
Abstract
Background
Sjögren’s syndrome (SjS) monocytes have a pro-inflammatory phenotype, which may influence SjS pathogenesis. MicroRNAs (miRNAs) are small endogenously expressed molecules that can inhibit protein expression of their targeted genes and have important functions in regulating cell signaling responses. We profiled miRNAs in SjS monocytes to identify a SjS-specific miRNA profile and determine the potential roles of miRNAs in SjS pathogenesis.
Methods
Total RNA was extracted from healthy control (HC, n = 10), SjS (n = 18), systemic lupus erythematosus (SLE, n = 10), and rheumatoid arthritis (RA, n = 10) peripheral blood CD14+ monocytes for miRNA microarray analysis. To validate select miRNAs from the microarray analysis, the original cohort and a new cohort of monocyte RNA samples from HC (n = 9), SjS (n = 12), SLE (n = 8), and RA (n = 9) patients were evaluated by quantitative reverse transcription (RT)-PCR. Functional predictions of differentially expressed miRNAs were determined through miRNA target prediction database analyses. Statistical analyses performed included one-way analysis of variance with Bonferroni post tests, linear regression, and receiver operating characteristic curve analyses.
Results
MiRNAs were predominantly upregulated in SjS monocytes in comparison with controls. Quantitative RT-PCR confirmations supported co-regulation of miR-34b-3p, miR-4701-5p, miR-609, miR-300, miR-3162-3p, and miR-877-3p in SjS monocytes (13/30, 43.3 %) in comparison with SLE (1/17, 5.8 %) and RA (1/18, 5.6 %). MiRNA-target pathway predictions identified SjS-associated miRNAs appear to preferentially target the canonical TGFβ signaling pathway as opposed to pro-inflammatory interleukin-12 and Toll-like receptor/NFkB pathways.
Conclusions
Our results underscore a novel underlying molecular mechanism where SjS-associated miRNAs may collectively suppress TGFβ signaling as opposed to pro-inflammatory interleukin-12 and Toll-like receptor/NFκB pathways in SjS pathogenesis.
Similar content being viewed by others


Background
Sjögren’s syndrome (SjS) is a chronic autoimmune disorder that affects the exocrine glands and multiple organ systems. The pathobiology of SjS involves the activation of the innate immune system and development of autoimmunity to exocrine tissues, resulting in severe dryness. In general, SjS can be divided into primary or secondary SjS (pSjS or sSjS) depending on co-morbid rheumatologic or autoimmune diseases [1]. In addition, although SjS is the second most common autoimmune condition to be diagnosed next to rheumatoid arthritis (RA), SjS diagnosis usually is difficult due to multiple symptoms resembling other conditions and a lack of disease-specific diagnostic markers [2, 3]. Devastating consequences of SjS have prompted research endeavors to expedite SjS diagnosis and guide SjS intervention strategies.
The precise mechanisms and sequence of events leading to salivary and ocular dysfunction are still unclear. Inflammatory responses in autoimmune conditions are assumed to be limited based on the tissue microenvironment, where tissue homeostatic processes and regulation of inflammation are critical in curtailing chronic inflammation. The innate immune cells involved in these processes, such as macrophages, dendritic cells (DCs), and epithelial cells, have been shown to shape lymphocyte proliferation and functions during adaptive immune regulation [4]. Monocytes and their derivatives of macrophages and DCs have abnormalities in autoimmune diseases such as systemic lupus erythematosus (SLE), RA, and SjS [5]. Monocytes represent a relatively homogenous population of important precursor cells to peripheral macrophages and DCs. Monocytes have innate immune functions including immunomodulatory functions, cytokine production, and phagocytosis [6]. Among SjS patient cell subsets, monocytes secrete increased levels of pro-inflammatory cytokines such as interleukin(IL)-6 and B cell-activating factor (BAFF) upon stimulation [7], express type I interferon (IFN)-regulated genes [5, 8–10], reduced NFκB inhibitor (IκBα) [11], and show decreased function in phagocytosis of apoptotic cells [12]. Thus, monocytes reflect the inflammatory state in SjS patients [10] and mature monocytes are proposed to contribute to salivary gland inflammation in SjS [13].
MicroRNAs (miRNAs) are naturally occurring post-transcriptional regulators of their targeted genes and can directly inhibit protein translation by a variety of mechanisms including direct cleavage of messenger RNAs (mRNAs) and translational repression due to disruption of translation initiation or premature translation termination [14, 15]. MiRNAs can regulate a wide range of processes including pro-apoptotic pathways, sensitization of innate immune receptor signaling, and cytokine expression in a variety of autoimmune processes and disease [16, 17]. One of the most intriguing aspects is how miRNAs function to cooperatively modulate cell processes by limiting specific pathway components, and thus in recent years extensive research has been performed to characterize miRNAs and their regulation of immune responses and immune cell development. Ours and other studies reported miRNAs are associated with SjS salivary gland tissue inflammation [18–21] and are shown to be upregulated in SjS peripheral blood mononuclear cells [20–23] and in long-term cultured salivary gland-derived epithelial cells [21]. Due to the complexity of cell subsets and miRNA-mRNA interactions involved in autoimmunity, it is still not well understood how miRNA dysregulation contributes to autoimmune disease pathogenesis in human patients.
Considering possible roles of miRNAs as mediators of inflammation, we hypothesized profiling of miRNAs in SjS monocytes provides insight into functional implications of miRNAs in shaping the observed dysregularities. No miRNA studies have been conducted on SjS monocytes to date, let alone clarified implications of miRNAs in SjS pathogenesis. Therefore, the objective of this study was to profile miRNAs in SjS monocytes, focusing on identifying SjS-specific miRNAs and predicting their potential roles in SjS pathogenesis. Our study is the first to characterize the differentially expressed miRNAs in SjS in comparison to HC, SLE, and RA autoimmune conditions and identify a potential SjS-specific miRNA profile in SjS monocytes. Elucidation of the pathobiology of SjS is critical not only to guide intervention strategies, but also to expedite SjS diagnosis.
Methods
Patient and control participants
The University of Florida Institutional Review Board approved this study and written permission was obtained from all who participated. Peripheral blood CD14+ monocytes were isolated using CD14+ whole blood isolation microbeads (Miltenyi Biotec, Bergisch Gladbach, Germany) for miRNA microarray and validation experiments. Patients were diagnosed with SjS according to the 2002 modified European-American diagnostic criteria [1], pSjS (n = 21) and sSjS ( n = 9) patients were included. Patients with pSjS were recruited when no other rheumatic or underlying disease was identified and sSjS participants were patients diagnosed with rheumatic or additional autoimmune disease. Autoimmune control participants diagnosed with SLE (n = 17) or RA (n = 18) alone were diagnosed based on the American College of Rheumatology criteria [24, 25]. Healthy donors with no history of autoimmune diseases were included as healthy controls (HC, n = 17). Detailed patients’ demographic, clinical and laboratory characteristics are summarized in Additional file 1.
RNA extraction and sample preparation
Total RNA including small RNAs from frozen CD14+ peripheral blood monocytes were extracted with mirVana miRNA Isolation kit (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. All RNA sample preparations’ RNA yield and A260/280 ratios were quantified using a NanoDrop1000 spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA) and RNA integrity was assessed by 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) prior to microarray profiling and stored at -80 °C until use.
MiRNA microarray analysis
Microarray miRNA expression profiling experiments were performed for each sample using one-color hybridizations on μParaflo™ microfluidic biochip miRNA microarray platform (LC Sciences, Houston, TX, USA) through LC Sciences’ miRNA microarray service. The array contained 2019 sequence-specific probes covering mature human miRNAs (hsa-) listed in Sanger miRBase Release v.19.0 and 44 Epstein-Barr virus miRNA-specific probes. Hybridized microarrays were scanned using an Axon GenePix 4000B Microarray Scanner and images digitized using Array-Pro Analyzer (MediaCybernetics, Rockville, MD, USA). Microarray data files were subjected background subtraction and to robust LOWESS (locally weighted regression) multi-array average background correction normalization. Data adjustment included signal significance analysis and data filtering that removed miRNAs with undetectable intensity values in more than half of control samples (Additional file 2). The log2-fold change values were calculated for each miRNA between any two participant groups. The threshold value used to define upregulation or downregulation of miRNA was a log2 ratio > ± 0.5 in expression in comparison to HC group and a value of P < 0.05 by one-way analysis of variance (ANOVA) with Bonferroni post tests (log2-transformed intensity values) among participant groups.
Verification of miRNA expression by quantitative reverse transcription (qRT)-PCR
Based on highest ranked increase in expression for SjS in comparison to HC, RA, or SLE controls, six selected candidate miRNAs were verified by qRT-PCR to validate the microarray results and in a separate validation cohort (Additional file 3). MiRNA quantification was carried out applying two-step reaction using commercially available TaqMan miRNA assays (Applied Biosystems, Foster City, CA, USA) according to manufacturer’s protocols. Microarray control samples HC#3727 and #3839, RA#3933, and SLE#3205 were excluded from validation by RT-PCR due to insufficient sample volume. For relative quantification of miRNA expression levels, all samples were normalized against endogenous reference RNA RNU6b using the comparative delta delta cycle threshold (ΔΔCt) method (relative expression level (fold change) = 2-(∆Ctsample-∆Ctcontrol)). To minimize experimental variation, matched samples of disease and controls were analyzed on the same reaction plates.
Verification of gene expression by qRT-PCR
Based on miRNA target prediction three genes SMAD2, SMAD3, and SMAD4 were selected for further verification by qRT-PCR in HCs and pSjS monocytes. Gene quantification was carried out using random primers for reverse transcription (high-capacity cDNA reverse transcription kit; Applied Biosystems) and TaqMan gene assay PCR primers (Applied Biosystems) according to manufacturer’s protocols. For relative quantification of gene expression, all samples were normalized against endogenous reference 18S ribosomal RNA using the comparative ΔΔCt method. Values were log2-transformed to display uniform scale for up- and downregulated genes.
Data analysis and statistical methods
Sample size determinations were performed prior to microarray analyses using software PS:Power and Sample Size Calculation version 3.1.2 [26]. With 80 % power to detect a mean effect of 0.50 with two-sided adjusted type I error probability α = 0.008333 and assuming within-group standard deviation estimate of 0.30 [26], required a sample size of ten subjects per group corresponding to HC, SjS, SLE, and RA. One-way ANOVA followed by Bonferroni post tests were performed as two-sided and adjusted P < 0.05 was considered significant (GraphPad Prism v.5 software, GraphPad Software, Inc, La Jolla, CA, USA). Receiver operator characteristics (ROC) curve and binary logistic regression analyses were used to determine the potential miRNA expression value cutoff to discriminate between HC and SjS patients (Additional file 4). Clustering analysis of miRNA microarray signal values was performed using the unsupervised hierarchical clustering method with the Cluster 3.0 program [27] and a clustering plot generated using TreeView software v. 1.6 [28]. MiRNA target prediction was analyzed through use of a variety of databases including validated miRNA targets (DIANA-TarBase v.7.0 [29] and miRSystem program [30]) and precomputed predicted miRNA targets (microRNA.org 2010 release (miRANDA) [31], TargetScan 6.2, RNA22 (miRANDA) [32]), summarized in Additional file 5. Potential biological functions for selected miRNAs were identified by miRSystem program [30]. Hsa-miR-588 has identical seed sequence (position 1–8) with miR-4701-5p and was utilized when database information was not available. Kyoto Encyclopedia of Genes and Genomes (KEGG) program (www.genome.jp/kegg/) was used to map predictions of individual mRNA-miRNA interactions. In addition, previously predicted and validated alternative polyadenylation sites in genes were indicated using NCBI AceView database for human genes [33] and RNAhybrid program (bibiserv.techfak.uni-bielefeld.de/rnahybrid/) was utilized to evaluate predicted miRNA and target gene interactions.
Results
MiRNAs are differentially upregulated in SjS monocytes
Microarray expression analyses were performed with HC, SjS, SLE, and RA monocyte samples to identify differentially expressed miRNAs among groups (Table 1).
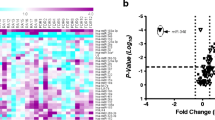
In general, SjS samples showed the highest level of miRNA upregulation, followed by SLE samples with an intermediate level of miRNA expression. Additionally, HC and RA samples showed lower expression levels for these miRNAs. Unsupervised hierarchical clustering (Fig. 1) of the microarray data comparing HC, SjS, SLE and RA CD14+ monocytes showed p SjS (8/12, 66.7 %), s SjS (3/6, 50 %), and SLE (7/10, 70 %) samples formed a cluster. Otherwise, most HC (7/10, 70 %) and RA (8/10, 80 %) samples were grouped within a second cluster.

MiRNA expression signature in CD14+ monocytes. Microarray heat map showing unsupervised hierarchical clustering analysis of differentially expressed miRNAs (one-way ANOVA with Bonferroni post tests, P < 0.05) for SjS (pSjS n = 12; sSjS n = 6), HC (n = 10), SLE (n = 10), and RA (n = 10) samples. Green, low expression levels; red, high expression levels. HC healthy control, pSjS primary Sjögren’s syndrome, RA rheumatoid arthritis, SLE systemic lupus erythematosus, SjS Sjögren’s syndrome, sSjS secondary Sjögren’s syndrome
We next used qRT-PCR to verify our microarray findings for selected miRNAs. These six miRNAs included miR-34b-3p, miR-300, miR-609, miR-877-3p, miR-3162-3p, and miR-4701-5p. The expression levels of these miRNAs from the combined cohort of samples are presented in Fig. 2. Of note, miR-34b-3p displayed a significant differential expression between SjS and HC or RA from the combined cohort (Fig. 2a). In addition, miR-4701-5p, and miR-3162-3p were significantly upregulated in SjS patient samples with comparison to RA (Fig. 2b,e). Although miR-609 and miR-300 tended to be upregulated in SjS compared with other control groups (Fig. 2c,d), there were no significant differences between relative expressions. Furthermore, ROC curve analyses were performed to determine cutoff values that maximized the specificity of discrimination between HC and SjS samples (Fig. 2, Additional file 4).

SjS-associated miRNAs are upregulated in CD14+ monocytes. Expression verification by qRT-PCR for combined microarray and validation cohorts including HC (circle, n = 17), SjS (pSjS closed square, n = 21; sSjS open square, n = 9), SLE (triangle, n = 17), and RA (diamond, n = 18) samples. Horizontal lines and bars represent mean ± SEM. Dotted line indicates cutoff value determined by ROC curve analyses (Additional file 4: Figure S2). True positive rate (TPR, sensitivity) = true positive/(true positive + false negative), true negative rate (TNR, specificity) = true negative/(true negative + false positive). * P < 0.05, *** P < 0.001, **** P < 0.0001 by one-way ANOVA with Bonferroni post tests. HC healthy control, pSjS primary Sjögren’s syndrome, RA rheumatoid arthritis, SLE systemic lupus erythematosus, SjS Sjögren’s syndrome, sSjS secondary Sjögren’s syndrome
SjS-associated monocyte miRNAs are co-expressed
MiRNAs are unlikely to be unique to a particular disease process and co-regulation may indicate similar functions. Therefore, we pursued characterization of their respective co-expression to determine whether a predictive panel could be identified. Utilizing cutoff values reported in Fig. 2 for our six selected miRNA markers, we evaluated the number of upregulated miRNAs in our combined cohort (Fig. 3a). Of note, 43.3 % of SjS patients (13/30) had upregulated expression of at least two of the six SjS-associated miRNAs. In contrast, 5.8 % SLE (1/17) and 5.6 % RA (1/18) had at least two upregulated SjS-associated miRNAs (Fig. 3a).

SjS-associated miRNA expression levels are positively associated in CD14+ monocytes. a Number and percent distribution of upregulated miRNAs in SjS (n = 30), SLE (n = 17), and RA (n = 18) patients. b Linear regression analyses were used to define associations between miRNA expression levels in SjS patients (pSjS closed square, n = 21; sSjS open square, n = 9). Cutoff values determined by ROC curve analyses are indicated by dotted box. P < 0.05 was considered statistically significant. HC healthy control, miRNA microRNA, RA rheumatoid arthritis, SLE systemic lupus erythematosus, SjS Sjögren’s syndrome
Since differentially expressed miRNAs tended to be co-increased in SjS monocytes, we tested whether any pairs of miRNAs displayed significant relationships in their expression levels. Linear regression analyses indicated miR-34b-3p and miR-4701-5p; miR-609 and miR-300; and miR-3162-3p and miR-877-3p expression levels were positively associated in SjS patients (Fig. 3b). Additionally, miR-609 and miR-300 expression values were positively associated in HC, SLE, and RA samples and miR-34b-3p and miR-4701-5p were also associated in RA patients (Additional file 6).
SjS-associated monocyte miRNAs preferentially target TGFβ signaling pathways
High-throughput miRNA target validation strategies are currently limited. Therefore, multiple miRNA target prediction algorithms and validated miRNA-target interaction databases were incorporated to identify potential functions of SjS-associated miRNAs in monocytes (summarized in Additional file 5). MiRNA target prediction was analyzed through databases for validated miRNA targets (direct evidence) and precomputed predicted miRNA targets (indirect evidence (weak) = 2 programs and indirect evidence (strong) = 3 or more programs). Our analyses further incorporated the KEGG program to map potential miRNA-regulated pathways. The canonical transforming growth factor beta (TGFβ) signaling pathway showed an average pathway coverage of 40.5 % ± 23.1 (SD) for the six SjS-associated miRNAs (Fig. 4a). Key factors such as TGFBR3 and SMAD2 involved in TGFβ signaling have direct evidence for targeting by miR-609 and miR-877-3p, respectively (Fig. 4b).

SjS-associated miRNAs are predicted to target canonical TGFβ signaling network. a Summary of average pathway-coverage scores for the six SjS-associated miRNAs for TGFβ, MAPK, JAK-STAT, and TLR-NFκB signal transduction pathways. Bars represent mean percent coverage ± SD for the six miRNAs. ** P < 0.01 by one-way ANOVA with Bonferroni post tests. MiRNA target predictions for TGFβ (b), MAPK (c), JAK-STAT (d), and TLR-NFκB (e) were analyzed through databases for validated miRNA targets (black, direct evidence) and precomputed predicted miRNA targets (light gray, indirect evidence (weak) = 2 programs and dark gray, indirect evidence (strong) = 3 or more programs). f Visualization of predicted miRNA targeting of canonical TGFβ signal transduction pathway
Additionally, MAPK pathways, which are also activated through TGFβ signaling receptors [34], also tended to show potential regulation (27.4 % ± 16.4 pathway coverage) by SjS-associated miRNAs (Fig. 4a). MiR-34b-3p has direct evidence for targeting GRB2, P38/MAPK13, and MEKK1/MAP3K1 and miR-877-3p has direct evidence for targeting SOS1, NRAS, ERK1/MAPK1, RAC1, and HGK/MAP4K4 (Fig. 4c). Visualization of the MAPK signaling pathways indicated critical components of the classical MAPK, JNK, and p38 MAPK signaling pathways may be affected by SjS-associated miRNAs (Additional file 7A).
Furthermore, evaluation of JAK-STAT signaling cascades, which are important for cytokine signaling cascades, also showed potential regulation (20.0 % ± 12.2 pathway coverage) by SjS-associated miRNAs (Fig. 4a). Of note, miR-877-3p had direct evidence for targeting STAT6 (Fig. 4d), which codes for the IL-4/IL-13 pathway transcription factor. In contrast, none of the SjS-associated miRNAs were predicted to target IL-12 receptor (IL12RB1, IL12RB2), TYK2, or STAT4 (Fig. 4d), which are involved in the IL-12 signaling cascade (Additional file 7B). Furthermore, the pro-inflammatory TLR/NFκB cascades averaged 4.2 % ± 3.2 pathway coverage (Fig. 4a,e) and had no direct evidence for targeting by SjS-associated miRNAs.
To identify impact of SjS-associated miRNAs on TGFβ signaling molecule gene levels in CD14+ monocytes, relative gene expression of SMAD2, SMAD3, and SMAD4 were evaluated by qRT-PCR. SMAD2 and SMAD3 gene expression levels were significantly increased in pSjS monocytes compared with HCs, although SMAD4 gene expression tended to be reduced (Fig. 5a). Further sequence analyses of predicted miRNA target sites revealed multiple alternative polyadenylation sites upstream of the predicted miRNA binding sites in the SMAD2 and SMAD3 3′ untranslated region (UTR) [33]. In contrast, analyses of SMAD4 3′UTR indicated miR-300 could possibly interact at four sites (positions 745, 833, 1094, 1185 3’UTR) upstream of the first alternative polyadenylation site (position 1499 [33]) and miR-609 to interact downstream (position 2810) in the SMAD4 3’UTR (Fig. 5b). Regression analyses indicated significant associations for both miR-300 and miR-609 with reductions in SMAD4 gene expression in pSjS patient monocytes (Fig. 5c).

MiR-300 and miR-609 are associated with inhibition of SMAD4 gene expression. a Relative expression of SMAD2, SMAD3, and SMAD4 were evaluated by qRT-PCR for SjS (n = 12) and HC (n = 10) CD14+ monocytes. Data represents log2-transformed gene expression. ** P < 0.01, *** P < 0.001 by two-tailed t test. b Target prediction of SMAD4 3′UTR by miR-300 and miR-609 represents target-miRNA alignments with the highest predicted minimum free energy (MFE). Position of alternative polyadenylation sites are indicated in the SMAD4 3′UTR. c Linear regression analyses of miR-300 and miR-609 with SMAD4 gene expression. HC healthy control, pSjS primary Sjögren’s syndrome, SjS Sjögren’s syndrome
Discussion
Research on gene expression in SjS CD14+ monocytes has yielded valuable findings for the role of inflammatory genes in SjS [5]. However, identifying master genes to be targeted to reverse disease processes has been a challenge in the field of SjS. MiRNAs can modulate protein expression levels by inhibiting gene translation into protein. MiRNA expression dysregulation has been reported in a variety of autoimmune diseases, including SjS [18–20, 22, 23]. However, the vast majority of previous studies evaluated miRNA expression levels in mixed cell populations or tissues, which may confound the further evaluation of specific miRNA-mRNA target relationships. Therefore, our study utilized miRNA microarray analyses to screen for differentially expressed miRNAs in SjS CD14+ monocytes in comparison with HC, RA, and SLE. In addition, we selected six SjS-associated miRNAs to be validated by RT-PCR and performed intensive pathway analyses to identify potential target pathways of SjS-associated miRNAs in CD14+ monocytes.
In general, primary and secondary SjS patients were undifferentiated from each other and showed the highest level of miRNA expression, whereas SLE patients tended to have an intermediate level of expression in comparison to SjS patients. Unsupervised hierarchical clustering of these miRNAs showed SjS and SLE patients clustered together independent from HCs and RA patient miRNA profiles (Fig. 1). These findings indicate miRNA alterations in monocytes are more common in SjS regardless of co-morbid autoimmune conditions and their dysregulation may play a role in SjS and SLE. In addition, although the healthy control group tended to be much younger than autoimmune groups (Additional file 1), they did not differ significantly based on miRNA associations with age (Additional file 8), which suggests age is not likely to confound our results.
It is important to note the monocytes utilized in this study were freshly isolated from peripheral blood. Monocytes once activated will rapidly adhere and migrate into affected tissues. Therefore, it was unsurprising a variety of previously reported miRNAs associated with monocyte activation and differentiation into DCs and macrophages, including miR-155-5p, miR-146a-5p, miR-146b-5p, miR-21-5p, miR-22-3p, miR-221-3p, miR-222-3p, and miR-424-5p [35, 36] were not differentially expressed miRNAs between SjS and control groups in our study. Indicating upregulated SjS-associated miRNAs in monocytes are most likely independent of immune activation stimuli controlling monocyte maturation and differentiation.
Validation of six selected SjS-associated miRNAs showed miR-34b-3p was upregulated in SjS patient monocytes in comparison to HCs and RA patient monocytes (Fig. 2a). MiRNAs significantly upregulated in SjS patient monocytes in comparison to RA patients were miR-3162-3p and miR-4701-5p (Fig. 2b,e). Notably, miR-300 and miR-609 tended to be elevated in SjS samples, although two RA patients from the validation cohort expressed higher levels of miR-300 and miR-609 (Fig. 2c,d). Conceivably, this observed overlap in upregulated SjS-associated miRNAs for SLE and RA patients could be related to the observed co-aggregation and similarities in serologic profiles of SjS patients with SLE and RA [37, 38].
Because SjS-associated miRNAs were similarly upregulated in SjS patients, we hypothesized SjS-associated miRNAs may also be co-expressed in individuals. Interestingly, 43.3 % of SjS patients, in comparison to 5.8 % SLE and 5.6 % RA patients, had upregulated expression of at least two of the six SjS-associated miRNAs analyzed (Fig 3a). Furthermore, regression analyses indicated definite pairs of miRNAs showed positive associations in SjS patients (Fig. 3b). Therefore, our data suggests co-expression of SjS-associated miRNAs may be especially useful in distinguishing cases of SjS. In addition, since these SjS-associated miRNAs are not genetically linked (Additional file 5), co-expression and upregulation of these miRNAs in SjS may indicate shared functions of regulating molecular pathways in monocytes.
In general, co-expressed miRNAs can share functions either by co-targeting individual genes or by targeting different components of the same pathway. Our in-depth analyses of target prediction algorithms and verified miRNA-mRNA interactions were utilized to reconstruct potential pathways targeted by SjS-associated miRNAs. Interestingly, canonical TGFβ signaling pathway showed the greatest coverage of predicted targets, averaging 40.5 % ± 23.1 (SD) pathway coverage for the six SjS-associated miRNAs (Fig. 4a). Moreover, TGFβ receptors are well known to activate a variety of associated signal transduction cascades, including the canonical MAP kinase and p 38/JNK MAP kinase pathways [34]. Targeting of the MAPK pathways averaged 27.4 % ± 16.4 coverage for the six SjS-associated miRNAs tested (Fig. 4a). Previously, miR-34 family members [39, 40], miR-609 [40], and miR-300 [41] were independently predicted to target the TGFβ signaling pathway. Specifically, experimental data support miR-34b-3p directly targets cAMP response element-binding (CREB)-1 transcription factor [42] and bone morphogenetic protein (BMP)-2 [43]. In addition, miR-609 directly targeted the full-length gene transcript of TGFβ1 [44] and TGFBR3 gene transcript [29, 45]. MiR-300 overexpression inhibited the TGFβ-mediated epithelial-to-mesenchymal transition in tumor cells [46]. And miR-877-3p has been shown to directly target SMAD2 gene transcript [29, 47]. Furthermore, we observed considerable overlap in evidence for SjS-associated miRNAs to target ACVR2B, SMAD2, SMAD3, and SMAD4 (Fig. 4b).
Based on our target prediction analyses, SMAD2, SMAD3 and SMAD4 were selected for initial screening for changes in gene expression in SjS monocytes compared with HCs. Intriguingly, SMAD2 and SMAD3 gene expression indicated significant increases in SjS monocytes compared with HCs (Fig. 5a), which has not been previously reported. Both SMAD2 and SMAD3 3’UTRs contain multiple alternative polyadenylation sites upstream of predicted miRNA binding sites according to AceView gene database [33], which we speculate could result in failure in miRNA-directed inhibition of gene transcript levels. Alternatively, miRNAs can also regulate protein translation by mechanisms other than transcript degradation [15]. However, at this time our study is limited to RNA-based analyses and monocytes are collected analyses. Relationships between miRNAs and their post-transcriptional inhibition of target genes will be addressed as soon as sufficient human monocytes are collected for protein analyses.
Nevertheless, SMAD4 gene expression exhibited repression in a subpopulation of pSjS patients (Fig. 5a). The SMAD4 protein of the family of TGFβ signaling molecules acts as an essential common mediator for receptor-regulated SMADs to enter the nucleus. MiR-300 and miR-609, which showed the strongest co-association, are likely to target SMAD4 directly based on target prediction analyses (Fig. 4b). Four miR-300 putative interaction sites are upstream of the first alternative polyadenylation site and the best predicted interaction of miR-300 with the lowest minimum free energy (MFE) -15.1 kcal/mole begins at nucleotide position 833 of the SMAD4 3’UTR as is shown in Fig. 4b. The best predicted interaction for miR-609, MFE -28.6 kcal/mole, is relatively nearby downstream to miR-300 predicted target sites and begins at position 2810 of the SMAD4 3’UTR (Fig. 4b). Regression analyses showed SMAD4 gene expression is significantly associated with both miR-300 and miR-609 (Fig. 4c), indicating possible cooperative targeting of SMAD4. Although further studies to identify target protein levels are required to define specific targets and functions of SjS-associated miRNAs in monocytes, our results indicate SjS-associated miRNAs could cooperate to modulate gene expression.
Our extensive data analyses pinpointing the TGFβ signaling network prompted us to revisit potential roles of TGFβ signaling in SjS pathogenesis. Until now the involvement of TGFβ in SjS patients has been largely unappreciated due to previous inconsistencies in reported TGFβ protein expression levels in SjS patient lip biopsy tissues [48–52] and in circulation [53, 54]. Conditional knockout of TGFβ receptors in mice have been utilized to better identify roles of TGFβ signaling in autoimmunity. Interestingly, TGFβ receptor 1 (TGFβR1) conditional knockout in mice salivary glands developed glandular inflammation resembling SjS only in female mice [55]. Of interest to our current study, bone marrow-derived DC-specific deletion of TGFβ receptor 2 (TGFβR2) resulted in multi-organ inflammation and activation of autoreactive T and B cells [56]. TGFβR2-deficient DCs are more pro-inflammatory (increased stimulated TNF, IL-6, IL-12, IL-1β, and IFNγ gene expression [56, 57]) and also have impaired IL-4 polarization of M2 alternatively activated regulatory macrophages [58]. Altogether these studies signify intact TGFβ signal transduction is critical for maintaining a regulatory immune phenotype and reinforcing DC/macrophage control of autoimmune progression.
Our target prediction analyses also identified SjS-associated miRNAs were predicted to target members of the JAK-STAT signaling family, averaging 20.0 % ± 16.4 coverage of signaling molecules (Fig. 4a). The MAPK and JAK-STAT signaling pathways are well known to affect a variety of pro-inflammatory and immune-modulatory cascades beyond TGFβ family signaling. As such, we propose that SjS-associated miRNAs also function by limiting the availability of shared signaling molecules, which creates conditions for competition that would ultimately favor pathways with greater abundance of signaling molecules. Substrate competition is found in many types of biological processes and can influence dynamics and steady state concentrations of a pathway [59] as well as serve as a general signal integration strategy for networks where enzymes interact with multiple regulators and substrates [60].
Interestingly, pro-inflammatory IL-12 and IL-23 signaling pathway molecules IL12RB1, IL12RB2, IL23R, TYK2, and STAT4, were not predicted to be targeted by SjS-associated miRNAs (Fig. 4d). Previously, IL-12p40 secretion in SjS patient monocyte-derived DCs was increased and correlated with increased NFκB/RELB protein levels [61]. Likewise, our analyses of the NFκB signaling pathway showed relatively low level of pathway coverage (7.3 % ± 5.8) by SjS-associated miRNAs (Fig. 4a,e). Intriguingly, SjS patient monocytes are previously shown to have increased gene expression of NFκB transcription factors (NFKB1 and NFKB2) and display increased inflammatory responses to BAFF and IFNγ [7, 9, 61]. Notably, SjS patients’ monocytes also exhibit reduced levels of inhibitor IκBα, which is a central regulator of NFkB activation [11] and TGFβ signaling is critical for IκBα maintenance [62–64]. Therefore, pro-inflammatory IL-12 and NFκB signaling pathways are presumed to be well maintained, whereas SjS-associated miRNAs skew regulatory TGFβ signaling responses in SjS monocytes.
Conclusions
Our study identified for the first time a miRNA signature for SjS in CD14+ monocytes, which appears to preferentially target TGFβ signaling responses as opposed to pro-inflammatory signaling pathways in monocytes. The interplay of the opposing NFκB and TGFβ signaling pathways is essential for coordinated cellular responses and numerous studies have established a critical role for intact TGFβ signaling mechanisms in controlling autoimmunity. As such, our study provides critical insight into the observed increase in inflammatory responses in SjS patient monocytes, which may possibly be related to changes in the salivary gland as well. Furthermore, we propose a paradigm shift in favor of defective regulatory signaling as an underlying mechanism for the observed pro-inflammatory phenotype in SjS monocytes. Clinical associations with miRNAs will need to be evaluated from a larger data set. Further examination of predicted miRNA-targeted protein levels and signaling responses in monocytes and in SjS salivary gland cell subsets is required to define the role of these upregulated miRNAs in SjS pathogenesis.
Abbreviations
- ANOVA:
-
analysis of variance
- BAFF:
-
B cell-activating factor
- BMP-2:
-
bone morphogenetic protein 2
- CREB1:
-
cAMP response element binding-1
- DC:
-
dendritic cell
- HC:
-
healthy control
- Hsa:
-
Homo sapiens
- IFN:
-
interferon
- IL:
-
interleukin
- IκBα:
-
nuclear factor of kappa light polypeptide gene enhancer in B cells inhibitor, alpha
- JAK:
-
Janus kinase
- KEGG:
-
Kyoto Encyclopedia of Genes and Genomes
- LOWESS:
-
locally weighted scatterplot smoothing
- MAPK:
-
mitogen-activated protein kinase
- MFE:
-
minimum free energy
- miRNA:
-
microRNA
- mRNA:
-
messenger RNA
- NFκB:
-
nuclear factor kappa-light-chain-enhancer of activated B cells
- pSjS:
-
primary SjS
- RA:
-
rheumatoid arthritis
- ROC:
-
receiver operating characteristic
- RT:
-
reverse transcription
- SjS:
-
Sjögren’s syndrome
- SLE:
-
systemic lupus erythematosus
- SMAD:
-
Sma and Mad (mothers against decapentaplegic)
- sSjS:
-
secondary SjS
- STAT:
-
signal transducer and activator of transcription
- TGFβ:
-
transforming growth factor beta
- TGFβR1:
-
transforming growth factor beta receptor type 1
- TGFβR2:
-
transforming growth factor beta receptor type 2
- TLR:
-
Toll-like receptor
- UTR:
-
untranslated region
- ΔΔCt:
-
delta delta cycle threshold
References
Vitali C, Bombardieri S, Jonsson R, Moutsopoulos HM, Alexander EL, Carsons SE, et al. Classification criteria for Sjogren’s syndrome: a revised version of the European criteria proposed by the American-European Consensus Group. Ann Rheum Dis. 2002;61(6):554–8.
Kassan SS, Moutsopoulos HM. Clinical manifestations and early diagnosis of Sjogren syndrome. Arch Intern Med. 2004;164(12):1275–84.
Gomes Pde S, Juodzbalys G, Fernandes MH, Guobis Z. Diagnostic Approaches to Sjogren’s syndrome: a literature review and own clinical experience. J Oral Maxillofac Res. 2012;3(1):e3.
Sun M, He C, Cong Y, Liu Z. Regulatory immune cells in regulation of intestinal inflammatory response to microbiota. Mucosal Immunol. 2015;8(5):969–78.
Brkic Z, Olthof ED, Drexhage HA, Versnel MA. Monocyte gene expression signatures in rheumatic diseases: biomarkers for disease activity and tools for diagnosis and classification. Open Arthr J. 2010;3(1):13–7.
Yang J, Zhang L, Yu C, Yang XF, Wang H. Monocyte and macrophage differentiation: circulation inflammatory monocyte as biomarker for inflammatory diseases. Biomark Res. 2014;2(1):1.
Yoshimoto K, Tanaka M, Kojima M, Setoyama Y, Kameda H, Suzuki K, et al. Regulatory mechanisms for the production of BAFF and IL-6 are impaired in monocytes of patients of primary Sjogren’s syndrome. Arthritis Res Ther. 2011;13(5):R170.
Wildenberg ME, van Helden-Meeuwsen CG, van de Merwe JP, Drexhage HA, Versnel MA. Systemic increase in type I interferon activity in Sjogren’s syndrome: a putative role for plasmacytoid dendritic cells. Eur J Immunol. 2008;38(7):2024–33.
Brkic Z, Maria NI, van Helden-Meeuwsen CG, van de Merwe JP, van Daele PL, Dalm VA, et al. Prevalence of interferon type I signature in CD14 monocytes of patients with Sjogren’s syndrome and association with disease activity and BAFF gene expression. Ann Rheum Dis. 2013;72(5):728–35.
Maria NI, Brkic Z, Waris M, van Helden-Meeuwsen CG, Heezen K, van de Merwe JP, et al. MxA as a clinically applicable biomarker for identifying systemic interferon type I in primary Sjogren’s syndrome. Ann Rheum Dis. 2014;73(6):1052–9.
Lisi S, Sisto M, Lofrumento DD, D’Amore M. Altered IκBα expression promotes NF-κB activation in monocytes from primary Sjogren’s syndrome patients. Pathology (Phila). 2012;44(6):557–61.
Hauk V, Fraccaroli L, Grasso E, Eimon A, Ramhorst R, Hubscher O, et al. Monocytes from Sjogren’s syndrome patients display increased vasoactive intestinal peptide receptor 2 expression and impaired apoptotic cell phagocytosis. Clin Exp Immunol. 2014;177(3):662–70.
Wildenberg ME, Welzen-Coppens JM, van Helden-Meeuwsen CG, Bootsma H, Vissink A, van Rooijen N, et al. Increased frequency of CD16+ monocytes and the presence of activated dendritic cells in salivary glands in primary Sjogren syndrome. Ann Rheum Dis. 2009;68(3):420–6.
Valencia-Sanchez MA, Liu J, Hannon GJ, Parker R. Control of translation and mRNA degradation by miRNAs and siRNAs. Genes Dev. 2006;20(5):515–24.
Valinezhad Orang A, Safaralizadeh R, Kazemzadeh-Bavili M. Mechanisms of miRNA-mediated gene regulation from common downregulation to mRNA-specific upregulation. Int J Genomics. 2014;2014:970607.
Jansson MD, Lund AH. MicroRNA and cancer. Mol Oncol. 2012;6(6):590–610.
Qu Z, Li W, Fu B. MicroRNAs in autoimmune diseases. Biomed Res Int. 2014;2014:527895.
Alevizos I, Alexander S, Turner RJ, Illei GG. MicroRNA expression profiles as biomarkers of minor salivary gland inflammation and dysfunction in Sjogren’s syndrome. Arthritis Rheum. 2011;63(2):535–44.
Tandon M, Gallo A, Jang SI, Illei GG, Alevizos I. Deep sequencing of short RNAs reveals novel microRNAs in minor salivary glands of patients with Sjogren’s syndrome. Oral Dis. 2012;18(2):127–31.
Kapsogeorgou EK, Gourzi VC, Manoussakis MN, Moutsopoulos HM, Tzioufas AG. Cellular microRNAs (miRNAs) and Sjogren’s syndrome: candidate regulators of autoimmune response and autoantigen expression. J Autoimmun. 2011;37(2):129–35.
Gourzi VC, Kapsogeorgou EK, Kyriakidis NC, Tzioufas AG. Study of microRNAs (miRNAs) that are predicted to target the autoantigens Ro/SSA and La/SSB in primary Sjogren’s syndrome. Clin Exp Immunol. 2015;182(1):14–22.
Pauley KM, Stewart CM, Gauna AE, Dupre LC, Kuklani R, Chan AL, et al. Altered miR-146a expression in Sjogren’s syndrome and its functional role in innate immunity. Eur J Immunol. 2011;41(7):2029–39.
Peng L, Ma W, Yi F, Yang YJ, Lin W, Chen H, et al. MicroRNA profiling in Chinese patients with primary Sjogren syndrome reveals elevated miRNA-181a in peripheral blood mononuclear cells. J Rheumatol. 2014;41(11):2208–13.
Aletaha D, Neogi T, Silman AJ, Funovits J, Felson DT, Bingham 3rd CO, et al. Rheumatoid arthritis classification criteria: an American College of Rheumatology/European League Against Rheumatism collaborative initiative. Arthritis Rheum 2010. 2010;62(9):2569–81.
Hochberg MC. Updating the American College of Rheumatology revised criteria for the classification of systemic lupus erythematosus. Arthritis Rheum. 1997;40(9):1725.
Dupont WD, Plummer Jr WD. Power and sample size calculations. A review and computer program. Control Clin Trials. 1990;11(2):116–28.
de Hoon MJ, Imoto S, Nolan J, Miyano S. Open source clustering software. Bioinformatics. 2004;20(9):1453–4.
Page RD. TreeView: an application to display phylogenetic trees on personal computers. Comput Appl Biosci. 1996;12(4):357–8.
Vlachos IS, Paraskevopoulou MD, Karagkouni D, Georgakilas G, Vergoulis T, Kanellos I, et al. DIANA-TarBase v7.0: indexing more than half a million experimentally supported miRNA:mRNA interactions. Nucleic Acids Res. 2015;43(Database issue):D153–9.
Lu TP, Lee CY, Tsai MH, Chiu YC, Hsiao CK, Lai LC, et al. miRSystem: an integrated system for characterizing enriched functions and pathways of microRNA targets. PLoS ONE. 2012;7(8):e42390.
Betel D, Wilson M, Gabow A, Marks DS, Sander C. The microRNA.org resource: targets and expression. Nucleic Acids Res. 2008;36(Database issue):D149–53.
Miranda KC, Huynh T, Tay Y, Ang YS, Tam WL, Thomson AM, et al. A pattern-based method for the identification of microRNA binding sites and their corresponding heteroduplexes. Cell. 2006;126(6):1203–17.
Thierry-Mieg D, Thierry-Mieg J. AceView: a comprehensive cDNA-supported gene and transcripts annotation. Genome Biol. 2006;7 Suppl 1:S12.1–14.
Zhang YE. Non-Smad pathways in TGF-beta signaling. Cell Res. 2009;19(1):128–39.
Forrest AR, Kanamori-Katayama M, Tomaru Y, Lassmann T, Ninomiya N, Takahashi Y, et al. Induction of microRNAs, mir-155, mir-222, mir-424 and mir-503, promotes monocytic differentiation through combinatorial regulation. Leukemia. 2010;24(2):460–6.
Wang J, Xiang G, Mitchelson K, Zhou Y. Microarray profiling of monocytic differentiation reveals miRNA-mRNA intrinsic correlation. J Cell Biochem. 2011;112(9):2443–53.
Scheinfeld N. Sjogren syndrome and systemic lupus erythematosus are distinct conditions. Dermatol Online J. 2006;12(1):4.
He J, Ding Y, Feng M, Guo J, Sun X, Zhao J, et al. Characteristics of Sjogren’s syndrome in rheumatoid arthritis. Rheumatology. 2013;52(6):1084–9.
Zhao F, Wang Z, Lang H, Liu X, Zhang D, Wang X, et al. Dynamic expression of novel miRNA candidates and miRNA-34 family members in early- to mid-gestational fetal keratinocytes contributes to scarless wound healing by targeting the TGF-beta pathway. PLoS ONE. 2015;10(5):e0126087.
Lhakhang TW, Chaudhry MA. Interactome of radiation-induced microRNA-predicted target genes. Comp Funct Genomics. 2012;2012:569731.
Kumar S, Kumar A, Shah PP, Rai SN, Panguluri SK, Kakar SS. MicroRNA signature of cis-platin resistant vs. cis-platin sensitive ovarian cancer cell lines. J Ovarian Res. 2011;4(1):17.
Pigazzi M, Manara E, Baron E, Basso G. miR-34b targets cyclic AMP-responsive element binding protein in acute myeloid leukemia. Cancer Res. 2009;69(6):2471–8.
Fotinos A, Nagarajan N, Martins AS, Fritz DT, Garsetti D, Lee AT, et al. Bone morphogenetic protein-focused strategies to induce cytotoxicity in lung cancer cells. Anticancer Res. 2014;34(5):2095–104.
Das S, Kumar M, Negi V, Pattnaik B, Prakash YS, Agrawal A, et al. MicroRNA-326 regulates profibrotic functions of transforming growth factor-beta in pulmonary fibrosis. Am J Respir Cell Mol Biol. 2014;50(5):882–92.
Balakrishnan I, Yang X, Brown J, Ramakrishnan A, Torok-Storb B, Kabos P, et al. Genome-wide analysis of miRNA-mRNA interactions in marrow stromal cells. Stem Cells. 2014;32(3):662–73.
Haga CL, Phinney DG. MicroRNAs in the imprinted DLK1-DIO3 region repress the epithelial-to-mesenchymal transition by targeting the TWIST1 protein signaling network. J Biol Chem. 2012;287(51):42695–707.
Kameswaran V, Bramswig NC, McKenna LB, Penn M, Schug J, Hand NJ, et al. Epigenetic regulation of the DLK1-MEG3 microRNA cluster in human type 2 diabetic islets. Cell Metab. 2014;19(1):135–45.
Ogawa N, Dang H, Lazaridis K, McGuff HS, Aufdemorte TB, Talal N. Analysis of transforming growth factor beta and other cytokines in autoimmune exocrinopathy (Sjogren’s syndrome). J Interferon Cytokine Res. 1995;15(9):759–67.
Kizu Y, Sakurai H, Katagiri S, Shinozaki N, Ono M, Tsubota K, et al. Immunohistological analysis of tumour growth factor beta 1 expression in normal and inflamed salivary glands. J Clin Pathol. 1996;49(9):728–32.
Koski H, Konttinen YT, Gu XH, Hietanen J, Malmstrom M. Transforming growth factor beta 2 in labial salivary glands in Sjogren’s syndrome. Ann Rheum Dis. 1995;54(9):744–7.
Mason GI, Hamburger J, Bowman S, Matthews JB. Salivary gland expression of transforming growth factor beta isoforms in Sjogren’s syndrome and benign lymphoepithelial lesions. Mol Pathol. 2003;56(1):52–9.
Kawanami T, Sawaki T, Sakai T, Miki M, Iwao H, Nakajima A, et al. Skewed production of IL-6 and TGFbeta by cultured salivary gland epithelial cells from patients with Sjogren’s syndrome. PLoS ONE. 2012;7(10):e45689.
Perrier S, Serre AF, Dubost JJ, Beaujon G, Plazonnet MP, Albuisson E, et al. Increased serum levels of interleukin 10 in Sjogren’s syndrome; correlation with increased IgG1. J Rheumatol. 2000;27(4):935–9.
Eriksson P, Andersson C, Ekerfelt C, Ernerudh J, Skogh T. Relationship between serum levels of IL-18 and IgG1 in patients with primary Sjogren’s syndrome, rheumatoid arthritis and healthy controls. Clin Exp Immunol. 2004;137(3):617–20.
Nandula SR, Amarnath S, Molinolo A, Bandyopadhyay BC, Hall B, Goldsmith CM, et al. Female mice are more susceptible to developing inflammatory disorders due to impaired transforming growth factor beta signaling in salivary glands. Arthritis Rheum. 2007;56(6):1798–805.
Ramalingam R, Larmonier CB, Thurston RD, Midura-Kiela MT, Zheng SG, Ghishan FK, et al. Dendritic cell-specific disruption of TGF-beta receptor II leads to altered regulatory T cell phenotype and spontaneous multiorgan autoimmunity. J Immunol. 2012;189(8):3878–93.
Laouar Y, Town T, Jeng D, Tran E, Wan Y, Kuchroo VK, et al. TGF-beta signaling in dendritic cells is a prerequisite for the control of autoimmune encephalomyelitis. Proc Natl Acad Sci U S A. 2008;105(31):10865–70.
Gong D, Shi W, Yi SJ, Chen H, Groffen J, Heisterkamp N. TGFbeta signaling plays a critical role in promoting alternative macrophage activation. BMC Immunol. 2012;13:31.
Schauble S, Stavrum AK, Puntervoll P, Schuster S, Heiland I. Effect of substrate competition in kinetic models of metabolic networks. FEBS Lett. 2013;587(17):2818–24.
Kim Y, Andreu MJ, Lim B, Chung K, Terayama M, Jimenez G, et al. Gene regulation by MAPK substrate competition. Dev Cell. 2011;20(6):880–7.
Vogelsang P, Karlsen M, Brun JG, Jonsson R, Appel S. Altered phenotype and Stat1 expression in Toll-like receptor 7/8 stimulated monocyte-derived dendritic cells from patients with primary Sjogren’s syndrome. Arthritis Res Ther. 2014;16(4):R166.
Ghafoori P, Yoshimura T, Turpie B, Masli S. Increased IkappaB alpha expression is essential for the tolerogenic property of TGF-beta-exposed APCs. FASEB J. 2009;23(7):2226–34.
Cavin LG, Romieu-Mourez R, Panta GR, Sun J, Factor VM, Thorgeirsson SS, et al. Inhibition of CK2 activity by TGF-beta1 promotes IkappaB-alpha protein stabilization and apoptosis of immortalized hepatocytes. Hepatology. 2003;38(6):1540–51.
Arsura M, Wu M, Sonenshein GE. TGF beta 1 inhibits NF-kappa B/Rel activity inducing apoptosis of B cells: transcriptional activation of I kappa B alpha. Immunity. 1996;5(1):31–40.
Acknowledgements
This work was supported by National Institutes of Health/National Institute of Dental and Craniofacial Research grants DE019644 (SC) and T90DE21990 (AEGW), and National Institutes of Health/National Center for Advancing Translational Sciences, Clinical and Translational Science Award UL1 TR000064/TL1 TR000066 (AEGW). We would also like to thank Dr. Kaleb M. Pauley for contributing to the experimental conception and design of the work presented.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
AEGW contributed to the experimental design, acquisition of the data set, analysis and interpretation of the work presented, and drafting and revision of the manuscript. KC acquired the data from qRT-PCR and assisted drafting the manuscript. YL contributed to data interpretation and manuscript revision. ALC, WHR, and MRB made contributions to study design, and acquisition of clinical data. CMS contributed to the design of the study. SC made substantial contributions to the study’s conception and design, coordination, drafting and revision of the manuscript. All authors read and approved the final manuscript.
Additional files
Additional file 1: Table S1.
Demographic and clinical parameters from the microarray, validation, and combined cohorts. (DOCX 25 kb)
Additional file 2: Table S2.
MicroRNA microarray data. (XLSX 4872 kb)
Additional file 3: Figure S1.
Differential miRNA expression verification by qRT-PCR from microarray and an independent validation cohort for CD14+ monocyte samples. HC, circles; pSjS, closed squares; sSjS, open squares; SLE, triangle; RA, diamond. Dotted line indicates cutoff value established by ROC curve analyses. * P < 0.05 by one-way ANOVA with Bonferroni post tests. (PDF 64 kb)
Additional file 4: Figure S2.
SjS-associated miRNA ROC curve analyses summary from qRT-PCR data. ROC curve analyses were performed for indicated miRNAs comparing HC (n = 17) and primary SjS (n = 21) samples. P < 0.05 was considered statistically significant. Cutoff values were determined to maximize specificity. (PDF 118 kb)
Additional file 5: Table S3.
Database analysis for miRNA target prediction summary. †Databases contain direct miRNA-mRNA target interaction information. (DOC 32 kb)
Additional file 6: Figure S3.
SjS-associated miRNA expression levels are positively associated in CD14+ monocytes. Linear regression analyses were used to define associations between miRNAs in HC (circle, n = 17), SLE (triangle, n = 17), and RA (diamond, n = 18) patient groups. Cutoff values established by ROC curve analyses are indicated by dotted lines. P < 0.05 was considered statistically significant. (PDF 125 kb)
Additional file 7: Figure S4.
MAP-kinase and JAK-STAT signaling components are predicted targets of SjS-associated miRNAs. Visualization of predicted miRNA targeting of the MAPK (A) and JAK-STAT (B) signaling pathways. Specific mRNA-miRNA interaction results were based on direct evidence obtained from DIANA Tarbase V.6 database (underlined) and from multiple target prediction programs. (PDF 83 kb)
Additional file 8: Figure S5.
Analysis of association of age with miRNA expression level indicates no significant differences between HC, SjS, SLE, and RA groups. Based on linear regression analyses, the differences between slopes were not significant among HC, SjS, SLE, and RA groups for miR-34b-3p (P = 0.1974), miR-4701-5p (P = 0.3065), miR-609 (P = 0.511), miR-300 (P = 0.1904), miR-3162-3p (P = 0.4748), and miR-877-3p (0.9712). In addition, although HCs tended to be younger than autoimmune patients on average, they are within the age range of autoimmune patients. (PDF 68 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Williams, A.E.G., Choi, K., Chan, A.L. et al. Sjögren’s syndrome-associated microRNAs in CD14+ monocytes unveils targeted TGFβ signaling. Arthritis Res Ther 18, 95 (2016). https://doi.org/10.1186/s13075-016-0987-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13075-016-0987-0