Amiens-Desneux E, Desneux N (2014) Nitrogen and water availability to tomato plants triggers bottom-up effects on the leafminer Tuta absoluta. Sci Rep 4:4455. doi:10.1038/srep04455
Han Y, Lie W, Wen L, Hou M (2015) Silicon mediated resistance in a susceptible rice variety to the rice leaf folder Cnaphalocrocis medinalis Guenée (Lepidoptera: Pyralidae). PLoS ONE 10(4):e0120557. doi:10.1371/journal.pone.0120557
Huberty FA, Denno RF (2006) Consequences of nitrogen and phosphorus limitation for the performance of two planthoppers with divergent life-history strategies. Oecologia 149:444–455
Jauset AM, Sarasua MJ, Avilla J, Albajes R (2000) Effect of nitrogen fertilization level applied to tomato on the greenhouse whitefly. Crop Prot 19:255–261
Kaneshiro LN, Johnson MW (1996) Tritropic effects of leaf nitrogen on Liriomyza trifolii (Burgess) and an associated parasitoid Chrysocharis oscinidis (Ashmead) on bean. Biol Control 6:186–192
Kerpel SM, Soprano E, Moreira GRP (2006) Effect of nitrogen on Passiflora suberosa L. (Passifloraceae) and consequences for larval performance and oviposition in Helconius eratophyllis (Fabricius) (Lepidoptera: Nymphalidae). Neotrop Entomol 35:192–200
Kvedaras OL, Keeping MG (2007) Silicon impedes stalk penetration by the borer Eldana saccharina in sugarcane. Entomol Exp Appl 125:103–110
Kvedaras OL, An M, Choi YS, Gurr GM (2010) Silicon enhances natural enemy attraction and biological control through induced plant defences. Bull Entomol Res 100:367–371
Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the folin phenol reagent. J Biol Chem 193:265–275
Lu ZX, Heong KL (2009) Effects of nitrogen-enriched rice plants on ecological fitness of planthoppers. In: Heong KL, Hardy B (eds) Planthoppers: New threats to the sustainability of intensive rice production systems in Asia. International Rice Research Institute, Los Baños, pp 247–256
Lu ZX, Heong KL, Yu XP, Hu C (2004) Effects of plant nitrogen on ecological fitness of the brown planthopper, Nilaparvata lugens in rice. J Asia-Pacific Entomol 7:97–104
Lu ZX, Yu XP, Heong KL, Hu C (2007) Effect of nitrogen fertilizer on herbivores and its stimulation to major insect pests in rice. Rice Sci 14:56–66
Marschner P (2012) Marschner’s mineral nutrition of higher plants, 3rd edn. Academic, London, p 672
Nikpay A, Nejadian ES, Goldasteh S, Farazmand H (2015) Response of sugarcane and sugarcane stalk borers Sesamia spp (Lepidoptera: Noctuidae) to calcium silicate fertilization. Neotrop Entomol. doi:10.1007/s13744-015-0298-1
Oliveira MD, Barbosa PRR, Silva-Torres CSA, Silva RR, Barros EM, Torres JB (2014) Reproductive performance of striped mealybug Ferrisia virgata Cockerell (Hemiptera: Pseudococcidae) on water-stressed cotton plants subjected to nitrogen fertilization. Arthropod Plant Interact 8:461–468
Prasad BR, Pasalu IC, Raju NBT, Lingaiah T (2005) Effect of nitrogen levels and rice varieties on brown planthopper adult weight and amount of honeydew excretion. Ann Plant Prot Sci 13:243–245
Preap V, Zalucki MP, Nesbitt HJ, Jahn GC (2001) Effect of fertilizer, pesticide treatment and plant variety on realized fecundity and survival rates of Nilaparvata lugens (Stål); generating outbreak in Cambodia. J Asia-Pacific Entomol 4:75–84
Prestidge RA (1982) Instar duration, adult consumption, oviposition and nitrogen utilization efficiencies of leafhoppers feeding on different quality food. Ecol Entomol 7:91–101
Rashid MM, Jahan M, Islam KS (2016a) Impact of nitrogen, phosphorus and potassium on brown planthopper and tolerance of its host rice plants. Rice Sci 23:119–131
Rashid MM, Jahan M, Islam KS (2016b) Response of adult brown planthopper Nilaparvata lugens (Stål) to rice nutrient management. Neotrop Entomol 45:588–596
Rashid MM, Jahan M, Islam KS (2017) Effect of nitrogen, phosphorus and potassium on host choice behavior of brown planthopper, Nilaparvata lugens (Stål) on rice cultivar. J Insect Behav 30:1–15. doi:10.1007/s10905-016-9594-9
Reynolds OL, Keeping MG, Meyer JH (2009) Silicon-augmented resistance of plants to herbivorous insects: A review. Ann Appl Biol 155:171–186
Salim M (2002) Effects of potassium nutrition on growth, biomass and chemical composition of rice plants and on host-insect interaction. Pakistan J Agric Res 17:14–21
Sarwar M (2012) Effects of potassium fertilization on population build up of rice stem borers (lepidopteron pests) and rice (Oryza sativa L.) yield. J Cereals Oilseeds 3:6–9
Sarwar M, Ahmad N, Tofique M (2011) Impact of soil potassium on population buildup of aphid (Homoptera: Aphididae) and crop yield in canola (Brassica napus L.) field. Pakistan J Zool 43:15–19
Schoonhoven LM, van Loon JJA, Dicke M (2005) Insect-plant biology. Oxford University Press, Oxford, p 440
Slansky FJ, Scriber JM (1985) Food consumption and utilization. In: Comprehensive insect physiology, biochemistry and pharmacology. Ed. by Kerkut GA, Gilbert LI, vol 4, Pergamon Press, Oxford, pp 87–163
Sogawa K (1982) The rice brown planthopper: Feeding physiology and host plant interactions. Ann Rev Entomol 27:49–73
Sogawa K (2015) Planthopper outbreaks in different paddy ecosystems in Asia: man-made hopper plagues that threatened the green revolution in rice. In: Heong KL et al. (eds) Rice Planthoppers: Ecology, management, socio-economics and policy. Zhejiang University Press; Hangzhou and Springer Science + Business Media Dordrecht, DOI: 10.1007/978-94-017-9535-7_2
SPSSInc (2007) SPSS for windows, version 16. SPSS Inc, Chicago
Sterner RW, Elser JJ (2002) Ecological stoichiometry: the biology of elements from molecules to the biosphere. Princeton University Press, Princeton, p 584
Urabe J, Clasen J, Sterner RW (1997) Phosphorus limitation of Daphnia: is it real? Limnol Oceanog 42:1436–1443
Wang M, Zheng Q, Shen Q, Guo S (2013) The critical role of potassium in plant stress response. Int J Mol Sci 14:7370–7390. doi:10.3390/ijms14047370
Wier AT, Boethel DJ (1995) Feeding, growth, and survival of soybean looper in response to nitrogen fertilization of nonnodulating soybean. Environ Entomol 24:326–331
Yoshida S, Forno DA, Cock JH, Gomez KA (1976) Laboratory manual for physiological studies of rice, 3rd edn. International Rice Research Institute, Los Baños, p 83
Acknowledgements
The authors wish to thank the authority of Bangladesh Agriculture Research Council for providing financial support to perform the research activities.
Authors’ contributions
MMR generated the idea, designed the study, carried out the experiment works, conducted the sample collection and laboratory analysis, collected and analyzed the data, and wrote the manuscript. MJ designed the study and wrote and revised the manuscript draft. KSI participated in its design and coordination and helped to draft the manuscript. MAL provided the statistical assistance and revised the manuscript draft. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Rashid, M.M., Jahan, M., Islam, K.S. et al. Ecological fitness of brown planthopper, Nilaparvata lugens (Stål), to rice nutrient management. Ecol Process 6, 15 (2017). https://doi.org/10.1186/s13717-017-0080-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13717-017-0080-x