
Abstract
Fndc5, a well-defined myokine and also identified as an adipokine, has a critical role in modulation of metabolism and protection against obesity. These important functions are mediated by irisin, a secretory peptide produced from proteolytic processing of Fndc5. The other beneficial physiological effects of irisin are alleviation of oxidative stress, neuroprotective effects, and anti-inflammatory properties and associated anti-metastatic effects. Fndc5/irisin exerts its biological effects through several intracellular signaling pathways. The major signaling pathway is thought to be MAPK signaling pathways which are involved in neural differentiation, browning of white adipocytes, as well as osteoblast proliferation and differentiation. Other essential functions of Fndc5/irisin are mediated through additional pathways including AMPK pathway, PI3K/AKT, and STAT3/Snail. Thorough understanding of the mechanisms of irisin actions are essential in order to develop Fndc5/irisin for therapeutic purposes. In the present review, we focus on the current knowledge of the signaling pathways that elicit irisin actions.
Similar content being viewed by others
Background
Fibronectin type III domain-containing protein 5 (FNDC5), also called fibronectin type III repeat containing protein (FRCP2) and Peroxisomal Protein (Pep) was first discovered and characterized in 2002 by two independent groups [1, 2]. Böstrom and colleagues first reported increased FNDC5 transcript levels in the skeletal muscle of mice and humans after exercise. FNDC5 encodes a PGC1α-dependent myokine, as a part Fndc5 protein is proteolytically processed and secreted as irisin, which can promote conversion of white adipose tissue (WAT) to brown adipose tissue (BAT) by increased UCP1 expression [3]. Fndc5 transcript is expressed in multiple tissues including the heart, brain, ovary, testis, kidney, stomach and liver [4]. Literature mining indicates that Fndc5 not only plays a vital role in energy metabolism but also it has crucial roles in a variety of processes such as inflammation, proliferation, metastasis and neural differentiation. In this review we cover the best-understood cellular signaling pathways that Fndc5/irisin acts to elicit these physiological effects.
Fndc5 and MAPK signaling pathways
The mitogen-activated protein kinases (MAPKs) regulate a variety of cellular processes by relaying extracellular signals to intracellular responses [5]. MAPK signaling impacts multiple fundamental cellular processes such as gene expression, mitosis, metabolism, motility, survival, apoptosis, and differentiation. The best understood MAPKs are the conventional MAPKs: The c-Jun N-terminal kinases 1–3 (JNK1-3), extracellular signal-regulated kinase 1 and 2 (ERK1/2), the p38 isoforms (α, β, γ, and δ) and ERK5 families. Among these, the less-understood MAPKs are Erk3/4, and Erk7/8 and stress activated protein kinases (SAPK1A, 1B, 1C) [5, 6]. The extracellular stimuli include environmental stressors, growth factors, and cytokines, which activate MAPKs via both receptor-dependent and -independent mechanisms. Each group of conventional MAPKs is composed of a set of three evolutionarily conserved, sequentially acting kinases: a MAPK, a mitogen-activated protein kinase kinase (MAPKK), and a mitogen-activated protein kinase kinase kinase (MAPKKK) [5]. The major functions regulated by the MAPKs are mediated through their phosphorylation of a variety of protein substrates; including members of a family of protein kinases termed MAPK activated protein kinases (MAPKAPKs). Recent studies indicate that Fndc5 acts mostly through MAPK signaling pathways in numerous cellular processes (Fig. 1). A list of physiological effects of irisin and downstream pathways are shown in Table 1.

Schematic representation of the the main physiological activities mediated by Fndc5/Irisin through MAP-kinase signaling pathways. The variety of cell differentiation and physiological activities of Fndc5/irisin and the MAPK pathways they elicit are depicted. As shown, through this signaling pathway, irisin is not only responsible for neural cells and osteocytes differentiation but also triggers glucose uptake by the muscles and browning of WAT
Fndc5 plays a vital role in neural differentiation through ERK1/2 pathway
The vital role of Fndc5 in the process of neural differentiation and protection have been shown in many studies [7,8,9,10,11,12,13,14,15]. Fndc5 expression elevates after retinoic acid (RA) treatment of mouse embryonic stem cells (mESCs) in the process of neural differentiation [7]. The importance of Fndc5 in neural differentiation process was shown by loss and gain of function studies [9, 10]. RA binds to its nuclear receptor, retinoic acid receptor (RAR), and then acts as a transcription factor to affect RA-responsive genes, including induction of the genes encoding MAPKs (ERK1/2, JNK, P38) [16,17,18,19]. RA treatment of mouse embryoid bodies (EBs) elevated ERK1/2 activity, triggering an increase in Fndc5 and BDNF transcript levels in neural differentiation of mouse and human embryonic stem cells. Consistently, ERK1/2 loss of function significantly decreased Fndc5 and BDNF expression during neural differentiation [8] (Table 1).
Browning of white adipocytes is mediated by Fndc5 through p38 and ERK MAP kinase signaling
In its critical role regulating energy metabolism, irisin exerts beneficial effects through the conversion of WAT to BAT that is associated with weight loss and improved glucose homeostasis [3]. Owing to this important physiological regulation, irisin shows great therapeutic potential in diabetes and obesity [20]. Irisin induces the browning of WAT through p38 and ERK signaling [21]. Phosphorylated p38 (P-p38) and phosphorylated ERK (P-ERK) were both significantly increased following treatment with recombinant irisin in both primary rat and 3T3-L1 adipocytes. Irisin up-regulated uncoupling protein-1 (UCP1) in this process, a response that was blocked with drugs that inhibit p38 or ERK [21] (Table 1).
Irisin triggers osteoblast proliferation and differentiation via p38 and ERK signaling pathways
Bone metabolic diseases are a diverse group of bone metabolism disorders, mostly characterized by decreased bone mineral density in calcium or phosphorous and vitamin D [22, 23]. These diseases have a significant impact on the elderly population [23]. Exercise, by maintaining bone mass and strength, prevents bone cell diminishment and acts as a vital anti-aging factor for preserving bone integrity [24,25,26]. Several studies recently have shown that irisin acts as an exercise induced hormone, which promotes osteoblast proliferation and differentiation through activating the p38 and ERK [27,28,29]. Involvement of these signaling pathways in osteogenesis was confirmed by inhibition of each signaling pathways, using their inhibitors [28]. On the other hand, bone loss was prevented and osteoporosis was blocked in mice lacking irisin. Kim et al. showed that irisin binds directly to αv integrin receptors on osteocyte cells and induces the expression of sclerostin which involves bone resorption by increasing osteoclasts activity [30]. Therefore, the therapeutic potential of irisin in bone metabolism disease is unclear but has strong potential (Table 1).
p38–PGC-1α–irisin–betatrophin axis decreases insulin resistance
A new hormone, betatrophin, has recently been identified that contributes to pancreatic β-cell regeneration and specifically increases β-cell mass in mice. Betatrophin is connected by a new pathway involved in insulin resistance [31]. Irisin basically acts on WAT cells after endurance exercise training through induction of UCP1 expression and energy expenditure [3]. Irisin not only induces UCP1 expression through MAPK pathways (P38 and ERK) but also promotes the expression of betatrophin through these pathways [31] (Table 1).
Fndc5 regulates endothelial cell proliferation through the ERK1/2 MAPK pathway
The integrity of endothelial cells is critical, as their dysfunction is responsible for a variety of vascular diseases like chronic metabolic disease [32]. Endothelial cell proliferation is vital for new blood vessel growth during angiogenesis, especially in diseases like type II diabetes to support wound healing [33]. In addition to its role in regulating metabolic homeostasis, irisin may stimulate proliferation in some cell types. Irisin increases human umbilical vein endothelial cell (HUVEC) proliferation by activating ERK signaling pathways, potentially supporting new blood vessel growth. Irisin also promotes INS-1 cell proliferation via the ERK and p38 MAPK signaling pathways [34] (Table 1).
Glucose uptake and homeostasis of Fndc5 through P38 MAPK and ERK pathways
Skeletal muscle is the main source of irisin production. Irisin has diverse physiological functions including thermogenesis, glucose metabolism, increased metabolism, differentiation, and proliferation [3]. A recent study showed that irisin stimulates glucose uptake in muscle cells through p38 signaling activated by ROS-mediated AMPK activation [35]. β-arrestin-2 also has an active role in irisin induced glucose metabolism in type 2 diabetes mellitus (T2DM) by controlling the p38 MAPK signaling. Irisin not only elevates glucose uptake but also plays an essential role in glucose homeostasis by direct effects on adipose tissue, muscle, liver and pancreas and sometimes by indirect effects through synergistic effects with other hormones. In adipose tissue in addition to increase in UCP1 expression via p38 MAPK and ERK pathways, glucose homeostasis, is mediated by lipolysis stimulation through the cyclic AMP–protein kinase A (PKA)–perilipin–hormone-sensitive lipase (HSL) pathway [36]. These findings present a novel therapeutic avenue for potential treatment of diabetes [37] (Table 1).
Relevance between irisin and insulin signaling through p38 and ERK MAPKs
A positive association between irisin and insulin resistance is reported in muscle [4, 38, 39]. Palmitate treatment of the C2C12 cell line resulted in decreased AKT and ERK signaling and irisin antagonized the reduced or diminished phosphorylation of AKT and ERK after palmitate treatment [40]. To understand the fundamental role of irisin in the amelioration of insulin resistance in involved tissues especially muscle tissue, more experiments are needed (Table 1).
Taken together, it seems that MAPK signaling pathways which are involved in cellular energy expenditure, cell proliferation and differentiation in a variety of cell and tissue types could be activated by irisin. This feature of irisin function is a very critical aspect physiologically for coordination between metabolic rate of tissues and organs which is governed by muscle and fat tissues as the main secretory sources of irisin.
Other signaling pathways for Fndc5/irisin function
Fndc5/irisin could act through alternative signaling pathways. The outline of these pathways is shown in Fig. 2 as follows:

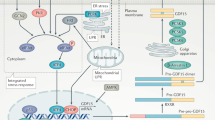
Graphic summary of physiological activities of Fndc5/Irisin that are elicited through pathways other than MAP-kinase signaling. Alternative signaling pathways that transmit the effects of Fndc5/Irisin are AMP-kinase, STAT3 and the TLR4 pathways. These pathways are involved in a set of cellular functions responsible for proliferation, anti-metastatic and anti-inflammatory activities. Irisin improves cardiac hypertrophy by inducing protective autophagy via mTOR independent activation of AMPK-ULK1 and AMPK- mTOR pathways. On the other hand, Irisin has an antidepressant-like effect in hippocampus. Of note that, Irisin lowers blood pressure via the AMPK-Akt-eNOS-NO pathway in endothelial cells. Anti-inflammatory properties of irisin are connected to TLR4/MyD88 signaling pathway activation in macrophages. For detailed information please see the text
Adipogenesis is suppressed by irisin through Wnt signaling
In addition to myokine activity, Irisin can also act as type of adipokine. Therefore, besides of the main role of irisin which is browning of WAT, irisin also prohibits accumulation of lipids through up-regulation of adipose triglyceride lipase (ATGL) and down regulation of fatty acid synthase (FAS) [41]. This modulatory effect is mediated by the PPARγ, C/EBPa, and FABP4 axis under control of Wnt signaling, as FNDC5/irisin up regulates Wnt6 and Wnt10a and Wnt 10b [42]. Wnt signaling is inhibitory for adipocyte differentiation [43]. This aspect of Irisin function through Wnt signaling is complementary to browning of WAT which is mediated by MAPK signaling pathways to suppress of body fat percentage (BFP) in the human body. Therefore, it can be concluded that the main aspect of irisin secretion through the muscle is governing of fat size of the body and thereby governing the metabolic rate of fat tissue.
Irisin has a neuroprotective role through stimulation of cAMP/PKA/CREB pathway
The role of FNDC5/irisin in learning and memory is mediated through the expression of brain-derived neurotrophic factor (BDNF) in the hippocampus [44]. This finding supports the potential role for FNDC5/irisin in preventing brain disorders such as Alzheimer’s disease (AD).
CREB (cAMP response element-binding protein) is a cellular transcription factor which has a well-documented role in neuronal plasticity and long-term memory formation in the brain [45]. Recent studies have revealed that recombinant irisin stimulates the cAMP/PKA/CREB pathway not only in humane cortical slices but also in mouse hippocampal slices [46].
Irisin increases H19-7 cell proliferation by activating STAT3 pathway
Irisin is expressed in the human brain [4, 13]. We have also shown that knockdown of Fndc5 decreased neural differentiation of mouse embryonic stem cells whereas its overexpression increased the rate of neural differentiation [9, 10]. To clarify whether irisin plays an important role in neurogenic regulation, Moon and colleagues showed that irisin increases cell proliferation in mouse H19-7 HN cells via STAT3, but not AMPK and/or ERK, whereas irisin has no dose-dependent effect on neurite outgrowth and synaptogenesis in these cells [47] (Table 1). Together, these results demonstrate that irisin mediates neuroprotective effects partly through activation of the STAT3 signaling pathway. Through a combination of its neuroprotective property and its induced neural differentiation through MAPK signaling pathway, irisin likely serves important function to support neuronal health.
Fndc5 attenuates inflammation and insulin resistance via AMPK pathway
Obesity is a complex disease that triggers inflammation and macrophage accumulation in adipose tissue and subsequently leads to metabolic diseases, including type 2 diabetes and insulin resistance [48,49,50]. Recent studies have shown that improved hyperlipidemia and increased lipolysis are two consequence of Fndc5 overexpression in adipose tissues of obese mice [51]. Guo-Qing Zhu’s research group demonstrated that Fndc5 deficiency reduced insulin sensitivity in obese mice. Their findings showed that Fndc5 plays a critical role in attenuating adipose tissue inflammation and insulin resistance [51]. The improvement effects of Fndc5 are significantly alleviated by the AMPK inhibitor Compound C (CC), but did not change by utilizing an AMPK activator, AICAR. These data demonstrate that AMPK reduces inflammation and M1 macrophage polarization by Fndc5 [52] (Table 1). This function of irisin, compared with those described already for irisin through MAPK signaling, demonstrated that irisin exerts its effects through alternative pathways.
Irisin exerts its anti-metastatic effects via the PI3K/AKT pathway in lung cancer tissue
One of the pivotal pathways in cancer cell growth, proliferation, and survival, is the PI3K/AKT pathway which is elevated in a variety of cancers including ovarian, breast and pancreatic cancers [53,54,55]. Irisin inhibits the migration, proliferation, and invasion of lung cancer cells and reduced the expression of EMT markers by inhibiting the PI3K/AKT pathway. From a mechanistic perspective, irisin can reverse the activity of epithelial–mesenchymal transition (EMT) and inhibits the expression of Snail via the PI3K/AKT pathway [56]. Specifically, irisin inhibited EMT and reduced the invasion of lung cancer cells via the PI3K/AKT/Snail pathway [56]. Conversely however, increased irisin levels may have protective roles in liver cancer cells through partial activation of the PI3K/AKT pathway, which may facilitate liver cancer progression and decrease the sensitivity to chemotherapy [57] (Table 1).
Effect of irisin on migration and invasion of osteosarcoma cells through the STAT3/Snail signaling pathway
EMT is a cellular process which occurs during normal embryonic development and wound healing. It is a highly conserved process, by which epithelial cells lose their cell polarity and cell–cell adhesion [58]. IL-6 appears to promote the proliferation, metastasis and angiogenesis of osteosarcoma through several downstream signals including AKT, ERK1/2 MAPK and STAT3 [59,60,61]. Kong and colleagues showed that irisin treatment of osteosarcoma cells inhibited the proliferation, migration and invasion of osteosarcoma cells by reversing IL-6-induced EMT. In a further study, it was ascertained that irisin inhibits IL-6-induced STAT3 phosphorylation [62] (Table 1).
Irisin suppresses pancreatic cancer cell growth via the activation of AMPK
AMPK-mTOR is a major signaling pathway in progress of pancreatic cancer. Irisin administration reportedly suppresses pancreatic cancer cell growth via the activation of AMPK and downregulation of the mTOR pathways, thereby inhibiting EMT of pancreatic cancer cells [63]. Moreover, irisin is responsible for increasing caspase activity in a process called “attenuation of cell death resistance”. Notably, irisin also suppresses other hallmarks of cancer such as “maintaining proliferative signaling” through targeting the PI3K/Akt pathway and also “evading growth suppressors” through the AMPK-mTOR pathway. In summary, irisin exerts cancer suppression through reduction in proinflammatory cytokines and adipokines linked to obesity status.
Browning of WAT in myostatin-knockout mouse through activating AMPK-PGC1α-FNDC5 signaling pathway
Myostatin, a myokine released by myocytes, acts on muscle cells by inhibition of myogenesis, muscle cell growth and differentiation. Myostatin (Mstn) knockout mouse showed significantly more muscle mass [64]. It has been shown that WAT of Mstn−/− mice show attributes of BAT through significantly increased expression of BAT marker genes, including Pgc1α and Ucp1 [65]. Moreover, in muscles of Mstn−/− mice, the level of total AMPK and activated pAMPK increased significantly as AMPK-PGC1α-Fndc5 pathway was activated in the muscle of Mstn−/− mice, leading to increased production of irisin [65] (Table 1).
Anti-inflammatory properties of irisin are connected to TLR4/MyD88 signaling pathway activation
Irisin also acts as an adipokine and exerts a potential protective effect on the progress of obesity-related diseases, such as arteriosclerosis, insulin resistance, and type 2 diabetes. Potential anti-inflammatory properties of irisin have been demonstrated [66]. Macrophage RAW 264.7 cells stimulation with lipopolysaccharide (LPS; 100 ng/mL) and irisin pretreatment caused a dramatic decrease in Toll like receptor (TLR4) and Myeloid differentiation primary response protein 88 (MyD88) levels, and decreased the phosphorylation of nuclear factor-κB (NF-κB), thereby reducing the release of vital pro-inflammatory cytokines (IL-6, TNF-α, and IL-1β) and keratinocyte chemo attractant (KC) and monocyte chemotactic protein 1 (MCP-1). Moreover, irisin exerts this anti-inflammatory effect through phosphorylation of MAPKs, where a significant reduction in p-JNK and p-ERK but not p-p38 was observed [66]. In conclusion, potential protective effects of irisin against the development of diseases associated with obesity, may be attributed in part to irisin’s anti-inflammatory properties (Table 1).
Negative regulation of serum irisin and skeletal muscle Fndc5 by SMAD3 during exercise
Smad3−/− mice transform WAT to BAT-like cell phenotype, thereby inferring protection against high fat diet-induced obesity and type 2 diabetes mellitus [67]. Irisin induces WAT browning similar to that observed in SMAD3-deficient mice [3]. SMAD3 represses Fndc5 and Pgc1α expression in skeletal muscle and in Smad3−/− mice exercise increases serum irisin and skeletal muscle Fndc5 as well as its upstream activator Pgc1α to a greater extent than in wild-type mice [67] (Table 1).
Irisin lowers blood pressure via the AMPK-Akt-eNOS-NO pathway in endothelial cells
Hypertension, affecting approximately one billion people worldwide, is a major risk factor for a variety of diseases including coronary artery disease, stroke, heart failure, atrial fibrillation, peripheral vascular disease, vision loss, chronic kidney disease, and dementia [68]. Exercise is a nonpharmacological anti-hypertensive factor responsible for lowering blood pressure through unknown mechanisms. Due to the close relationships between metabolic diseases and hypertension, it is postulated that exercise may act through irisin to elicit lowering of blood pressure [68]. AMPK could be activated by irisin, thereby down regulating intracellular ATP levels through increasing reactive oxygen species (ROS) or intracellular calcium concentrations [35]. In the vasculature, activated endothelial AMPK phosphorylates eNOS, stimulating NO release and subsequent vasodilation of both large conduit and resistance arteries [68]. This was demonstrated in the spontaneously hypertensive rat, where irisin lowers blood pressure by ameliorating endothelial dysfunction of the mesenteric artery through the AMPK-Akt-eNOS-NO signaling pathway [68]. In a similar study, irisin improved endothelial function in aortas of high fat diet-induced obese mice through activation of AMPK-eNOS signaling [69] (Table 1).
Antidepressant-like effect mediated by Fndc5/BDNF/Akt in mice by modulating hippocampal signaling pathway
There is mounting evidence for significant involvement of creatine in the pathophysiology of major depressive disorder (MDD) [70]. Several studies have shown that Akt and its downstream molecular targets effect MDD and may be targets for depression treatment [71,72,73]. It has shown that the acute antidepressant-like effect of creatine is dependent, at least in part, on PI3K/Akt signaling pathway activation [74]. AKT induces activation of BDNF [75, 76]. BDNF is a member of the neurotrophin family of growth factors, which regulates the survival and growth of neurons and has recently received attention in relation to the therapeutic action of antidepressant treatment [77, 78]. Several studies demonstrated that physical exercise increases BDNF levels in the hippocampus through PGC-1α activation and FNDC-5 expression modulation [44, 79, 80]. Subchronic administration of creatine in the hippocampus caused increased expression of PGC-1α, FNDC5 and BDNF through the FNDC5/BDNF/Akt pathway [81] (Table 1).
Irisin improves cardiac hypertrophy by inducing protective autophagy via mTOR independent activation of AMPK-ULK1 and AMPK- mTOR pathways
FNDC5 overexpression attenuated damage to transverse aortic constriction induced hypertrophy in the heart, demonstrating a protective effect of irisin against cardiomyocyte hypertrophy induced by angiotensin II or phenylephrine. Irisin deficiency decreased autophagy, whereas irisin overexpression elevated autophagic flux. ULK1 plays an essential role in the initiation of autophagy and can be regulated by AMPK and mTOR via direct phosphorylation at Ser555 and Ser757, respectively. Irisin increased the activity of AMPK but not Akt and MAPK in hypertrophic hearts and cultured cardiomyocytes which triggered further activation of ULK1 at Ser555 but not Ser757 and did not affect the mTOR-S6K axis [82]. Irisin may also display anti-fibrotic therapeutic potential to counter angiotensin II-related cardiac fibrosis. In skeletal muscle cells the ADAM family of metalloendopeptidases, especially ADAM10, is responsible for the cleavage of FNDC5 into irisin and for irisin-induced cardiac autophagy through activation of the AMPK-mTOR pathway. Since FNDC5 expression is significantly decreased in ischemic cardiomyopathy, in severe chronic heart failure mice, application of irisin may be beneficial as a novel therapeutic approach for treatment of heart disease [83] (Table 1).
In addition, FNDC5/irisin is responsible for the repair of cardiac tissue after an ischemic heart disease episode as it induces cell proliferation through activation of cardiac progenitor cells. In this context, there is a significant increase in proliferative markers such as Ki67 and phosphorylated histone 3, a reduction of histone deacetylase 4, and increased p38 acetylation in Irisin-treated cardiac progenitor cells [84].
Discussion
Irisin is a myokine that is secreted from skeletal muscle in response to exercise and stimulates convertion of WAT to BAT [3]. There is rising evidence to support a vital role for irisin in the regulation of metabolism and body fat reduction [85]. These effects can raise energy expenditure, increase oxygen consumption and reduce insulinemia [86].
However, more recent studies revealed additional vital roles for Fndc5/irisin in other tissues [36, 87, 88]. Recent studies have revealed that Irisin has anti-cancer, -depression, -hypertension and -cardiac hypertrophy properties [57, 62, 68, 81,82,83].
The inhibitory effects of irisin on inflammation is mediated by significant decrease in the release of vital pro-inflammatory cytokines. This property is associated with hyperphosphorylation of MAPKs induced by irisin [66]. Therefore, irisin acts as an important regulator of tissue cross-talk, mainly between muscle and other tissues/organs. Therefore, irisin is a potential option for preventing/treating a wide range of diseases including cancer [89]. In the present review we tried to delineate molecular mechanism of intracellular functions of irisin. However, more investigations are needed to clarify the precise mechanism of irisin actions and its spectrum of physiological effects.
Conclusion
Numerous studies demonstrate the physiological properties of irisin, pointing to its beneficial health potential in the maintenance of a variety of tissues and organs. With the promise of its health benefits, further studies to investigate and test the therapeutic applications of this signaling peptide are highly anticipated. Understanding the precise underlying mechanisms of Fndc5 is required to fully appreciate and appropriately apply Fndc5/irisin in cancer, aging and other metabolic diseases.
Availability of data and materials
Not applicable.
Abbreviations
- AD:
-
Alzheimer’s disease
- Ang II:
-
Angiotensin II
- ATGL:
-
Adipose triglyceride lipase
- BAT:
-
Brown adipose tissue
- BDNF:
-
Brain-derived neurotrophic factor
- BFP:
-
Body fat percentage
- CC:
-
Compound C
- EBs:
-
Embryoid bodies
- ERK1/2:
-
Extracellular signal-regulated kinase 1 and 2
- EMT:
-
Epithelial–mesenchymal transition
- FAS:
-
Fatty acid synthase
- FNDC5:
-
Fibronectin type III domain-containing 5 protein
- FRCP2:
-
Fibronectin type III repeat containing protein
- JNK:
-
c-Jun N-terminal kinases
- HUVEC:
-
Human umbilical vein endothelial cell
- IL-1β:
-
Interleukin 1β
- IL-6:
-
Interleukin 6
- KC:
-
Keratinocyte chemoattractant
- LPS:
-
Lipopolysaccharide
- MAPKs:
-
Mitogen-activated protein kinases
- MAPKAPKs:
-
MAPK activated protein kinases
- MAPKK:
-
Mitogen-activated protein kinase kinase
- MAPKKK:
-
Mitogen-activated protein kinase kinase kinase
- MCP-1:
-
Monocyte chemotactic protein 1
- MDD:
-
Major depressive disorder
- MyD88:
-
Myeloid differentiation primary response protein 88
- mESCs:
-
Mouse embryonic stem cells
- NF-κB:
-
Nuclear factor-κB
- NLK:
-
Nemo-like kinase
- P-ERK:
-
Phosphorylated ERK
- PE:
-
Phenylephrine
- Pep:
-
Peroxisomal protein
- PGC-1α:
-
Peroxisome proliferator-activated receptor-γ coactivator-1 alpha
- PKA:
-
Protein kinase A
- P-p38:
-
Phosphorylated p38
- PPAR-γ:
-
Peroxisome proliferator-activated receptor-γ
- RA:
-
Retinoic acid
- RAR:
-
Retinoic acid receptor
- RARE:
-
Retinoic acid response element
- ROS:
-
Reactive oxygen species
- SAPK:
-
Stress activated protein kinases
- TAC:
-
Transverse aortic constriction
- T2DM:
-
Type 2 diabetes mellitus
- TNFα:
-
Tumor necrosis factor α
- UCP:
-
Uncoupling protein
- WAT:
-
White adipose tissue
References
Ferrer-Martínez A, Ruiz-Lozano P, Chien KR. Mouse PeP: a novel peroxisomal protein linked to myoblast differentiation and development. Dev Dyn. 2002;224(2):154–67.
Teufel A, Malik N, Mukhopadhyay M, Westphal H. Frcp1 and Frcp2, two novel fibronectin type III repeat containing genes. Gene. 2002;297(1):79–83.
Boström P, Wu J, Jedrychowski MP, Korde A, Ye L, Lo JC, et al. A PGC1-α-dependent myokine that drives brown-fat-like development of white fat and thermogenesis. Nature. 2012;481(7382):463.
Huh JY, Panagiotou G, Mougios V, Brinkoetter M, Vamvini MT, Schneider BE, et al. FNDC5 and irisin in humans: I. Predictors of circulating concentrations in serum and plasma and II. mRNA expression and circulating concentrations in response to weight loss and exercise. Metabolism. 2012;61(12):1725–38.
Cargnello M, Roux PP. Activation and function of the MAPKs and their substrates, the MAPK-activated protein kinases. Microbiol Mol Biol Rev. 2011;75(1):50–83.
Sun Y, Liu W-Z, Liu T, Feng X, Yang N, Zhou H-F. Signaling pathway of MAPK/ERK in cell proliferation, differentiation, migration, senescence and apoptosis. J Recept Signal Transduction. 2015;35(6):600–4.
Ostadsharif M, Ghaedi K, Nasr-Esfahani MH, Mojbafan M, Tanhaie S, Karbalaie K, et al. The expression of peroxisomal protein transcripts increased by retinoic acid during neural differentiation. Differentiation. 2011;81(2):127–32.
Farahabadi SH, Ghaedi K, Zadegan FG, Karbalaie K, Rabiee F, Nematollahi M, et al. ERK1/2 is a key regulator of Fndc5 and PGC1α expression during neural differentiation of mESCs. Neuroscience. 2015;297:252–61.
Hashemi M-S, Ghaedi K, Salamian A, Karbalaie K, Emadi-Baygi M, Tanhaei S, et al. Fndc5 knockdown significantly decreased neural differentiation rate of mouse embryonic stem cells. Neuroscience. 2013;231:296–304.
Forouzanfar M, Rabiee F, Ghaedi K, Beheshti S, Tanhaei S, Shoaraye Nejati A, et al. Fndc5 overexpression facilitated neural differentiation of mouse embryonic stem cells. Cell Biol Int. 2015;39(5):629–37.
Tanhaei S, Nikpour P, Ghaedi K, Rabiee F, Homayouni Moghadam F, Nasr-Esfahani MH. RNA/protein discordant expression of Fndc5 in central nervous system is likely to be mediated through microRNAs. DNA Cell Biol. 2018;37(4):373–80.
Zhang J, Zhang W. Can irisin be a linker between physical activity and brain function? Biomol Concepts. 2016;7(4):253–8.
Ghahrizjani FA, Ghaedi K, Salamian A, Tanhaei S, Nejati AS, Salehi H, et al. Enhanced expression of FNDC5 in human embryonic stem cell-derived neural cells along with relevant embryonic neural tissues. Gene. 2015;557(2):123–9.
Farshbaf MJ, Ghaedi K, Megraw TL, Curtiss J, Faradonbeh MS, Vaziri P, et al. Does PGC1α/FNDC5/BDNF elicit the beneficial effects of exercise on neurodegenerative disorders? NeuroMol Med. 2016;18(1):1–15.
Wrann CD. FNDC5/Irisin–their role in the nervous system and as a mediator for beneficial effects of exercise on the brain. Brain Plasticity. 2015;1(1):55–61.
Amura CR, Marek L, Winn RA, Heasley LE. Inhibited neurogenesis in JNK1-deficient embryonic stem cells. Mol Cell Biol. 2005;25(24):10791–802.
Leppä S, Saffrich R, Ansorge W, Bohmann D. Differential regulation of c-Jun by ERK and JNK during PC12 cell differentiation. EMBO J. 1998;17(15):4404–13.
Morooka T, Nishida E. Requirement of p38 mitogen-activated protein kinase for neuronal differentiation in PC12 cells. J Biol Chem. 1998;273(38):24285–8.
Saffar AS, Dragon S, Ezzati P, Shan L, Gounni AS. Phosphatidylinositol 3-kinase and p38 mitogen-activated protein kinase regulate induction of Mcl-1 and survival in glucocorticoid-treated human neutrophils. J Allergy Clin Immunol. 2008;121(2):492–8.
Novelle MG, Contreras C, Romero-Picó A, López M, Diéguez C. Irisin, two years later. Int J Endocrinol. 2013;2013:506.
Zhang Y, Li R, Meng Y, Li S, Donelan W, Zhao Y, et al. Irisin stimulates browning of white adipocytes through mitogen-activated protein kinase p38 MAP kinase and ERK MAP kinase signaling. Diabetes. 2014;63(2):514–25.
Varacallo MA, Fox EJ. Osteoporosis and its complications. Med Clin. 2014;98(4):817–31.
Golob AL, Laya MB. Osteoporosis: screening, prevention, and management. Med Clin. 2015;99(3):587–606.
De Souza MJ, Arce JC, Nulsen JC, Puhl JL. Exercise and bone health across the life span. Muscatine: Women and Sport Interdisciplinary Perspectives Champaign (USA), Human Kinetics; 1994.
Cosman F, De Beur S, LeBoff M, Lewiecki E, Tanner B, Randall S, et al. Clinician’s guide to prevention and treatment of osteoporosis. Osteoporos Int. 2014;25(10):2359–81.
Papaioannou A, Morin S, Cheung AM, Atkinson S, Brown JP, Feldman S, et al. 2010 clinical practice guidelines for the diagnosis and management of osteoporosis in Canada: summary. Can Med Assoc J. 2010;182(17):1864–73.
Ma Y, Qiao X, Zeng R, Cheng R, Zhang J, Luo Y, et al. Irisin promotes proliferation but inhibits differentiation in osteoclast precursor cells. FASEB J. 2018;32:5813–23.
Qiao X, Nie Y, Ma Y, Chen Y, Cheng R, Yin W, et al. Irisin promotes osteoblast proliferation and differentiation via activating the MAP kinase signaling pathways. Sci Rep. 2016;6:18732.
Colaianni G, Cuscito C, Mongelli T, Oranger A, Mori G, Brunetti G, et al. Irisin enhances osteoblast differentiation in vitro. Int J Endocrinol. 2014;2014:902186.
Kim H, Wrann CD, Jedrychowski M, Vidoni S, Kitase Y, Nagano K, et al. Irisin mediates effects on bone and fat via αV integrin receptors. Cell. 2018;175(7):1756–68.
Crunkhorn S. Metabolic disorders: betatrophin boosts β-cells. Nat Rev Drug Discovery. 2013;12(7):504.
Song H, Wu F, Zhang Y, Zhang Y, Wang F, Jiang M, et al. Irisin promotes human umbilical vein endothelial cell proliferation through the ERK signaling pathway and partly suppresses high glucose-induced apoptosis. PLoS ONE. 2014;9(10):e110273.
Tepper OM, Galiano RD, Capla JM, Kalka C, Gagne PJ, Jacobowitz GR, et al. Human endothelial progenitor cells from type II diabetics exhibit impaired proliferation, adhesion, and incorporation into vascular structures. Circulation. 2002;106(22):2781–6.
Liu S, Du F, Li X, Wang M, Duan R, Zhang J, et al. Effects and underlying mechanisms of irisin on the proliferation and apoptosis of pancreatic β cells. PLoS ONE. 2017;12(4):e0175498.
Lee HJ, Lee JO, Kim N, Kim JK, Kim HI, Lee YW, et al. Irisin, a novel myokine, regulates glucose uptake in skeletal muscle cells via AMPK. Mol Endocrinol. 2015;29(6):873–81.
Perakakis N, Triantafyllou GA, Fernández-Real JM, Huh JY, Park KH, Seufert J, et al. Physiology and role of irisin in glucose homeostasis. Nat Rev Endocrinol. 2017;13(6):324.
Pang Y, Zhu H, Xu J, Yang L, Liu L, Li J. β-arrestin-2 is involved in irisin induced glucose metabolism in type 2 diabetes via p38 MAPK signaling. Exp Cell Res. 2017;360(2):199–204.
Stengel A, Hofmann T, Goebel-Stengel M, Elbelt U, Kobelt P, Klapp BF. Circulating levels of irisin in patients with anorexia nervosa and different stages of obesity–correlation with body mass index. Peptides. 2013;39:125–30.
Roca-Rivada A, Castelao C, Senin LL, Landrove MO, Baltar J, Crujeiras AB, et al. FNDC5/irisin is not only a myokine but also an adipokine. PLoS ONE. 2013;8(4):e60563.
Yang Z, Chen X, Chen Y, Zhao Q. Decreased irisin secretion contributes to muscle insulin resistance in high-fat diet mice. Int J Clin Exp Pathol. 2015;8(6):6490.
Huh J, Dincer F, Mesfum E, Mantzoros C. Irisin stimulates muscle growth-related genes and regulates adipocyte differentiation and metabolism in humans. Int J Obesity. 2014;38(12):1538–44.
Ma EB, Sahar NE, Jeong M, Huh JY. Irisin exerts inhibitory effect on adipogenesis through regulation of Wnt signaling. Front Physiol. 2019;10:1085.
Rosen ED, MacDougald OA. Adipocyte differentiation from the inside out. Nat Rev Mol Cell Biol. 2006;7(12):885–96.
Wrann CD, White JP, Salogiannnis J, Laznik-Bogoslavski D, Wu J, Ma D, et al. Exercise induces hippocampal BDNF through a PGC-1α/FNDC5 pathway. Cell Metab. 2013;18(5):649–59.
Silva AJ, Kogan JH, Frankland PW, Kida S. CREB and memory. Annu Rev Neurosci. 1998;21(1):127–48.
Lourenco MV, Frozza RL, de Freitas GB, Zhang H, Kincheski GC, Ribeiro FC, et al. Exercise-linked FNDC5/irisin rescues synaptic plasticity and memory defects in Alzheimer’s models. Nat Med. 2019;25(1):165–75.
Moon H-S, Dincer F, Mantzoros CS. Pharmacological concentrations of irisin increase cell proliferation without influencing markers of neurite outgrowth and synaptogenesis in mouse H19-7 hippocampal cell lines. Metabolism. 2013;62(8):1131–6.
Donath MY, Shoelson SE. Type 2 diabetes as an inflammatory disease. Nat Rev Immunol. 2011;11(2):98.
Tsai S-Y, Rodriguez AA, Dastidar SG, Del Greco E, Carr KL, Sitzmann JM, et al. Increased 4E-BP1 expression protects against diet-induced obesity and insulin resistance in male mice. Cell Rep. 2016;16(7):1903–14.
Lips MA, van Klinken JB, Pijl H, Janssen I, van Dijk KW, Koning F, et al. Weight loss induced by very low calorie diet is associated with a more beneficial systemic inflammatory profile than by Roux-en-Y gastric bypass. Metabolism. 2016;65(11):1614–20.
Xiong X-Q, Chen D, Sun H-J, Ding L, Wang J-J, Chen Q, et al. FNDC5 overexpression and irisin ameliorate glucose/lipid metabolic derangements and enhance lipolysis in obesity. Biochimica et Biophysica Acta (BBA) Mol Basis Dis. 2015;1852(9):1867–75.
Xiong X-Q, Geng Z, Zhou B, Zhang F, Han Y, Zhou Y-B, et al. FNDC5 attenuates adipose tissue inflammation and insulin resistance via AMPK-mediated macrophage polarization in obesity. Metabolism. 2018;83:31–41.
Vivanco I, Sawyers CL. The phosphatidylinositol 3-kinase–AKT pathway in human cancer. Nat Rev Cancer. 2002;2(7):489.
Ung MH, Wang GL, Varn FS, Cheng C. Application of pharmacologically induced transcriptomic profiles to interrogate PI3K-Akt-mTOR pathway activity associated with cancer patient prognosis. Oncotarget. 2016;7(51):84142.
Courtney KD, Corcoran RB, Engelman JA. The PI3K pathway as drug target in human cancer. J Clin Oncol. 2010;28(6):1075.
Shao L, Li H, Chen J, Song H, Zhang Y, Wu F, et al. Irisin suppresses the migration, proliferation, and invasion of lung cancer cells via inhibition of epithelial-to-mesenchymal transition. Biochem Biophys Res Commun. 2017;485(3):598–605.
Shi G, Tang N, Qiu J, Zhang D, Huang F, Cheng Y, et al. Irisin stimulates cell proliferation and invasion by targeting the PI3K/AKT pathway in human hepatocellular carcinoma. Biochem Biophys Res Commun. 2017;493(1):585–91.
Rosanò L, Cianfrocca R, Spinella F, Di Castro V, Nicotra MR, Lucidi A, et al. Acquisition of chemoresistance and EMT phenotype is linked with activation of the endothelin a receptor pathway in ovarian carcinoma cells. Clin Cancer Res. 2011;17:23250.
Tu B, Du L, Fan Q-M, Tang Z, Tang T-T. STAT3 activation by IL-6 from mesenchymal stem cells promotes the proliferation and metastasis of osteosarcoma. Cancer Lett. 2012;325(1):80–8.
Dou L, Wang S, Sui X, Meng X, Shen T, Huang X, et al. MiR-301a mediates the effect of IL-6 on the AKT/GSK pathway and hepatic glycogenesis by regulating PTEN expression. Cell Physiol Biochem. 2015;35(4):1413–24.
Che Q, Liu B-Y, Wang F-Y, He Y-Y, Lu W, Liao Y, et al. Interleukin 6 promotes endometrial cancer growth through an autocrine feedback loop involving ERK–NF-κB signaling pathway. Biochem Biophys Res Commun. 2014;446(1):167–72.
Kong G, Jiang Y, Sun X, Cao Z, Zhang G, Zhao Z, et al. Irisin reverses the IL-6 induced epithelial–mesenchymal transition in osteosarcoma cell migration and invasion through the STAT3/Snail signaling pathway. Oncol Rep. 2017;38(5):2647–56.
Liu J, Song N, Huang Y, Chen Y. Irisin inhibits pancreatic cancer cell growth via the AMPK-mTOR pathway. Sci Rep. 2018;8(1):1–10.
Guo T, Jou W, Chanturiya T, Portas J, Gavrilova O, McPherron AC. Myostatin inhibition in muscle, but not adipose tissue, decreases fat mass and improves insulin sensitivity. PLoS ONE. 2009;4(3):e4937.
Shan T, Liang X, Bi P, Kuang S. Myostatin knockout drives browning of white adipose tissue through activating the AMPK-PGC1α-Fndc5 pathway in muscle. FASEB J. 2013;27(5):1981–9.
Mazur-Bialy AI, Pocheć E, Zarawski M. Anti-inflammatory properties of irisin, mediator of physical activity, are connected with TLR4/MyD88 signaling pathway activation. Int J Mol Sci. 2017;18(4):701.
Tiano JP, Springer DA, Rane SG. SMAD3 negatively regulates serum irisin and skeletal muscle FNDC5 and peroxisome proliferator-activated receptor γ coactivator 1-α (PGC-1α) during exercise. J Biol Chem. 2015;290(12):7671–84.
Fu J, Han Y, Wang J, Liu Y, Zheng S, Zhou L, et al. Irisin lowers blood pressure by improvement of endothelial dysfunction via AMPK-Akt-eNOS-NO pathway in the spontaneously hypertensive Rat. J Am Heart Assoc. 2016;5(11):e003433.
Han F, Zhang S, Hou N, Wang D, Sun X. Irisin improves endothelial function in obese mice through the AMPK-eNOS pathway. Am J Physiol-Heart Circ Physiol. 2015;309(9):H1501–8.
Kious BM, Kondo DG, Renshaw PF. Creatine for the treatment of Depression. Biomolecules. 2019;9(9):406.
Duric V, Duman RS. Depression and treatment response: dynamic interplay of signaling pathways and altered neural processes. Cell Mol Life Sci. 2013;70(1):39–53.
Wu Y, Wang L, Hu K, Yu C, Zhu Y, Zhang S, et al. Mechanisms and therapeutic targets of depression after intracerebral hemorrhage. Front Psychiatry. 2018;9:682.
Duman RS, Voleti B. Signaling pathways underlying the pathophysiology and treatment of depression: novel mechanisms for rapid-acting agents. Trends Neurosci. 2012;35(1):47–56.
Cunha MP, Budni J, Ludka FK, Pazini FL, Rosa JM, Oliveira Á, et al. Involvement of PI3K/Akt signaling pathway and its downstream intracellular targets in the antidepressant-like effect of creatine. Mol Neurobiol. 2016;53(5):2954–68.
Croll S, Suri C, Compton D, Simmons M, Yancopoulos G, Lindsay R, et al. Brain-derived neurotrophic factor transgenic mice exhibit passive avoidance deficits, increased seizure severity and in vitro hyperexcitability in the hippocampus and entorhinal cortex. Neuroscience. 1999;93(4):1491–506.
Jankowsky JL, Patterson PH. Cytokine and growth factor involvement in long-term potentiation. Mol Cell Neurosci. 1999;14(4–5):273–86.
Schmidt HD, Duman RS. The role of neurotrophic factors in adult hippocampal neurogenesis, antidepressant treatments and animal models of depressive-like behavior. Behav Pharmacol. 2007;18(5–6):391–418.
Seitz F, Schmidt M, Shum C. Signals of extreme weather conditions in Central Europe in GRACE 4-D hydrological mass variations. Earth Planet Sci Lett. 2008;268(1–2):165–70.
Xu B. BDNF (I) rising from exercise. Cell Metab. 2013;18(5):612–4.
Yau SY, Li A, Hoo RL, Ching YP, Christie BR, Lee TM, et al. Physical exercise-induced hippocampal neurogenesis and antidepressant effects are mediated by the adipocyte hormone adiponectin. Proc Natl Acad Sci. 2014;111(44):15810–5.
Cunha MP, Pazini FL, Lieberknecht V, Rodrigues ALS. Subchronic administration of creatine produces antidepressant-like effect by modulating hippocampal signaling pathway mediated by FNDC5/BDNF/Akt in mice. J Psychiatr Res. 2018;104:78–87.
Li R-L, Wu S-S, Wu Y, Wang X-X, Chen H-Y, Xin J-J, et al. Irisin alleviates pressure overload-induced cardiac hypertrophy by inducing protective autophagy via mTOR-independent activation of the AMPK-ULK1 pathway. J Mol Cell Cardiol. 2018;121:242–55.
Yu Q, Kou W, Xu X, Zhou S, Luan P, Xu X, et al. FNDC5/Irisin inhibits pathological cardiac hypertrophy. Clin Sci. 2019;133(5):611–27.
Zhou X, Xu M, Bryant JL, Ma J, Xu X. Exercise-induced myokine FNDC5/irisin functions in cardiovascular protection and intracerebral retrieval of synaptic plasticity. Cell Biosci. 2019;9(1):32.
Cao RY, Zheng H, Redfearn D, Yang J. FNDC5: a novel player in metabolism and metabolic syndrome. Biochimie. 2019;158:111–6.
Kurdiova T, Balaz M, Vician M, Maderova D, Vlcek M, Valkovic L, et al. Effects of obesity, diabetes and exercise on Fndc5 gene expression and irisin release in human skeletal muscle and adipose tissue: in vivo and in vitro studies. J Physiol. 2014;592(5):1091–107.
Askari H, Rajani SF, Poorebrahim M, Haghi-Aminjan H, Raeis-Abdollahi E, Abdollahi M. A glance at the therapeutic potential of irisin against diseases involving inflammation, oxidative stress, and apoptosis: an introductory review. Pharmacol Res. 2018;129:44–55.
Aguiar AS Jr, Bristot V, Alves A, Cardoso L, Scheffer D. The role of PGC-1α/UCP2 signaling in the beneficial effects of physical exercise on the brain. Front Neurosci. 2019;13:292.
Maalouf GE, El Khoury D. Exercise-induced Irisin, the fat browning myokine, as a potential anticancer agent. J Obesity. 2019;2019:6561726.
Acknowledgements
We thank our colleagues at Royan Institute for their supportive discussions.
Funding
There is no funding to report.
Author information
Authors and Affiliations
Contributions
The design of study was done by FR, MHNE and KG, data mining was performed by FR, SG, LL, Interpretation of the obtained information was done by MHNE, TLM and K.G., The manuscript was written by FR, LL, SG and approved by MHNE, TLM and KG. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
There is no competing interests to report.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Rabiee, F., Lachinani, L., Ghaedi, S. et al. New insights into the cellular activities of Fndc5/Irisin and its signaling pathways. Cell Biosci 10, 51 (2020). https://doi.org/10.1186/s13578-020-00413-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13578-020-00413-3