
Abstract
Background
Adult mosquitoes of the genus Anopheles are important vectors of Plasmodium parasites, causative agents of malaria. The aim of this review was to synthesize the overall and species-specific proportion of Anopheles species infected with sporozoites and their geographical distribution in the last 2 decades (2001–2021).
Methods
A comprehensive search was conducted using databases (PubMed, Google Scholar, Science Direct, Scopus, African Journals OnLine) and manual Google search between January 1 and February 15, 2022. Original articles describing work conducted in Ethiopia, published in English and reporting infection status, were included in the review. All the required data were extracted using a standardized data extraction form, imported to SPSS-24, and analyzed accordingly. The quality of each original study was assessed using a quality assessment tool adapted from the Joanna Briggs Institute critical appraisal checklist. This study was registered on PROSPERO (International Prospective Register of Systematic Reviews; registration no. CRD42022299078).
Results
A search for published articles produced a total of 3086 articles, of which 34 met the inclusion criteria. Data on mosquito surveillance revealed that a total of 129,410 anophelines comprising 25 species were captured, of which 48,365 comprising 21 species were tested for sporozoites. Anopheles arabiensis was the dominant species followed by An. pharoensis and An. coustani complex. The overall proportion infected with sporozoites over 21 years was 0.87%. Individual proportions included Anopheles arabiensis (1.09), An. pharoensis (0.79), An. coustani complex (0.13), An. funestus (2.71), An. demeilloni (0.31), An. stephensi (0.70), and An. cinereus (0.73). Plasmodium falciparum sporozoites accounted 79.2% of Plasmodium species. Mixed infection of Plasmodium vivax and P. falciparum was only reported from one An. arabiensis sample.
Conclusions
Anopheles arebiensis was the dominant malaria vector over the years, with the highest sporozoite infection proportion of 2.85% and an average of 0.90% over the years. Other species contributing to malaria transmission in the area were An. pharoensis, An. coustani complex, An. funestus, An. stephensi, and An. coustani. The emergence of new vector species, in particular An. stephensi, is particularly concerning and should be investigated further.
Graphical Abstract

Similar content being viewed by others


Background
Malaria is one of the most serious public health problems worldwide. In 2020, there were 241 million malaria cases (5.0% higher than 2019) and 627,000 deaths (34.8% higher than 2019) in 85 malaria-endemic countries worldwide [1]. Plasmodium falciparum, P. vivax, P. ovale, P. malariae, and P. knowlesi are the five Plasmodium species that cause malaria. Of them, P. falciparum and P. vivax are predominant and responsible for frequent mortalities and morbidities [2]. The disease transmission is aggravated by poverty and climate change [3,4,5].
Adult mosquitoes of the genus Anopheles are important vectors of Plasmodium parasites. Globally, there are over 500 identified species of Anopheles mosquitoes and approximately 70–80 transmit malaria in humans [6, 7]. In Africa, there are 140 Anopheles species and only 20 are known malaria vectors [8]. Their long-life span and strong human-biting behavior explain why over 90% of the world’s malaria cases are recorded from WHO African region each year [2]. More than 40 species of Anopheles mosquitoes have been identified in Ethiopia [9,10,11]. Of them, Anopheles gambiae complex is the primary vector of malaria, while An. pharoensis, An. funestus, An. coustani complex, and An. nili are secondary vectors [12, 13]. Anopheles arabiensis is the major vector species of the An. gambiae complex [14].
Recently, An. stephensi (the Asian malaria vector) has emerged on the African continent as a potential vector of both P. falciparum and P. vivax. It was first detected in Djibouti (2012) [15], Ethiopia (2016) [16], Sudan (2019) [17], Somalia (2019) [18], Nigeria (2020) [18], and most recently Kenya (2022) [19]. Detection was made morphologically and/or by sequencing recombinant DNA (rDNA) internal transcribed spacer region 2 (ITS2) and mitochondrial DNA cytochrome c oxidase subunit 1 (cox1) [16, 20].
Assessment of infection status of the malaria vectors in their salivary glands is crucial for vector incrimination and to estimate the number of infectious mosquitoes carrying Plasmodium sporozoites, which with human biting rate is important to evaluate entomological inoculation rate (EIR). Plasmodium sporozoite proportions can be estimated by detecting sporozoites under a microscope [21], using Circumsporozoite-ELISA (CS-ELISA) [22] or polymerase chain reaction (PCR) [23,24,25] from mosquito samples. In earlier times, microscopy was used to detect sporozoites in salivary glands for calculation of the percentage/proportion of infectious mosquitoes. However, this method requires fresh samples, more time, and skilled microscopists to dissect many mosquitoes, and it also fails to discriminate among Plasmodium species [26, 27]. Though CS-ELISA is the routinely used and widely accepted technique to detect sporozoites from naturally infected Anopheles mosquitoes, studies reported that it lacks specificity and overestimates the sporozoite rate (real infection) by detecting circumsporozoite protein in the hemolymph [23] and from the developing oocysts in the abdomen [21, 28].
Ethiopian malaria entomological data from 1930 to 2000 reported that the sporozoite infection rate of An. arabiensis ranges from 0 to 5.43% and the maximum sporozoite rate of An. nili was 1.57% [29]. In the last 2 decades (2001–2021), several entomological studies have been conducted to determine the infection rate of Anopheles mosquitoes by detecting Plasmodium CSPs using ELISA or any other methods (PCR and microscopy). However, no record is available on the overall and species-specific infection proportion of Anopheles species in Ethiopia. The present review aimed to summarize the past 21 years (2001–2021) overall and species-specific sporozoite infection proportion and regional distribution of Anopheles mosquitoes.
Methods
Search design and strategy
This systematic review has employed the Preferred Reporting Items for Systematic Reviews and Meta-Analysis (PRISMA) guidelines (Additional file 1: Table S1) [30]. The review protocol was registered in PROSPERO (International Prospective Register of Systematic Reviews; registration no. CRD42022299078). To find potentially relevant articles, a comprehensive search was conducted using electronic databases (PubMed, Google Scholar, Science direct, Scopus, African Journals OnLine) and manual Google search between January 1 and February 15, 2022. The search used the following search terms (MeSH and Text Words): ‘Anopheles species,’ ‘Anopheles mosquito,’ ‘malaria vector,’ ‘sporozoite infection,’ and ‘Ethiopia.’ The search terms were used in combination using Boolean operators like “OR” or “AND.” The reference lists of included studies were screened for additional eligible studies. Search results from different databases were exported to EndNote-X8 and then combined after removing duplicates.
Study selection
Two researchers screened the selected articles for eligibility by reading the titles, abstracts, and then the full-text. Articles that did not report the outcome of interest were removed.
Eligibility criteria
Inclusion criteria
Original articles conducted in Ethiopia, published in English between January 2001 and December 2021, reporting on female Anopheles mosquitoes of any species tested for circumsporozoite protein and sporozoite infection proportions.
Exclusion criteria
Mosquitoes other than Anopheles (e.g. Culex and Aedes) were not considered. Eligibility of each article was initially assessed based on titles and abstracts. Relevant articles were further evaluated using full texts. Studies without full texts were excluded after attempting to contact the primary author at least twice via email with no response. Additionally, studies lacking reported sporozoite infection statuses were excluded after reviewing their full texts.
Data extraction
Two authors (YA and AG) extracted all the required data independently using a standardized data extraction form adapted from the JBI data extraction form for systematic reviews in a Microsoft Excel spreadsheet. Data were extracted from texts and tables of the original papers. Discrepancy between investigators on the data to be extracted was resolved via discussion or a third researcher. If still not resolved and additional clarification is required, the primary author of the included studies was contacted via email at least two times. Nominal and numerical data such as primary author, year of publication, study site, altitude of the study area, collection methods, number of Anopheles mosquito collected, Anopheles species identified, number of tested Anopheles mosquitoes, number of infected Anopheles mosquitoes, overall Plasmodium sporozoite rate, Plasmodium species detected, and methods used to detect sporozoite were extracted in Excel format.
Assessment of methodological quality
Methodological quality and possibility of bias were assessed before inclusion in the review by undertaking critical appraisal to incorporate into the review process. Two authors (YA and AG) independently assessed the quality of each article using a standard quality assessment tool adapted from the JBI critical appraisal checklist for studies reporting prevalence data (Additional file 2: Table S2) [31]. Disagreements between two assessors were resolved via discussion and by contacting the primary author of the included studies via email. The tool has nine quality items that focus on sample size, methods used for identification of the outcome, and statistical analysis. Articles with a score ≥ 70% were considered high quality. Articles with a score between 69 and 51% and ≤ 50% were considered as moderate and poor quality respectively. None of the 34 articles was poor quality. The cut-off point was determined after reading other literature.
Data processing and analysis
The data were extracted using Microsoft Excel 2010 spreadsheet and imported to SPSS version 24 software for analysis. The detailed description of the included original articles is presented on a table. Frequency and percentage were calculated to determine the overall and species-specific sporozoite infection proportion, frequency of occurrence, and to analyze regional distribution of anophelines. Mean and range were computed to describe the range and mean infection proportion of each Anopheles species over 21 years. The results were then presented in charts and tables.
Results
Literature search results
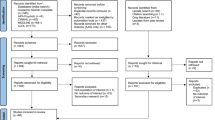
The search of international databases and manual Google provided a total of 3086 published studies. After the removal of duplicates, 2819 remained. Of these, 2776 studies were discarded after reading their titles and abstracts since they did not meet the inclusion criteria. Of the 43 studies assessed for eligibility, one study was discarded because of the inaccessibility of the full text at the time of review and the full text of 42 studies was assessed. Then, six studies were discarded since they did not report the outcome of interest, and two studies were discarded because of incomplete data. Finally, 34 studies were included in the review process (Fig. 1).

Flow chart of study selection for systematic review of sporozoite infection rate of Anopheles mosquitoes in Ethiopia, 2001–2021
Study characteristics
All experiments included in this review were conducted based on standardized procedures and guidelines for CSP or DNA detection [6, 22, 25, 32,33,34]. The studies were published between January 1, 2001, and December 31, 2021, and conducted in different (western, eastern, central, southern, and northern) parts of Ethiopia. Twenty-eight studies were longitudinal studies, four were cross-sectional, and two were randomized control trials (Additional file 3: Table S3) [20, 35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67].
Species diversity and frequency of Anopheles species
According to this review, 129,410 Anopheles mosquito specimens were captured from 2001 to 2021. A total of 25 Anopheles species have been recorded in different parts of Ethiopia, with an altitude ranging from 751 to 2660 m above sea level. They were collected from both indoor and outdoor sites. The collection methods employed were CDC light trap, human landing catch (HLC), pyrethrum spray catch (PSC), manual aspiration, artificial shelter, pitfall trap, clay pots, exit traps, human-baited double net traps, and cattle-baited traps (Additional file 3: Table S3). In all studies, sub-speciation using molecular testing has been performed on An. gambiae complex, and it has been confirmed to be An. arabiensis.
Anopheles arabiensis comprised the highest frequency (recorded in 32 out of 34 studies), followed by An. pharoensis (recorded in 25/34 studies), An. coustani complex (16/34), and An. funestus (13/34). Anopheles stephensi and An. cinereus have also been recorded in two (2/34) and one (1/34) study respectively (Fig. 2).

Frequency of Anopheles species in the four original studies from 2001 to 2021
Sporozoite infection proportions of the tested Anopheles mosquitoes
Of the 25 Anopheles species identified, 21 species had been tested for Plasmodium sporozoite or DNA. The overall Plasmodium sporozoite infection proportion ranged from 0.0 to 3.88%. A total of 48,365 (37.4%) Anopheles mosquito samples were tested for circumsporozoite protein or DNA among 129,410 Anopheles mosquitoes captured. Of these, CSP/DNA was detected in 423 (0.87%) samples using one of the three methods (CS-ELISA, PCR, or microscopy). Only seven Anopheles species (An. arabiensis, An. pharoensis, An. stephensi, An. coustani complex, An. funestus, An. cinereus, and An. demeilloni) were found positive for sporozoites. None of the Anopheles dthali, An. rhodiensis, An. longipalpis, and An. wellcomei was tested for Plasmodium sporozoite. Anopheles arabiensis, An. pharoensis, An. coustani, and An. funestus had sporozoite infection proportion of 1.09%, 0.79%, 0.13%, and 2.71% respectively. Though non-significant, there was a slight variation in sporozoite infection proportion of these three vectors over 21 years. Anopheles arabiensis varied between 0 and 2.85% (mean = 0.90%), An. pharoensis varied between 0 and 2.13% (mean = 0.42%), and An. coustani complex varied between 0 and 0.61% (mean = 0.18%) (Table 1).
Geographical distribution of tested Anopheles species
The majority of the tested anophelines were from central Ethiopia (39.2%), followed by southern Ethiopia (35.4%) and western Ethiopia (22.4%). The lowest numbers of anophelines were tested from eastern (1.8%) and northern (1.2%) Ethiopia. Anopheles arabiensis was recorded and tested in all parts of Ethiopia. Anopheles stephensi and An. cinereus were identified and reported as a vector of malaria in eastern and northern Ethiopia respectively.
Geographical distribution of sporozoite-infected Anopheles species
The highest numbers of sporozoite-infected Anopheles species were recorded in central Ethiopia (246/423), followed by southern Ethiopia (120/423). The fewest infected Anopheles species were recorded in northern Ethiopia (4/423). Infected An. arebiensis was recorded in all parts of Ethiopia except eastern Ethiopia. Sporozoite-infected An. cinereus, An. stephensi, and An. demeilloni were recorded in northern, eastern, and southern Ethiopia, respectively (Fig. 3).

Distribution of sporozoite-infected Anopheles species in different parts of Ethiopia from 2001 to 2021
Frequency of sporozoite detection methods and species detected
Overall, three detection methods (ELISA, PCR, and wet mount) were used in the 34 studies. Thirty studies (88.2%) used direct/sandwich ELISA, three studies (8.8%) used nested PCR, and one study (3.0%) used direct microscopy to detect Plasmodium sporozoites (Fig. 4).

Frequency of sporozoite detection methods and sporozoite species detected from 2001 to 2021. CS-ELISA, circumsporozoite enzyme-linked immunosorbent assay; PCR, polymerase chain reaction
Plasmodium falciparum, P. vivax (both vivax-210 and vivax-247 variants), and mixed sporozoites were detected from Anopheles samples tested positive. Either P. vivax or P. falciparum was isolated from examined anopheline samples in each study (Additional file 3: Table S3). Plasmodium falciparum sporozoites comprised the higher percentage (79.2%). Mixed infection of P. vivax and P. falciparum was only reported from one An. arabiensis sample (Fig. 4).
Discussion
Vector control is a very effective approach to combat malaria transmission. Hence, it is an essential element of malaria control and elimination plans. The spread of native vectors, such as An. stephensi, into new areas is currently a challenge for WHO malaria elimination programs. Information about competent vectors of malaria with their geographical distribution is vital to public health and helps to mitigate and monitor the malaria threat in a certain area. For improved management and control of malaria, an update on the entomological profile is important to identify focus areas where vector surveillance is limited.
This review presented the previous 21-year (2001–2021) overall and species-specific sporozoite positivity rate and regional distribution profile of Anopheles species across Ethiopia. Twenty-five Anopheles species were recorded in different parts of the country with an altitude ranging from 751 to 2660 m above sea level. A previous review aiming to assess Anopheles species composition, abundance, and distribution reported a record of 35 Anopheles species in different parts of Ethiopia [68]. The difference in composition of Anopheles species is that the current study aimed to assess sporozoite infection proportion and included only studies that were conducted to determine infection status and reported sporozoite proportions.
Of the 129,410 Anopheles mosquitoes captured, 48,365 (37.4%) anophelines comprising 21 species were tested for circumsporozoite protein or parasite DNA using CS-ELISA and PCR techniques respectively from 2001 to 2021. This might be due to lack of resources and/or the remaining samples might not be eligible (nulliparous) for the determination of infection status which can be done via examination of abdominal status. The present review confirmed that An. arabiensis, An. pharoensis, An. coustani complex, An. funestus, An. stephensi, An. cinereus, and An. demeilloni were important malaria vectors of both P. falciparum and P. vivax in the area. Even though An. nili was incriminated as a vector of malaria in 1970 [69] and has been considered a secondary malaria vector in the Gambella area, none of the tested An. nili was sporozoite positive over 21 years.
According to the current review, the overall proportion of infected Anopheles mosquitoes from 2001 to 2021 was 0.87% with a mean overall proportion of 0.73%. Only seven species were found positive among 21 species tested. The sporozoite infection proportion of An. arabiensis, An. pharoensis, and An. coustani complex over 21 years ranged from 0 to 2.85% (mean = 0.90%), 0 to 2.13% (mean = 0.42%), and 0–0.61% (mean = 0.18%) respectively. The maximum sporozoite infection proportion of An. arabiensis in the present review was slightly lower than in a previous Federal Ministry of Health (FMoH) report (5.43%) [29]. Anopheles funestus, An. cinereus, and An. demeilloni had a mean sporozoite infection proportion of 0.48% (range 0–3.84%), 0.31% (range 0–0.92%), and 0.09% (range 0–0.58%) respectivly over 21 years.
Most the tested anophelines were from central Ethiopia (39.2%), followed by southern Ethiopia (35.4%). The fewest anophelines were tested from eastern (1.8%) and northern (1.2%) Ethiopia. This finding is similar to a previous review report focused on the abundance and distribution of Anopheles mosquitoes, which reported that the most abundant Anopheles mosquitoes were recorded in central Ethiopia and the fewest in eastern Ethiopia [68]. Similarly, the highest numbers of sporozoite-infected anophelines were recorded in central Ethiopia (n = 246) followed by southern Ethiopia (n = 120) compared to other areas, and the fewest sporozoite-infected anophelines were recorded in northern (n = 4) and eastern (n = 6) Ethiopia. Although several entomological studies were conducted in northern and eastern Ethiopia, they were focused on species composition, seasonal abundance, distribution, longevity, parous rate, and insecticide resistance despite the high prevalence of plasmodial infections among the population in these regions [70,71,72,73,74].
As shown in Fig. 4, the majority (88.6%) of studies have estimated sporozoite rates using CS-ELISA despite several limitations of this method. False-positive results have been reported by CS-ELISA compared with cytochrome b PCR (Cytb-PCR) [24]. A high rate of false positives due to cross-reactivity with non-Plasmodium antigens (host blood) has also been observed [75]. In Senegal, false-positive results were reported in An. gambiae sensu lato with bovine and/or sheep blood meals tested for the presence of P. malariae and P. ovale circumsporozoite protein [76]. In Thailand, false-positive results for P. falciparum and P. vivax have been found when testing the plasma fractions of pig and bovine blood [77].
Hasan et al. [24] revealed that a novel PCR method targeting Plasmodium mitochondrial cytochrome b (Mt Cytb-PCR) is highly sensitive and reliable for identifying Plasmodium species from Anopheles mosquitoes. It can detect the sporozoites as efficiently as CS-ELISA and nested PCR and detects low parasite density. Hendershot et al. [78] also confirmed that PCR-based method targeting mitochondrial cytochrome C oxidase subunit I (Mt COX‑I PCR) is highly sensitive in detecting Plasmodium DNA in mosquitoes. However, it is not a good candidate for sporozoite rate estimation because it detects DNA presence in all life stages of the parasite and hence has limited Plasmodium life-stage specificity. In this case, use of Mt COX‑I PCR together with CS-ELISA is recommended to confirm the real infection [78]. During sporozoite targeted DNA extraction and CS-ELISA assay, the head and thorax are commonly used, and infectious sporozoite stages are targeted from these organs [79, 80]. This helps to eliminate misreporting of positivity rates, which is mostly due to oocyst presence in the abdomen and/or rings,’ trophozoites,’ and gametocytes’ DNA in freshly ingested blood or incompletely digested blood.
Regarding the sporozoite species detected, the tested anophelines were infected with either P. vivax or P. falciparum sporozoites. Mixed infection of P. vivax and P. falciparum was only reported from one An. arabiensis sample. This supports the fact that P. vivax and P. falciparum are the predominant Plasmodium species causing malaria but are rarely present at the same time. Even though P. ovale and P. malariae cases have been reported in Ethiopia [81, 82], none of the tested samples were found infected with P. ovale and P. malariae sporozoites.
Strength and limitations of the study
This is the first systematic review to our knowledge of the sporozoite infection proportion of Anopheles mosquitoes in Ethiopia conducted by reviewing original articles published in the last 2 decades. A recent systematic review by Adugna et al. [68] assessed only Anopheles species composition, abundance, and distribution in Ethiopia. However, our study might not provide an updated list of Anopheles mosquito species in Ethiopia since we have included only studies that were conducted to determine infection status and reported sporozoite proportions.
Conclusions
The current review revealed that An. arebiensis has been the dominant malaria vector over the years, with the highest sporozoite infection proportion being 2.85% and an average of 0.90% over the study period. Other species contributing to malaria transmission in the area were An. pharoensis, An. coustani complex, An. funestus, An. stephensi, An. cinereus, and An. demeilloni. The emergence of new vector species, in particular An. stephensi, is particularly concerning and should be investigated further. Continued assessment of the specific role of different vector species is important to guide more effective control of malaria in the region. The use of head and thorax samples is recommended for sporozoite targeted DNA extraction and CS-ELISA assay to exclude contamination of oocyst sporozoites present in the abdomen. Moreover, Anopheles mosquito samples used for estimation of sporozoite infection proportion should be caught by human landing catch to avoid cross-reactivity since most mosquitoes that approach humans are expected to be unfed.
Availability of data and materials
The data supporting the findings of the study must be available within the article and/or its supplementary materials, or deposited in a publicly available database.
Abbreviations
- DNA:
-
Deoxyribonucleic acid
- PCR:
-
Polymerase chain reaction
- CSP:
-
Circumsporozoite protein
- ELISA:
-
Enzyme linked immunosorbent assay
- HLC:
-
Human landing catch
- JBI:
-
Joanna Briggs Institute
References
WHO. World malaria report 2020. Geneva: World Health Organization; 2020.
WHO. World malaria report 2018. Geneva: World Health Organization; 2018.
Zhou G, Minakawa N, Githeko AK, Yan G. Association between climate variability and malaria epidemics in the East African highlands. Proc Natl Acad Sci. 2004;1:2375–80.
Reiter P. Climate change and highland malaria in the tropics. Abstract of presentation to avoiding dangerous climate change, international symposium on the stabilization of greenhouse gas concentration. Exeter: Hadley Centre, Met officer; 2004.
Hay SI, Rogers DG, Randpolph SE, Stern DI, Cox J, Shanks GD, et al. Hot topic or hot air? Climate change and malaria resurgence in East African highlands. Trends Parasitol. 2002;8:530–4.
Williams J, Pinto J. Training manual on malaria entomology for entomology and vector control technicians (basic level). USAID. Washington, DC; 2012:78.
Harbach RE. The Phylogeny and classification of Anopheles. Anopheles mosquitoes—new insights into malaria vectors. InTech; 2013.
Sinka ME, Bangs MJ, Manguin S, Rubio-Palis Y, Chareonviriyaphap T, Coetzee M, et al. A global map of dominant malaria vectors. Parasit Vectors. 2012;5:69.
Irish SR, Kyalo D, Snow RW, Coetzee M. Updated list of Anopheles species (Diptera: Culicidae) by country in the Afrotropical region and associated islands. Zootaxa. 2020;4747:4747.
Gaffigan TV, Wilkerson RC, Pecor JE, Stoffer JA, Anderson T. Systematic catalog of Culicidae. Walter reed biosystematics unit, division of entomology, Walter reed army institute of research, Silver Spring; 2013.
Kyalo D, Amratia P, Mundia CW, Mbogo CM, Coetzee M, Snow RW. A geo-coded inventory of anophelines in the Afrotropical region south of the Sahara: 1898–2016. Wellcome Open Res. 2017;2:57.
Jaleta KT, Hill SR, Seyoum E, Balkew M, Gebre-Michael T, Ignell R, et al. Agro-ecosystems impact malaria prevalence: large-scale irrigation drives vector population in western Ethiopia. Malar J. 2013;12:350.
Solomon K, Yihenew A, Eline B, Habte T, Dawit A, Beyene P. The impact of a small-scale irrigation scheme on malaria transmission in Ziway area, Central Ethiopia. Trop Med Int Health. 2010;15:41–50.
White GB, Tesfaye F, Boreham PFL, Lemma G. Malaria vector capacity of Anopheles arabiensis and Anopheles quadriannulatus in Ethiopia; chromosomal interactions of after 6 years storage of field preparations. Trans R Soc Trop Med Hyg. 1980;74:683–4.
Faulde MK, Rueda LM, Khaireh BA. First record of the Asian malaria vector Anopheles stephensi and its possible role in the resurgence of malaria in Djibouti. Horn of Africa Acta Trop. 2014;139:39–43.
Carter TE, Yared S, Gebresilassie A, Bonnell V, Damodaran L, Lopez K, et al. First detection of Anopheles stephensi Liston, 1901 (Diptera: culicidae) in Ethiopia using molecular and morphological approaches. Acta Trop. 2018;188:180–6.
WHO. Vector alert: Anopheles stephensi invasion and spread. Geneva: World Health Organization; 2019.
WHO. WHO initiative to stop the spread of Anopheles stephensi in Africa. Geneva: World Health Organization; 2022.
Ndenga BA, Wambua S, Owuor KO, Omukuti R, Chemutai S, Arabu D, et al. Serendipitous detection of Anopheles stephensi in Kisumu, Kenya in June 2022. MedRxiv. 2022;2023:2023–105.
Lemma W, Alemu K, Birhanie M, Worku L, Niedbalski J, McDowell MA, et al. Anopheles cinereus implicated as a vector of malaria transmission in the highlands of north-west Ethiopia. Parasit Vectors. 2019;12:557.
Fontenille D, Meunier JY, Nkondjio CA, Tchuinkam T. Use of circumsporozoite protein enzyme-linked immunosorbent assay compared with microscopic examination of salivary glands for calculation of malaria infectivity rates in mosquitoes (Diptera: Culicidae) from Cameroon. J Med Entomol. 2001;38:451–4.
Burkot TR, Williams JL, Schneider I. Identification of Plasmodium falciparum-infected mosquitoes by a double antibody enzyme-linked immunosorbent assay. Am J Trop Med Hyg. 1984;33:783–8.
Marie A, Boissiere A, Tsapi MT, Poinsignon A, Awono-Ambéné PH, Morlais I, et al. Evaluation of a real-time quantitative PCR to measure the wild Plasmodium falciparum infectivity rate in salivary glands of Anopheles gambiae. Malar J. 2013;12:224.
Hasan AU, Suguri S, Sattabongkot J, Fujimoto C, Amakawa M, Harada M, et al. Implementation of a novel PCR based method for detecting malaria parasites from naturally infected mosquitoes in Papua New Guinea. Malar J. 2009;8:182.
Pinheirob VE, Thaithongc S, Browna KN. High sensitivity of detection of human malaria parasites by the use of nested polymerase chain reaction. Mol Biochem Parasitol. 1993;61:315–20.
Wilson MD, Ofosu-Okyere A, Okoli AU, McCall PJ, Snounou G. Direct comparison of microscopy and polymerase chain reaction for the detection of Plasmodium sporozoites in salivary glands of mosquitoes. Trans R Soc Trop Med Hyg. 1998;92:482–3.
Li F, Niu C, Ye B. Nested polymerase chain reaction in detection of Plasmodium vivax sporozoites in mosquitoes. Chin Med J. 2001;114:654–7.
Vaughan JA, Noden BH, Beier JC. Population dynamics of Plasmodium falciparum sporogony in laboratory-infected Anopheles gambiae. J Parasitol. 1992;1992:716–24.
Federal Ministry of Health. Entomological profile of malaria in Ethiopia. FMoH. Addis Ababa, Ethiopia. 2007.
Liberati A, Altman DG, Tetzlaff J, Mulrow C, Gotzsche PC, Ioannidis JP, et al. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. Ann Intern Med. 2009;151:W65.
Joanna Briggs Institute. Joanna briggs institute critical appraisal checklist for studies reporting prevalence data. Adelaide: JBI Libr Syst Rev. 2017;2017:1.
Wirtz RA, Zavala F, Charoenvit Y, Campbell GH, Burkot TR, Schneider I, et al. Comparative testing of monoclonal antibodies against Plasmodium falciparum sporozoites for ELISA development. Bull World Health Organ. 1987;65:39.
Beier JC, Perkins PV, Wirtz RA, Whitmire RE, Mugambi M, Hockmeyer WT. Field evaluation of an Enzyme-Linked Immunosorbent Assay (ELISA) for Plasmodium falciparum sporozoite detection in anopheline mosquitoes from Kenya. Am J Trop Med Hyg. 1987;36:459–68.
Wirtz RA, Sattabongkot J, Hall T, Burkot TR, Rosenberg R. Development and evaluation of an enzyme-linked immunosorbent assay for Plasmodium vivax-VK247 sporozoites. J Med Entomol. 1992;29:854–7.
Tadesse FG, Ashine T, Teka H, Esayas E, Messenger LA, Chali W, et al. Anopheles stephensi mosquitoes as vectors of Plasmodium vivax and falciparum, Horn of Africa, 2019. Emerg Infect Dis. 2021;27:603.
Tesfaye S, Belyhun Y, Teklu T, Mengesha T, Petros B. Malaria prevalence pattern observed in the highland fringe of Butajira, southern Ethiopia: a longitudinal study from parasitological and entomological survey. Malar J. 2011;10:153.
Kibret S, Wilson GG, Tekie H, Petros B. Increased malaria transmission around irrigation schemes in Ethiopia and the potential of canal water management for malaria vector control. Malar J. 2014;13:360.
Getaneh A, Yimer M, Alemu M, Dejazmach Z, Alehegn M, Tegegne B. Species composition, parous rate, and infection rate of Anopheles mosquitoes (Diptera: Culicidae) in Bahir Dar city administration, northwest Ethiopia. J Med Entomol. 2021;58:1874–9.
Taye A, Hadis M, Adugna N, Tilahun D, Wirtz RA. Biting behavior and Plasmodium infection rates of Anopheles arabiensis from Sille. Ethiopia Acta Trop. 2006;97:50–4.
Animut A, Balkew M, Gebre-Michael T, Lindtjørn B. Blood meal sources and entomological inoculation rates of anophelines along a highland altitudinal transect in south-central Ethiopia. Malar J. 2013;12:76.
Gari T, Kenea O, Loha E, Deressa W, Hailu A, Balkew M, et al. Malaria incidence and entomological findings in an area targeted for a cluster-randomized controlled trial to prevent malaria in Ethiopia: results from a pilot study. Malar J. 2016;15:145.
Habtewold T, Walker AR, Curtis CF, Osir EO, Thapa N. The feeding behaviour and Plasmodium infection of Anopheles mosquitoes in southern Ethiopia in relation to use of insecticide-treated livestock for malaria control. Trans R Soc Trop Med Hyg. 2001;95:584–6.
Kenea O, Balkew M, Tekie H, Gebre-Michael T, Deressa W, Loha E, et al. Human-biting activities of Anopheles species in south-central Ethiopia. Parasit Vectors. 2016;9:527.
Taye B, Lelisa K, Emana D, Asale A, Yewhalaw D. Seasonal dynamics, longevity, and biting activity of anopheline mosquitoes in southwestern Ethiopia. J Insect Sci. 2016;16:6.
Kibret S, Alemu Y, Boelee E, Tekie H, Alemu D, Petros B. The impact of a small-scale irrigation scheme on malaria transmission in Ziway area, Central Ethiopia. Trop Med Int Health. 2010;15:41–50.
Lelisa K, Asale A, Taye B, Emana D, Yewhalaw D. Anopheline mosquitoes behaviour and entomological monitoring in southwestern Ethiopia. J Vector Borne Dis. 2017;54:240.
Massebo F, Balkew M, Gebre-Michael T, Lindtjørn B. Entomologic inoculation rates of Anopheles arabiensis in Southwestern Ethiopia. Am J Trop Med Hyg. 2013;89:466.
Abraham M, Massebo F, Lindtjørn B. High entomological inoculation rate of malaria vectors in area of high coverage of interventions in Southwest Ethiopia: implication for residual malaria transmission. Parasit Epidemiol Control. 2017;2:61–9.
Degefa T, Zeynudin A, Godesso A, Michael YH, Eba K, Zemene E, et al. Malaria incidence and assessment of entomological indices among resettled communities in Ethiopia: a longitudinal study. Malar J. 2015;14:24.
Eba K, Habtewold T, Yewhalaw D, Christophides GK, Duchateau L. Anopheles arabiensis hotspots along intermittent rivers drive malaria dynamics in semi-arid areas of Central Ethiopia. Malar J. 2021;20:154.
Daygena TY, Massebo F, Lindtjørn B. Variation in species composition and infection rates of Anopheles mosquitoes at different altitudinal transects, and the risk of malaria in the highland of Dirashe Woreda, south Ethiopia. Parasit Vectors. 2017;10:343.
Getachew D, Gebre-Michael T, Balkew M, Tekie H. Species composition, blood meal hosts and Plasmodium infection rates of Anopheles mosquitoes in Ghibe River Basin, southwestern Ethiopia. Parasit Vectors. 2019;12:257.
Zemene E, Belay DB, Tiruneh A, Lee MC, Yewhalaw D, Yan G. Malaria vector dynamics and utilization of insecticide-treated nets in low-transmission setting in Southwest Ethiopia: implications for residual transmission. BMC Infect Dis. 2021;21:882.
Dugassa S, Murphy M, Chibsa S, Tadesse Y, Yohannes G, Lorenz LM, et al. Malaria in migrant agricultural workers in western Ethiopia: entomological assessment of malaria transmission risk. Malar J. 2021;20:95.
Degefa T, Githeko AK, Lee MC, Yan G, Yewhalaw D. Patterns of human exposure to early evening and outdoor biting mosquitoes and residual malaria transmission in Ethiopia. Acta Trop. 2021;216:105837.
Fettene M, Hunt RH, Coetzee M, Tessema F. Behaviour of Anopheles arabiensis and Anopheles quadriannulatus sp. B mosquitoes and malaria transmission in southwestern Ethiopia. Afr Entomol. 2004;12:83–7.
Aklilu E. Species composition, seasonal variation and roles of Anopheles mosquitoes in the transmission of malaria in Koka villages, Central Ethiopia (Doctoral dissertation, Addis Ababa University).
Kenea O, Balkew M, Tekie H, Deressa W, Loha E, Lindtjørn B, et al. Impact of combining indoor residual spraying and long-lasting insecticidal nets on Anopheles arabiensis in Ethiopia: results from a cluster randomized controlled trial. Malar J. 2019;18:182.
Kibret S, Wilson GG, Ryder D, Tekie H, Petros B. Malaria impact of large dams at different eco-epidemiological settings in Ethiopia. Trop Med Health. 2017;45:4.
Getawen SK, Ashine T, Massebo F, Woldeyes D, Lindtjørn B. Exploring the impact of house screening intervention on entomological indices and incidence of malaria in Arba Minch town, southwest Ethiopia: a randomized control trial. Acta Trop. 2018;181:84–94.
Yewhalaw D, Kelel M, Getu E, Temam S, Wessel G. Blood meal sources and sporozoite rates of Anophelines in Gilgel-Gibe dam area, Southwestern Ethiopia. J Parasitol Vector Biol. 2014;2014:1.
Kibret S, Lautze J, Boelee E, McCartney M. How does Ethiopian dam increase malaria? Entomological determinants around the Koka reservoir. Trop Med Int Health. 2012;17:1320–8.
Kindu M, Aklilu E, Balkew M, Gebre-Michael T. Study on the species composition and ecology of anophelines in Addis Zemen, South Gondar, Ethiopia. Parasit Vectors. 2018;11:215.
Nigatu W, Asale A, Massebo F, Yohannes M, Mekuriaw W, Wuletaw Y, et al. Entomological surveillance in the context of malaria elimination in some selected sentiniel sites of Ethiopia. Ethiop J Public Health Nutr. 2020;3:62–71.
Balkew M, Mumba P, Yohannes G, Abiy E, Getachew D, Yared S, et al. An update on the distribution, bionomics, and insecticide susceptibility of Anopheles stephensi in Ethiopia, 2018–2020. Malar J. 2021;20:263.
Tirados I, Costantini C, Gibson G, Torr SJ. Blood-feeding behaviour of the malarial mosquito Anopheles arabiensis: implications for vector control. Med Vet Entomol. 2006;20:425–37.
Bekele D, Belyhun Y, Petros B, Deressa W. Assessment of the effect of insecticide-treated nets and indoor residual spraying for malaria control in three rural kebeles of Adami Tulu District, South Central Ethiopia. Malar J. 2012;11:127.
Adugna F, Wale M, Nibret E. Review of Anopheles mosquito species, abundance, and distribution in Ethiopia. J Trop Med. 2021;2021:6726622.
Krafsur ES. Anopheles nili as a vector of malaria in a lowland region of Ethiopia. Bull World Health Organ. 1970;42:466.
Aschale Y, Ayehu A, Worku L, Addisu A, Zeleke AJ, Bayih AG, et al. Anopheles gambiae s.l (Diptera: Culicidae) seasonal abundance, abdominal status and parity rates in Metema-Armachiho lowland, Northwest Ethiopia. BMC Infect Dis. 2020;20:333.
Adugna T, Getu E, Yewhelew D. Parous rate and longevity of anophelines mosquitoes in Bure district, northwestern Ethiopia. PLoS ONE. 2022;17:e0263295.
Adugna T, Getu E, Yewhalaw D. Species diversity and distribution of Anopheles mosquitoes in Bure district, Northwestern Ethiopia. Heliyon. 2020;6:e05063.
Balkew M, Gebre-Michael T, Hailu A. Insecticide susceptibility level of Anopheles arabiensis in two agro-development localities in eastern Ethiopia. Parassitologia. 2003;45:1–3.
Carter TE, Yared S, Hansel S, Lopez K, Janies D. Sequence-based identification of Anopheles species in eastern Ethiopia. Malar J. 2019;18:135.
Durnez L, Van Bortel W, Denis L, Roelants P, Veracx A, Dinh Trung H, et al. False positive circumsporozoite protein ELISA: a challenge for the estimation of the entomological inoculation rate of malaria and for vector incrimination. Malar J. 2011;10:195.
Lochouarn L, Fontenille D. ELISA detection of malaria sporozoites: false positive results in Anopheles gambiae s.l. associated with bovine bloodmeals. Trans R Soc Trop Med Hyg. 1999;93:101–2.
Somboon P, Morakote N, Koottathep S, Trisanarom U. Detection of sporozoites of Plasmodium vivax and Plasmodium falciparum in mosquitoes by ELISA: false positivity associated with bovine and swine blood. Trans R Soc Trop Med Hyg. 1993;87:322–4.
Hendershot AL, Esayas E, Sutcliffe AC, Irish SR, Gadisa E, Tadesse FG, et al. A comparison of PCR and ELISA methods to detect different stages of Plasmodium vivax in Anopheles arabiensis. Parasit Vectors. 2021;14:473.
Echeverry DF, Deason NA, Makuru V, Davidson J, Xiao H, Niedbalski J, et al. Fast and robust single PCR for Plasmodium sporozoite detection in mosquitoes using the cytochrome oxidase I gene. Malar J. 2017;16:230.
Kumpitak C, Nguitragool W, Cui L, Sattabongkot J, Bantuchai S. Detection of Plasmodium sporozoites in Anopheles mosquitoes using an enzyme-linked immunosorbent assay. J Vis Exp. 2021;30:e63158.
Alemu A, Fuehrer HP, Getnet G, Tessema B, Noedl H. Plasmodium ovale curtisi and Plasmodium ovale wallikeri in North-West Ethiopia. Malar J. 2013;12:346.
Assefa A, Ahmed AA, Deressa W, Wilson GG, Kebede A, Mohammed H, et al. Assessment of subpatent Plasmodium infection in northwestern Ethiopia. Malar J. 2020;19:108.
Acknowledgements
We would like to acknowledge the authors of the original studies.
Funding
No funding to report.
Author information
Authors and Affiliations
Contributions
The review was conceived by YA. Data extraction, data analysis, interpretation, and drafting of the manuscript were conducted by YA and AG. All authors contributed to critical review of the manuscript. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1
. PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analysis) checklist for systematic review of sporozoite infection rate of Anopheles mosquitoes in Ethiopia, 2001–2021.
Additional file 2: Table S2
. Methodological qualities of original studies included in the systematic review of sporozoite infection rate of Anopheles mosquito in Ethiopia, 2001–2021.
Additional file 3: Table S3
. Descriptive summary of original studies included in the systematic review of sporozoite infection rate of Anopheles mosquito in Ethiopia, 2001–2021. CBT, cattle-baited trap; HDNT, human-baited double net traps; MA, mouth aspiration; PSC, pyrethrum spray sheet collection.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Aschale, Y., Getachew, A., Yewhalaw, D. et al. Systematic review of sporozoite infection rate of Anopheles mosquitoes in Ethiopia, 2001–2021. Parasites Vectors 16, 437 (2023). https://doi.org/10.1186/s13071-023-06054-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-06054-y