
Abstract
Background
It has been hypothesized that the ParaHox gene Gsx patterned the foregut of the last common bilaterian ancestor. This notion was corroborated by Gsx expression in three out of four lophotrochozoan species, several ecdysozoans, and some deuterostomes. Remarkably, Gsx is also expressed in the bilaterian anterior-most central nervous system (CNS) and the gastropod and annelid apical organ. To infer whether these findings are consistent with other mollusks or even lophotrochozoans, we investigated Gsx expression in developmental stages of representatives of two other molluscan classes, the scaphopod Antalis entalis and the cephalopod Idiosepius notoides.
Results
Gsx is not expressed in the developing digestive tract of Antalis entalis and Idiosepius notoides. Instead, it is expressed in cells of the apical organ in the scaphopod trochophore and in two cells adjacent to this organ. Late-stage trochophores express Aen-Gsx in cells of the developing cerebral and pedal ganglia and in cells close to the pavilion, mantle, and foot. In postmetamorphic specimens, Aen-Gsx is expressed in the cerebral and pedal ganglia, the foot, and the nascent captacula. In early squid embryos, Ino-Gsx is expressed in the cerebral, palliovisceral, and optic ganglia. In late-stage embryos, Ino-Gsx is additionally expressed close to the eyes and in the supraesophageal and posterior subesophageal masses and optic lobes. Developmental stages close to hatching express Ino-Gsx only close to the eyes.
Conclusions
Our results suggest that Gsx expression in the foregut might not be a plesiomorphic trait of the Lophotrochozoa as insinuated previously. Since neither ecdysozoans nor deuterostomes express Gsx in their gut, a role in gut formation in the last common bilaterian ancestor appears unlikely. Gsx is consistently expressed in the bilaterian anterior-most CNS and the apical organ of lophotrochozoan larvae, suggesting a recruitment of Gsx into the formation of this organ in the Lophotrochozoa. The cephalopod posterior subesophageal mass and optic ganglia and the scaphopod pedal ganglia also express Gsx. In summary, Gsx expression only appears to be conserved in the anterior-most brain region during evolution. Accordingly, Gsx appears to have been recruited into the formation of other expression domains, e.g., the apical organ or the foregut, in some lophotrochozoans.
Similar content being viewed by others

Background
The Hox and ParaHox gene clusters are considered to be derived from a hypothetical ProtoHox cluster by duplication [1]. Both belong to the homeobox gene family and exhibit highly conserved amino acid sequences in phylogenetically distantly related animals [1, 2]. In the majority of bilaterians investigated, it has been shown that Hox genes are expressed in tempo-spatial collinearity during development, in particular in neuroectodermal domains [2, 3]. Cephalopod and gastropod mollusks were among the first examples among bilaterians that apparently do not exhibit such a collinear mode of Hox gene expression [4, 5]. Tempo-spatial collinear expression of the three ParaHox genes has also been proposed for the last common bilaterian ancestor [1]. It has been hypothesized that Gsx was expressed in the foregut, Xlox in the midgut, and Cdx in the hindgut in the last common bilaterian ancestor [1, 6]. While Xlox expression in the midgut and Cdx expression in the hindgut was found in various bilaterians, no Gsx expression has been reported in the foregut of any deuterostome representative to date [1, 6]. This was explained by the fact that the blastopore does not develop into the prospective mouth in deuterostomes. Deuterostomes instead evolved a new mouth and hence Gsx might have lost its role in patterning the anterior-most region of the digestive tract. Interestingly, the deuterostome hemichordate Ptychodera flava, for example, does express Gsx around the blastopore, however, apparently not in the digestive tract of subsequent developmental stages [7]. Holland anticipated that protostome invertebrates may show Gsx expression in the foregut since their blastopore usually does become the future mouth [1, 6].
Data on the ecdysozoan and lophotrochozoan condition show, however, an ambiguous picture. While all ecdysozoans investigated so far do not appear to express Gsx in their digestive tract, the situation in lophotrochozoans is less clear (Table 1). The annelids Platynereis dumerilii and Nereis virens and the gastropod Gibbula varia express Gsx in their foregut [12–14], while the annelid Capitella teleta does not [15]. Comparisons with the condition in the Cnidaria, the putative bilaterian sister group, do not appear to contribute to inferring the ancestral state of Gsx expression in the Bilateria since the different germ layers cannot be homologized convincingly among the Cnidaria and the Bilateria. In addition, Gsx expression patterns are not consistent among cnidarians. In the planula larvae of Nematostella vectensis, Clytia hemisphaerica, and Podocoryne carnea [9–11], Gsx is expressed in the endoderm, while it is expressed in the ectoderm of the planula of Acropora millepora [8].
Gsx is also involved in the development of the CNS in bilaterians, and it is expressed in distinct cells of the apical organ in the gastropod mollusk G. varia and the annelid P. dumerilii (Table 1; [12, 13]). In addition, Gsx expression was also found in the radula sac, a molluscan evolutionary novelty [12]. Recent phylogenomic analyses on mollusks have revived a classical hypothesis placing the Aculifera, i.e., the worm-shaped and spicule-bearing aplacophorans and the eight-shelled polyplacophorans, as a sister group to the Conchifera [30–32]. The Conchifera is an anatomically diverse clade comprising scaphopods, gastropods, bivalves, monoplacophorans, and cephalopods. Until now, conchiferan interrelationships are unsettled, and attempts to infer the evolution of their body plans are scarce (c.f. [31, 32]; but see [33, 34]).
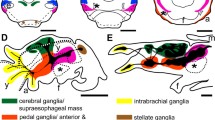
The present study deals with two conchiferans, the scaphopod Antalis entalis Jeffreys 1869 and the cephalopod squid Idiosepius notoides Berry, 1921 (Fig. 1). Adult scaphopods and cephalopods exhibit a pronounced dorso-ventral body axis as opposed to the majority of bilaterians that exhibit a pronounced antero-posterior body axis (Fig. 1). In adult scaphopods, the mouth and foot are located ventrally, while the pavilion (i.e., the mantle cavity opening on the opposite side) marks the dorsal pole (Fig. 1b). In adult cephalopods, the funnel and (parts of) the circumoral brachial crown are considered to be homologous to the foot of other mollusks [35] (Fig. 1c). The brachial crown and the funnel define the ventral side, while the mantle apex is located dorsally (Fig. 1c). Thus, the dorso-ventral axis constitutes the major body axis in these animals. In both clades, the cerebral ganglia are located anteriorly (labeled blue in Fig. 1), while the statocysts are located at the posterior pole (dashed circles in Fig. 1b, c).

Dorso-ventral and antero-posterior body axes in adult mollusks. Dorsal (d)–ventral (v), anterior (a)-posterior (p) axes indicate the orientation. The (anlagen of the) cerebral ganglia/commissure (blue) are located anteriorly, while the statocysts (sc) are located posteriorly (statocysts are absent in polyplacophorans). The trajectory of the digestive tract is lined in red. a An adult polyplacophoran with a pronounced antero-posterior axis, a dorsal shell (sh) and ventral mouth (mo) and foot (f). b The mouth and foot of the adult scaphopod Antalis entalis are located ventrally, while the scaphopod pavilion (pv) lies dorsally. c The mouth and brachial crown (brc) of the adult cephalopod Idiosepius notoides are located ventrally, while the mantle apex (ma) is located dorsally. an anus, fn funnel, m mantle and pt prototroch
Ontogeny of the scaphopod Antalis entalis and the cephalopod Idiosepius notoides
In the scaphopod, A. entalis gastrulation occurs at 12 h after fertilization (hpf) at 21–23 °C (Fig. 2a). At 14 hpf, a trochophore larva develops that exhibits an episphere with an apical organ and tuft (red dashed circles in Fig. 2). The episphere is divided from the hyposphere by a prototroch (Fig. 2b; see also [36–38]). The gastropod trochophore resembles the latter, but while the apical region develops into the prospective anterior region in gastropods, it develops into the prospective ventral region in scaphopods (see scaphopod condition in Fig. 2; [12, 36–38]). The blastopore of the gastrula develops into the mouth in A. entalis and lecithotrophic early-stage trochophore larvae already possess a through-gut with mouth and anus (Fig. 2a, b). The apical organ exhibits two serotonin-like immunoreactive cell somata (labeled red in Fig. 2b), and the nascent shell field is located in the anterior region of the hyposphere (Fig. 2b; [39]). The apical organ of mid-stage trochophore larvae (21 hpf) possesses four serotonin-like immunoreactive cells that are located next to two lateral cells that do not belong to this sensory organ (Fig. 2c). The episphere including the apical organ migrates in direction of the dorsal side and the cerebral ganglia develop below the latter and ventrally to the esophagus (Fig. 2c) [40]. In mid-stage trochophore larvae, the statocysts become visible in the foot (black dashed circles in Fig. 2c), and the dorsal-most region of the mantle, the pavilion, serves as second opening of the mantle cavity. In late-stage trochophore larvae (63 hpf) and advanced developmental stages, the dorso-ventral body axis elongates considerably and the foot grows out into ventral direction (Fig. 2d). The apical organ migrates in dorsal direction and most probably disappears with all serotonin-like immunoreactive cells in metamorphic competent trochophore larvae (Fig. 2e). The cerebral ganglia are located anteriorly (blue domain in Fig. 2e) and connect to the pedal ganglia that are located ventrally to the statocysts (green domain in Fig. 2e). During metamorphosis, trochophores settle and are able to retract their prototroch and foot into the shell. Postmetamorphic individuals do not exhibit a prototroch and possess two captacula anlagen. These are the forerunners of the multiple cephalic tentacles that are used to collect food (Fig. 2f). Settled individuals show a well-differentiated midgut gland, a pronounced trilobed foot, and a buccal cone with a mouth (Fig. 2f). Notably, adult scaphopods generally lack eyes and a distinct head.

Ontogeny of the scaphopod Antalis entalis. All lateral views and dorsal (d)–ventral (v), anterior (a)–posterior (p) axes indicate the orientation. Note the prototroch (dark gray), the statocysts (black dashed encircled), the apical organ (red dashed circle) with serotonin-like immunoreactive cells (red) and apical tuft (at), the cerebral ganglia (blue) and the pedal ganglia with connectives (green). The mouth (mo) is located ventrally to the foot (f), while the anus (an) is located ventrally to the pavilion (pv; dorsal mantle opening). The light blue line outlines the shell. a Gastrula with blastopore (bp) (12 hpf). b The early-stage trochophore (14 hpf) exhibits an episphere (ep) with an apical organ with two serotonin-like immunoreactive cells. The prototroch (pt) divides the episphere from the hyposphere (hp). In the anterior hyposphere, the shell field (shf) develops. c Mid-stage trochophore (21 hpf) possesses four serotonin-like immunoreactive cells in addition to two lateral cells (not shown) that do not belong to the apical organ. d Late-stage trochophore (63 hpf). e Metamorphic competent trochophore (70 hpf). f Postmetamorphic and settled specimen (114 hpf). Data on serotonin-like immunoreactive cells and on the location of the CNS derive from [36–38, 40]. cp captacula, mg midgut gland. Scale bars 50 µm
In the cephalopod I. notoides, cleavage only occurs on the cytoplasmic cap of the yolk-rich embryo (stages 2–13 according to [41]; reviewed in [42]). During the gastrulation process at stage 13, the outermost blastomere rows migrate below the inner blastomeres and a two-layered epithelium is formed on the yolk syncytium. Stage 18–19 individuals are roundish in shape and various organ systems are formed as placodes, among others the CNS, the arms, the funnel, the eyes, the mantle, and the arms (Fig. 3a). The brachial ganglia are located in the anlagen of the arms, the stellate ganglia are situated in the anterior portion of the mantle, and the optic ganglia are connected to both eyes (Fig. 3a). The cerebral ganglia develop dorsally to the mouth opening, the palliovisceral ganglia lie between the mantle and the statocysts, and the pedal ganglia are located ventrally to the statocysts. The dorso-ventral axis of stage 23 individuals is elongated compared to earlier stages (Fig. 3b). The esophagus is situated adjacent to the inner yolk duct and the individual ganglia connected to each other. Stage 25 embryos exhibit a more centralized brain and all individual central ganglia are termed brain masses herein in accordance with the classical literature (Fig. 3c). The cerebral ganglia give rise to the supraesophageal mass, the pedal ganglia develop into the anterior and middle subesophageal masses, and the palliovisceral ganglia are then termed posterior subesophageal mass [43]. Contrary, the peripheral stellate and brachial ganglia are still termed ganglia. The dorso-ventral body axis of stage 30 hatchlings is more elongated, and the CNS is more centralized than in earlier stages (Fig. 3d).

Ontogeny of the cephalopod Idiosepius notoides. All lateral views and dorsal (d)–ventral (v), anterior (a)–posterior (p) axes indicate the orientation. Core brain regions are highlighted, while innervation of peripheral target is omitted. The major ganglia are color coded: cerebral ganglia (blue), pedal ganglia (green), palliovisceral ganglia (violet), brachial ganglia (orange), and stellate ganglia (brown). The digestive tract is only indicated in the brain region, and the inner yolk duct and the entire external yolk sac are omitted for clarity. a Stage 19 embryos exhibit anlagen of all major ganglia. b The individual ganglia are connected in stage 23 embryos. Note that it is difficult to identify the origin of the brain mass connecting cerebral and pedal ganglia (herein labeled turquoise). c Stage 25 embryos exhibit a more centralized brain with the supraesophageal mass (blue), the posterior subesophageal mass (violet), the middle and anterior subesophageal mass (green), the brachial ganglia (orange), and the stellate ganglia (brown). d Hatchlings (stage 30) possess a CNS that largely resembles the one of adults. Please note that the stellate ganglia are not shown. ar arm, ey eye, fn funnel, m mantle, mo mouth and y yolk. Scale bars 150 µm
In this study, we describe hitherto unknown Gsx orthologs and their expression domains in the scaphopod Antalis entalis and the cephalopod squid Idiosepius notoides (Fig. 1). Our results question the widely assumed role of Gsx in patterning the foregut of the last common bilaterian ancestor and highlight similarities as well as differences among mollusks, lophotrochozoans, and bilaterians.
Methods
Collection and culture of animals
Adults of the scaphopod Antalis entalis were collected from approximately 30 m depth by the staff of the research vessel Neomys off the coast of Roscoff (France). Individuals were immediately transferred into dishes filled with seawater (see also [39]). Spawning occurred spontaneously or was induced by heat shocks, i.e., individuals were exposed to alternating water temperatures. Unfertilized eggs were rinsed several times and fertilized with sperm. Early- and mid-stage trochophore larvae were cultured in Millipore-filtered seawater (MFSW) with 50 mg streptomycin sulfate and 60 mg penicillin G per liter MFSW. Early cleavage stages, metamorphic competent larvae, and settled individuals were cultured in MFSW without antibiotics. Water was changed every other day. Metamorphosis occurred spontaneously or was induced by adding shell-gravel from the collection site.
Adults of the pygmy squid Idiosepius notoides were dip-netted in the sea grass beds of Moreton Bay, Queensland, Australia. Embryos were cultured and staged as described previously [43]. Development from freshly laid fertilized eggs (stage 1) to hatchlings (stage 30) takes 9–10 days at 25 °C.
RNA extraction and fixation of animals
For Antalis entalis, a total of several hundred individuals of mixed developmental stages including early cleavage stages, trochophore larvae, metamorphic competent individuals, and early juveniles were collected and stored at −20 to −80 °C in RNAlater (Lifetechnologies, Vienna, Austria). RNA was extracted with a RNA extraction kit (Qiagen, Roermond, Netherlands) and stored at −80 °C.
For Idiosepius notoides, the egg jelly and chorion were removed from approximately 300 specimens covering freshly laid zygotes (stage 1) to hatchlings (stage 30). RNA was extracted using TriReagent according to the manufacturer’s instructions (Astral Scientific Pty. Ltd., Caringbah, Australia, see also [44]). Individuals of all the above-described developmental stages were fixed for in situ hybridization experiments as previously described [44].
RNAseq and transcriptome assembly
Total RNA from pooled developmental stages of Antalis entalis was sequenced by Illumina technology (Eurofins, Ebersberg, Germany). Paired-end reads of an average read length of 100 bp were obtained and subsequently filtered (rRNA removal). Adapter and low-quality sequences were trimmed, normalized, and assembled de novo into contigs with the assembler Trinity [45].
RNA from developmental stages of Idiosepius notoides was sequenced by 454 and Illumina technology (both Eurofins) as described previously [44]. After filtering, the adapter and low-quality reads were trimmed, normalized, and assembled de novo by Eurofins (454 transcriptome) or using Trinity (Illumina transcriptome).
Alignment and phylogenetic analysis
Known amino acid sequences of bilaterian Gsx orthologs were retrieved from the National Center for Biotechnology Information (NCBI) and used in BLAST searches against both assembled transcriptomes. Amino acid sequences were aligned using ClustalX v.2.0 [46], trimmed by hand with the program AliView [47], and only conserved regions were retained (Fig. 4; untrimmed alignments are available upon request). This alignment was used to construct the neighbor-joining tree shown in Fig. 5 using the JTT matrix with 1000 bootstrap replicates within the Phylip v.3.695 [48] suite of programs.

Alignment of amino acid sequences of bilaterian ParaHox and Hox gene orthologs. The Gsx amino acid sequences of Idiosepius notoides and Antalis entalis (red-labeled) cluster with their bilaterian orthologs. Molluscan Gsx orthologs are labeled in red and annelid orthologs in blue. GenBank or JGI accession numbers of amino acid sequences used for the alignment and the phylogenetic tree: Pmi-Xlox (Patiria miniata) AGK89734.1; Bfl-Xlox (Branchiostoma floridae) AAC39016.1; Pdu-Xlox (P. dumerilii) ACH87541.1; Gva-Xlox (G. varia) ADJ18240.1; Lgi-Xlox (Lottia gigantea) JGI e_gw1.80.260.1; Ino-Gsx (I. notoides) KT380894; Lgi-Gsx (L. gigantea) JGI e_gw1.30.54.1; Aen-Gsx (A. entalis) KT380895; Pdu-Gsx (P. dumerilii) ACH87538.1; Gva-Gsx (G. varia) KT380896; Pmi-Gsx (P. miniata) AGK89736.1; Dme-Ind (intermediate neuroblasts defective, D. melanogaster) AAC97116.1; Tca-Ind (intermediate neuroblasts defective, Tribolium castaneum) NP_001034494.1; Bfl-Gsx (Branchiostoma floridae) AAC39015.1; Pdu-Cdx (P. dumerilii) ABA29777.3; Dme-Cdx (D. melanogaster) AAF53923.1; Pmi-Cdx (P. miniata) AGK89735.1; Bfl-Cdx (B. floridae) AAC39017.1; Tca-Cdx (T. castaneum) NP_001034498.1; Gva-Cdx (G. varia) ADJ18241.1; Lgi-Cdx (L.gigantea) JGI estExt_Genewise1.C_sca_850005; Pdu-Hox1 (P. dumerilii) AFJ91921.1; Bla-Hox1 (B. lanceolatum) ACJ74382.1. Pdu-Lox5 (P. dumerilii) AFJ91925.1; Pdu-Post2 (P. dumerilii) AFJ91927.1

Phylogenetic analysis including the amino acid sequences of bilaterian ParaHox and Hox gene orthologs. This phylogenetic analysis of amino acid sequences of bilaterian orthologs of Gsx, Cdx, Xlox, and Hox1 genes confirms the identity of Aen-Gsx and Ino-Gsx (red-labeled/bold). Molluscan Gsx orthologs are labeled in red and annelid orthologs in blue. Percentage of bootstrap support is shown when over 50 % and only for the major clades. See capture of Fig. 4 for accession numbers
Molecular isolation of RNA transcripts
First-strand cDNA synthesis of the RNA pooled from different developmental stages of Antalis entalis and Idiosepius notoides, respectively, was carried out by reverse transcription using the First-strand cDNA Synthesis Kit for rt-PCR (Roche Diagnostics GmbH, Mannheim, Germany). Identified Gsx orthologs of A. entalis and I. notoides were used to design gene-specific primers, and PCR products were size-fractioned by gel electrophoresis. Gel bands of the expected length were excised and cleaned up using a QIAquick Gel Extraction Kit (QIAgen, Hilden, Germany). By insertion into pGEM-T Easy Vectors (Promega, Mannheim, Germany), cleaned-up products were cloned. Plasmid minipreps were grown overnight, cleaned up with the QIAprep Spin MiniprepKit (QIAgen), and sent for sequencing. The sequenced minipreps matched both transcripts identified as Aen-Gsx and Ino-Gsx in the phylogenetic analysis (Figs. 4, 5).
Probe synthesis and whole-mount in situ hybridization
Riboprobe templates were amplified via standard PCR from miniprepped plasmids using M13 forward and reverse primers. In vitro transcription reactions were performed with these templates, digoxigenin-UTP (DIG RNA Labeling Kit, Roche Diagnostics) and SP6/T7 polymerase (Roche Diagnostics GmbH) for the syntheses of antisense riboprobes according to the manufacturer’s instructions. For whole-mount in situ hybridization experiments, specimens were rehydrated into PBT (PBS + 0.1 % Tween-20) and treated with Proteinase-K (25 µg/ml for Idiosepius notoides and 45 µg/ml for Antalis entalis) in PBT at 37 °C for 10 min. Specimens were prehybridized in hybridization buffer for 4 h at 50 °C (A. entalis) or 65 °C (I. notoides), and hybridization with a probe concentration of 0.5 μg/ml (I. notoides) to 1 μg/ml (A. entalis) was carried out overnight at 50 °C (A. entalis) or 65 °C (I. notoides). For A. entalis as well as I. notoides, a minimum of 20 individuals per stage were investigated, and negative controls were carried out with sense probes for all genes and developmental stages. The majority of whole-mount preparations were cleared in a solution of benzyl benzoate/benzyl alcohol (2:1), mounted on objective slides, and analyzed. Preparations were documented with an Olympus BX53 Microscope (Olympus, Hamburg, Germany). In addition, scaphopod developmental stages were scanned with a Leica confocal SP5 II microscope (Leica Microsystems, Wetzlar, Germany) using bright-field, autofluorescence, and reflection mode scans [49]. If necessary, images were processed with Adobe Photoshop 9.0.2 software (San Jose, CA, USA) to adjust contrast and brightness.
Histology
After in situ hybridization experiments, developmental stages of Antalis entalis were post-fixed in 100 % EtOH and embedded in agar low viscosity resin (Agar Scientific, Essex, United Kingdom). Specimens were semithin sectioned with a diamond knife (Histo Jumbo Diatome) at a thickness of 0.5 µm with an ultramicrotome (Leica EM UC6, Wetzlar, Germany). Sections were mounted on objective slides, stained with Eosin using standard histological protocols, and covered with cover slips. Alternatively, after in situ hybridization, specimens were embedded in O.C.T. medium (VWR, Vienna, Austria) and cut into 15–30 µm cryosections with a cryotome (Leica CM 3050S). Sections were stained with Dapi (Sigma-Aldrich, St. Louis, MO, USA) and Cellmask Green plasma membrane stain (ThermoFisher, Waltham, MA, USA) in order to stain cell nuclei and cell membranes. Sections were mounted in Fluoromount G (Southern Biotech, Birmingham, Alabama, USA) and covered with cover slips. Semithin as well as cryotome sections was documented with an Olympus BX53 Microscope (Olympus).
Statement of ethical approval
Developmental stages and adults of the pygmy squid Idiosepius notoides were collected, anesthetized, and fixed according to internationally recognized standards (University of Queensland Animal Welfare Permit No. 158/09 “The cultivation of Idiosepius (pygmy squid) for studies in developmental biology” to BMD).
Results
Aen-Gsx expression in developmental stages of the scaphopod Antalis entalis
The alignment of multiple amino acid sequences shows that Aen-Gsx and Ino-Gsx exhibit high sequence similarity with their bilaterian orthologs (Fig. 4). Aen-Gsx as well as Ino-Gsx clusters with their bilaterian orthologs in the phylogenetic analysis (Fig. 5).
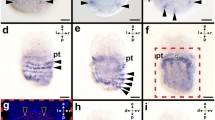
Aen-Gsx is first expressed in two cells in the episphere of early-stage trochophore larvae (14 hpf) (arrowheads in Fig. 6a, b, f–h). In addition, each one Aen-Gsx-expressing cell is located laterally to the anus (“4” in Fig. 6c–e, g). In mid-stage trochophore larvae (19 hpf), Aen-Gsx is expressed in two flask-shaped cells of the apical organ (“1” in Figs. 7a, b, 8a, b) and two lateral cells (“2” in Figs. 7a, b, 8a, b). Mid-stage trochophore larvae at 21 hpf also exhibit both above-mentioned groups of cells (Fig. 7c, d). While two Aen-Gsx-expressing cells are located in the apical organ (“1” in Figs. 7d, g, 8a, b), both lateral Aen-Gsx-expressing cells do not appear to belong to the latter (“2” in Figs. 7d, f, 8a, b). Another pair of Aen-Gsx-expressing cells is present on the posterolateral side of the mantle (“3” in Figs. 7d, 8a, b) and below the mantle laterally to the anus (“4” in Figs. 7e, 8a, b). In late-stage trochophore larvae, two clusters of Aen-Gsx-expressing cells are present at the base of both captacula, in a region where the future cerebral ganglia develop (Fig. 8c, d; black dashed circle in Fig. 9a). Two additional clusters of Aen-Gsx-expressing cells are located ventro-laterally to the statocysts (Fig. 8c, d; red dashed circle in Fig. 9b). Two flask-shaped Aen-Gsx-expressing cells are located in the region of the pavilion (Fig. 8c, d; arrowheads in Fig. 9b). Another group of Aen-Gsx-expressing cells is located in the ventral portion of the foot (Fig. 8c; green dashed circle in Fig. 9c). In some individuals, one or two flask-shaped Aen-Gsx-expressing cells are visible in the region close to the cerebral ganglia (data not shown). In metamorphic competent trochophore larvae, each one Aen-Gsx-expressing cell cluster is situated ventro-laterally to the statocysts in the region of the pedal ganglia (green dashed circles in Fig. 10b, c) and in the region of the cerebral ganglia (red dashed circles in Fig. 10b, c). Other Aen-Gsx-expressing cell clusters are located in the region of the nascent captacula (black dashed circles in Fig. 10). Postmetamorphic specimens exhibit a similar distribution of Aen-Gsx-expressing cells in the regions of the cerebral and pedal ganglia (Figs. 8e, f, 11a–c). Aen-Gsx-expressing cells are also present in the region of the nascent captacula (arrowheads in Fig. 11a, b) and in the ventral foot region (Figs. 8e, f, 11a–c).

Expression of Aen-Gsx in early-stage trochophore larvae (14 hpf) of the scaphopod Antalis entalis. Dorsal (d)–ventral (v), anterior (a)–posterior (p), and left (l)–right (r) axes indicate the orientation. a–d, f, h, i are whole-mount preparations, while e and g are cryosections (cell membranes stained green, cellmask/cell nuclei labeled blue, Dapi). a Each two cells (arrowheads) express Aen-Gsx on both sides of the episphere. b–e Each one cell (4) expresses Aen-Gsx bilaterally to the anus. f Four Aen-Gsx-expressing cells (arrowheads) are located in episphere. g Aen-Gsx-expressing cells on the left side of the episphere (both cells on the right side are on another cryosection) and close to the anus (4). h, i BBA cleared specimen highlighting the position of mouth (mo), anus, and digestive tract (red stippled line) in relation to Aen-Gsx-expressing cells (arrowhead). an anus, pt prototroch. Scale bars 50 µm

Expression of Aen-Gsx in mid-stage trochophore larvae of the scaphopod Antalis entalis. Dorsal (d)–ventral (v), anterior (a)–posterior (p), and left (l)–right (r) axes indicate the orientation. Asterisks mark the mouth opening. a Two cells (1) of the apical organ express Aen-Gsx in a 19 hpf old larva. Another two cells (2) faintly express Gsx and are located laterally to the former ones (the right cell is located in another cryotome section). b Apical view of 19 hpf old, same staged specimen showing both cells (1) in the apical organ and faintly both lateral cells 2. c Lateral view of a 21 hpf mid-stage trochophore larva. In this optical section, both Aen-Gsx-expressing cells in the apical organ and both Aen-Gsx-expressing cells (4) on both sides below the mantle lateral to the anus. d Same specimen as shown in c showing a pair of Aen-Gsx-expressing cells (3) on the latero-posterior-most side of the mantle. Both cells in the apical organ (1) as well as lateral to the latter (2) express Aen-Gsx. Inset: magnification of the Aen-Gsx-expressing cells (3) in a same staged specimen. e Same specimen as shown in c, d with both Gsx-expressing cells (4) on both sides below the mantle lateral to the anus. f This histological section shows a lateral Aen-Gsx-expressing cell (2) that penetrates the epidermis. g This histological section shows an Aen-Gsx-expressing cell (1) belonging to the apical organ. an anus, f foot, m mantle, mc mantle cavity and pt prototroch. Scale bars 50 µm

Summary of Gsx expression (blue) during the development of the scaphopod Antalis entalis and the cephalopod Idiosepius notoides. Dorsal (d)–ventral (v), anterior (a)–posterior (p), and left (l)–right (r) axes indicate the orientation and are the same in each of the six columns. Shown Gsx-expressing cell somata do not represent absolute numbers. a–f Sketch drawing depicting a mid-stage trochophore (a, b), a late-stage trochophore (c, d), and a postmetamorphic (settled) individual (e, f) of Antalis entalis. Gsx-expressing cell somata are labeled with red numbers 1–4 (c.f. Figures 6, 7). g–l Sketch drawing depicting developmental of stage 19 (g, h), stage 25 (i, j), and stage 28 (k, l, yolk sac removed) of the pygmy squid Idiosepius notoides. ab anterior basal lobe, ao apical organ, ar arm, bc buccal cone, cg cerebral ganglion, cp captacula, ey eye, f foot, fn funnel, ib inferior buccal lobe, if inferior frontal lobe, m mantle, mf mantle fold, mo mouth, o optic ganglion/lobe, pb posterior basal lobe, pg pedal ganglion, pt prototroch, pv pavilion, pvg palliovisceral ganglion, ps posterior subesophageal mass and y yolk. Scale bars a–f 50 µm, g–l 150 µm

Expression of Aen-Gsx during late-stage trochophores of the scaphopod Antalis entalis. Dorsal (d)–ventral (v) and left (l)–right (r) axes indicate the orientation. The statocysts are labeled with asterisks. Optical sections from anterior (a) to posterior (b) in a 63 hpf old trochophore (all with same orientation and scale bar as indicated in a). a One cell cluster on each side expresses Aen-Gsx in the region of the cerebral ganglia (black dashed circles). b Two cell clusters (red dashed circles) express Aen-Gsx ventro-laterally to the statocysts, in the region of the prospective pedal ganglia. A pair of Aen-Gsx-expressing cells is located in the region of the pavilion (pv). c Aen-Gsx is expressed in the ventral portion of the foot (green dashed circle). m mantle, pt prototroch. Scale bar 50 µm

Expression of Aen-Gsx in metamorphic competent larvae (70 hpf) of the scaphopod Antalis entalis. In all three cryosections, anterior (a)–posterior (p), dorsal (d)–ventral (v) and left (l)–right (r) axes indicate the orientation. The statocysts are labeled with asterisks. a Aen-Gsx is expressed in the region of the cerebral (cg) and pedal (pg) ganglia as well as in the region of the anterior foot (f) (arrow). Note the Aen-Gsx expression in the region of the nascent captacula (black dashed circle). b, c Aen-Gsx expression in the region of the nascent captacula (black dashed circle), the cerebral ganglia (red dashed circle), and pedal ganglia (green dashed circle). The bright-field micrograph (b) and the nuclear counterstain (blue, Dapi) show the same individual of the same orientation and size. Note the perikaryal layers around the cerebral and pedal ganglia shown in c. m mantle, mg midgut gland, pt prototroch, pv pavilion. Scale bars 100 µm

Expression of Aen-Gsx during postmetamorphic development (114 hpf) of the scaphopod Antalis entalis. Anterior (a)–posterior (p), dorsal (d)–ventral (v), and left (l)–right (r) axes indicate the orientation. The statocysts are labeled with asterisks. a, b Aen-Gsx expression in the region of the cerebral (cg) and pedal ganglia (pg) as well as the ventral foot (f) and the ventral region of the nascent captacula (arrowheads). c Inverted bright-field image of Aen-Gsx expression (green) with nuclear counterstain (blue, Dapi) of a cryosection (see inset for original stain). Aen-Gsx is expressed in the cerebral and pedal ganglia as well as the foot. Faint staining is also visible in the region of the nascent captacula. pv pavilion. Scale bars 50 µm
Ino-Gsx expression in Idiosepius notoides
In stage 19–20 individuals, Ino-Gsx is expressed in the region of the optic and palliovisceral ganglia (Figs. 8g, h, 12a–c). The cerebral ganglia, which are located dorsally to the mouth and expand anteroventrally in direction of the eyes, also express Ino-Gsx (Fig. 8h; arrowheads in Fig. 12b). In subsequent developmental stages, the expression domains remain the same and stage 23 individuals express Ino-Gsx in the optic and palliovisceral ganglia (Fig. 12d). The expression domain in the cerebral ganglia is relatively smaller compared to the domain reported for previous stages, and it is restricted to two patches ventro-laterally to the eye and close to the forming buccal mass (double arrowheads in Fig. 12d). In subsequent developmental stages, individual lobes of the supraesophageal mass as well as the posterior subesophageal mass and the optic lobes express Ino-Gsx (Fig. 13a–e). In the supraesophageal mass of stage 25 individuals, Ino-Gsx expression occurs in the inferior frontal and precommissural lobes as well as in the anterior basal and posterior basal lobes including the dorsal basal and dorsolateral lobes (Figs. 8i, j, 13a–c). In addition, Ino-Gsx is still expressed around the eyes and laterally of the buccal mass. This area might correspond to the region where the inferior buccal lobes develop (Figs. 8i, j, 13a, b). No expression was observed in the vertical, subvertical, and the superior frontal lobes or the anterior or middle subesophageal masses (Figs. 8i, j, 13a–c). Compared to stage 25 individuals, stage 26 individuals strongly express Ino-Gsx in their optic lobes (Figs. 8i, j, 13a–c). In addition, lobes of the supraesophageal mass such as the peduncle lobes or the buccal lobes express Ino-Gsx (Figs. 8i, j, 13a–c). Stronger Ino-Gsx expression is also observed laterally of the buccal mass, most likely corresponding to the inferior buccal lobes (Fig. 13e). Subsequent developmental stages until hatching only express Ino-Gsx around the eyes but not in the CNS (Figs. 8k, l, 13f, g).

Expression of Ino-Gsx during development of the cephalopod Idiosepius notoides. Dorsal (d)–ventral (v), anterior (a)–posterior (p), and left (l)–right (r) axes indicate the orientation. a Gsx is expressed in the region of the cerebral (cg) and palliovisceral ganglia (pvg) of stage 19 individuals. Inserted image depicts other optical section highlighting the mantle (m). b In addition to the optic ganglia, Ino-Gsx is expressed in the cerebral ganglia that expand anteroventrally in direction of the eyes of stage 19 individuals (arrowheads). c Optical section through the middle of the specimen shown in a and b which depicts the connection of the optic and palliovisceral ganglia. d Stage 22 individuals express Ino-Gsx in two domains of the cerebral ganglia, among others in the region of the future buccal lobes (double arrowheads). Note the unspecific signal in the intestines of this specimen (asterisk). Note unspecific staining (purple) in yolk sac (removed in d). ar arm, ey eye, mo mouth, o optic ganglion/lobe, y yolk. Scale bars 150 µm

Expression of Ino-Gsx during late development of the cephalopod Idiosepius notoides. Dorsal (d)–ventral (v), anterior (a)–posterior (p), and left (l)–right (r) axes indicate the orientation. Yolk sac removed in a–e. a Cephalic region of a stage 25 individual which expresses Ino-Gsx widely in the supraesophageal mass including the inferior frontal (if), anterior basal (ab), dorsal basal (db), dorsolateral (dl), and precommissural lobes (pc). Ino-Gsx expression is also observed around the eyes (arrowheads). b Same specimen as seen in a with Ino-Gsx expression around the eye (arrowhead), with faint expression on the lateral sides of the buccal region (arrow) and in the optic lobes (double-arrowhead). Note the lack of Ino-Gsx expression in the middle subesophageal mass (ms). c Same specimen as seen in a and b with Ino-Gsx expression in the posterior subesophageal mass (ps) but not in the middle subesophageal mass. Note the lack of Ino-Gsx expression in the vertical lobe (arrow) that is the anterior-most brain region. d Ino-Gsx is strongly expressed in the supraesophageal mass of stage 26 individuals including the peduncle lobes (pd) (here only cephalic region visible). Note the Ino-Gsx expression domain around the eyes (arrowheads). e Same specimen as seen in H with strong expression in the optic lobes and, among others, in the median basal lobes (mb). Note the strong Ino-Gsx expression on the lateral sides of the buccal mass (arrows). f Stage 28 individuals up to hatchlings express Ino-Gsx in the circular domain around the eyes (arrowhead); however, expression is lacking in the CNS. Note the unspecific staining in the statocysts. g Optical section along the sagittal plane of the same specimen as shown in j. No Ino-Gsx expression is visible in the supraesophageal (sp) or subesophageal masses (sb) of the CNS. Note the unspecific staining in the statocysts. ar arm, bm buccal mass, ey eye, fn funnel, mo mouth, o optic lobe, y yolk. Scale bars a–e 150 µm and f, g 200 µm
Discussion
Gsx does not pattern the digestive tract of scaphopods and cephalopods
To date, it is commonly hypothesized that the digestive tract of the last common bilaterian ancestor expressed Gsx in a collinear fashion together with the two other ParaHox genes, Cdx and Xlox [1, 6, 12, 20, 50]. This hypothesis is seemingly corroborated by the fact that among the Lophotrochozoa, the annelids Platynereis dumerilii and Nereis virens, as well as the gastropod Gibbula varia, express Gsx in their anterior digestive tract (Table 1; [12–14]). Our results for the scaphopod Antalis entalis and the cephalopod Idiosepius notoides, however, show that this is not the case for all mollusks, and therefore, neither for all lophotrochozoans, a scenario that was already suggested by data on the annelid Capitella teleta ([15]; Table 1). Moreover, all ecdysozoan representatives investigated lack Gsx expression in their digestive tract, and among the deuterostomes investigated, only the hemichordate Ptychodera flava expresses Gsx around the blastopore [7]. The lack of Gsx expression in the foregut of the other deuterostomes has been explained by the fate of the blastopore that does not transform into the definite mouth in deuterostomes as it does in protostomes, but, instead, into the anus [6]. Accordingly, the latter hypothesis would argue for Gsx expression in the deuterostome hindgut which, however, appears to be absent ([50]; Table 1). It is important to mention that Gsx orthologs have either not been found or are indeed absent in representatives of the Acoelomorpha, which are characterized by having a single mouth/anus opening in their digestive tract and may form the sister taxon to all remaining Bilateria (the so-called Nephrozoa; [51, 52]; but see [53] for a controversial view). In cnidarians, Gsx is endodermally expressed in the planula larva of Nematostella vectensis, Clytia hemisphaerica, and Podocoryne carnea [9–11]. In the coral Acropora millepora, Gsx is expressed in the ectoderm of the planula larva [8]. Comparisons of the cnidarian and nephrozoan expression domains are difficult since mouth and digestive system cannot be easily homologized. Hence, the data currently available argue for a last common nephrozoan and probably also bilaterian ancestor without Gsx expression in the digestive tract and for a recruitment of Gsx into foregut patterning in selected lineages. Accordingly, the gastropod G. varia and the polychaete annelids N. virens and P. dumerilii have acquired Gsx expression in the foregut secondarily during evolution (Table 1). In contrast, other genes such as Brachyury, Nkx2.1, or FoxA appear to be evolutionary highly conserved in the digestive system within the Lophotrochozoa [54–59].
Gsx is expressed in the anterior-most portion of the molluscan CNS
In contrast to the digestive tract, Gsx is consistently expressed in the anterior CNS of bilaterians and hence an ancestral role in CNS development was proposed (Table 1; [2]). Shared Gsx expression domains among mollusks are the cerebral ganglia that subsequently develop into the supraesophageal mass in cephalopods (present study; [12]). In scaphopod and gastropod larvae, the apical organ is located in the anterior-most region. In the scaphopod Antalis entalis, Gsx is expressed in two flask-shaped cells of this organ and in two cells that are located laterally to it but do not constitute a part of the apical organ (Fig. 4d). With two apical tuft cells and further putative sensory cells, the larva of the gastropod Gibbula varia possesses more Gsx-expressing cells in the apical organ than the one of A. entalis (present study; [12]). The flask-shaped Gsx-expressing cells of A. entalis do not appear to be homologous to any of the Gsx-expressing cell types of G. varia judging by their morphology. However, detailed ultrastructural studies and molecular fingerprints on the various cell types occurring in lophotrochozoan apical organs are necessary to further assess homologies in this organ on the cellular level. Among all metazoans with an apical organ (Cnidaria, Ambulacraria, and Lophotrochozoa), only both above-mentioned mollusks and the annelid Platynereis dumerilii possess Gsx-expressing cells in the apical organ, suggesting that Gsx has been recruited into the patterning of this sensory organ in lophotrochozoans only (Table 1; present study; [12, 13]).
Gsx expression has also been reported for the polychaete annelids Nereis virens and Capitella teleta [14, 15]. As far as known, both species lack an apical organ as do cephalopods as direct developers (present study; [3, 15]). The vertical lobe as the anterior-most portion of the cephalopod CNS does not express Gsx (Figs. 9d, 10). This resembles the expression patterns of other homeobox genes such as Otx or the POU genes which are consistently expressed in the gastropod cerebral ganglia and large parts of the cephalopod cerebral ganglia/supraesophageal mass but not in the vertical lobe [44, 60, 61]. The vertical lobe is considered an evolutionary innovation of coleoid cephalopods, i.e., all cephalopods except the nautiluses as basal cephalopod offshoots [62]. As an evolutionary younger brain region confined to coleoid cephalopods, the vertical lobe also differentiates relatively late during ontogeny compared to other brain regions [63]. Hence, the vertical lobe probably evolved after Otx expression domains had already been established in the supraesophageal mass of coleoid cephalopods.
Gsx is expressed in the posterior portion of the molluscan CNS
Idiosepius notoides and Antalis entalis express Gsx in posterior portions of their CNS such as the scaphopod pedal ganglia and the cephalopod palliovisceral ganglia (the latter develop into the future posterior subesophageal mass). This is in contrast to the gastropod Gibbula varia and the annelid Capitella teleta, where Gsx expression is restricted to the anterior CNS [12, 15]. The scaphopod and cephalopod condition is, however, similar to the condition found in Platynereis dumerilii and certain vertebrates insofar that both mollusks and the polychaete express Gsx in more posterior regions of their nervous system. These domains comprise the scaphopod pedal ganglia, the cephalopod palliovisceral lobe/posterior subesophageal mass, the polychaete nerve cord, and the hindbrain of vertebrates [13, 24–27]. Interestingly, Gsx is also expressed in portions of the developing visual system of few representatives of all three bilaterian superphyla. The mollusks I. notoides and Nereis virens, the arthropods Drosophila melanogaster, as well as the teleost fish Oryzias latipes, express Gsx in portions of their visual system (Table 1; present study; [14, 17, 28]). Further studies on other bilaterian representatives are needed to assess if Gsx expression in the eyes and related brain regions may be an ancestral trait among nephrozoans or bilaterians.
Conclusions
This study suggests that Gsx expression in the foregut is not a molluscan plesiomorphy and together with already published data argues against Gsx expression in the foregut of the last common bilaterian ancestor. It is therefore most likely that Gsx has been independently recruited into the development of the foregut in some lophotrochozoan representatives. Gsx is consistently expressed in the developing anterior nervous system of bilaterians, which is probably an apomorphy of Bilateria. In contrast to other metazoan taxa, Gsx expression was only found in the larval apical organ in lophotrochozoans, indicating that Gsx expression in the apical organ may be a lophotrochozoan synapomorphy.
Abbreviations
- a:
-
Anterior
- ab:
-
Anterior basal lobe
- Aen :
-
Antalis entalis
- A. entalis :
-
Antalis entalis
- an:
-
Anus
- ar:
-
Arm
- at:
-
Apical tuft
- ao:
-
Apical organ
- BBA:
-
Benzyl benzoate
- bc:
-
Buccal cone
- Bfl:
-
Branchiostoma floridae
- Bla:
-
Branchiostoma lanceolatum
- bm:
-
Buccal mass
- bp:
-
Blastopore
- brc:
-
Brachial crown
- c.f.:
-
Confer
- cg:
-
Cerebral ganglia
- cp:
-
Captacula
- CNS:
-
Central nervous system
- d:
-
Dorsal
- db:
-
Dorsal basal lobe
- dl:
-
Dorsolateral lobe
- Dme:
-
Drosophila melanogaster
- ep:
-
Episphere
- ey:
-
Eye
- f:
-
Foot
- fn:
-
Funnel
- Gsx :
-
Genomic screened homeobox protein
- G. varia :
-
Gibbula varia
- Gva:
-
Gibbula varia
- hpf:
-
Hours after fertilization
- hp:
-
Hyposphere
- ib:
-
Inferior buccal lobe
- i.e.:
-
Id est
- if:
-
Inferior frontal lobe
- ind:
-
Intermediate neuroblasts defective
- Ino :
-
Idiosepius notoides
- I. notoides :
-
Idiosepius notoides
- l:
-
Left
- Lgi:
-
Lottia gigantea
- m:
-
Mantle
- ma:
-
Mantle apex
- mb:
-
Median basal lobe
- mc:
-
Mantle cavity
- mf:
-
Mantle fold
- mg:
-
Midgut gland
- MFSW:
-
Millipore-filtered seawater
- mo:
-
Mouth
- ms:
-
Middle subesophageal mass
- NCBI:
-
National Center for Biotechnology Information
- o:
-
Optic ganglion/lobe
- p:
-
Posterior
- P. dumerilii :
-
Platynereis dumerilii
- Pdu:
-
Platynereis dumerilii
- pb:
-
Posterior basal lobe
- pc:
-
Precommissural lobe
- pd:
-
Peduncle lobe
- pg:
-
Pedal ganglion
- Pmi:
-
Patiria miniata
- ps:
-
Posterior subesophageal mass
- pt:
-
Prototroch
- pv:
-
Pavilion
- pvg:
-
Palliovisceral ganglia
- r:
-
Right
- sb:
-
Subesophageal mass
- sh:
-
Shell
- shf:
-
Shell field
- sp:
-
Supraesophageal mass
- Tca:
-
Tribolium castaneum
- v:
-
Ventral
- y:
-
Yolk
References
Holland PWH. Evolution of homeobox genes. WIREs Dev Biol. 2013;2:31–45.
Duboule D, Dollé P. The structural and functional organization of the murine Hox gene family resembles that of Drosophila homeotic genes. EMBO J. 1989;8:1497–505.
Kulakova MA, Bakalenko N, Novikova E, Cook CE, Eliseeva E, Steinmetz PRH, Kostyuchenko RP, Dondua A, Arendt D, Akam M, Andreeva T. Hox gene expression in the larval development of the polychaetes Nereis virens and Platynereis dumerilii (Annelida, Lophotrochozoa). Dev Genes Evol. 2007;217:39–54.
Lee PN, Callaerts P, de Couet HG, Martindale MQ. Cephalopod Hox genes and the origin of morphological novelties. Nature. 2003;424:1061–5.
Samadi L, Steiner G. Involvement of Hox genes in shell morphogenesis in the encapsulated development of a top shell gastropod (Gibbula varia L.). Dev Genes Evol. 2009;219:523–30.
Holland PW. Beyond the Hox: how widespread is homeobox gene clustering? J Anat. 2001;199:13–24.
Ikuta T, Chen YC, Annunziata R, Ting HC, Tung CH, Koyanagi R, Tagawa K, Humphreys T, Fujiyama A, Saiga H, Satoh N, Yu IK, Arnone MI, Su YH. Identification of an intact ParaHox cluster with temporal collinearity but altered spatial collinearity in the hemichordate Ptychodera flava. BMC Evol Biol. 2013;13:129.
Hayward DC, Catmull J, Reece-Hoyes JS, Berghammer H, Dodd H, Hann SJ, Miller DJ, Ball EE. Gene structure and larval expression of cnox-2Am from the coral Acropora millepora. Dev Genes Evol. 2001;211:10–9.
Finnerty JR, Paulson D, Burton P, Pang K, Martindale MQ. Early evolution of a homeobox gene: the parahox gene Gsx in the Cnidaria and the Bilateria. Evol Dev. 2003;5:331–45.
Quiquand M, Yanze N, Schmich J, Schmid V, Galliot B, Piraino S. More constraint on ParaHox than Hox gene families in early metazoan evolution. Dev Biol. 2009;328:173–87.
Yanze N, Spring J, Schmidli C, Schmid V. Conservation of Hox/ParaHox-related genes in the early development of a cnidarian. Develop Biol. 2001;236:89–98.
Samadi L, Steiner G. Conservation of ParaHox genes’s function in patterning of the digestive tract of the marine gastropod Gibbula varia. BMC Dev Biol. 2010;10:74.
Hui JHL, Raible F, Korchagina N, Dray N, Samain S, Magdelenat G, Jubin C, Segurens B, Balavoine G, Arendt D, Ferrier DEK. Features of the ancestral bilaterian inferred from Platynereis dumerilii ParaHox genes. BMC Biol. 2009;7:43.
Kulakova MA, Cook CE, Andreeva TF. ParaHox gene expression in larval and postlarval development of the polychaete Nereis virens (Annelida, Lophotrochozoa). BMC Dev Biol. 2008;8:61.
Fröbius AC, Seaver EC. Capitella sp. I homeobrain-like, the first lophotrochozoan member of a novel paired-like homeobox gene family. Gene Expr Patt. 2006; 6:895–91.
Weiss JB, Von Ohlen T, Mellerick DM, Dressler G, Doe CQ, Scott MP. Dorsoventral patterning in the Drosophila central nervous system: the intermediate neuroblasts defective homeobox gene specifies intermediate column identity. Genes Dev. 1998;12:3591–602.
Urbach R, Technau GM. Segment polarity and DV patterning gene expression reveals segmental organization of the Drosophila brain. Development. 2003;130:3607–20.
Wheeler SR, Carrico ML, Wilson BA, Skeath JB. The Tribolium columnar genes reveal conservation and plasticity in neural precursor patterning along the embryonic dorsal-ventral axis. Genomes Dev Control. 2005;279:491–500.
Arnone MI, Rizzo F, Annunciata R, Cameron RA, Peterson KJ, Martínez P. Genetic organization and embryonic expression of the ParaHox genes in the sea urchin S. purpuratus: insights into the relationship between clustering and collinearity. Dev Biol. 2006;300:63–73.
Annunziata R, Martinez P, Arnone MI. Intact cluster and chordate-like expression of ParaHox genes in a sea star. BMC Biol. 2013;11:68.
Brooke NM, Garcia-Fernàndez J, Holland PWH. The ParaHox gene cluster is an evolutionary sister of the Hox gene cluster. Nature. 1998;392:920–2.
Osborne PW, Benoit G, Laudet V, Schubert M, Ferrier DEK. Differential regulation of ParaHox genes by retinoic acid in the invertebrate chordate amphioxus (Branchiostoma floridae). Dev Biol. 2009;327:252–62.
Hudson C, Lemaire P. Induction of anterior neural fates in the ascidian Ciona intestinalis. Mech Dev. 2001;100:189–203.
Hsieh-Li HM, Witte DP, Szucsik JC, Weinstein M, Li H, Potter SS. Gsh-2, a murine homeobox gene expressed in the developing brain. Mech Dev. 1995; 50:177–86.
Valerius MT, Li H, Stock JL, Weinstein M, Kaur S, Singh G, Potter SS. Gsh-1: a novel murine homeobox gene expressed in the central nervous system. Dev Dyn. 1995;203:337–51.
Szucsik JC, Witte DP, Li H, Pixley SK, Small KM, Potter SS. Altered forebrain and hindbrain development in mice mutant for the Gsh-2 homeobox gene. Dev Biol. 1997;191:230–42.
Cheesman SE, Eisen SE. Gsh1 demarcates hypothalamus and intermediate spinal cord in zebrafish. Gene Expr Patt. 2004;5:107–12.
Deschet K, Bourrat F, Chourrout D, Joly JS. Expression domains of the medaka (Oryzias latipes) Ol-Gsh 1 gene are reminiscent of those of clustered and orphan homeobox genes. Dev Genes Evol. 1998;208:235–44.
Illes CJ, Winterbottom E, Isaacs HV. Cloning and expression analysis of the anterior ParaHox genes, Gsh1 and Gsh2 from Xenopus tropicalis. Dev Dyn. 2009;238:194–203.
Scheltema AH. Aplacophora as progenetic aculiferans and the coelomate origin of mollusks as the sister taxon of Sipuncula. Biol Bull. 1993;184:57–78.
Kocot KM, Cannon JT, Todt C, Citarella MR, Kohn AB, Meyer A, Santos SR, Schander C, Moroz LL, Lieb B, Halanych KM. Phylogenomics reveals deep molluscan relationships. Nature. 2011;477:452–6.
Smith SA, Wilson NG, Goetz FE, Feehery C, Andrade SCS, Rouse GW, Giribet G, Dunn CW. Resolving the evolutionary relationships of molluscs with phylogenomic tools. Nature. 2011;480:364–9.
Wanninger A, Haszprunar G. Muscle development in Antalis entalis (Mollusca, Scaphopoda) and its significance for scaphopod relationships. J Morph. 2002;254:53–64.
Steiner G, Dreyer H. Molecular phylogeny of Scaphopoda (Mollusca) inferred from 18S rDNA sequences: support for a Scaphopoda-Cephalopoda clade. Zool Scripta. 2003;32:343–56.
Shigeno S, Sasaki T, Moritaki T, Kasugai T, Vecchione M, Agata K. Evolution of the cephalopod head complex by assembly of multiple molluscan body parts: evidence from Nautilus embryonic development. J Morph. 2008;269:1–17.
Van Dongen CAM, Geilenkirchen WLM. The development of Dentalium with special reference to the significance of the polar lobe. I, II, III. Division chronology and development of the cell pattern in Dentalium dentale (Scaphopoda). Proc Kongl Ned Akad v Wet Ser C. 1974;77:57–100.
Van Dongen CAM, Geilenkirchen WLM. The development of Dentalium with special reference to the significance of the polar lobe. IV. Division chronology and development of the cell pattern in Dentalium dentale after removal of the polar lobe at first cleavage. Proc Kongl Ned Akad v Wet Ser C. 1975;78:358–75.
Van Dongen CAM, Geilenkirchen WLM. The development of Dentalium with special reference to the significance of the polar lobe. V and VI. Differentiation of the cell pattern in lobeless embryos of Dentalium vulgare (da Costa) during late larval development. Proc Kongl Ned Akad v Wet Ser C. 1976;79:245–66.
Wanninger A, Haszprunar G. The expression of an engrailed protein during embryonic shell formation of the tusk-shell, Antalis entalis (Mollusca, Scaphopoda). Evol Dev. 2001;3:312–21.
Wanninger A, Haszprunar G. The development of the serotonin-like immunoreactive and FMRFamidergic nervous system in Antalis entalis (Mollusca, Scaphopoda). Zoomorph. 2003;122:77–85.
Yamamoto M. Normal embryonic stages of the pygmy cuttlefish, Idiosepius pygmaeus paradoxus Ortmann. Zool Sci. 1988;5:989–98.
Wanninger A, Wollesen T. Mollusca. In: Wanninger A, editor. Evolutionary developmental biology of invertebrates, vol. 2 Lophotrochozoa (Lophotrochozoa). Wien: Springer; 2015. pp. 103–53.
Wollesen T, Cummins SF, Degnan BM, Wanninger A. FMRFamide gene and peptide expression during central nervous system development of the cephalopod Idiosepius notoides. Evol Dev. 2010;12:113–30.
Wollesen T, McDougall C, Degnan BM, Wanninger A. POU genes are expressed during the formation of individual ganglia of the cephalopod central nervous system. EvoDevo. 2014;5:41.
Grabherr MG, Haas BJ, Yassour M, Levin JZ, Thompson DA, Amit I, Adiconis X, Fan L, Raychowdhury R, Zeng Q, Chen Z, Mauceli E, Hacohen N, Gnirke A, Rhind A, Rhind N, di Palma F, Birren BW, Nusbaum C, Lindblad-Toh K, Friedman N, Regev A. Full-length transcriptome assembly from RNA-seqdata without a reference genome. Nat Biotechnol. 2011;15:644–52.
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, Thompson JD, Gibson TJ, Higgins DG. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23:2947–8.
Larsson A. AliView: a fast and lightweight alignment viewer and editor for large data sets. Bioinformatics. 2014;30:3276–8.
Felsenstein J. PHYLIP—Phylogeny Inference Package (Version 3.2). Cladistics. 1989;5:164–6.
Jékely G, Arendt D. Confocal detection of NBT/BCIP in situ hybridization samples by reflection microscopy. Biochemica. 2007;4:12–4.
Grapin-Botton A. Antero-posterior patterning of the vertebrate digestive tract: 40 years after Nicole Le Douarin´s Ph.D. thesis. Int J Dev Biol. 2005;49:335–47.
Hejnol A, Obst M, Stamatakis A, Ott M, Rouse GW, Edgecombe GD, Martinez P, Baguna J, Bailly X, Jondelius U, Wiens M, Müller WE, Seaver E, Wheeler WC, Martindale MQ, Giribet G, Dunn CW. Assessing the root of bilaterians animals with scalable phylogenomic methods. Proc Roy Soc B Biol Sci. 2009;276:4261–70.
Moreno E, Permanyer J, Martinez P. The origin of patterning systems in Bilateria—insights from the Hox and ParaHox genes in Acoelomorpha. Genomics Proteomics Bioinform. 2011;9:65–76.
Philippe H, Brinkmann H, Copley R, Moroz LL, Nakano H, Poustka AJ, Wallberg A, Peterson KJ, Telford MJ. Acoelomorph flatworms are deuterostomes related to Xenoturbella. Nature. 2011;470:255–60.
Arendt D, Technau U, Wittbrodt J. Evolution of the bilaterian larval foregut. Nature. 2001;409:81–5.
Boyle MJ, Seaver EC. Developmental expression of FoxA and GATA genes during gut formation in the polychaete annelid. Capitella sp I Evol Dev. 2008;10:89–105.
Boyle MJ, Seaver EC. Expression of FoxA and GATA transcription factors correlates with regionalized gut development in two lophotrochozoan marine worms: Chaetopterus (Annelida) and Themiste lageniformis (Sipuncula). EvoDevo. 2010;1:2.
Boyle MJ, Yamaguchi E, Seaver EC. Molecular conservation of metazoan gut formation: evidence from expression of endomesoderm genes in Capitella teleta (Annelida). EvoDevo. 2014;5:39.
Martin-Duran JM, Amaya E, Romero R. Germ layer specification and axial patterning in the embryonic development of the freshwater planarian Schmidtea polychroa. Dev Biol. 2010;340:145–58.
Martin-Duran JM, Romero R. Evolutionary implications of morphogenesis and molecular patterning of the blind gut in the planarian Schmidtea polychroa. Dev Biol. 2011;352:164–76.
O’Brien EK, Degnan BM. Pleiotropic developmental expression of HasPOU-III, a class III POU gene, in the gastropod Haliotis asinina. Mech Dev. 2002;114:129–32.
Buresi A, Baratte S, Da Silva C, Bonnaud L. Orthodenticle/otx ortholog expression in the anterior brain and eyes of Sepia officinalis (Mollusca, Cephalopoda). Gene Expr Patt. 2012;12:109–16.
Nixon M, Young JZ. The Brains and Lives of Cephalopods. New York: Oxford University Press; 2003.
Shigeno S, Tsuchiya K, Segawa S. Embryonic and paralarval development of the central nervous system of the loliginid squid Sepioteuthis lessoniana. J Comp Neurol. 2001;437:449–75.
Authors’ contributions
TW designed the project together with AW. TW reared and fixed all developmental stages of Idiosepius notoides and Antalis entalis, extracted the RNA, and assembled the transcriptomes. TW cloned all genes, and TW and SVRM carried out the in situ hybridization experiments. CMD performed the phylogenetic analysis. TW analyzed all data and drafted the manuscript. AW contributed to data interpretation and writing of the manuscript. BMD commented on a later version of the manuscript. All authors read and approved the final version of the manuscript.
Acknowledgements
The staff from the research vessel Neomys (SBR in Roscoff) is thanked for collecting adult scaphopods. Thomas Eder and Thomas Rattei (Vienna) are thanked for advice with transcriptome assembly. Julia Bauder is thanked for help with semithin sectioning. We kindly thank three anonymous reviewers for their constructive criticism. During his stay at the SBR in Roscoff (France), TW was generously supported by the Faculty of Life Science, University of Vienna, and an ASSEMBLE (Association of European Marine Biological Laboratories) Grant. This work was also supported by Grants to BMD of the Australian Research Council. Research in the lab of AW also benefitted from a Grant of the Austrian Science Fund (FWF) on comparative aspects of molluscan EvoDevo to AW (Grant number: P24276-B22).
Competing interests
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Wollesen, T., Rodríguez Monje, S.V., McDougall, C. et al. The ParaHox gene Gsx patterns the apical organ and central nervous system but not the foregut in scaphopod and cephalopod mollusks. EvoDevo 6, 41 (2015). https://doi.org/10.1186/s13227-015-0037-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13227-015-0037-z