
Abstract
Chicken cathelicidin-2 (CATH-2) as a host defense peptide has been identified to have potent antimicrobial and immunomodulatory activities. Here, we reported the mechanism by which CATH-2 modulates NLRP3 inflammasome activation. Our results show that CATH-2 and ATP as a positive control induced secretion of IL-1β and IL-1α in LPS-primed macrophages but did not affect secretion of IL-6, IL-12 and TNF-α. Furthermore, CATH-2 induced caspase-1 activation and oligomerization of apoptosis-associated speck-like protein containing a carboxy- terminal caspase recruitment domain (ASC), which is essential for NLRP3 inflammasome activation. However, CATH-2 failed to induce IL-1β secretion in Nlrp3−/−, Asc−/− and Casp1−/− macrophages. Notably, IL-1β and NLRP3 mRNA expression were not affected by CATH-2. In addition, CATH-2-induced NLRP3 inflammasome activation was mediated by K+ efflux but independent of the P2X7 receptor that is required for ATP-mediated K+ efflux. Gene interference of NEK7 kinase which has been identified to directly interact with NLRP3, significantly reduced IL-1β secretion and caspase-1 activation induced by CATH-2. Furthermore, confocal microscopy shows that CATH-2 significantly induced lysosomal leakage with the diffusion of dextran fluorescent signal. Cathepsin B inhibitors completely abrogated IL-1β secretion and caspase-1 activation as well as attenuating the formation of ASC specks induced by CATH-2. These results all indicate that CATH-2-induced activation of NLRP3 inflammasome is mediated by K+ efflux, and involves the NEK7 protein and cathepsin B. In conclusion, our study shows that CATH-2 acts as a second signal to activate NLRP3 inflammasome. Our study provides new insight into CATH-2 modulating immune response.
Similar content being viewed by others


Introduction
Antimicrobial resistance is one of the major challenges for human and animal health in the twenty-first century and new alternative strategies are needed to combat this global problem [1, 2]. So far, discovering new antibiotics based on conventional targets of microorganisms has failed to produce new classes of antibiotics [3, 4]. Therefore, there is an urgent need to find an innovative method to develop new drugs against microbial infections [5, 6]. Boosting host defense to kill pathogens rather than kill pathogens itself could be such a new potential therapy.
Host defense peptides (HDP) also known as antimicrobial peptides, are identified to control infections by directly killing pathogens or by modulating the host immune response [7]. Cathelicidins as one of the main families of HDP, are broadly expressed in different animals and could be developed as potential immunomodulatory molecules to treat microbial infection [8]. Immunomodulatory functions of chicken cathelicidin-2 (CATH-2) has been extensively studied in different species, such as in human, murine and porcine cells [9,10,11,12,13,14,15]. It efficiently neutralizes LPS and inhibits Toll like receptor-4 (TLR-4) activation in a non-species specific manner [14, 15]. Furthermore, the treatment of D-analog of CATH-2 in ovo reduced mortality and morbidity in chickens from E. coli infection up to 7 days after hatching [16]. These studies indicate that CATH-2 protects the host beyond antimicrobials itself but also modulates immune response and exerts its function cross-species.
Inflammasomes are an intracellular complex of proteins composed of a central protein, an adaptor (ASC) and an effector (caspase-1), which play an important role in the host defense against microbial infection. NOD-, LRR- and pyrin domain-containing protein 3 (NLRP3) are intracellular sensors detecting a broad range of microbial motifs, resulting in assembly and activation of an inflammasome, which finally leads to caspase-1‑mediated IL-1β maturation and secretion [17]. The NLRP3 inflammasome has been demonstrated to be important in response to infection by different pathogens, such as influenza virus, Streptococcal pneumoniae, Candida albicans, Pasteurella multocida and Staphylococcus aureus [18,19,20,21]. NLRP3 full activation requires two signals for priming and inflammasome formation. In addition, NLRP3 activation is thought to be involved in multiple upstream signals including efflux of potassium ions (K+), NIMA-related kinase 7 (NEK7) and lysosomal disruption [17]. It has been reported that human cathelicidin LL-37 can promote NLRP3 activation via the P2X7 receptor [22], but that is the only study that shows involvement of a cathelicidin in inflammasome formation; therefore data on whether other cathelicidins, including chicken CATH-2 can also regulate NLRP3 activation and especially whether the same mechanism is used, is still absent. A previous study has shown that mouse macrophages can be used as a good model to investigate the mechanism by which CATH-2 modulates innate immune response due to its non-species specificity [14, 15].
In this study, we investigated the mechanism through which CATH-2 modulates NLRP3 activation in LPS-primed mice primary peritoneal macrophages. Our results show that CATH-2 promoted IL-1β maturation and secretion via the NLRP3 pathway. Furthermore, CATH-2 induced ASC oligomerization, the formation of ASC specks and caspase-1 activation, leading to NLRP3 inflammasome activation. This process depended on K+ efflux. In addition, NEK7 and cathepsin B were also required for CATH-2-induced NLRP3 inflammasome activation. Our study reveals a novel modulatory role of CATH-2 and provides the basis for the development of novel therapeutic immunomodulatory strategies from the host perspective against antibiotic-resistant pathogens for cross species application.
Materials and methods
Animals
The wild-type (WT) C57BL/6 mice were purchased from Chongqing Academy of Chinese Material Medica (Chongqing, China). Nlrp3−/−, Asc−/− and Casp1−/− mice were a kind gift from Dr. Feng Shao, the NIBS (National Institute of Biological Sciences, Beijing, China). All gene knockout mice were on a C57BL/6 background and maintained in Specific Pathogen Free (SPF) conditions before being used at 8–10 weeks of age. All the animal experiments were approved by the Southwest University Ethics Committee, Chongqing, China (IACUC-2019-0627-05).
Peptides
All the peptides were synthesized by China Peptides (Shanghai, China) using Fmoc-chemistry. All peptides were purified by reverse phase high-performance liquid chromatography to a purity > 95%.
Preparation of macrophages and LPS stimulation in vitro
Mice were injected intraperitoneally with 2–3 mL of 4% thioglycolate medium (Eiken, Tokyo, Japan). After 3–4 days, the mice peritoneal exudate cells were collected by peritoneal lavage and suspended with RPMI 1640 supplemented with 10% FCS or Opti-MEM (Gibco, USA) as reported previously [21]. Then, the cells were seeded at 2 × 105 cells/well for 48-well plates or 1 × 106 cells/well for 12-well plates. These cells were maintained at a humidified 37 °C incubator with 5% CO2. After 2 h incubation, the nonadherent cells were removed and the adherent cells were used for assays described below. Subsequently, cells were stimulated with E. coli LPS (50 ng/mL) (Beyotime, China) for 3 h. After stimulation, peptides (5 μM) were added for an additional 21 h and ATP (1.5 mg/mL) (Beyotime, China) was added as the positive control. After 24 h, supernatants and cell lysates were collected for the assays described below. To inhibit K+ efflux and P2X7 receptor, KCl (5 mM and 50 mM) and P2X7 inhibitor (A-740003, 100 μM) (MedChemExpress, USA) were added after LPS stimulation. To inhibit CATH-2 endocytosis pathway, bafilomycin A1 (100 nM) and Cytochalasin B (10 μM) were added to incubate 1 h prior to CATH-2 treatment. Furthermore, to inhibit cathepsin activities, cathepsin B inhibitor (CA-074-Me, 20 μM) and cathepsin D inhibitor (pepstatin A, 20 μM) were added for 1 h incubation before LPS stimulation and CATH-2 treatment.
Cell viability
Cells were prepared and treated as described above. After incubation, 10% WST-reagent was added according to the manufacturer’s protocols. Finally, absorbance was measured at 450 nm with a microplate reader (Bio-Rad, Japan) and was corrected for absorbance at 630 nm.
Enzyme linked immunosorbent assay (ELISA)
Cells were prepared in 48-well plates and treated as described above. After incubation as described above, supernatants were collected and cytokine secretion was determined by ELISA according to the manufacturer’s instructions. The kits used in this study included IL-1β, TNF-α, IL-1α, IL-6 and IL-12 (Invitrogen, CA, USA).
Western blot analysis
Cells were prepared in 12-well plates and treated as described above. After incubation as indicated, supernatants were collected and concentrated using 20% (w/v) trichloroacetate, and the cells were lysed with 1 × SDS loading buffer (Beyotime, China) and then cell lysates were collected. Subsequently, concentrated supernatants and cell lysates were subjected to 12% SDS-PAGE and then transferred onto a polyvinylidene difluoride (PVDF) membrane by electroblotting. Next, the membranes were blocked with 5% nonfat dry milk and then immunoblotted with the indicated antibodies (Abs) including anti-IL-1β (R&D, USA), anti-caspase-1 p20 (AdipoGen, USA), anti-pro-IL-1β, anti-pro-caspase-1, anti-NEK7 (Abcam, Cambridge, UK) and anti-β-actin (Beyotime, Beijing, China). Finally, the distinct protein bands were detected by ECL detection reagent (Biosharp, China).
ASC oligomerization
Cells were prepared in 12-well plates and treated as described above. After incubation, cells were lysed with cold PBS containing 0.5% Triton X-100 and then cell lysates were centrifuged at 13 000 rpm for 15 min at 4 ℃ to obtain the cell pellets. The pellets were washed twice with cold PBS and suspended in 200 μL PBS. The resuspended pellets were cross-linked with 4 mM fresh disuccinimidyl suberate (DSS) at 37 ℃ for 30 min and then the pellets were centrifuged at 13 000 rpm for 15 min at 4 ℃. Finally, the cross-linked pellets were dissolved in 30 μL 1 × SDS-PAGE sample loading buffer and samples were boiled for 5 min before the Western blot analysis.
Quantitative real time polymerase chain reaction (RT-PCR)
Cells were prepared in 12-well plates and treated as described above. After 6 h incubation, cells were lysed and total RNA were extracted by TRIzol Reagent (Life Technologies Carlsbad, CA, USA) according to the manufacturers’ instructions. Then, cDNA was synthesized using PrimeScript® RT reagent Kit (Perfect Real Time) (Takara, Japan). Finally, RT-PCR was performed using the CFX96 (Bio-Rad, USA). Primers were used as follows: IL-1β forward 5′-GAA ATG CCA CCT TTT GAC AGT G and reverse 5′-TGG ATG CTC TCA TCA GGA CAG, NLRP3 forward 5′-CTT TCT GGA CTC TGA CCG GG and reverse 5′-CTC CCA TTC TGG CTC TTC CC, β-actin forward 5′-TGG AAT CCT GTG GCA TCC ATG AAA C and reverse 5′-TAA AAC GCA GCT CAG TAA CAG TCC G. Relative gene expression levels were normalized against the expression levels of β-actin.
SiRNA interference
Cells were prepared as described above and transfected using Lipofectamine 3000 (Thermo Scientific, USA) with 60 nM of Nek7 siRNA (Sangon Biotech, 5′-GAUAGACUGUGUUUAUAGATT-3′) or 60 nM of control siRNA (Sangon Biotech, 5′-UUCUCCGAACGUGUCACGUTT-3′) for 48 h. Subsequently, cells were primed with LPS and treated with CATH-2 as described above. After incubation, cell supernatants and lysates were collected for ELISA and Western blot analysis.
Immunofluorescence staining
Cells were prepared in 48-well plates and treated with LPS and CATH-2 as described above. After 6 h incubation, cells were washed three times with PBS and fixed in 4% paraformaldehyde (Sango Biotech, Shanghai, China) for 20 min at room temperature (RT). After three wash steps, cells were permeabilized with 0.1% Triton X-100 in PBS for 10 min. Subsequently, cells were blocked with 5% Bovine Serum Albumin (BSA) in PBS for 30 min. Then, cells were stained with primary antibody containing anti-NLRP3 (Bioss, Beijing, China) and anti-ASC (Santa cruz, CA, USA) for 1 h at RT. After the wash steps, cells were incubated with Goat anti-mouse IgG (H&L) Alexa fluor 488 and Goat anti-rabbit IgG (H&L) Alexa fluor 594 (Abcam, UK) for 1 h. DAPI (Beyotime Biotechnology, Shanghai, China) was added for 5 min to visualize cell nuclei. Finally, cells were washed and maintained in antifading medium (Solarbio, Beijing, China). Cells were observed using the fluorescence microscopy (Olympus, Tokyo, Japan).
Lysosomal leakage
Cells were prepared in 48-well plates as described above. Then, cells were loaded with Alexa 488 Dextran (20 μM), 10 kDa (Life Technologies) for 3 h prior to LPS and CATH-2 treatment for further 6 h. After incubation, the cells were observed using fluorescence microscopy (Olympus, Tokyo, Japan).
Statistical analysis
Data are represented as mean ± SEM of three independent experiments for each group (n = 3). One-way ANOVA were used to analyze statistical significance among different groups. Statistical significance was shown as *p < 0.05, **p < 0.01, ***P < 0.001, ns = no significance.
Results
CATH-2 induces IL-1β and IL-1α secretion in LPS-primed macrophages
To investigate immunomodulatory properties of CATH-2 after LPS stimulation, different cytokines were measured in mice primary peritoneal macrophages. As shown in Figure 1, LPS induced high levels of IL-6, IL-12 and TNF-α secretion in macrophages but it failed to induce IL-1β and IL-1α secretion (Figures 1A, B, C). Under the treatment of CATH-2 and ATP, IL-1β and IL-1α were produced in LPS-primed macrophages while IL-6, IL-12 and TNF-α production was not affected by CATH-2. However, ATP significantly decreased LPS-induced production of IL-6 and IL-12 (Figures 1C, D). Moreover, CATH-2 had no effect on cell viability but ATP significantly reduced cell viability (Figure 1F), suggesting that ATP induces cell death. These results indicate that CATH-2 specifically acts on the IL-1β signaling pathway.

CATH-2 induces IL-1β and IL-1α secretion in LPS-primed macrophages. Cells were primed with LPS for 3 h and then CATH-2 (5 μM) and ATP (1.5 mg/mL) as positive control were added for an additional 21 h. After 24 h, supernatants were collected to determine cytokine secretion of IL-1β (A), IL-1α (B), IL-6 (C), IL-12 (D) and TNF-α (E) using ELISA; WST-1 was used to detect metabolic activity of cells (F). Data are represented as mean ± SEM of three independent experiments of triplicate samples per experiment. *P ≤ 0.05; ** P ≤ 0.01; *** P ≤ 0.005.
CATH-2 induces NLRP3 activation via ASC oligomerization and caspase-1 activation in LPS-primed macrophages
NLRP3 activation leads to IL-1β secretion via the assembly of adaptor protein ASC to recruit caspase-1 which cleaves pro-IL-1β. To further investigate whether CATH-2 induced IL-1β secretion via the NLRP3 pathway, macrophages from WT, Nlrp3−/−, Asc−/−, and Casp1−/− mice were used in this study. The results show that CATH-2 and ATP-induced IL-1β secretion was completely abrogated in LPS-primed Nlrp3−/−, Asc−/−, and Casp1−/− macrophages while TNF-α secretion was not affected (Figures 2A, B), indicating CATH-2-induced IL-1β secretion via an NLRP3-dependent mechanism. Subsequently, caspase-1 and ASC oligomerization were detected by Western blot. The results show that CATH-2 induced caspase-1 activation and ASC oligomerization in LPS-primed macrophages (Figures 2C, D). However, CATH-2 did not promote LPS-induced mRNA expression of IL-1β and NLRP3 (Figures 2E, F). These results demonstrate that CATH-2 acts as a second signal to activate NLRP3, resulting in IL-1β maturation and secretion.

CATH-2 induces NLRP3 activation via ASC oligomerization and caspase-1 activation in LPS-primed macrophages. Cells from WT, Nlrp3−/−, Asc−/−, Casp1−/− mice were primed with LPS for 3 h and then CATH-2 (5 μM) and ATP (1.5 mg/mL) as positive control were added for an additional 3 h or 21 h. After 24 h, cell supernatants and lysates were collected for different assays. ELISA was used to determine cytokine secretion of IL-1β (A) and TNF-α (B); A Representative image of the Western blot assay is presented to show protein expression including IL-1β, caspase-1, pro-IL-1β, pro-caspase-1 and β-action (C) as well as ASC protein expression (D); qPCR was used to measure mRNA expression of IL-1β (E) and NLRP3 (F). Data in A, B, D, F are represented as mean ± SEM of three independent experiments from triplicate samples per experiment. * P ≤ 0.05; ** P ≤ 0.01; *** P ≤ 0.005.
CATH-2 induces NLRP3 activation via a P2X7R-independent manner but dependent on K+ efflux in LPS-primed macrophages
It has been reported that ATP-mediated activation of the P2X7 receptor (P2X7R), a ligand-gated ion channel of the purinergic receptor family, promotes NLRP3 activation dependent on K+ efflux [23]. According to the findings that the role of CATH-2 as a 2nd signal to activate NLRP3 is similar to the role of ATP, we identified the effect of P2X7R and K+ efflux on CATH-2-induced NLRP3 activation in LPS-primed macrophages.
To block these two signaling pathways, P2X7R inhibitor A-740003 and KCl were used in this study. The results show that the P2X7R inhibitor had no effect on CATH-2 induced IL-1β secretion in LPS-primed macrophages while the inhibitor completely abolished ATP-induced IL-1β secretion (Figure 3A). On the contrary, TNF-α secretion that is not related to inflammasome formation, was not affected by the P2X7R inhibitor (Figure 3B). Notably, the results of the Western blot analysis show that CATH-2-induced IL-1β and caspase-1 protein expression were not affected by the P2X7R inhibitor (Figure 3C), suggesting CATH-2-induced NLRP3 activation independent of the P2X7 receptor. In addition, after K+ efflux was blocked by KCl in LPS-primed macrophages, both CATH-2 and ATP-induced IL-1β secretion were significantly inhibited but TNF-α secretion was not affected (Figures 3D, E). Furthermore, CATH-2-induced caspase-1 activation was also attenuated by KCl (Figure 3F). These results indicate that CATH-2 promotes K+ efflux independently of P2X7R, which leads to NLRP3 activation.

CATH-2 induces NLRP3 activation via a P2X7R-independent manner but dependent on K+ efflux in LPS-primed macrophages. Cells were primed with LPS for 3 h and then CATH-2 (5 μM) and ATP (1.5 mg/mL) as positive control were added for an additional 21 h. To inhibit K + efflux and the P2X7 receptor, cells were untreated (UT) or treated with KCl (5 mM and 50 mM) and the P2X7R inhibitor (A-740003, 100 μM) after LPS stimulation. Finally, cell supernatants and lysates were collected for different assays. ELISA was used to determine cytokine secretion of IL-1β (A, D) and TNF-α (B, E). A representative image (Western blot) is presented to show protein expression including IL-1β, caspase-1, pro-IL-1β, pro-caspase-1 and β-action (C, F). Data in A, B, D, E are represented as mean ± SEM from three independent experiments with triplicate samples per experiment. * P ≤ 0.05; ** P ≤ 0.01; *** P ≤ 0.005.
NEK7 is involved in CATH-2-induced NLRP3 activation in LPS-primed macrophages
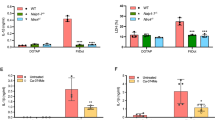
NEK7 is a serine threonine kinase which is found to be critical for NLRP3 inflammasome activation and which specifically interacts with NLRP3 but not with the other inflammasomes [24]. To identify whether CATH-2-induced NLRP3 activation requires NEK7, its gene expression was silenced using siRNA knock-out. The results show that CATH-2 induced-IL-1β secretion was significantly decreased after NEK7 siRNA transfection in LPS-primed macrophages, which was consistent with the results in Western blot analysis (Figures 4A, C). IL-6 secretion was not affected by silencing of NEK7 confirming the specific role of NEK7 in NLRP7 inflammasome formation (Figure 4B). Moreover, CATH-2-induced caspase-1 protein expression was also significantly reduced in NEK7 knock out macrophages (Figure 4C). These results demonstrate that CATH-2 promotes NEK7-NLRP3 interaction, leading to NLRP3 inflammasome activation.

NEK7 is involved in CATH-2-induced NLRP3 activation in LPS-primed macrophages. Cells were transfected with NEK7 siRNA and control siRNA for 24 h. Subsequently, cells were primed with LPS for 3 h and then CATH-2 (5 μM) were added for an additional 21 h. Cell supernatants and lysates were collected for different assays. ELISA was used to determine cytokine secretion of IL-1β (A) and IL-6 (B). A representative image (Western blot) is presented to show protein expression including IL-1β, caspase-1, pro-IL-1β, pro-caspase-1 and β-action (C). Data in A, B are represented as mean ± SEM from three independent experiments with triplicate samples per experiment. * P ≤ 0.05; ** P ≤ 0.01; *** P ≤ 0.005.
Lysosomal leakage of cathepsin B mediates CATH-2-induced NLRP3 activation in LPS-primed macrophages
To further investigate the mechanism by which CATH-2 exerts its function, bafilomycin A1 (Baf.A1) and cytochalasin B (Cyto.B) were used to inhibit the CATH-2 endocytosis pathway. The results show that Baf.A1 significantly decreased CATH-2-induced IL-1β secretion while Cyto.B did not affect IL-1β secretion (Figure 5A). However, IL-6 secretion was not altered by Baf.A1 and Cyto.B (Figure 5B). These results reveal that CATH-2 endosomal acidification is required for NLRP3 activation.

Lysosomal leakage of cathepsin B mediates CATH-2-induced NLRP3 activation in LPS-primed macrophages. Cells were primed with LPS for 3 h and then Baf.A1 (100 nM) and Cyto.B (10 μM) were added for 1 h incubation prior to CATH-2 treatment for a further 21 h. ELISA was used to determine cytokine secretion of IL-1β (A) and IL-6 (B). Confocal microscopy images of cells loaded with Alexa 488 Dextran (green) treatment with LPS and CATH-2, with dashed white arrows indicating fluorescent signal in lysosomes and solid white arrows indicating lysosomal leakage (C). Furthermore, cells were pretreated with cathepsin B inhibitor (CA-074-Me, 20 μM) and cathepsin D inhibitor (pepstatin A, 20 μM) for 1 h. Subsequently, cells were primed with LPS for 3 h and then CATH-2 (5 μM) was added for an additional 21 h. ELISA was used to determine cytokine secretion of IL-1β (D). A representative image of the Western blot assay is present to show protein expression including IL-1β, caspase-1, pro-IL-1β, pro-caspase-1 and β-action (E); Additionally, after 6 h incubation, immunofluorescence staining was performed using primary antibodies anti-NLRP3 and anti-ASC as well as secondary antibodies Goat anti-mouse IgG (H&L) Alexa fluor 488 and Goat anti-rabbit IgG (H&L) Alexa fluor 594. Representative images of ASC, ASC specks (white arrows), NLRP3 and cell nucleus (F). Data from figure A are represented as mean ± SEM of three independent experiments of triplicate samples per experiment. * P ≤ 0.05; ** P ≤ 0.01; *** P ≤ 0.005.
It has been reported that lysosomal disruption is crucial for NLRP3 activation. To investigate whether CATH-2 induces lysosomal leakage, fluorescently labelled Dextran was loaded in the macrophages, which resulted in punctate accumulation of fluorescent signal in lysosomes (Figure 5C, dashed white arrows). Subsequent CATH-2 treatment significantly reduced punctate accumulation of the fluorescent signal in lysosomes, evidenced by the diffusion of the fluorescent signal (Figure 5C, solid white arrows), indicating the lysosomal leakage.
To further investigate the significance of leakage of lysosomal cathepsin B and D, cathepsin B inhibitor (CA-074-Me) and cathepsin D inhibitor (pepstatin A) were used to pretreat cells before LPS stimulation. The results show that the cathepsin B inhibitor abrogated CATH-2-induced IL-1β secretion while cathepsin D inhibitor did not affect CATH-2-induced IL-1β secretion (Figure 5D). Similarly, the cathepsin B inhibitor completely inhibited CATH-2-induced caspase-1 protein expression (Figure 5E). Early studies have revealed that polymerization of the adaptor ASC and the assembly of ASC specks recruit and activate caspase-1 [25]. Our immunofluorescent staining shows that CATH-2 induced formation of ASC specks (Figure 5F, white arrows) in LPS-primed macrophages. However, CATH-2-induced formation of ASC specks was also decreased by the cathepsin B inhibitor (Figure 5C, white arrows). These results indicate that CATH-2 promotes lysosomal leakage to release cathepsin B and then induces assembly of ASC specks, which results in NLRP3 inflammasome activation.
Discussion
Cathelicidins exhibit potent antimicrobial activity against a broad spectrum of pathogens, such as bacteria, parasites, viruses and fungi [7, 26]. An additional function of cathelicidins is to modulate the host immune response, which contributes to host immune homeostasis [27]. The immunomodulatory activity of human cathelicidin LL-37 has been extensively studied. For example, LL-37 promotes cell differentiation, enhances killing and clearance of bacteria and regulates the production of inflammatory mediators in different cell types [27,28,29,30,31]. These responses are modulated by cell surface receptors including FPR2/ALX/P2X7R [27, 29]. However, the biological role of other cathelicidins such as the ones present in livestock, avian and fish is rarely studied. An exception is probably the chicken cathelicidin-2 for which the biological activities have also been extensively studied. Except for strong antimicrobial activity, CATH-2 to some extent shows partially similar immunomodulatory activities to LL-37 in human and mouse cells. Importantly, this function of CATH-2 has not been found to be species specific. CATH-2 has for example been found to play a role in LPS neutralization and modulation of TLR, but whether CATH-2 is involved in the modulation of intracellular signaling pathways remains unknown. Therefore, in this study, we explored the mechanism by which CATH-2 regulates NLRP3 inflammasome activation in mice peritoneal macrophages primed with LPS.
The NLRP3 inflammasome is the protein complex that recognizes different pathogens including bacteria and viruses, which plays an important role in the induction of an inflammatory response [17]. It has been reported that LL-37 internalizes into macrophages to induce the NLRP3 inflammasome [22]. Similarly, our study shows that CATH-2 activates NLRP3 inflammasome through the induction of ASC oligomerization and caspase-1 activation in LPS-primed macrophages. Furthermore, CATH-2 does not affect NLRP3 and IL-1β transcription. These results indicate that CATH-2 has similar immunomodulatory properties as LL-37 and both of them can act as a second signal to activate the NLRP3 inflammasome. Importantly, it has been reported that LL-37 activates NLRP3 inflammasome to generate protective epithelial cell inflammatory responses against Pseudomonas aeruginosa infection [32]. Due to the similar property with LL-37, it is reasonably speculated that CATH-2 could be used to enhance protective inflammatory responses for the clearance of immune tolerant pathogens.
The P2X7 receptor is highly expressed in macrophages and mediates the influx of Ca2+, Na+ and K+ ions, which controls the release of proinflammatory cytokines [17]. Our data show that ATP induces NLRP3 activation via the P2X7 receptor, which is consistent with the knowledge that ATP induces K+ efflux to activate the NLRP3 inflammasome [33]. Unlike LL-37’s internalization via the P2X7 receptor to activate the NLRP3 inflammasome in macrophages [23], our study shows that CATH-2 induces NLRP3 activation independently of the P2X7 receptor. However, some researchers have identified that the endocytosis of LL-37 via the P2X7 receptor depends on the cell type studied. In PBMC and epithelial cells, the P2X7 receptor was not actually involved in LL-37 internalization [32, 34]. In addition, Coorens et al. found that CATH-2/DNA complexes can be taken up by macrophages in a clathrin-dependent manner, providing clear evidence that CATH-2, at least in complex with DNA is endocytosed [13]. Combined with our results, it can be speculated that CATH-2 is internalized by macrophages and can then activate the NLRP3 pathway. However, the exact mechanism of CATH-2 endocytosis by macrophages needs to be further studied.
Efflux of K+ is thought to be a common trigger for NLRP3 inflammasome activation [33]. However, several reports have shown that NLRP3 inflammasome activation is independent of K+ efflux, possibly depending on the exact species that is studied as well [35]. In human monocytes, LPS-triggered NLRP3 inflammasome activation relies on NLRP3-ASC-caspase-1 signaling but is independent of K+ efflux [36]. In addition, imiquimod and CL097-mediated NLRP3 inflammasome activation directly targets mitochondria instead of induction of K+ efflux [36]. In contrast with these findings, our study demonstrates that the addition of extracellular high concentrations of KCl blocks efflux of K+ and thereby significantly inhibits CATH-2-induced NLRP3 inflammasome activation, indicating that CATH-2 might induce efflux of K+. Recent research has shown that K+ efflux is mediated by the K+ channel 2 (TWIK2) and the chloride intracellular channel (CLIC) protein acts downstream of K+ efflux to promote Cl− efflux, leading to NLRP3 inflammasome activation [37, 38]. However, whether CATH-2 interacts with these proteins that control ion channels needs to be further studied.
Recently, NEK7 was proposed as an essential component of the NLRP3 inflammasome where it regulates assembly and activation of the inflammasome [39,40,41]. It has been identified that NEK7 protein binding to NLRP3 mediates the activation of the NLRP3 inflammasome. Some reports have shown that NEK7-NLRP3 interaction, as revealed by Co-IP and GST pull-down assays, can be induced by different stimuli, such as LPS + ATP, bacterial and pathogen toxins [24, 42, 43]. Although our data does not directly show NEK7-NLRP3 interaction, NEK7 knockdown significantly reduced CATH-2-induced caspase-1 activation and IL-1β secretion, indicating that CATH-2 might promote NEK7-NLRP3 interaction. However, it has been recently reported that K+ efflux regulates the NEK7-NLRP3 interaction [24]. Therefore, CATH-2-induced NLRP3 inflammasome activation might also be mediated by upstream signaling events of NLRP3 inflammasome.
Lysosomal disruption, as the upstream signal of NLRP3 inflammasome can also induce its activation [44,45,46]. Phagocytosis of particulates causes lysosomal acidification and rupture. In addition, studies that used cathepsin inhibitors indicate that cathepsins resident in the lysosomes are important to activate the NLRP3 inflammasome in response to external stimuli. Considering the endosomal uptake of CATH-2 it is possible that (at least part) of the role of CATH-2 in inflammasome activation lies in its possible role in disruption of lysosomes, thereby releasing cathepsins [13]. This is in line with the observation from our study that CATH-2-induced IL-1β secretion is required for CATH-2 endosome acidification and CATH-2 induced lysosomal leakage. In addition, CATH-2-induced IL-1β secretion is completely blocked by cathepsin B inhibitors, although this inhibition might also result from suppressed pro-IL-1β synthesis. Actually, it has been reported that cathepsin B-deficient macrophages have similar IL-1β secretion as normal macrophages [45], indicating that IL-1β secretion is independent of cathepsin B. However, CATH-2-induced caspase-1 activation and formation of ASC specks inhibited by cathepsin B inhibitors suggest that CATH-2-induced NLRP3 inflammasome activation is mediated by lysosomal leakage of cathepsin B. This finding is similar to the effect of LL-37 [32]. It will be interesting to discover the basis of these seemingly contradicting results on the involvement of Cathepsin B, but it is likely that differences in species or cell types studied, or even experimental setup can have an effect on the observations. Our results at least, reveal that cathepsin B is not only involved in the priming step of pro-IL-1β synthesis, but also mediates NLRP3 inflammasome activation.
In conclusion, CATH-2 acts as a second signal to activate the NLRP3 inflammasome in LPS-primed macrophages, leading to ASC oligomerization and the formation of ASC specks as well as caspase-1 activation, finally resulting in IL-1β maturation and secretion. This activation process is mediated by K+ efflux. Furthermore, NEK7 and cathepsin B are also involved in CATH-2-induced NLRP3 inflammasome activation. Our study provides new insight on the immunomodulatory activity of CATH-2 and contributes to the development of CATH-2 as an immunomodulatory adjuvant against microbial infection not only in chickens, but also in other mammalian species.
Availability of data and materials
All data sets generated for this study are included in the article.
References
Christaki E, Marcou M, Tofarides A (2020) Antimicrobial resistance in bacteria: mechanisms, evolution, and persistence. J Mol Evol 88:26–40
Ferri M, Ranucci E, Romagnoli P, Giaccone V (2017) Antimicrobial resistance: a global emerging threat to public health systems. Crit Rev Food Sci Nutr 57:2857–2876
Silver LL (2011) Challenges of antibacterial discovery. Clin Microbiol Rev 24:71–109
Payne DJ, Gwynn MN, Holmes DJ, Pompliano DL (2007) Drugs for bad bugs: confronting the challenges of antibacterial discovery. Nat Rev Drug Discov 6:29–40
Baym M, Stone LK, Kishony R (2016) Multidrug evolutionary strategies to reverse antibiotic resistance. Science 351:aad3292
Durand GA, Raoult D, Dubourg G (2019) Antibiotic discovery: history, methods and perspectives. Int J Antimicrob Agents 53:371–382
Mookherjee N, Anderson MA, Haagsman HP, Davidson DJ (2020) Antimicrobial host defence peptides: functions and clinical potential. Nat Rev Drug Discov 19:311–332
van Harten RM, van Woudenbergh E, van Dijk A, Haagsman HP (2018) Cathelicidins: immunomodulatory antimicrobials. Vaccines 6:63
Kraaij MD, van Dijk A, Scheenstra MR, van Harten RM, Haagsman HP, Veldhuizen EJA (2020) Chicken CATH-2 increases antigen presentation markers on chicken monocytes and macrophages. Protein Pept Lett 27:60–66
Kraaij MD, van Dijk A, Haagsman HP (2017) CATH-2 and LL-37 increase mannose receptor expression, antigen presentation and the endocytic capacity of chicken mononuclear phagocytes. Mol Immunol 90:118–125
Scheenstra MR, van den Belt M, Tjeerdsma-van Bokhoven JLM, Schneider VAF, Ordonez SR, van Dijk A, Veldhuizen EJA, Haagsman HP (2019) Cathelicidins PMAP-36, LL-37 and CATH-2 are similar peptides with different modes of action. Sci Rep 9:4780
van Dijk A, van Eldik M, Veldhuizen EJ, Tjeerdsma-van Bokhoven HL, de Zoete MR, Bikker FJ, Haagsman HP (2016) Immunomodulatory and anti-inflammatory activities of chicken cathelicidin-2 derived peptides. PLoS One 11:e0147919
Coorens M, van Dijk A, Bikker F, Veldhuizen EJ, Haagsman HP (2015) Importance of endosomal cathelicidin degradation to enhance DNA-induced chicken macrophage activation. J Immunol 195:3970–3977
van Harten RM, Veldhuizen EJA, Haagsman HP, Scheenstra MR (2022) The cathelicidin CATH-2 efficiently neutralizes LPS- and E coli-induced activation of porcine bone marrow derived macrophages. Vet Immunol Immunopathol 244:110369
Coorens M, Schneider VAF, de Groot AM, van Dijk A, Meijerink M, Wells JM, Scheenstra MR, Veldhuizen EJA, Haagsman HP (2017) Cathelicidins inhibit Escherichia coli-induced TLR2 and TLR4 activation in a viability-dependent manner. J Immunol 199:1418–1428
Cuperus T, van Dijk A, Matthijs MG, Veldhuizen EJ, Haagsman HP (2016) Protective effect of in ovo treatment with the chicken cathelicidin analog D-CATH-2 against avian pathogenic E. coli. Sci Rep 6:26622
Swanson KV, Deng M, Ting JP (2019) The NLRP3 inflammasome: molecular activation and regulation to therapeutics. Nat Rev Immunol 19:477–489
Sarvestani ST, McAuley JL (2017) The role of the NLRP3 inflammasome in regulation of antiviral responses to influenza A virus infection. Antiviral Res 148:32–42
Surabhi S, Cuypers F, Hammerschmidt S, Siemens N (2020) The role of NLRP3 inflammasome in pneumococcal infections. Front Immunol 11:614801
Hise AG, Tomalka J, Ganesan S, Patel K, Hall BA, Brown GD, Fitzgerald KA (2009) An essential role for the NLRP3 inflammasome in host defense against the human fungal pathogen Candida albicans. Cell Host Microbe 5:487–497
Fang R, Du H, Lei G, Liu Y, Feng S, Ye C, Li N, Peng Y (2019) NLRP3 inflammasome plays an important role in caspase-1 activation and IL-1β secretion in macrophages infected with Pasteurella multocida. Vet Microbiol 231:207–213
Kahlenberg JM, Carmona-Rivera C, Smith CK, Kaplan MJ (2013) Neutrophil extracellular trap-associated protein activation of the NLRP3 inflammasome is enhanced in lupus macrophages. J Immunol 190:1217–1226
Karmakar M, Katsnelson MA, Dubyak GR, Pearlman E (2016) Neutrophil P2X7 receptors mediate NLRP3 inflammasome-dependent IL-1β secretion in response to ATP. Nat Commun 7:10555
He Y, Zeng MY, Yang D, Motro B, Núñez G (2016) NEK7 is an essential mediator of NLRP3 activation downstream of potassium efflux. Nature 530:354–357
Lu A, Magupalli VG, Ruan J, Yin Q, Atianand MK, Vos MR, Schröder GF, Fitzgerald KA, Wu H, Egelman EH (2014) Unified polymerization mechanism for the assembly of ASC-dependent inflammasomes. Cell 156:1193–1206
Peng L, Du W, Balhuizen MD, Haagsman HP, de Haan CAM, Veldhuizen EJA (2020) Antiviral activity of chicken cathelicidin B1 against influenza A virus. Front Microbiol 11:426
Hancock RE, Haney EF, Gill EE (2016) The immunology of host defence peptides: beyond antimicrobial activity. Nat Rev Immunol 16:321–334
Li Y, Cai L, Wang H, Wu P, Gu W, Chen Y, Hao H, Tang K, Yi P, Liu M, Miao S, Ye D (2011) Pleiotropic regulation of macrophage polarization and tumorigenesis by formyl peptide receptor-2. Oncogene 30:3887–3899
Tang X, Basavarajappa D, Haeggström JZ, Wan M (2015) P2X7 receptor regulates internalization of antimicrobial peptide LL-37 by human macrophages that promotes intracellular pathogen clearance. J Immunol 195:1191–1201
Pahar B, Madonna S, Das A, Albanesi C, Girolomoni G (2020) Immunomodulatory role of the antimicrobial LL-37 peptide in autoimmune diseases and viral infections. Vaccines 8:517
Minns D, Smith KJ, Alessandrini V, Hardisty G, Melrose L, Jackson-Jones L, MacDonald AS, Davidson DJ, Gwyer Findlay E (2021) The neutrophil antimicrobial peptide cathelicidin promotes Th17 differentiation. Nat Commun 12:1285
McHugh BJ, Wang R, Li HN, Beaumont PE, Kells R, Stevens H, Young L, Rossi AG, Gray RD, Dorin JR, Gwyer Findlay EL, Brough D, Davidson DJ (2019) Cathelicidin is a “fire alarm”, generating protective NLRP3-dependent airway epithelial cell inflammatory responses during infection with Pseudomonas aeruginosa. PLoS Pathog 15:e1007694
Muñoz-Planillo R, Kuffa P, Martínez-Colón G, Smith BL, Rajendiran TM, Núñez G (2013) K+ efflux is the common trigger of NLRP3 inflammasome activation by bacterial toxins and particulate matter. Immunity 38:1142–1153
Mookherjee N, Lippert DN, Hamill P, Falsafi R, Nijnik A, Kindrachuk J, Pistolic J, Gardy J, Miri P, Naseer M, Foster LJ, Hancock REW (2009) Intracellular receptor for human host defense peptide LL-37 in monocytes. J Immunol 183:2688–2696
Gaidt MM, Ebert TS, Chauhan D, Schmidt T, Schmid-Burgk JL, Rapino F, Robertson AA, Cooper MA, Graf T, Hornung V (2016) Human monocytes engage an alternative inflammasome pathway. Immunity 44:833–846
Groß CJ, Mishra R, Schneider KS, Médard G, Wettmarshausen J, Dittlein DC, Shi H, Gorka O, Koenig PA, Fromm S, Magnani G, Ćiković T, Hartjes L, Smollich J, Robertson AAB, Cooper MA, Schmidt-Supprian M, Schuster M, Schroder K, Broz P, Traidl-Hoffmann C, Beutler B, Kuster B, Ruland J, Schneider S, Perocchi F, Groß O (2016) K(+) efflux-independent NLRP3 inflammasome activation by small molecules targeting mitochondria. Immunity 45:761–773
Tang T, Lang X, Xu C, Wang X, Gong T, Yang Y, Cui J, Bai L, Wang J, Jiang W, Zhou R (2017) CLICs-dependent chloride efflux is an essential and proximal upstream event for NLRP3 inflammasome activation. Nat Commun 8:202
Di A, Xiong S, Ye Z, Malireddi RKS, Kometani S, Zhong M, Mittal M, Hong Z, Kanneganti TD, Rehman J, Malik AB (2018) The TWIK2 potassium efflux channel in macrophages mediates NLRP3 inflammasome-induced inflammation. Immunity 49:56-65.e54
Sharif H, Wang L, Wang WL, Magupalli VG, Andreeva L, Qiao Q, Hauenstein AV, Wu Z, Núñez G, Mao Y, Wu H (2019) Structural mechanism for NEK7-licensed activation of NLRP3 inflammasome. Nature 570:338–343
Shi H, Wang Y, Li X, Zhan X, Tang M, Fina M, Su L, Pratt D, Bu CH, Hildebrand S, Lyon S, Scott L, Quan J, Sun Q, Russell J, Arnett S, Jurek P, Chen D, Kravchenko VV, Mathison JC, Moresco EM, Monson NL, Ulevitch RJ, Beutler B (2016) NLRP3 activation and mitosis are mutually exclusive events coordinated by NEK7, a new inflammasome component. Nat Immunol 17:250–258
Schmid-Burgk JL, Chauhan D, Schmidt T, Ebert TS, Reinhardt J, Endl E, Hornung V (2016) A genome-wide CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) screen identifies NEK7 as an essential component of NLRP3 inflammasome activation. J Biol Chem 291:103–109
Boal-Carvalho I, Mazel-Sanchez B, Silva F, Garnier L, Yildiz S, Bonifacio JP, Niu C, Williams N, Francois P, Schwerk N, Schöning J, Carlens J, Viemann D, Hugues S, Schmolke M (2020) Influenza A viruses limit NLRP3-NEK7-complex formation and pyroptosis in human macrophages. EMBO Rep 21:e50421
Liu R, Liu Y, Liu C, Gao A, Wang L, Tang H, Wu Q, Wang X, Tian D, Qi Z, Shen Y (2021) NEK7-Mediated Activation of NLRP3 Inflammasome is coordinated by potassium efflux/Syk/JNK signaling during Staphylococcus aureus infection. Front Immunol 12:747370
Hornung V, Bauernfeind F, Halle A, Samstad EO, Kono H, Rock KL, Fitzgerald KA, Latz E (2008) Silica crystals and aluminum salts activate the NALP3 inflammasome through phagosomal destabilization. Nat Immunol 9:847–856
Orlowski GM, Colbert JD, Sharma S, Bogyo M, Robertson SA, Rock KL (2015) Multiple cathepsins promote pro-IL-1β synthesis and NLRP3-Mediated IL-1β activation. J Immunol 195:1685–1697
Katsnelson MA, Lozada-Soto KM, Russo HM, Miller BA, Dubyak GR (2016) NLRP3 inflammasome signaling is activated by low-level lysosome disruption but inhibited by extensive lysosome disruption: roles for K+ efflux and Ca2+ influx. Am J Physiol Cell Physiol 311:C83–C100
Acknowledgements
We would like to thank Dr Feng Shao from NIBS (National Institute of Biological Sciences, Beijing) to provide the Casp1/11−/−, Asc−/− and Nlrp3−/− mice. This study was supported by National Natural Science Foundation of China (32102684, 32172850), the Fundamental Research Funds for the Central Universities (SWU120057), the Chongqing Pig Industry Technology System (20211105) and the Chongqing Science and Technology Commission (cstc2021jcyj-msxm2218, cstc2021jcyj-msxmX0504).
Author information
Authors and Affiliations
Contributions
LP and HT performed all experiments, wrote the manuscript and designed the experiments. YL, KJ, JR, QT, GL, CW and CY helped perform parts of the experiments. EV, HC and RF contributed to the supervision of the experiments and corrected the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Peng, L., Tian, H., Lu, Y. et al. Chicken cathelicidin-2 promotes NLRP3 inflammasome activation in macrophages. Vet Res 53, 69 (2022). https://doi.org/10.1186/s13567-022-01083-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13567-022-01083-4