
Abstract
Background
Mesenchymal stem cell (MSC)-derived exosomes have shown comprehensive application prospects over the years. Despite performing similar functions, exosomes from different origins present heterogeneous characteristics and components; however, the relative study remains scarce. Lacking of a valuable reference, researchers select source cells for exosome studies mainly based on accessibility and personal preference.
Methods
In this study, exosomes secreted by MSCs derived from different tissues were isolated, by ultracentrifugation, and proteomics analysis was performed. A total of 1014 proteins were detected using a label-free method.
Results
Bioinformatics analysis revealed their shared function in the extracellular matrix receptor. Bone marrow MSC-derived exosomes showed superior regeneration ability, and adipose tissue MSC-derived exosomes played a significant role in immune regulation, whereas umbilical cord MSC-derived exosomes were more prominent in tissue damage repair.
Conclusions
This study systematically and comprehensively analyzes the human MSC-derived exosomes via proteomics, which reveals their potential applications in different fields, so as to provide a reference for researchers to select optimal source cells in future exosome-related studies.
Similar content being viewed by others

Background
In recent years, mesenchymal stem cells (MSCs) have been widely used in clinical treatment owing to their multi-differentiation potential [1, 2]. These cells play a significant role in immune regulation [3] and angiogenesis [4], especially in the heart, bone, cartilage, nerves, and skin [5,6,7]. There are many tissue-derived MSCs that have been used in therapeutic research, such as the bone marrow (BM), adipose tissue (AT), fetal lung, dental pulp, and umbilical cord (UC) [8, 9]. Paracrine secretion, particularly exosomes, is one of the main mechanisms of therapeutic action of MSCs [10], which has received much attention for its important roles in cellular communication and regenerative medicine.
Exosomes are lipid bilayer vesicles of 30–200 nm in diameter originating from endocytosis [11], which are secreted by multiple cells and appear in the supernatant of cell cultures or body fluids, such as the blood, saliva, urine, breast milk, cerebrospinal fluid, bile, and lymph [11,12,13]. Many types of molecules, including proteins, lipids, and nucleic acids, are present in exosomes [11]. When exosomes are absorbed by other cells, these molecules are transferred to the recipient cells and affect their physiological function, thus acting as intercellular communication vehicles and mediating many physiological and pathological processes [2, 14]. Studies have described several limitations to the direct use of MSCs, such as low survival rate, immunological rejection, and safety, and ethical issues [15,16,17]. Nevertheless, exosomes represent a safe and effective stem cell-free alternative therapeutic modality [18,19,20].
Previous studies have shown that MSC-derived from different tissues, as well as their secretome, have significant differences [21, 22]. As important components of the secretome, exosomes from different sources deserve further investigation. Most studies on exosomes have focused on RNA molecules (miRNA, lncRNA, and circRNA) [23,24,25]. However, MSC-derived exosomes are likely to play functional roles because of their proteins rather than RNAs, owing to their biologically relevant concentration, biochemical functionality, and potential to elicit an appropriate timely biochemical response [26].
Presently, exosomes derived from BM-, AT-, and UC-MSCs have gained the most attention. Therefore, in the present study, these three types of exosomes were selected for further investigation (SF. 1), and their morphology and surface markers were analyzed. Using a label-free method and bioinformatics tools, the systematic proteomic characteristics of exosomes were explored (SF. 1). This study aimed to provide novel insights for future human MSC-derived exosome research and treatment selection.
Methods
Culture and identification of MSCs
MSCs derived from different tissues (BM, AT, and UC) were obtained from Cyagen Biosciences (Suzhou, China). The cells were seeded in a specific culture medium (HUXMA-90011, HUXMD-90011, and HUXUC-90011, respectively; Cyagen Biosciences) and maintained at 37 °C and 5% CO2. The culture medium was changed every day. MSCs of the third passage were used in the following experiments.
To evaluate their multiple differentiation potential, MSCs were induced to differentiate by culturing in osteogenic, adipogenic, and chondrogenic differentiation medium (Cyagen), which confirmed by Alizarin red, Oil red O, and Alcian blue staining. MSC phenotypic analysis was performed using flow cytometry (BD Biosciences, San Jose, CA, USA), and the percentage of cells expressing mesenchymal markers—CD105, CD29, CD73, CD34, CD45, CD11b, and CD44 (Abcam, Cambridge, UK), was determined.
Isolation and characterization of exosomes
Exosomes were isolated by ultracentrifugation as described previously [27]. Briefly, MSCs were swilled thrice with PBS and cultured in serum-free medium for 48 h. The supernatant was collected and used for gradient centrifugation at 300×g for 10 min, 2000×g for 10 min, and 10,000×g for 30 min to remove cells and debris. The supernatant was filtered through a 0.45-μm porous membrane and centrifuged at 120,000×g for 70 min, in an L-80XP ultracentrifuge equipped with a Ti70 rotor (Beckman Coulter, Brea, CA, USA). The pellet was resuspended in PBS and centrifuged again as described above. The exosomes were resuspended in PBS and stored at − 80 °C until further analysis.
Nanoparticle tracking analysis (NTA) through the NanoSight NS300 system (Malvern Instruments, Malvern, UK) was used to measure the concentration and size of exosomes. The morphological characteristics of exosomes were detected using transmission electron microscopy (TEM). The expression of surface markers including calnexin (ab13504), CD9 (ab254175), CD81 (ab219209), and Tsg101 (ab125011) (Abcam) on exosomes was analyzed by western blot.
Protein extraction and trypsin digestion
The samples stored at − 80 °C were thawed and treated with lysis buffer (8 M urea, 1% protease inhibitor). After ultrasonic cracking and centrifugation, protein concentration was measured using a BCA kit (Tiangen Biotech, Beijing, China). Proteins were reduced with 5 mM dithiothreitol for 30 min at 56 °C. Iodoacetamide (11 mM) was used as a decontaminant at 24 °C in darkness, and 50 mM ammonium bicarbonate was added to reduce the urea concentration of dithiothreitol to 1 mM. Finally, the proteins were digested overnight with trypsin at 50:1 trypsin-to-protein mass ratio. The digested peptide was desalted using a C18 column.
Liquid chromatography with tandem mass spectrometry (LC–MS/MS) analysis
The peptides were dissolved in mobile phase A solvent (0.1% (v/v) formic acid solution) and separated using a NanoElute ultra-high-performance liquid phase system from a C18 trap column to a C18 analytical column at a speed of 300 nL/min. Mobile phase B was an acetonitrile solution containing 0.1% formic acid. The peptides were subjected to NSI source followed by tandem mass spectrometry (MS/MS) in Q ExactiveTM Plus (Thermo Fisher Scientific, Waltham, MA, USA) coupled online to the UPLC. The electrospray voltage applied was 2.0 kV. The m/z scan range was 350 to 1800 for a full scan, and intact peptides were detected in the Orbitrap at a resolution of 70,000.
Bioinformatics analysis
Maxquant software package (v1.6.6.0) was used to retrieve the secondary MS data, and the Swiss-Prot_Human data was used as reference (20,600 proteins; Proteome ID: UP000005640). The sequences of the identified proteins were mapped according to their Gene Ontology (GO), to determine their biological and functional properties, using InterProScan (v.5.14-53.0, http://www.ebi.ac.uk/interpro/). Protein-enriched pathways assessment was performed by the Kyoto Encyclopedia of Genes and Genomes (KEGG) database and the KAAS tool (v.2.0, http://www.genome.jp/kaas-bin/kaas_main). The identified proteins were compared with exosome available data from ExoCarta database (http://www.exocarta.org).
Statistical analysis
All experiments were performed three times. Principal component analysis (PCA), relative standard deviation, and Pearson’s correlation coefficient were used to evaluate the quantitative repeatability of proteins. GraphPad Prism 7.0 (GraphPad Software, San Diego, CA, USA) was used for all statistical analyses. Results with a P value < 0.05 were considered statistically significant.
Results
Characteristics of MSCs and exosomes
The MSCs showed good morphology and excellent proliferation rates. Results of flow cytometry showed that the MSCs were positive for CD105, CD29, CD73, and CD44, and negative for IgG1, IgG2b, CD11b, CD34, and CD45 (SF. 2a-c). Cytochemical staining data further showed the differentiation potential of MSCs for osteogenesis, lipid formation, and cartilage formation, respectively (Fig. 1a). These results confirmed that the experimental cells had typical MSC characteristics.

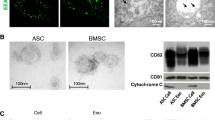
Quality control of MSCs and exosomes. a MSCs were stained with Alizarin red, Oil red O, and Alcian blue to confirm their differentiation into osteogenesis, adipogenesis, and chondrogenesis. (Scale bar 50 μm and 100 μm). b Size distribution of exosomes measured by NTA. c The MSCs and exosomes expression levels of Calnexin, CD9, CD81, and TSG101 were detected via western blot analysis. d TEM images of exosomes derived from MSCs. (Scale bar 100 nm, 500 nm)
The particle size distribution for exosomes was within 30–200 nm, with most exosomes (exo) having a 150-nm diameter. The concentration of AT-MSC exo was higher than that of BM-MSC exo or UC-MSC exo (Fig. 1b). Western blot analysis confirmed the presence of surface markers of the source cells and exosomes such as CD9, CD81, TSG101, and Calnexin, indicating that exosomes were representative (Fig. 1c). Typical cup-shaped vesicles were observed by TEM, and no differences in shape among the exosomes from the three different tissue sources were noted (Fig. 1d).
Bioinformatics analyses of exosomes from different sources
Most of the peptides comprised 7–20 amino acids, meeting the quality control requirements for LC–MS/MS analysis (SF. 3a). The molecular weight of the protein was negatively correlated with the coverage (SF. 3b). The Pearson’s diagram demonstrated a high correlation among exosomes of the same cell origin. Among exosomes from different sources, the correlation between BM-MSC exo and UC-MSC exo was low (SF. 3c). The PCA diagram indicated good repeatability (SF. 3d).
A detailed proteomic analysis of the differently sourced-exosomes revealed 771, 457, and 431 proteins in BM-MSC exo, AT-MSC exo, and UC-MSC exo, respectively. Almost 90% of proteins in the three types of exosomes matched ExoCarta, indicating that the results were significantly reliable, and their unique proteins enriched the existing exosome library (Figs. 2a, 3a, and 4a). The GO analysis grouped the results as per biological process, cellular component, and molecular function. In the cellular component, the identified proteins were significantly enriched in extracellular region and cytoplasm region (Fig. 2b, 3b, and 4b). In terms of biological process, BM-MSC exo proteins were mainly involved in granulocyte activation and regulation of cell migration, whereas AT-MSC exo and UC-MSC exo proteins were enriched in leukocyte activation involved in immune response. In addition, UC-MSC exo proteins were also enriched in collagen metabolic process. As for molecular function, AT-MSC exo and UC-MSC exo proteins were both significantly enriched in cell adhesion molecule binding, whereas BM-MSC exo proteins were mostly involved in protein complex binding and integrin binding. The KEGG pathway analysis showed that all three exosome proteins were enriched in extracellular matrix (ECM)-receptor interaction, with BM-MSC exo proteins being mainly enriched in proteasome (Figs. 2c, 3c and 4c).

Bioinformatics analysis of BM-MSC exo. a Venn diagram of BM-MSC-derived exosomes against ExoCarta. b GO analysis of BM-MSC exo. c KEGG analysis of BM-MSC exo

Bioinformatics analysis of AT-MSC exo. a Venn diagram of AT-MSC-derived exosomes against ExoCarta. b GO analysis of AT-MSC exo. c KEGG analysis of AT-MSC exo

Bioinformatics analysis of UC-MSC exo. a Venn diagram of UC-MSC-derived exosomes against ExoCarta. b GO analysis of UC-MSC exo. c KEGG analysis of UC-MSC exo
Horizontal bioinformatics analysis of the three sources of exosomes
Comparison of the three types of exosomes using a Venn diagram revealed 355 common proteins, and 341, 23, and 37proteins unique to BM-MSC exo, AT-MSC exo, and UC-MSC exo, respectively (Fig. 5a). Further GO analysis of the shared proteins showed that most of the proteins in the cellular component were concentrated in the extracellular matrix, anchoring junction, and adherent junction. Furthermore, proteins were enriched in secretion by cells at the biological process level and enriched in cadherin binding of molecular function (Fig. 5b). Moreover, in the KEGG pathway analysis, the shared proteins were significantly enriched in ECM-receptor interaction (Fig. 5c). According to the heat map, proteins in clusters 1 and 2, which comprised proteins involved in transport and pathways, were enriched in BM-MSC exo. Proteins in cluster 3, metabolic and immune proteins, were depleted in AT-MSC exo. Receptor and binding proteins (cluster 4) were enriched in UC-MSC exo, whereas BM-MSC exo was depleted of proteins involved in tissue development (clusters 5 and 6). Proteins in clusters 7 and 8, secretion and transport proteins, were depleted in UC-MSC exo. Proteins in cluster 9, proteins of immune and metabolic processes, were enriched in AT-MSC exo (Fig. 5d).

Horizontal bioinformatics analysis of three sources of exosome proteins. a Venn diagrams of detected proteins in exosomes derived from human MSCs. b GO analysis of the shared proteins among three sources of MSCs. (Red: biological process, green: cellular component, blue: molecular function.) c KEGG analysis of the shared proteins among three sources of MSCs. d Heat map of the protein level of shared proteins among three sources of MSCs. Clusters are assembled by GO analysis. The significantly enriched items are shown on the right
The volcanogram reflected the differential protein expression between every two groups (Fig. 6a). These differential proteins were mainly located in the extracellular region, followed by the cytoplasm (Fig. 6b). According to GO analysis, BM-MSC exo showed notable differences in terms of cellular components, when compared with AT-MSC exo and UC-MSC exo. Among them, the specific proteins of AT-MSC exo and UC-MSC exo were mostly enriched in the extracellular space and extracellular region, and BM-MSC exo was mostly enriched in the extracellular exosome and extracellular vesicle (Fig. 6c). In KEGG analysis, BM-MSC exo proteins were enriched in the proteasome, UC-MSC exo proteins were enriched in legionellosis, and AT-MSC exo proteins were enriched in cell cycle and oocyte meiosis (Fig. 6d).

Bioinformatics analysis of the specific proteins detected in each source of exosomes. a The volcanogram of differential proteins for BM-MSC exo vs AT-MSC exo, BM-MSC exo vs UC-MSC exo, and AT-MSC exo vs UC-MSC exo. b The cytolocalization of differential proteins for BM-MSC exo vs AT-MSC exo, BM-MSC exo vs UC-MSC exo, and AT-MSC exo vs UC-MSC exo. c GO analysis of specific proteins for each source of exosomes. (Red: biological process, green: cellular component, blue: molecular function). d KEGG analysis of specific proteins for each source of exosomes
Differentially expressed membrane proteins in exosomes
Lastly, the differentially expressed membrane proteins were analyzed in a line graph. Our analysis revealed that ATP2B1 and ATP1A1 showed high expression in AT-MSC exo, whereas ITGA2 and LRP1 showed low expression. LTGB3 and SLC44A1 showed low expression in UC-MSC exo. In contrast, ADAM9, ADAM10, CD81, CACNA2D1, NOTCH2, and HLA-A showed high expression in BM-MSC exo (Fig. 7).

Membrane protein enrichment in exosomes depends on source cell type. Relative expression of each membrane protein within a sample is depicted on slop charts and membrane protein grouped according to their enrichment or depletion in AT-MSC exo, enrichment in BM-MSC exo, and depletion in UC-MSC exo
Discussion
MSCs and their secretome have been regarded as a significant in many research areas, such as regeneration, immunity, and inflammation [14]. Extracellular vesicles (EVs) are important components of the secretome. Based on their biological origin and physical dimensions, there are two EV subgroups in particular, namely, exosomes and microvesicles (MVs). Although exosomes and MVs have various proteins in common [28], there are some important differences between them. Previous studies have shown that proteins related to extracellular matrix, adherens junctions and integrins, and vesicle-mediated transport are enriched in exosomes, whereas proteins related to mitochondria, endoplasmic reticulum, and cytoskeleton are enriched in MVs [29, 30]. With the recent advances in the field of extracellular vesicles, the methods for the isolation and purification of exosomes are constantly optimized, meeting the rigor needed for comparative studies [31]. Therefore, there is an increasing interest in the study of exosomes.
Compared with MSC therapy, exosomes have the advantages of improved safety, easier preservation, fewer side effects, and lesser ethical issues [15, 16]. Exosomes function by transversal transfer of proteins, mRNAs, and miRNAs [11]. Disease status can change the exosome contents [32], and the composition of exosomes from various sources varies. Exosomes isolated from AT-derived MSCs contained four times higher levels of neutral lysins than those from BM-MSCs [33]. Moreover, the exosomes isolated from BM- and UC-derived MSCs showed significantly different in vivo effects on U87MG glioblastoma cells [34].
In previous studies, researchers have mainly considered the accessibility of exosomes and selected exosomes specifically related to their research fields, paying less attention to the differences between exosomes derived from different sources. Some studies have explored the proteomic profile of different MSCs [8] and compared the proteomics of secretomes [22]. We specifically focused on the exosome proteome and systematically compared the characteristics of exosomes derived from MSCs of three different tissue sources to provide a reference for the selection of exosomes.
Exosomes from BM-, AT-, and UC-MSCs were similar in morphology and immunophenotype. Furthermore, the shared proteins were concentrated in the extracellular matrix receptor, which suggests common properties between the exosomes. Their corresponding functions, such as migration, proliferation, adhesion, and apoptosis, have been shown in exosomes of different sources [34,35,36].
BM-MSC exo, as the first studied stem cell exosome, were mainly evaluated in the field of the bone and cartilage regeneration, and angiogenesis [37,38,39]. Zuo et al. showed that exosomes derived from irradiated BM-MSCs could prevent bone mass loss via the Wnt/β-catenin pathway [38]. In our study, proteins related to collagen, extracellular matrix, bone regeneration, and muscle regeneration were enriched, revealing the excellent potential of exosomes in regeneration medicine. For example, Notch2 was enriched in BM-MSC exo. Notch signaling is an evolutionarily conserved pathway that can mediate critical cell communication for embryonic development and tissue regeneration. In particular, Notch2 increases skeletal remodeling in osteoprogenitor cells [40]. In addition, previous studies have shown that exosomes can be absorbed by peripheral nerve endings and transported in reverse, as well as partake in neuroplasticity [41, 42]. Interestingly, we found that ADAM10 and CA2D1 showed high expression in BM-MSC exo. ADAM10 is involved in neurodevelopment, synaptic plasticity, and dendritic spine morphology, and it has been well studied as a therapeutic target for brain diseases [43, 44]. CA2D1, an early biomarker of vestibular schwannoma, is observed in response to spinal injury and neuropathic pain [45,46,47]. Moreover, Pires et al. indicated that the secretome of BMSC might be the most advantageous choice for a therapy designed to reduce oxidative stress [22]. Further studies are needed to confirm this possible modulation in neurological diseases.
AT-MSCs were found to hold the most significant secretory function among the three types of MSCs evaluated. Considering their easy availability and fewer ethical issues, AT-MSCs may present a good choice for mass production of exosomes. Previous studies described the role of AT-MSC exo in immune dysregulation, oxidative stress, and inflammation [48, 49]. Lee et al. showed that AT-MSCs had the ability to stably produce exosomes, and PRDX 1, 2, 4, and 6 exerted antioxidant activity to protect the kidney during acute injury [49]. Our data further confirmed that exosomes play a role in immunity primarily by the regulation of leukocyte-mediated immunity. In addition, based on GO analysis, AT-MSC exo may facilitate clotting, which is consistent with the findings of Chance et al. [50]. However, the clinical application for this aspect is still rare.
Studies on UC-MSC exo have shown that they play a significant role in tissue damage repair; however, the specific mechanism are yet to be elucidated [51]. Among various research directions, skin regeneration is a hot spot [52, 53]. Zhang et al. found that 14-3-3ζ of the UC-MSC exo could coordinate Wnt4 signaling by promoting YAP phosphorylation to repair damaged skin tissue; however, there is no evidence indicating whether exosome-delivered Wnt4 is superior to exosome-free Wnt4 [52]. Herein, it was found that PAI-1 protein was enriched in UC-MSC exo. PAI-1 is a single-stranded globular glycoprotein that plays a significant role in maintaining endothelial homeostasis and regulating fibrosis [54, 55]. During tissue damage, PAI-1 contributes to faster wound healing by inhibiting uPA/tPA/plasminogen and plasminogen-dependent MMP activity [54].
Although the lack of validation is a major limitation of this study, we comprehensively analyzed the proteomic profile of exosomes from MSCs of three tissue origins and compared their shared and unique proteins. Significant differences in proteins were found among BM-, AT-, and UC-derived MSC exosomes, and molecular targets for potential therapeutic purposes were identified.
Conclusions
To the best of our knowledge, this is the first study to systematically and comprehensively analyze the human MSC-derived exosomes via proteomics. Our findings will provide helpful information for selecting optimal source cells for exosomes to improve research and therapeutic outcomes.
Availability of data and materials
All proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE partner repository with the dataset identifier PXD020948.
Abbreviations
- MSCs:
-
Mesenchymal stem cells
- BM:
-
Bone marrow
- AT:
-
Adipose tissue
- UC:
-
Umbilical cord
- NTA:
-
Nanoparticle tracking analysis
- TEM:
-
Transmission electron microscopy
- LC–MS/MS:
-
Liquid chromatography with tandem mass spectrometry
- GO:
-
Gene Ontology
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- exo:
-
Exosomes
- ECM:
-
Extracellular matrix
References
Zellner J, Johnstone B, Barry F. Mesenchymal stem cell based regenerative treatment of the knee: from basic science to clinics. Stem Cells Int. 2019;2019:7608718.
Mao XY, Pan T, Shen H. The rescue effect of mesenchymal stem cell on sodium iodate-induced retinal pigment epithelial cell death through deactivation of NF-kappaB-mediated NLRP3 inflammasome. Biomed Pharmacother. 2018;103:517–23.
Uccelli A, Moretta L, Pistoia V. Mesenchymal stem cells in health and disease. Nat Rev Immunol. 2008;8(9):726–36.
Piard C, Jeyaram A, Liu Y. 3D printed HUVECs/MSCs cocultures impact cellular interactions and angiogenesis depending on cell-cell distance. Biomaterials. 2019;222:119423.
Gazdic M, Arsenijevic A, Markovic BS. Mesenchymal stem cell-dependent modulation of liver diseases. Int J Biol Sci. 2017;13(9):1109–17.
Lin WP, Xu LL, Pan Q. Lgr5-overexpressing mesenchymal stem cells augment fracture healing through regulation of Wnt/ERK signaling pathways and mitochondrial dynamics. FASEB J. 2019;33(7):8565–77.
Zhang YL, Zhu WW, He HW. Macrophage migration inhibitory factor rejuvenates aged human mesenchymal stem cells and improves myocardial repair. Aging (Albany NY). 2019;11(24):12641–60.
Jeon YJ, Kim J, Cho JH. Comparative analysis of human mesenchymal stem cells derived from bone marrow, placenta, and adipose tissue as sources of cell therapy. J Cell Biochem. 2016;117(5):1112–25.
Kern S, Eichler H, Stoeve J. Comparative analysis of mesenchymal stem cells from bone marrow, umbilical cord blood, or adipose tissue. Stem Cells. 2006;24(5):1294–301.
Lötvall J, Hill AF, Hochberg F. Minimal experimental requirements for definition of extracellular vesicles and their functions: a position statement from the International Society for Extracellular Vesicles. J Extracell Vesicles. 2014;3:26913.
Simpson RJ, Lim JW, Moritz RL. Exosomes: proteomic insights and diagnostic potential. Expert Rev Proteomics. 2009;6(3):267–83.
Taylor DD, Taylor CG. MicroRNA signatures of tumor-derived exosomes as diagnostic biomarkers of ovarian cancer. Gynecol Oncol. 2008;110(1):13–21.
Gallo A, Tandon M, Alevizos I. The majority of microRNAs detectable in serum and saliva is concentrated in exosomes. PLoS One. 2012;7(3):e30679.
Mathieu M, Jaular LM, Lavieu G. Specificities of secretion and uptake of exosomes and other extracellular vesicles for cell-to-cell communication. Nat Cell Biol. 2019;21(1):9–17.
Lukomska B, Stanaszek L, Zuba-Surma E, Legosz P, Sarzynska S, Drela K. Challenges and controversies in human mesenchymal stem cell therapy. Stem Cells Int. 2019;2019:9628536.
Mani SA, Guo WJ, Liao MJ. The epithelial-mesenchymal transition generates cells with properties of stem cells. Cell. 2008;133(4):704–15.
Wang Y, Ma XB, Chai B. Multiscale stem cell technologies for osteonecrosis of the femoral head. Stem Cells Int. 2019;2019:8914569.
Lai RC, Arslan F, Lee MM. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010;4(3):214–22.
Zhang SP, Chuah SJ, Lai RC. MSC exosomes mediate cartilage repair by enhancing proliferation, attenuating apoptosis and modulating immune reactivity. Biomaterials. 2018;156:16–27.
Shao LB, Zhang Y, LAN BB. MiRNA-sequence indicates that mesenchymal stem cells and exosomes have similar mechanism to enhance cardiac repair. Biomed Res Int. 2017;2017:4150705.
Silva RCA, Pinheiro BM, Patrício P. Exploiting the impact of the secretome of MSCs isolated from different tissue sources on neuronal differentiation and axonal growth. Biochimie. 2018;155:83–91.
Pires AO, Pinheiro BM, Teixeira FG. Unveiling the differences of secretome of human bone marrow mesenchymal stem cells, adipose tissue-derived stem cells, and human umbilical cord perivascular cells: a proteomic analysis. Stem Cells Dev. 2016;25(14):1073–83.
Xin HQ, Li Y, Chopp M. Exosomes/miRNAs as mediating cell-based therapy of stroke. Front Cell Neurosci. 2014;8:377.
Fanale D, Taverna S, Russo A. Circular RNA in Exosomes. Adv Exp Med Biol. 2018;1087:109–17.
Dong RQ, Liu YM, Yang YX. MSC-derived exosomes-based therapy for peripheral nerve injury: a novel therapeutic strategy. Biomed Res Int. 2019;2019:6458237.
Toh WS, Lai RC, Zhang B. MSC exosome works through a protein-based mechanism of action. Biochem Soc Trans. 2018;46(4):843–53.
Théry C, Amigorena S, Raposo G. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protoc Cell Biol. 2006 Chapter 3:Unit 3.22.
Choi DS, Kim DK, Kim YK, Gho YS. Proteomics of extracellular vesicles: exosomes and ectosomes. Mass Spectrom Rev. 2015;34(4):474–90.
Haraszti RA, Didiot MC, Sapp E, et al. High-resolution proteomic and lipidomic analysis of exosomes and microvesicles from different cell sources. J Extracell Vesicles. 2016;5:32570.
Dozio V, Sanchez JC. Characterisation of extracellular vesicle-subsets derived from brain endothelial cells and analysis of their protein cargo modulation after TNF exposure. J Extracell Vesicles. 2017;6(1):1302705.
Merchant ML, Rood IM, Deegens JKJ, Klein JB. Isolation and characterization of urinary extracellular vesicles: implications for biomarker discovery. Nat Rev Nephrol. 2017;13(12):731–49.
Muntión S, Ramos TL, Campelo MD. Microvesicles from mesenchymal stromal cells are involved in HPC-microenvironment crosstalk in myelodysplastic patients. Plos One. 2016;11(2):e0146722.
Katsuda T, Suchiya R, Kosaka N. Human adipose tissue-derived mesenchymal stem cells secrete functional neprilysin-bound exosomes. Sci Rep. 2013;3:1197.
Fattore AD, Luciano R, Saracino R. Differential effects of extracellular vesicles secreted by mesenchymal stem cells from different sources on glioblastoma cells. Expert Opin Biol Ther. 2015;15(4):495–504.
Jin J, Shi YF, Gong JG. Exosome secreted from adipose-derived stem cells attenuates diabetic nephropathy by promoting autophagy flux and inhibiting apoptosis in podocyte. Stem Cell Res Ther. 2019;10(1):95.
Wang RS, Ding Q, Yaqoob U. Exosome adherence and internalization by hepatic stellate cells triggers sphingosine 1-phosphate-dependent migration. J Biol Chem. 2015;290(52):30684–96.
Furuta T, Miyaki S, Ishitobi H. Mesenchymal stem cell-derived exosomes promote fracture healing in a mouse model. Stem Cells Transl Med. 2016;5(12):1620–30.
Zuo R, Liu MH, Wang YQ. BM-MSC-derived exosomes alleviate radiation-induced bone loss by restoring the function of recipient BM-MSCs and activating Wnt/beta-catenin signaling. Stem Cell Res Ther. 2019;10(1):30.
Liang B, Liang JM, Ding JN. Dimethyloxaloylglycine-stimulated human bone marrow mesenchymal stem cell-derived exosomes enhance bone regeneration through angiogenesis by targeting the AKT/mTOR pathway. Stem Cell Res Ther. 2019;10(1):335.
Vollersen N, Borgmeyer IH, Cornils K. High bone turnover in mice carrying a pathogenic Notch2 mutation causing Hajdu-Cheney syndrome. J Bone Miner Res. 2018;33(1):70–83.
Ren R, Tan XH, Zhao JH. Bone marrow mesenchymal stem cell-derived exosome uptake and retrograde transport can occur at peripheral nerve endings. Artif Cells Nanomed Biotechnol. 2019;47(1):2918–29.
Zaldivar EER, Sapiéns MAH, Minjarez B. Potential effects of MSC-derived exosomes in neuroplasticity in Alzheimer’s disease. Front Cell Neurosci. 2018;12:317.
Marcello E, Borroni B, Pelucchi S, Gardoni F, Di Luca M. ADAM10 as a therapeutic target for brain diseases: from developmental disorders to Alzheimer’s disease. Expert Opin Ther Targets. 2017;21(11):1017–26.
Hsia HE, Tüshaus J, Brummer T, Zheng Y, Scilabra SD, Lichtenthaler SF. Functions of ‘A disintegrin and metalloproteases (ADAMs)’ in the mammalian nervous system. Cell Mol Life Sci. 2019;76(16):3055–81.
Huang X, Xu J, Shen Y, et al. Protein profiling of cerebrospinal fluid from patients undergoing vestibular schwannoma surgery and clinical significance. Biomed Pharmacother. 2019;116:108985.
Newton RA, Bingham S, Case PC, Sanger GJ, Lawson SN. Dorsal root ganglion neurons show increased expression of the calcium channel alpha2delta-1 subunit following partial sciatic nerve injury. Brain Res Mol Brain Res. 2001;95(1–2):1–8.
Yaksh TL. Calcium channels as therapeutic targets in neuropathic pain. J Pain. 2006;7(1 Suppl 1):S13–30.
Nojehdehi S, Soudi S, Hesampour A. Immunomodulatory effects of mesenchymal stem cell-derived exosomes on experimental type-1 autoimmune diabetes. J Cell Biochem. 2018;119(11):9433–43.
Lee JH, Ha DH, Kyu H. Goeproducible large-scale isolation of exosomes from adipose tissue-derived mesenchymal stem/stromal cells and their application in acute kidney injury. Int J Mol Sci. 2020;21(13):4774.
Chance TC, Rathbone CR, Kamucheka RM. The effects of cell type and culture condition on the procoagulant activity of human mesenchymal stromal cell-derived extracellular vesicles. J Trauma Acute Care Surg. 2019;87(1S Suppl 1):S74–82.
Yin SQ, Ji C, Wu PP. Human umbilical cord mesenchymal stem cells and exosomes: bioactive ways of tissue injury repair. Am J Transl Res. 2019;11(3):1230–40 eCollection 2019.
Zhang B, Shi YH, Gong A. HucMSC exosome-delivered 14-3-3zeta orchestrates self-control of the Wnt response via modulation of YAP during cutaneous regeneration. Stem Cells. 2016;34(10):2485–500.
Hoang DH, Nguyen TD, Nguyen PH. Differential wound healing capacity of mesenchymal stem cell-derived exosomes originated from bone marrow, Adipose Tissue and Umbilical Cord Under Serum- and Xeno-Free Condition. Front Mol Biosci. 2020;7:119.
Ghosh AK, Vaughan DE. PAI-1 in tissue fibrosis. J Cell Physiol. 2012;227(2):493–507.
Kanno Y. The role of fibrinolytic regulators in vascular dysfunction of systemic sclerosis. Int J Mol Sci. 2019;20(3):619.
Acknowledgements
We thank Pei Zhang for his help in proteomic analysis and manuscript preparation.
Funding
This work was supported by the National Natural Science Foundation of China [No. 81672168] and Hubei Province health and family planning scientific research project (Grant No. WJ2019Q028).
Author information
Authors and Affiliations
Contributions
ZG Wang: conception and design, collection and/or assembly of data, data analysis and interpretation, and manuscript writing; ZY He: collection and/or assembly of data, data analysis and interpretation, and manuscript writing; S Liang: conception and design, and collection and/or assembly of data; Q Yang: data analysis and interpretation; AM Chen and P Cheng: conception and design, manuscript writing, and final approval of manuscript. The authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable
Consent for publication
Not applicable
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: SF. 1.
Workflow of exosome preparation and mass spectrometry. Human bone marrow (BM), umbilical cord (UC), and adipose tissue (AT)-derived MSCs were cultured, and exosome samples were obtained from the different cell line types by ultracentrifugation. Exosomes were subjected to proteomic analysis (nine samples, LC–MS/MS). Proteins were quantified using the label-free quantification method iBAQ.
Additional file 2: SF. 2.
Flow cytometric analysis of cell immunophenotype of MSCs.
Additional file 3: SF. 3.
Sample repeatability test and quality control of mass spectrometry. (a) The distribution of peptide length is showed in a histogram. (b) Protein mass and coverage distribution. (c) Pearson correlation coefficient between all samples is presented. (d) Protein quantitative principal component analysis results of all samples.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, Zg., He, Zy., Liang, S. et al. Comprehensive proteomic analysis of exosomes derived from human bone marrow, adipose tissue, and umbilical cord mesenchymal stem cells. Stem Cell Res Ther 11, 511 (2020). https://doi.org/10.1186/s13287-020-02032-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13287-020-02032-8