
Abstract
Streptococcus suis serotype 2 is an important bacterial pathogen of swine, responsible for substantial economic losses to the swine industry worldwide. The knowledge on the pathogenesis of the infection caused by S. suis is still poorly known. It has been previously described that S. suis possesses at least one lipoprotein with double laminin and zinc (Zn)-binding properties, which was described in the literature as either laminin-binding protein (Lmb, as in the current study), lipoprotein 103, CDS 0330 or AdcAII. In the present study, the role of the Lmb in the pathogenesis of the infection caused by S. suis serotype 2 was dissected. Using isogenic mutants, results showed that Lmb does not play an important role in the laminin-binding activity of S. suis, even when clearly exposed at the bacterial surface. In addition, the presence of this lipoprotein does not influence bacterial adhesion to and invasion of porcine respiratory epithelial and brain endothelial cells and it does not increase the susceptibility of S. suis to phagocytosis. On the other hand, the Lmb was shown to play an important role as cytokine activator when tested in vitro with dendritic cells. Finally, this lipoprotein plays a critical role in Zn acquisition from the host environment allowing bacteria to grow in vivo. The significant lower virulence of the Lmb defective mutant may be related to a combination of a lower bacterial survival due to the incapacity to acquire Zn from their surrounding milieu and a reduced cytokine activation.
Similar content being viewed by others


Introduction
Streptococcus suis is one of the most important swine pathogens responsible for important economic losses, causing sudden death, meningitis, and arthritis among other clinical manifestations [1]. Moreover, S. suis is a zoonotic agent responsible for meningitis and septic shock in humans [1]. Although the knowledge on the pathogenesis of the infection caused by this pathogen has improved in recent years, our understanding on virulence factors and the pathogenesis of the infection remains incomplete [1]. A battery of surface proteins has been proposed as being important for virulence, although most of them have only been partially confirmed [2].
Bacterial attachment to host cells by mucosal pathogens like S. suis is frequently associated to the first steps of the pathogenesis, leading to colonization and infection [3, 4]. Bacterial adhesion and invasion of cells would be critical to allow successful infection of the host. Indeed, bacteria have evolved with several adhesins that specifically recognize and attach to host cell surface components, including extracellular matrix proteins, such as fibronectin, collagens, and laminin [2, 3]. Several laminin-binding proteins (Lmbs) expressed on the bacterial surface have been described for different pathogenic streptococci. Lmb was first identified in Streptococcus agalactiae (or group B Streptococcus) [5], and homologs of this lipoprotein were reported in other streptococci, such as Streptococcus pyogenes or Group A Streptococcus (named as Lbp or Lsp) [6, 7] and Streptococcus pneumoniae (called AdcAII, see below) [8]. Most studies demonstrated that recombinant Lmbs can bind purified laminin which is a large (900 kDa), highly glycosylated multidomain protein found in all human tissues [5, 7, 9,10,11]. In addition, in S. agalactiae, a mutant lacking Lmb showed decreased adherence to human laminin and reduced invasion of human brain microvascular endothelial cells [12]. Lsp-negative S. pyogenes mutants were shown to be defective in adhesion and invasion of epithelial cells in vitro and they were also highly attenuated in a murine subcutaneous ulcer model [7, 13].
Although many studies mostly focused on its function as an adhesin, Lmbs (or homologous proteins) may be also involved in zinc (Zn) uptake. Indeed, crystal structure analysis of Lmb homologs from different streptococci revealed that the two globular domains form a pocket to facilitate a tetrahedral geometry for Zn-binding [8, 11, 14]. Zinc is crucial in any normal metabolic condition, but during infection this element becomes particularly critical for bacterial pathogens [15, 16]. Indeed, both host and pathogen compete for the same essential metals, which are at very low concentration in infection sites [17]. In several pathogenic streptococci, deletion of one or more Zn-acquisition lipoproteins results in lower growth under Zn-restricted conditions as well as decreased virulence, adhesion, and biofilm formation, underlining the importance of zinc metabolism during colonization and infection.
In S. suis, an homologue of Lmb has been found in different strains [10] and a Lmb defective mutant was shown to be avirulent in a mouse model of infection [18]. In addition, lipoproteins involved in Zn-acquisition have also been described and reported to be crucial for S. suis virulence [19]. However, it seems to be some confusion in the literature concerning the proteins involved in both laminin adhesion and Zn uptake. Originally, a critical role as virulence factor of the so-called Zn-binding lipoprotein 103 was reported, without any mention of a possible role as a laminin-binding protein [18]. Later, Zhang et al. described a laminin-binding protein of S. suis (called CDS 0330) [10], without mentioning that this protein is the same Zn-binding lipoprotein protein 103 previously described by Aranda et al. [18]. Interestingly, both research groups independently reported this protein as being protective using a mouse model of infection [10, 18]. More recently, Zheng et al. reported that S. suis possesses two Zn-binding proteins, AdcA and AdcAII. Indeed, AdcAII corresponds to both lipoprotein 103 and the CDS 0330 previously described [10, 18, 19]; yet again, no mention of a potential laminin-binding ability of such protein was discussed [19].
In the present study, we further dissected the role of the Lmb/AdcAII/lipoprotein 103/CDS 0330 as both, laminin-binding and Zn-uptake protein in the pathogenesis of the infection caused by S. suis serotype 2.
Materials and methods
Bacterial strains and growth conditions
The strains and plasmids used in this study are listed in Table 1. The classical S. suis serotype 2 virulent European reference P1/7 strain (wild type) was used throughout this study including for construction of the isogenic deficient mutants [18]. S. suis strains were cultured in Todd Hewitt broth (THB; Becton Dickinson, Mississauga, ON, Canada) as previously described [20]. Escherichia coli strains, and different plasmids used in this study are also listed in Table 1. For in vitro cell culture assays, bacteria were prepared as previously described [21, 22] and resuspended in cell culture medium. When needed, antibiotics (Sigma-Aldrich, Oakville, ON, Canada) were added to the media at the following concentrations: for S. suis, spectinomycin (Spc) at 100 μg/mL; for E. coli, kanamycin (Km) and spectinomycin at 50 μg/mL and ampicillin (Ap) at 100 μg/mL.
DNA manipulations
Genomic DNA was extracted from the S. suis wild-type strain using InstaGene Matrix solution (BioRad Laboratories, Hercules, CA, USA). Mini preparations of recombinant plasmids were carried out using the QIAprep Spin Miniprep Kit (Qiagen, Valencia, CA, USA). Restriction enzymes and DNA-modifying enzymes (Fisher Scientific, Ottawa, ON, Canada) were used according to the manufacturer’s recommendations. Oligonucleotide primers (Table 2) were obtained from Integrated DNA Technologies (Coralville, IA, USA) and PCRs carried out with the iProof proofreading DNA polymerase (BioRad Laboratories, Mississauga, ON, Canada) or the Taq DNA polymerase (Qiagen). Amplification products were purified using the QIAquick PCR Purification Kit (Qiagen) and sequenced using an ABI 310 Automated DNA Sequencer and ABI PRISM Dye Terminator Cycle Sequencing Kit (Applied Biosystems, Carlsbad, CA, USA).
Construction of the laminin-binding defective/non-encapsulated double mutant
Precise in-frame deletion of cpsG gene from strain P1/7 Δlmb [18] was constructed using splicing-by-overlap-extension PCRs as previously described [27, 28]. Overlapping PCR product were cloned into pCR2.1 (Invitrogen, Burlington, ON, Canada), extracted with EcoRI, recloned into the thermosensitive E. coli–S. suis shuttle plasmid pSET4s, and digested with the same enzyme, giving rise to the knockout vector p4ΔcpsG. Electroporation of wild-type strain P1/7 procedures to obtain the mutant were previously described [25]. Allelic replacement was confirmed by PCR and DNA sequencing analyses. Amplification products were purified with the QIAgen PCR Purification Kit (Qiagen) and sequenced as described above.
Complementation of the laminin-binding defective mutant
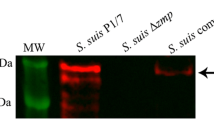
The pMX1 vector was used for the generation of recombinant plasmids for complementation analysis (Table 1). This vector is a derivative of the E. coli- S. suis shuttle cloning vector pSET2 [29] and possesses the S. suis malX promoter for transgene expression in S. suis. The entire lmb gene was amplified from genomic DNA of S. suis P1/7 strain and cloned into pMX1 via EcoRI and NcoI sites, generating complementation vector pMX1-lmb. This plasmid was introduced into E. coli MC1061 for verification of the sequence and then into the deletion mutant derived from S. suis P1/7 to construct lmb-complemented mutants.
Bacterial surface hydrophobicity assay
Relative surface hydrophobicity of the S. suis wild-type, Lmb and Lmb/non-encapsulated mutant strains was determined by measuring adsorption to n-hexadecane as previously described [30].
Microtiter plate laminin-binding assay
Laminin binding assay was performed as previously described [31]. Briefly, Maxisorp™ flat-bottom microtiter 96-well plates (Nunc, VWR, Mississauga, ON, Canada) were coated with 5 μg/mL of laminin from human placenta from Sigma Chemical Co. (St. Louis, MO, USA) in 0.1 M carbonate coating buffer (pH 9.6) and were incubated overnight at 4 °C. The plates were washed with PBS containing 0.05% (v/v) Tween 20 (PBST, pH 7.3), and 200 μL of 3% (w/v) non-fat dry milk in PBST was added to each well to prevent non-specific bacterial binding. After 1 h at 37 °C, the wells were washed with PBST. Formaldehyde-killed bacterial suspensions of wild-type or mutant strains were added (concentration equivalent to 108 CFU/mL), and the plates were incubated for 2 h at 37 °C. All unbound bacteria were subsequently removed by washing the wells with PBST. S. suis serotype 2-specific rabbit antiserum diluted 1/5000 in PBST prepared as previously described [31] was added to each well. This antiserum equally recognized both encapsulated and non-encapsulated serotype 2 S. suis as previously shown [31]. The plates were incubated for 1 h at 37 °C and wells were then washed, followed by the addition of horseradish peroxidase-labelled anti-rabbit IgG (Jackson Immunoresearch Laboratories Inc., West Grove, PA, USA) (diluted 1/8000 in PBST). The plates were incubated for 1 h at 37 °C with the secondary antibody. After washing, 3,3′,5,5′-tetramethylbenzidine (Zymed, San Francisco, CA, USA) was used as the enzyme substrate according to the manufacturer’s instructions. The reactions were stopped by adding 25 μL of H2SO4 (1 N) and were read at 450 nm. Uncoated wells served as background controls. Casein-coated wells served as a control for non-specific adhesion of S. suis to protein-coated wells. Each experiment was repeated at least 3 times.
Bacterial adhesion and invasion assays using porcine tracheal epithelial and brain microvascular endothelial cells
The neonatal porcine tracheal epithelial cell line (NPTr) and the porcine brain microvascular endothelial cell line (PBMEC) were used and cultured until confluence as previously described [32, 33]. Cells were infected with 1 × 106 CFU/well (multiplicity of infection (MOI) = 10) of the different S. suis strains and incubated for 2 h at 37 °C in 5% CO2. The adhesion assay, which quantifies total cell-associated bacteria (surface-adherent and intracellular bacteria), and invasion assay (using the antibiotic protection assay) were performed as previously described [32, 33].
S. suis virulence mouse model of systemic infection
A C57BL/6 J mouse model of infection was used. These studies were carried out in strict accordance with the recommendations of and approved by the University of Montreal Animal Welfare Committee guidelines and policies, including euthanasia to minimize animal suffering by the use of humane endpoints, applied throughout this study when animals were seriously affected (mortality was not an endpoint measurement). Thirty 6-week-old female C57BL/6 J (Jackson Research Laboratories, Bar Harbor, ME, USA) were used for these experiments (15 mice per group). Early stationary phase bacteria were washed twice in phosphate-buffered saline, pH 7.4, and resuspended in THB [34,35,36]. Bacterial cultures were appropriately diluted and plated on THB agar (THA) to accurately determine bacterial concentrations. Mice were inoculated with 1 × 107 CFU via the intraperitoneal route and health and behavior monitored at least thrice daily until 72 h post-infection and twice thereafter until the end of the experiment (12 days post-infection) for the development of clinical signs of sepsis, such as depression, swollen eyes, rough hair coat, prostration, and lethargy. For bacteremia studies, blood samples were collected from the caudal vein of surviving mice 12 h, 24 h and 48 h post-infection and plated as previously described [36].
Measurement of plasma (systemic) pro-inflammatory mediators
In addition, 8 mice per group were intraperitoneally mock-infected (THB) or infected with 1 × 107 CFU of wild-type or Δlmb mutant strains and blood samples were collected 12 h post-infection by intracardiac puncture following euthanasia and anti-coagulated with EDTA (Sigma-Aldrich) as previously described [36, 37]. Plasma supernatants were collected following centrifugation at 10 000 × g for 10 min at 4 °C and stored at −80 °C. The 12 h post-infection time point was selected to obtain maximal pro-inflammatory mediator production in the absence of significant mouse mortality as determined previously [36]. Plasmatic concentrations of interleukin (IL)-6 and C-X-C motif chemokine ligand (CXCL) 1 were quantified by sandwich. ELISA using pair-matched antibodies from R&D Systems (Minneapolis, MN, USA) according to the manufacturer’s recommendations.
Phagocytosis assay
J774A.1 murine macrophages (ATCC TIB-67; Rockville, MD, USA) were maintained in Dulbecco’s Modified Eagle’s Medium (Gibco, Burlington, ON, Canada) supplemented with 10% fetal bovine serum (Gibco) and grown at 37 °C with 5% CO2. Confluent cell cultures were scraped, seeded at 1 × 105 cells/mL, and incubated for 3 h at 37 °C with 5% CO2 to allow cell adhesion. Cells were infected by adding 1 × 107 CFU/mL of bacterial suspension in complete culture medium (MOI = 100), incubated for 2 h at 37 °C with 5% CO2, and phagocytosis assays performed as previously described using the antibiotic protection assay [21].
S. suis activation of marrow-derived dendritic cells (bmDC)
The femur and tibia from C57BL/6 J mice (Jackson Research Laboratories) were used to generate bmDCs, as previously described [21]. Briefly, hematopoietic bone marrow stem cells were cultured in complete culture medium (RPMI-1640 supplemented with 5% heat-inactivated fetal bovine serum, 10 mM HEPES, 2 mM L-glutamine, and 50 µM 2-mercaptoethanol (Gibco, Burlington, ON, Canada) and complemented with 20% granulocyte-macrophages colony-stimulating factor from mouse-transfected Ag8653 cells [38]. Cell purity was confirmed to be at least 90% CD11c + by flow cytometry as previously described [21]. Albeit this culture system cannot completely rule out the presence of other innate cells such as macrophages, it represents an enriched source of bmDCs [39].
All experiments were performed in the absence of endotoxin (lipopolysaccharide) contamination and under non-toxic conditions (data not shown), the latter being evaluated by the lactate dehydrogenase release with the CytoTox 96® Non-Radioactive Cytotoxicity Assay (Promega, Madison, WI, USA). Since the P1/7Δlmb mutant strain did not grow in the RPMI-1640 complete medium, heat-killed S. suis suspensions (prepared as previously described [40]) of all tested strains were used for bmDC stimulation at a concentration equivalent to 2 × 109 CFU/mL. Cells were resuspended at 1 × 106 cells/mL in complete medium and stimulated with the different heat-killed S. suis strains. Supernatants were collected at 4 h, 6 h, 8 h, 12 h and 16 h following stimulation with heat-kill S. suis, incubation times at which secreted cytokine levels were maximal in the absence of cytotoxicity (data not shown) [20, 21]. Mock-infected cells served as negative controls. Secreted levels of tumor necrosis factor (TNF), interleukin (IL)-6, C-C motif chemokine ligand (CCL) 3, and C-X-C motif chemokine ligand (CXCL) 1 were quantified by sandwich ELISA using pair-matched antibodies from R&D Systems (Minneapolis, MN, USA) according to the manufacturer’s recommendations.
Bacterial growth analysis
Bacterial growth experiments were performed in microtubes (500-μL culture volume). A bacterial overnight culture grown (1 × 104 CFU/mL) in THB was used to inoculate THB, plasma naturally poor in zinc, plasma with various ZnSO4 concentrations (from 0 to 50 μM) and plasma with ZnSO4 and the addition of the appropriate concentration of the chelating agent Tetrakis-(2-Pyridylmethyl) ethylenediamine (TPEN, Sigma Aldrich). Growth was followed during 24 h of incubation at 37 °C. The total numbers of CFU/mL were evaluated at different incubation times.
Statistical analyses
Normality of data was verified using the Shapiro–Wilk test. Accordingly, parametric (unpaired t-test) or non-parametric tests (Mann-Whitney rank sum test), where appropriate, were performed to evaluate statistical differences between groups. Log-rank test was used to compare survival rates between wild-type-infected mice and those infected with mutant strains. Each in vitro test was repeated in at least three independent experiments. p < 0.05 was considered as statistically significant.
Results
Surface hydrophobicity of the strains included in this study
Previous studies have demonstrated that deletion of either cpsG or cpsF biosynthesis genes from S. suis results in a non-encapsulated phenotype [30, 41]. An isogenic mutant in which the cpsG gene, encoding a glycosyltransferase, was deleted from the P1/7 Δlmb strain was constructed and compared to the wild-type P1/7, P1/7 Δlmb and P1/7 ΔcpsF. Surface hydrophobicity of P1/7 Δlmb was low (less than 6%) and comparable to that of wild-type P1/7 (Additional file 1). Meanwhile, and as expected, deletion of the cpsG gene significantly increased surface hydrophobicity, with similar values to those obtained with P1/7 ΔcpsF non-encapsulated mutant (Additional file 1). These results indicate that the Lmb does not play any role in the surface hydrophobicity of the strain tested.
Lack of lipoprotein Lmb does not affect S. suis-binding to laminin
Although previous studies revealed that purified Lmb of S. suis could bind human laminin [31], the ability of a Δlmb mutant to bind immobilized human placental laminin was not previously evaluated. Binding capacity to laminin of the Δlmb mutant was comparable to that of the wild-type strain (Figure 1). It was also confirmed that a non-encapsulated mutant strain is able to bind laminin more efficiently than the wild-type strain, as previously described [31]. Interestingly, adhesion to laminin of a double mutant Δlmb/non-encapsulated (Δlmb/ΔcpsG) was similar to that observed with the single non-encapsulated ΔcpsF mutant (Figure 1). Taken, together, these results show that Lmb would not be critical for the interaction between S. suis and laminin, even in the absence of the capsular polysaccharide (CPS).

The Δlmb mutant is not impaired in its capacity of adhesion to laminin. Adhesion to laminin of the wild-ype P1/7 strain (dark blue), Δlmb (light blue), ΔcpsF (dark brown) and Δlmb/ΔcpsG (light brown) mutant strains as evaluated by ELISA. Data represent the optical density (O.D.)450 mean ± SEM from at least three independent experiments. *Indicates a significant difference (p < 0.05).
Lack of lipoprotein Lmb does not impair the adhesion to and invasion of epithelial and endothelial swine cells
The role of the Lmb protein in the adhesion/invasion to NPTr and PBMEC cells was evaluated using the generated mutant strains. For both cell types, and as expected [42, 43], the non-encapsulated mutant strain significantly adhered and invaded cells more efficiently than the wild-type strain (Figures 2A–D). However, the Δlmb mutant similarly adhered and (weakly) invaded NPTr (Figures 2A, B) and PBMEC (Figures 2C, D) cells. Moreover, the Δlmb/ΔcpsG double mutant adhered and invaded the two cell types similarly to the non-encapsulated ΔcpsF mutant. These results indicate that the Lmb does not play a critical role in the adhesion/invasion of the epithelial and endothelial cells tested, even in the absence of the CPS.

The Δlmb mutant is not impaired in its capacity of adhesion to and invasion of epithelial and endothelial cells. Adhesion to and invasion of NPTr (A, C) and PBMEC (B, D) of wild-type P1/7 strain (dark blue), Δlmb (light blue), ΔcpsF (dark brown) and Δlmb/ΔcpsG (light brown) mutant strains after 2 h of incubation. *Indicates a significant difference (p < 0.05). Each bar represents the mean bacterial concentration (CFU/mL)+SEM from at least three independent experiments.
Presence of the lipoprotein Lmb is required for full virulence of S. suis
To confirm the role of Lmb in S. suis virulence and development of clinical disease, a well-characterized C56BL/6 mouse model of infection was used [36, 44]. Wild-type strain-infected mice rapidly developed clinical signs of systemic disease characteristic of septic shock with 60% of mice succumbing to infection within 48 h (Figure 3A). On the other hand, only 20% of mice infected with the Δlmb mutant succumbed to disease (Figure 3A). These mice developed transient signs of infection such as rough coat hair following inoculation of bacteria and rapidly recovered and presented a normal behavior. These results confirm that the Lmb lipoprotein is crucial for the virulence of S. suis serotype 2 strain P1/7.

Presence of the lipoprotein Lmb is important for S. suis systemic virulence and blood persistence following intraperitoneal inoculation. Survival (A) and blood bacterial burden at 12, 24 and 48 h post-infection (B) of C57BL/6 mice following intraperitoneal inoculation of the S. suis virulent wild-type P1/7 strain (dark blue) and Δlmb mutant strain (light blue). Data represent survival curves (A) (n = 15) or geometric mean (B) (n = survived mice at each time point). *(p < 0.05) indicates a significant difference between survival or blood bacterial burden of mice infected either with the wild-type or the Δlmb mutant strain.
Blood bacterial burden was also evaluated at the early infection times of 12 h, 24 h and 48 h (Figure 3B). Twelve hours following infection, mice infected with the wild-type strain presented elevated blood bacterial burdens averaging 1 × 107 CFU/mL (Figure 3B). Moreover, infection with the wild-type strain resulted in elevated bacterial burdens after 24 h and 48 h infection. On the other hand, mice infected with the Δlmb mutant show a significant decrease in blood bacterial burden at 12, 24 h and 48 h post-infection. This difference would have been more evident at 48 h after infection considering that almost 60% of mice infected with the wild-type P1/7 strain were already dead and the viable counts were performed with survived (less affected) mice only. These data indicate that the Δlmb mutant strain presents less survival capacity within the bloodstream of infected mice.
In addition, the inflammatory response of animals infected with the P1/7 and Δlmb mutant strain was evaluated. IL-6 and CXCL1 concentration were measured after 12 h of infection. The concentrations of these plasma mediators were significantly lower for the Δlmb mutant strain when compared to P1/7 wild-type strain (Figure 4).

Absence of the lipoprotein Lmb significantly decreases the cytokine levels in mouse plasma from infected animals. Plasma levels of IL-6 (A) and CXCL1 (B) in mice 12 h following intraperitoneal inoculation of the S. suis serotype 2 wild-type strain P1/7 (dark blue) or Δlmb mutant strain (light blue). Data represent mean ± SEM (n = 8). *p < 0.05 indicates a significant difference between plasma levels of mice infected with the mutant strain when compared to the wild-type P1/7 strain.
Lmb does not promote S. suis resistance to phagocytosis by murine macrophages
One of the hypotheses that may explain that the Δlmb mutant survives less in the bloodstream of infected mice is an increased susceptibility to phagocytosis. In vitro phagocytosis studies with murine macrophages showed that the wild-type strain was less phagocytosed than the non-encapsulated ΔcpsF mutant, as previously described [30] (Figure 5). The Δlmb mutant was internalized similarly to the wild-type P1/7 strain, whereas the Δlmb/ΔcpsG double mutant was phagocytosed similarly to the non-encapsulated ΔcpsF mutant. These results indicate that the lipoprotein Lmb does not play any role in the resistance to the phagocytosis.

The lipoprotein Lmb is not involved in S. suis phagocytosis resistance to J774A.1 murine macrophages. Internalization of the P1/7 wild-type strain (dark blue), Δlmb (light blue), ΔcpsF (dark brown) and Δlmb/ΔcpsG (light brown) mutant strains after 2 h of incubation. * (p < 0.05) indicate a significant difference between the wild-type and ΔcpsF mutant and a significant difference between the Δlmb and Δlmb/ΔcpsG mutants. Each bar represents the mean bacterial concentration (CFU/mL) + SEM from at least three independent experiments.
The lipoprotein Lmb partially regulates bmDC release of cytokines
The role of the Lmb lipoprotein in cytokine release was evaluated using bmDCs as an innate immune cell model, given that dendritic cells play a critical role during S. suis pathogenesis and that their inflammatory response to S. suis has been well-characterized [20, 21]. bmDCs were activated for up to 16 h with heat-killed bacteria. For all experiments and at all incubation times, control mock infected cells presented negligible cytokine values < 300 pg/mL (not shown). The Δlmb and Δlmb/cpsG mutant strains induced significant lower levels of IL-6 and TNF (Figures 6A, C) and, to a lesser extent, CXCL1 (Figure 6B), when compared to wild-type P1/7 and non-encapsulated ΔcpsF mutant strains, respectively (Figure 6). The complemented Δlmb:: pMX1-lmb strain restored the capacity of cytokine induction of the Δlmb mutant to values similar to those of the wild-type strain (Figure 6). On the other hand, the presence of Lmb did not affect the induction of CCL3 (Figure 6D). These results indicate that the Lmb lipoprotein is one of the S. suis components responsible for cytokine release.

Presence of the lipoprotein Lmb modulates S. suis induced dendritic cell (DC) cytokine production. Pro-inflammatory cytokine production by bmDCs following activation with heat killed bacteria of the wild-type P1/7 strain (dark blue), Δlmb (light blue), Δlmb:: pMX1-lmb (grey), ΔcpsF (dark brown) and Δlmb/ΔcpsG (light brown) mutant strains. Production of IL-6 (A), CXCL1 (B), TNF (C) and CCL3 (D). Data represent the mean+SEM of at least three independent experiments. *Indicates a significant difference (p < 0.05).
The lipoprotein Lmb is essential for S. suis growth under zinc starvation conditions
Considering the limited role observed for the Lmb as laminin-binding protein, the second characteristic already described in the literature for this lipoprotein (Zn uptake) was evaluated. First, the wild-type P1/7 and Δlmb strains cultured in rich media (THB) presented similar good growth rate, as evaluated by bacterial counts (Figure 7A). In contrast, when cultured in plasma (a poor medium that mimic in vivo conditions), growth of the Δlmb mutant strain was seriously reduced when compared to the wild-type strain (Figure 7B). The complemented Δlmb mutant completely restored the growth to the levels of the wild-type strain. To investigate if the loss of Lmb was associated with a reduced ability to grow under conditions of zinc starvation, ZnSO4 was added to the plasma at concentrations of 25 and 50 µM (Figure 7C). Growth of Δlmb mutant was partially and totally restored with 25 µM and 50 µM ZnSO4, respectively, indicating a dose-effect of zinc concentration (Figure 7C). Finally, the chelating agent N,N,N’,N’-Tetrakis-(2-pyridylmethyl)ethylenediamine (TPEN), a cell permeable high-affinity Zn2+ chelator used to reduce concentration of zinc in zinc homeostasis studies (15, 49, 51) was added to the plasma supplemented with ZnSO4 (Figure 7C) [13, 45, 46]. Addition of 20 μM TPEN in plasma supplemented with 50 µM ZnSO4 impaired the growth of the Δlmb mutant strain (Figure 7D).

Growth of Δlmb mutant strain under different conditions. Growth of the wild-type P1/7 strain (dark blue), Δlmb (light blue) and Δlmb::pMX1-lmb (grey, plasma only) in THB (A), plasma (B), plasma supplemented with 25 µM ZnSO4 (pink dash) or 50 µM ZnSO4 (pink dot) (C) and plasma supplemented with 50 µM ZnSO4 (pink dot) added with 20 µM TPEN (green). *(p < 0.05) indicate a significant difference between the wild-type and Δlmb mutant in plasma, between Δlmb mutant with and without supplementation of ZnSO4 in plasma or between Δlmb mutant with supplementation of ZnSO4 and with supplementation of ZnSO4 with addition of TPEN. Each point represents mean bacterial concentration (CFU/mL) + / − SEM of at least three different independent experiments.
Discussion
The pathogenesis of the infection caused by S. suis is only partially known and the definition of critical virulence factors is controversial [2]. During the first steps of the infection, the interaction of microorganisms with extracellular matrix proteins can promote bacterial colonization and invasion [47]. It has been shown that S. suis is able to interact with several of these proteins, including laminin [31]. On the other hand, in order for bacterial pathogens to survive inside their hosts, they need to efficiently acquire and use the available nutrients from their surrounding medium. One of these components playing an important role in the host-pathogen interplay is zinc (Zn) [48]. Zn-binding lipoproteins have also been described in S. suis as being critically important for bacterial fitness and survival during the infection [19]. Interestingly, one of these S. suis laminin- and Zn-binding proteins is a single lipoprotein with a double function previously identified as lipoprotein 103, Lmb CDS 0330 or AdcAII [18, 19]. This protein (or genes coding for it) presents high homology with that of other streptococci [10], in which this double function has also been described [5, 7, 13, 46]. In the case of S. suis, it is not clear whether the laminin-binding or the Zn-binding capacities of this protein (or both) plays an important role during the infection. In the present study, the more precise role of this lipoprotein (called Lmb in the current study for simplicity purposes) in the pathogenesis of the infection has been studied.
Based on the previous observation that some S. suis strains are able to bind laminin [31], Zhang et al. showed that S. suis possesses a lmb gene which is homologous to those of S. pyogenes, S. agalactiae and S. pneumoniae and the cloned and purified protein clearly adhered to laminin in vitro. However, it is not clear if this lipoprotein is mainly responsible for the binding of S. suis to laminin. Results from the current study using a Lmb-defective mutant indicate that the Lmb does not play a critical role in the bacterial binding to this extracellular matrix protein. Although Lmb was reported as a surface protein [10], it was previously described that the presence of the CPS may prevent S. suis to attach to laminin by hindering surface proteins [31]. Hence, a double mutant without Lmb and CPS was constructed and tested. The ΔcpsF non-encapsulated mutant adhered significantly more to laminin when compared to the wild-type strain confirming previous results. However, the double Δlmb/ΔcpsG mutant adhered similarly to the non-encapsulated mutant, indicating that even in the absence of CPS the surface expressed Lmb does not seem to play an important role in S. suis adhesion to the laminin. The presence of additional proteins able to bind laminin that have been described as being present in S. suis serotype 2 may compensate the absence of Lmb [49, 50].
The role of the Lmb in the adhesion/invasion of host cells seems to vary among different streptococci. For example, the presence of the Lmb seems to be important for S. agalactiae colonization of the epithelium and subsequent translocation into the bloodstream, having a tropism for the central nervous system [51]. The Lbp from S. pneumoniae has also been reported to play an important role in the adhesion of the pathogen to human microvascular endothelial cells [52] and S. pyogenes Δlmb (or Lsp, or Lbp) mutants show a decrease adhesion only [6] or a decrease in adhesion and invasion [7] to host cells. Results of the current study showed that the Lmb does not play any major role in S. suis adhesion to or invasion of swine respiratory epithelial and brain microvascular endothelial cells, which correlates with the fact that this protein is not critical for laminin binding. As mentioned, other proteins playing similar overlapped functions may compensate and allow S. suis to adhere to this extracellular matrix protein and host cells.
There are several studies on the role of a Lmb (or homologous proteins) as Zn-binding proteins of streptococci [5,6,7,8], with the exception of that of S. pneumoniae which plays an important role as a Zn-binding lipoprotein but it does not bind laminin [53]. As mentioned, two Zn-binding proteins have been described in S. suis: AdcA and AdcAII [19]. Although both lipoproteins may play a role in Zn-binding, Zhang et al. reported that AdcAII expression is significantly more up-regulated than AdcA, but only the absence of both lipoproteins affected survival under Zn-restricted conditions and virulence in a mouse model of infection [19]. Results of the current study confirm only partially such observations. Indeed, the Δlmb mutant (which corresponds to the ΔAdcAII previously described) could not grow under Zn-restricted conditions (plasma) and the addition of Zn restored this defect. In addition, the complemented-mutant was able to grow at similar levels than the wild-type strain. In addition, and also different from the study of Zhang et al. [19], in the current work, the Δlmb mutant presented a markedly lower virulence than the wild-type strain, confirming previous results reported by Aranda et al. [18], who used mice from a different background (BALB/c). The critical role of ΔAdcAII has also been observed in S. pneumoniae (16), whereas both ΔAdcA and ΔAdcAII presented less virulence than a wild-type strain of Group A Streptococcus [54]. In our hands, the presence of ΔAdcAII is critical for Zn uptake and virulence in S. suis serotype 2. Differences observed among the S. suis studies might be explained by the background of the S. suis strains used: the current study and that of Aranda et al. [18] used the virulent P1/7 sequence type (ST) 1 as wild-type reference strain, whereas Zhang et al. [19] used a ST7 highly virulent strain from China. Previous studies have also reported some differences within S. suis serotype 2 strains from different background in the pathogenesis of the infection [22, 55]. It may be hypothesized that regulation of Zn may vary depending on the virulence/phenotype of the strain.
The lower virulence of the Δlmb mutant can be explained by a lower survival (lower bacteremia) in vivo as shown in the current study. This would be directly related to the in-vivo Zn-restricted conditions rather to a higher susceptibility of the mutant to bacterial killing. Indeed, no difference in the phagocytosis rate between the Δlmb mutant and the wild-type strains could be observed. In addition, induction of production of large amounts of pro-inflammatory mediators leading to exacerbated inflammation is a hallmark of S. suis infections and is responsible, at least in part, for host death [56]. Although the current study clearly showed a lower cytokine concentration in mice infected with the Δlmb mutant, this effect might be the simple consequence of lower bacteremia. However, in addition to this, in vitro results showed that the presence of the Lmb is also important as cytokine activator. Similarly, the laminin-binding protein from S. pneumoniae (Lbp) was reported to activate human brain microvascular-endothelial cells [52], whereas that of S. agalactiae was not involved in IL-8 release when using the same cells [12]. As lipoproteins have been clearly shown to be responsible for the cytokine activation of cells by S. suis [57, 58], it can also be hypothesized that the lower cytokine production in vivo by the Δlmb mutant is a double combination of lower bacteremia and reduced activation due to the absence of this lipoprotein.
In summary, results from the current study showed that Lmb does not play an important role in the laminin-binding activity of S. suis serotype 2. In addition, the presence of this lipoprotein does not influence bacterial adhesion to and invasion of porcine respiratory epithelial and brain endothelial cells and it does not increase the susceptibility of S. suis serotype 2 to phagocytosis. On the other hand, this lipoprotein was shown to play an important role as cytokine activator when tested in vitro with dendritic cells. Finally, the Lmb plays a critical role in Zn acquisition from the host environment allowing bacteria to grow in vivo. The significant lower virulence of the Lmb defective mutant may be related to a combination of a lower bacterial survival due to the incapacity to acquire and use Zn from their surrounding milieu and a reduced cytokine activation in the absence of this lipoprotein. Since the gene coding for this protein is also present in other serotypes of S. suis (data from GeneBank, not shown), it would be interesting to confirm if the role of the Lmb described in the current study applies also to other pathogenic serotypes of S. suis.
Abbreviations
- Ap:
-
ampicillin
- CFU:
-
colony-forming unit
- CCL:
-
C-C motif chemokine ligand
- CXCL:
-
C-X-C motif chemokine ligand
- DC:
-
dendritic cell
- ELISA:
-
enzyme-linked immunosorbent assay
- Ig:
-
immunoglobulin
- IL:
-
interleukin
- Km:
-
kanamycin
- LB:
-
Luria-Bertani
- Lmb:
-
laminin binding protein
- Nptr:
-
Neonatal Porcine Tracheal Epithelial cell
- PBMEC:
-
Porcine Brain Microvascular Endothelial cell
- PCR:
-
polymerase chain reaction
- ST:
-
sequence type
- THA:
-
THB agar
- THB:
-
Todd Hewitt broth
- TNF:
-
tumor necrosis factor
- Zn:
-
zinc
- TPEN:
-
Tetrakis-(2-Pyridylmethyl) ethylenediamine
References
Gottschalk M, Xu J, Calzas C, Segura M (2010) Streptococcus suis: a new emerging or an old neglected zoonotic pathogen? Future Microbiol 5:371–391
Segura M, Fittipaldi N, Calzas C, Gottschalk M (2017) Critical Streptococcus suis virulence factors: are they all really critical? Trends Microbiol 25:585–599
Segura M, Calzas C, Grenier D, Gottschalk M (2016) Initial steps of the pathogenesis of the infection caused by Streptococcus suis: fighting against nonspecific defenses. FEBS Lett 590:3772–3799
Jiang Q, Zhou X, Cheng L, Li M (2019) The adhesion and invasion mechanisms of Streptococci. Curr Issues Mol Biol 32:521–560
Spellerberg B, Rozdzinski E, Martin S, Weber-Heynemann J, Schnitzler N, Lutticken R, Podbielski A (1999) Lmb, a protein with similarities to the LraI adhesin family, mediates attachment of Streptococcus agalactiae to human laminin. Infect Immun 67:871–878
Terao Y, Kawabata S, Kunitomo E, Nakagawa I, Hamada S (2002) Novel laminin-binding protein of Streptococcus pyogenes, Lbp, is involved in adhesion to epithelial cells. Infect Immun 70:993–997
Elsner A, Kreikemeyer B, Braun-Kiewnick A, Spellerberg B, Buttaro BA, Podbielski A (2002) Involvement of Lsp, a member of the LraI-lipoprotein family in Streptococcus pyogenes, in eukaryotic cell adhesion and internalization. Infect Immun 70:4859–4869
Loisel E, Jacquamet L, Serre L, Bauvois C, Ferrer JL, Vernet T, Di Guilmi AM, Durmort C (2008) AdcAII, a new pneumococcal Zn-binding protein homologous with ABC transporters: biochemical and structural analysis. J Mol Biol 381:594–606
Kikkawa Y, Sanzen N, Sekiguchi K (1998) Isolation and characterization of laminin-10/11 secreted by human lung carcinoma cells: laminin-10/11 mediates cell adhesion through integrin α3β1. J Biol Chem 273:15854–15859
Zhang YM, Shao ZQ, Wang J, Wang L, Li X, Wang C, Tang J, Pan X (2014) Prevalent distribution and conservation of Streptococcus suis Lmb protein and its protective capacity against the Chinese highly virulent strain infection. Microbiol Res 169:395–401
Linke C, Caradoc-Davies TT, Young PG, Proft T, Baker EN (2009) The laminin-binding protein Lbp from Streptococcus pyogenes is a zinc receptor. J Bacteriol 191:5814–5823
Tenenbaum T, Spellerberg B, Adam R, Vogel M, Kim KS, Schroten H (2007) Streptococcus agalactiae invasion of human brain microvascular endothelial cells is promoted by the laminin-binding protein Lmb. Microbes Infect 9:714–720
Weston BF, Brenot A, Caparon MG (2009) The metal homeostasis protein, Lsp, of Streptococcus pyogenes is necessary for acquisition of zinc and virulence. Infect Immun 77:2840–2848
Ragunathan P, Spellerberg B, Ponnuraj K (2009) Structure of laminin-binding adhesin (Lmb) from Streptococcus agalactiae. Acta Crystallogr D Biol Crystallogr 65:1262–1269
Braymer JJ, Giedroc DP (2014) Recent developments in copper and zinc homeostasis in bacterial pathogens. Curr Opin Chem Biol 19:59–66
Gammoh NZ, Rink L (2017) Zinc in infection and inflammation. Nutrients 9:624
Capdevila DA, Wang J, Giedroc DP (2016) Bacterial strategies to maintain zinc metallostasis at the host-pathogen interface. J Biol Chem 291:20858–20868
Aranda J, Teixido L, Fittipaldi N, Cortes P, Llagostera M, Gottschalk M, Barbe J (2012) Inactivation of the gene encoding zinc-binding lipoprotein 103 impairs the infectivity of Streptococcus suis. Can J Vet Res 76:72–76
Zheng C, Qiu J, Zhao X, Yu S, Wang H, Wan M, Wei M, Jiao X (2022) The AdcR-regulated AdcA and AdcAII contribute additively to zinc acquisition and virulence in Streptococcus suis. Vet Microbiol 269:109418
Lecours MP, Segura M, Fittipaldi N, Rivest S, Gottschalk M (2012) Immune receptors involved in Streptococcus suis recognition by dendritic cells. PLoS One 7:e44746
Lecours MP, Gottschalk M, Houde M, Lemire P, Fittipaldi N, Segura M (2011) Critical role for Streptococcus suis cell wall modifications and suilysin in resistance to complement-dependent killing by dendritic cells. J Infect Dis 204:919–929
Auger J-P, Santinón A, Roy D, Mossman K, Xu J, Segura M, Gottschalk M (2017) Type I interferon induced by Streptococcus suis serotype 2 is strain-dependent and may be beneficial for host survival. Front Immunol 8:1039
Slater JD, Allen AG, May JP, Bolitho S, Lindsay H, Maskell DJ (2003) Mutagenesis of Streptococcus equi and Streptococcus suis by transposon Tn917. Vet Microbiol 93:197–206
Casadaban MJ, Cohen SN (1980) Analysis of gene control signals by DNA fusion and cloning in Escherichia coli. J Mol Biol 138:179–207
Takamatsu D, Osaki M, Sekizaki T (2001) Thermosensitive suicide vectors for gene replacement in Streptococcus suis. Plasmid 46:140–148
Okura M, Osaki M, Fittipaldi N, Gottschalk M, Sekizaki T, Takamatsu D (2011) The minor pilin subunit Sgp2 is necessary for assembly of the pilus encoded by the srtG cluster of Streptococcus suis. J Bacteriol 193:822–831
Warrens AN, Jones MD, Lechler RI (1997) Splicing by overlap extension by PCR using asymmetric amplification: an improved technique for the generation of hybrid proteins of immunological interest. Gene 186:29–35
Auger J-P, Chuzeville S, Roy D, Mathieu-Denoncourt A, Xu J, Grenier D, Gottschalk M (2017) The bias of experimental design, including strain background, in the determination of critical Streptococcus suis serotype 2 virulence factors. PLoS One 12:e0181920
Takamatsu D, Osaki M, Sekizaki T (2001) Construction and characterization of Streptococcus suis-Escherichia coli shuttle cloning vectors. Plasmid 45:101–113
Auger JP, Dolbec D, Roy D, Segura M, Gottschalk M (2018) Role of the Streptococcus suis serotype 2 capsular polysaccharide in the interactions with dendritic cells is strain-dependent but remains critical for virulence. PLoS One 13:e0200453
Esgleas M, Lacouture S, Gottschalk M (2005) Streptococcus suis serotype 2 binding to extracellular matrix proteins. FEMS Microbiol Lett 244:33–40
Wang Y, Gagnon CA, Savard C, Music N, Srednik M, Segura M, Lachance C, Bellehumeur C, Gottschalk M (2013) Capsular sialic acid of Streptococcus suis serotype 2 binds to swine influenza virus and enhances bacterial interactions with virus-infected tracheal epithelial cells. Infect Immun 81:4498–4508
Vanier G, Segura M, Friedl P, Lacouture S, Gottschalk M (2004) Invasion of porcine brain microvascular endothelial cells by Streptococcus suis serotype 2. Infect Immun 72:1441–1449
Dominguez-Punaro M, Segura M, Plante MM, Lacouture S, Rivest S, Gottschalk M (2007) Streptococcus suis serotype 2, an important swine and human pathogen, induces strong systemic and cerebral inflammatory responses in a mouse model of infection. J Immunol 179:1842–1854
Dominguez-Punaro M, Segura M, Radzioch D, Rivest S, Gottschalk M (2008) Comparison of the susceptibilities of C57BL/6 and A/J mouse strains to Streptococcus suis serotype 2 infection. Infect Immun 76:3901–3910
Auger JP, Fittipaldi N, Benoit-Biancamano MO, Segura M, Gottschalk M (2016) Virulence studies of different sequence types and geographical origins of Streptococcus suis Serotype 2 in a mouse model of infection. Pathogens 5:48
Lachance C, Gottschalk M, Gerber PP, Lemire P, Xu J, Segura M (2013) Exacerbated type II interferon response drives hypervirulence and toxic shock by an emergent epidemic strain of Streptococcus suis. Infect Immun 81:1928–1939
Segura M, Su Z, Piccirillo C, Stevenson MM (2007) Impairment of dendritic cell function by excretory-secretory products: a potential mechanism for nematode-induced immunosuppression. Eur J Immunol 37:1887–1904
Lavagna A, Auger JP, Girardin SE, Gisch N, Segura M, Gottschalk M (2020) Recognition of lipoproteins by toll-like receptor 2 and DNA by the AIM2 inflammasome is responsible for production of interleukin-1β by virulent suilysin-negative Streptococcus suis serotype 2. Pathogens 9:147
Segura M, Stankova J, Gottschalk M (1999) Heat-killed Streptococcus suis capsular type 2 strains stimulate tumor necrosis factor alpha and interleukin-6 production by murine macrophages. Infect Immun 67:4646–4654
Zhang Y, Ding D, Liu M, Yang X, Zong B, Wang X, Chen H, Bei W, Tan C (2016) Effect of the glycosyltransferases on the capsular polysaccharide synthesis of Streptococcus suis serotype 2. Microbiol Res 185:45–54
Auger JP, Payen S, Roy D, Dumesnil A, Segura M, Gottschalk M (2019) Interactions of Streptococcus suis serotype 9 with host cells and role of the capsular polysaccharide: comparison with serotypes 2 and 14. PLoS ONE 14:e0223864
Vanier G, Segura M, Gottschalk M (2007) Characterization of the invasion of porcine endothelial cells by Streptococcus suis serotype 2. Can J Vet Res 71:81–89
Dominguez-Punaro Mde L, Segura M, Radzioch D, Rivest S, Gottschalk M (2008) Comparison of the susceptibilities of C57BL/6 and A/J mouse strains to Streptococcus suis serotype 2 infection. Infect Immun 76:3901–3910
Shumaker DK, Vann LR, Goldberg MW, Allen TD, Wilson KL (1998) TPEN, a Zn2+/Fe2+ chelator with low affinity for Ca2+, inhibits lamin assembly, destabilizes nuclear architecture and may independently protect nuclei from apoptosis in vitro. Cell Calcium 23:151–164
Tedde V, Rosini R, Galeotti CL (2016) Zn2+ uptake in Streptococcus pyogenes: characterization of adcA and lmb null mutants. PLoS One 11:e0152835
Tomlin H, Piccinini AM (2018) A complex interplay between the extracellular matrix and the innate immune response to microbial pathogens. Immunology 155:186–201
Shafeeq S, Kuipers OP, Kloosterman TG (2013) The role of zinc in the interplay between pathogenic streptococci and their hosts. Mol Microbiol 88:1047–1057
Li Q, Liu H, Du D, Yu Y, Ma C, Jiao F, Yao H, Lu C, Zhang W (2015) Identification of novel laminin- and fibronectin-binding proteins by Far-Western blot: capturing the adhesins of Streptococcus suis type 2. Front Cell Infect Microbiol 5:82
Li Q, Fu Y, He Y, Zhang Y, Qian Y, Yu Y, Yao H, Lu C, Zhang W (2017) Fibronectin-/fibrinogen-binding protein (FBPS) is not a critical virulence factor for the Streptococcus suis serotype 2 strain ZY05719. Vet Microbiol 208:38–46
Shabayek S, Spellerberg B (2018) Group B streptococcal colonization, molecular characteristics, and epidemiology. Front Microbiol 9:437
Jimenez-Munguia I, Tomeckova Z, Mochnacova E, Bhide K, Majerova P, Bhide M (2021) Transcriptomic analysis of human brain microvascular endothelial cells exposed to laminin binding protein (adhesion lipoprotein) and Streptococcus pneumoniae. Sci Rep 11:7970
Brown LR, Gunnell SM, Cassella AN, Keller LE, Scherkenbach LA, Mann B, Brown MW, Hill R, Fitzkee NC, Rosch JW, Hill R, Fitzkee NC, Rosch JW, Tuomanen EI, Thornton JA (2016) AdcAII of Streptococcus pneumoniae affects pneumococcal invasiveness. PLoS One 11:e0146785
Makthal N, Do H, Wendel BM, Olsen RJ, Helmann JD, Musser JM, Kumaraswami M (2020) Group A Streptococcus AdcR regulon participates in bacterial defense against host-mediated zinc sequestration and contributes to virulence. Infect Immun 88:e00097-e120
Lavagna A, Auger JP, Dumesnil A, Roy D, Girardin SE, Gisch N, Segura M, Gottschalk M (2019) Interleukin-1 signaling induced by Streptococcus suis serotype 2 is strain-dependent and contributes to bacterial clearance and inflammation during systemic disease in a mouse model of infection. Vet Res 50:52
Fittipaldi N, Segura M, Grenier D, Gottschalk M (2012) Virulence factors involved in the pathogenesis of the infection caused by the swine pathogen and zoonotic agent Streptococcus suis. Future Microbiol 7:259–279
Gisch N, Auger JP, Thomsen S, Roy D, Xu J, Schwudke D, Gottschalk M (2018) Structural analysis and immunostimulatory potency of lipoteichoic acids isolated from three Streptococcus suis serotype 2 strains. J Biol Chem 293:12011–12025
Payen S, Roy D, Boa A, Okura M, Auger JP, Segura M, Gottschalk M (2021) Role of maturation of lipoproteins in the pathogenesis of the infection caused by Streptococcus suis serotype 2. Microorganisms 9:2386
Acknowledgements
The authors would like to thank S. Lacouture for technical help and Dr N. Fittipaldi for constructive suggestions with mutant constructions.
Funding
This study was funded by the Natural Sciences and Engineering Research Council of Canada (NSERC) to MG (grant #-2022–03730) and to MS (Grant # 2021–03020).
Author information
Authors and Affiliations
Contributions
Conception of the work: SP, JAR, MS, MG; laboratory techniques: SP, JAR; acquisition, analysis and interpretation of data: SP, MS, MG; preparation of the manuscript: SP, JAR, MS, MG. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All experiments involving animals were conducted in accordance with the guidelines and policies of the Canadian Council on Animal Care and the principles set in the Guide for the Care and the Use of Laboratory Animals by the Animal Welfare Committee of the University of Montreal, which approved the protocols and procedures used herein (permit 550 number RECH-1570). A total of 46 mice were used in this study.
Competing interests
The authors declare that they have no competing interests.
Data availability
The materials and data not presented in this manuscript are available from the corresponding author upon request.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Absence of Lmb does not influence
S. suis surface hydrophobicity. Surface hydrophobicity of of the wild-type P1/7 strain (dark blue), Δlmb (light blue), ΔcpsF (dark brown) and Δlmb/ΔcpsG (light brown) strains was determined using n-hexadecane. Data represent the mean ± SEM from at least three independent experiments. *Indicates a significant difference (p < 0.05).
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Payen, S., Rrodriguez, J.A., Segura, M. et al. Laminin-binding protein of Streptococcus suis serotype 2 influences zinc acquisition and cytokine responses. Vet Res 54, 1 (2023). https://doi.org/10.1186/s13567-022-01128-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-022-01128-8