
Abstract
Background
The oriental eyeworm Thelazia callipaeda (Spirurida: Thelaziidae) is an emerging parasitic ocular nematode of carnivores and humans. In domestic animals and humans, the infection causes varying degrees of inflammation and lacrimation, and wild carnivores represent an important reservoir. In this study we examined the infection status and molecular characterization of T. callipaeda in two urban carnivores, raccoons Procyon lotor and wild Japanese raccoon dogs Nyctereutes viverrinus, in the Kanto region of Japan.
Methods
From January 2020 to December 2021, 193 carcasses including 178 raccoons and 15 raccoon dogs were examined for the presence of worms in the eye. The worms from infected animals (one worm per host) were morphologically identified as T. callipaeda. Worms (1–5 worms per host) were subjected to genetic analysis using mitochondrial cytochrome c oxidase subunit I gene sequences.
Results
The prevalence of T. callipaeda in raccoons and Japanese raccoon dogs was 20.2% (36/178) and 13.3% (2/15), respectively. The cox1 sequences from 56 worms from 38 animals revealed three haplotypes (h9, h10, and h12). Analysis of multiple worms for five raccoons showed co-infection of two different haplotypes (h9 and h10) in a single host. Comparing our data with published sequences, three sequences obtained from raccoons and raccoon dogs shared the same haplotypes as those reported in humans, dogs, and cats in Japan.
Conclusions
Our findings show a high prevalence of T. callipaeda in raccoons, suggesting that this invasive carnivore species serves as an important natural reservoir of T. callipaeda in the Kanto region of Japan, an area with the highest human population of the country.
Graphical Abstract

Similar content being viewed by others

Background
The Oriential eyeworm, Thelazia callipaeda Ralliet and Henry, 1910, is a parasitic nematode that infects the eyes of wildlife, domestic animals, and humans [1, 2]. Adult T. callipaeda live in the conjunctival sac, and ovoviviparous T. callipaeda females release L1 in lacrimal secretions [3,4,5,6]. These L1 are ingested by fruit flies of the genera Phortica and Amiota (Diptera: Drosophilidae) when they feed on tears and conjunctival fluids. Ingested L1 grow into L3 in a fly and are transmitted to the eyes of susceptible vertebrate hosts [4, 6].
Human and canine thelaziosis caused by T. callipaeda has been reported in many countries in recent years. Thelaziosis appears to be particularly prevalent in European countries and the Russian Far East, where the parasite has been detected in wild animals including gray wolves Canis lupus, European rabbits Oryctolagus cuniculus, golden jackals Canis aureus, common raccoon dogs Nyctereutes procyonoides, brown bears Ursus arctos, Asian black bears Ursus thibetanus, lynx Lynx lynx, sable Martes zibellina, beech marten Martes foina, and European wildcat Felis silvestris, suggesting a rapid expansion of the parasite [7,8,9,10,11,12,13,14,15]. In Japan, the first report of T. callipaeda was on Kyusyu, located in the southwest portion of the Japanese island chain, and the worms have been sporadically reported since the 1950s in humans and domestic dogs and cats [16,17,18]. It is generally believed that T. callipaeda spread to the eastern part of Japan, including the Kanto region, after the 1970s, where it has been detected in humans, dogs, cats, and Japanese raccoon dogs [19, 20].
The raccoon Procyon lotor is a medium-sized carnivore originally native to North and Central America. Raccoons were first introduced in Japan in the late 1960s [21], and they are now one of the most problematic non-native species in Japan. The Japanese government investigated feral raccoon distribution in 2006, confirming their dispersion across most of the prefectures of Japan. Feral raccoons are now considered an invasive species and prey on endangered amphibians and crustaceans, damage crops and houses, and compete for resources with sympatric mammals [21]. Meanwhile, the Japanese raccoon dog Nyctereutes viverrinus is an endemic medium-sized carnivore found throughout Japan [22]. This species lives in forested areas, but is also frequently found in agricultural land and urban areas [23]. In urban and suburban environments, the distributions of raccoons and raccoon dogs overlap, and both species may harbor pathogens, including parasites [7, 24, 25].
In Japan, although infection with T. callipaeda in domestic animals and humans has been reported sporadically in many areas [17,18,19,20, 26], the prevalence of T. callipaeda in wildlife has not been thoroughly investigated. Here, we examined the infection status of T. callipaeda in feral raccoons and wild raccoon dogs in the Kanto region of Japan. We also observed the genetic differences between T. callipaeda strains in Japan and its neighboring countries.
Methods
Study area
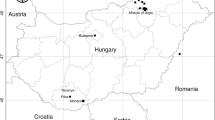
This survey was conducted in the Kanto region, which is located in the southeastern part of the main island of Japan and is dominated by the Kanto Plain, the largest plain in Japan (Fig. 1). The Kanto region has a humid subtropical climate, and the four seasons are sharply delineated. This region is the most populous and urbanized in Japan. The native carnivorous species, Japanese weasel Mustela itatsi, Japanese marten Martes melampus, Japanese badger Meles anakuma, Japanese red fox Vulpes vulpes japonica, and Japanese raccoon dogs have been observed only in the suburbs of the Kanto region, where woodlands and grasslands remain. Invasive carnivore species, such as raccoons and masked palm civets Paguma larvata, have been observed in residential areas [27].

Geographical distribution of Thelazia callipaeda-infected animals (red marker) and non-infected animals
Animal samples
From January 2020 to December 2021, 193 carcasses, including of 178 raccoons and 15 Japanese raccoon dogs, were examined for the presence of worms in the eye. All animals were caught by licensed hunters using box traps (Havahart, Model 1089; Woodstream, PA) and killed using a combination of xylocaine and pentobarbital sodium or carbon dioxide gas. These carcasses were bagged in two or three layers of plastic bags immediately after death, sent to Nippon Veterinary and Life Science University under freezing or frozen conditions, and subsequently necropsied. The age of all the animals was estimated according to the extent of tooth eruption. We classified the raccoons into two groups: pre-weaned juvenile raccoons (< 5 months old) and weaned subadult and adult raccoons (≥ 5 months old), hereafter referred to as “adult raccoons” [28]. Japanese raccoon dogs were divided into two groups: juvenile raccoons (< 6 months old) and subadult and adult raccoons (≥ 6 months old), hereafter referred to as “adult raccoon dogs” [29, 30]. Detailed information regarding the place of capture, as well as the weight, estimated age by stage, and sex of the animals is included in Table 1.
GIS mapping
The geographical distribution of T. callipaeda-infected and non-infected animals was displayed on the maps. ArcGIS Pro software (ESRI, Inc., West Redland, CA, USA) was used to project the capture locations of the examined animals.
Nematode collection
The third eyelid and conjunctival sacs of the animals were carefully examined for the presence of adult eyeworms. The worms were collected mechanically using sterile swabs and stored in 70% ethanol. The worms were mounted on slides in 50% glycerol and 40% ethanol solutions, observed using a BX53 microscope (Olympus, Tokyo), and identified as T. callipaeda according to morphological keys [31].
Molecular analysis
A total of 55 adults of T. callipaeda collected from the infected animals (1–5 worms per host) were used for molecular analysis. Genomic DNA was extracted from the middle part of each worm using a QIAGEN DNA mini kit (Qiagen, Germany), according to the manufacturer’s instructions. Double-distilled water was used as the negative control.
Partial fragments of mitochondrial cox1 were amplified using the specific primers NTF (5′-TGATTGGTGGTTTTGGTAA-3′) and NTR (5′-ATAAGTACGAGTATCAATATC-3′) [32]. PCR was performed using 20 μl reaction volumes, each containing 0.2 μl TaKaRa ExTaq polymerase (TaKaRa, Japan), 2 μl 10 × buffer, 1.6 μl dNTPs (2.5 mM of each), 0.2 μl of each primer (50 μM), 1.0 μl template, and 14.8 μl double-distilled water. The thermocycling conditions were as follows: initial denaturation at 95 °C for 5 min followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 40 °C for 30 s, and extension at 72 °C for 1 min. The final extension was performed at 72 °C for 10 min, followed by a holding step at 4 °C. PCR products were separated by electrophoresis on 1.5% agarose gels and visualized under an LED transilluminator after staining with GRGreen (BioCraft, Japan). The sizes of the PCR products were estimated by comparison with a 100-bp DNA ladder (Maestrogen, Taiwan). The PCR products were purified using ExoSAP-IT (Macrogen Corp., Japan) and analyzed using an ABI 3730xl DNA analyzer (Thermo Fisher Scientific, USA) with the same PCR primers.
Sequence similarity was studied using the BLASTN program available on the National Center for Biotechnology Information database (http://www.ncbi.nlm.nih.gov/Blast.cg). Multiple sequence alignment was performed using MAFFT ver. 7.505 with the Q-INS-I option [33]. For phylogenetic analysis, a 642-bp dataset generated from multiple alignments of 117 cox1 sequences (62 from the public database and 55 determined in this study) was used for haplotype and network analyses (Additional files 1 and 2). Definition and numbering of haplotypes were based on previous studies [31, 34]. Haplotype networks were generated using the medium-joining network algorithm in PopART ver. 1.7 [35, 36]. The haplotype diversity and nucleotide diversity at each site were calculated using DnaSP ver. 6.12.03 [37].
Results
Prevalence and geographical distribution of Thelazia callipaeda
A total of 233 T. callipaeda individuals were collected from 36 raccoons (Table 1). The mean worm burden, SD and range of parasite number (in parentheses) were 6.5 ± 8.4 (1–44). Thirty infected individuals (16 males and 14 females) were captured in Tokyo, and six individuals (4 males and 2 females) were captured in Kanagawa. Eight worms were collected from two male Japanese raccoon dogs (Table 1).
In raccoons, the overall infection rate was 20.2% (36/178; 95% CI 15.0–26.8) (Table 2). Juvenile male raccoons showed the highest infection rate of 36.4% (8/22; 95% CI 19.6–57.1), followed by adult female raccoons at 23.5% (12/51; 95% CI 13.9–36.9), adult male raccoons at 14.9% (13/87; 95% CI 8.8–24.0), and juvenile female raccoons at 11.1% (3/18; 95% CI 5.2–40.2). The overall infection rate in Japanese raccoon dogs was 13.3% (2/15; 95% CI 2.5–39.1) (Table 3). In Japanese raccoon dogs, T. callipaeda was detected only in adult male individuals (25.0%, 2/8; 95% CI 6.3–59.9).
Molecular identification
Partial cox1 sequences (667 bp) were also determined separately for each individual worm, and sequence alignments differentiated the three haplotypes. Nineteen worms from 16 raccoons and one worm from a Japanese raccoon dog shared one sequence type, which was 100% identical to T. callipaeda sequences reported from humans in the Okayama Prefecture, Japan (640/640 bp; accession no. AB538283) and a dog in Tokyo, Japan (660/660 bp; accession no. AB852544). Zhang et al. [34] referred to the former sequence reported from Okayama as haplotype h9 and the latter reported from Tokyo as h11. However, since these sequences are indistinguishable from each other (635/635 bp), we have unified them and identified as h9. The second sequence, obtained from 32 worms from 22 individual raccoons and one worm from a Japanese raccoon dog, was identical to the sequence reported in humans in Kumamoto (596/569 bp; accession no. LC565618) and Oita Prefectures (585/585 bp; accession no. LC565619) and dogs (660/660 bp; accession nos. AB852543, AB852546–AB852548, AB852550) and a cat (660/660 bp, accession no. AB852549) in Tokyo, Japan. The third sequence type, obtained from two worms and two raccoons, was identical to a sequence recorded from a dog in Tokyo (660/660 bp; accession no. AB852545). The second and third haplotypes were formerly known as h10 and h12, respectively [34]. In the five raccoons analyzed, with 4–5 worms per individual, all had mixed infection with h9 and h10 haplotypes (Table 1). Representative sequences were deposited in the DNA Data Bank of Japan under accession numbers LC46896–LC46898.
Multiple alignment of the available sequences and sequences obtained in this study revealed 24 haplotypes (Table 4); 3 from Japan (h9, h10, and h12), 17 from China, 3 from Korea, and 1 from European countries (Germany, Greece, The Netherlands, Italy, Romania, Slovakia, and Serbia). Upon comparison of haplotypes recorded from Japan and China, which had larger sample sizes, both haplotype and nucleotide diversity were higher in China.
A median-joining network of T. callipaeda haplotypes revealed that the Chinese haplotypes were centrally located in the network and were accompanied by some haplotypes reported from China and Korea (Fig. 2). The three Japanese haplotypes and the European haplotype (h1) were located outside this group and were not connected directly to each other, except for h9 and h12.

Haplotype network of cox1 haplotypes found in Thelazia callipaeda from Asian and European countries. Each cycle represents a haplotype with the size proportional to its sampling frequency. Colors represent the locality of the host reported. Black dots and bars on lines represent hypothetical haplotypes and the number of additional mutation steps between two haplotypes
Discussion
To the best of our knowledge, T. callipaeda has not been detected in raccoons in North America or any countries where raccoons have been introduced [7]. Meanwhile, T. callipaeda has reportedly been detected in multiple individuals of common raccoon dogs in Russia [9, 10], but only one case in Japanese raccoon dogs has been reported in Japan [38]. In this study, 20.2% of raccoons and 13.3% of Japanese raccoon dogs in the Kanto region of Japan were infected with T. callipaeda. Few studies have examined the infection rate of T. callipaeda in animals in Japan: a survey in 1981 in the Hyogo Prefecture found a prevalence of 0.2% (1/521) in outdoor dogs [26]; a survey in 1992 of outdoor dogs and cats in the Hiroshima Prefecture found a prevalence of 1.0% (5/500) and 0.4% (2/250), respectively [39]; a survey in 2009 of outdoor dogs and cats in Tokyo found 4.0% (4/100) and 2.0% (2/100) prevalence, respectively [20]. Compared to these facts, this study reports the first investigation of thelaziosis in raccoons, and the infection rates in raccoons and sympatric Japanese raccoon dogs are higher than other regions of Japan, suggesting that these animals are more involved in the transmission of T. callipaeda in the Kanto region of Japan.
Genetic analysis showed that haplotype diversity in T. callipaeda collected from Japan was lower than among that from the continental origin, as Japan has only three haplotypes of T. callipaeda (h9, h10, and h12). These three Japanese haplotypes appear to have low host specificity, as h9 has been reported in humans, domestic dogs, raccoons, and Japanese raccoon dogs; h10 has been reported in humans, dogs, cats, raccoons, and Japanese raccoon dogs; and h12 has been reported in dogs and raccoons. Of these haplotypes, h9 and h12 are genetically related, whereas h10 has a separate lineage. The low number of haplotypes and the genetic diversity of T. callipaeda in Japan may be a result of its relatively recent introduction. A similar situation was observed during its initial introduction in Italy [40], followed by France [41] and Spain [42], where the same haplotype (h1) was detected. Future studies on the infection status of T. callipaeda and its haplotypes from other native Japanese carnivores, such as Japanese weasels, martens, badgers, and red foxes, are required.
Feral raccoons in Japan emerged after individuals escaped from zoos in the 1960s or were kept as exotic pets in the 1970s, and their distribution has spread from urban areas to suburban areas [21]. Thus, raccoons are now part of the urban wildlife of Japan and are found in residential areas, green spaces, and rivers in suburban areas. Raccoon dogs frequently visit residential areas but use woodlands and grasslands more frequently than raccoons [27]. The detection of T. callipaeda in raccoons and raccoon dogs and the identical molecular characteristics between our samples and the samples detected in humans, dogs, and cats indicate that this zoonotic nematodes is able to infect and spread between many species frequently found in urban areas in Japan.
Conclusion
Feral raccoons and wild Japanese raccoon dogs in the Kanto region of Japan showed high T. callipaeda infection rates and the same T. callipaeda with haplotypes identical to those identified for the parasites found in humans, dogs, and cats. The results of this study indicate that invasive raccoons are additional definitive hosts for T. callipaeda in Japan. The feral raccoon is a successful urban wildlife species in multiple countries and thus may play a role in transmitting the oriental worm to domestic dogs, cats, and possibly to humans.
Availability of data and materials
The manuscript and its supporting information files contain all necessary information.
Abbreviations
- CI:
-
Confidence interval
- cox1 :
-
Cytochrome c oxidase subunit I gene
- L1:
-
First stage larvae
- L3:
-
Third stage larvae
- PCR:
-
Polymerase chain reaction
- SD:
-
Standard deviation
References
Shen J, Gasser RB, Chu D, Wang Z, Cantacessi C, Otranto D. Human thelaziasis—a neglected parasitic disease of the eye. J Parasitol. 2006;92:872–5.
Otranto D, Mendoza-Roldan JA, Dantas-Torres F. Thelazia callipaeda. Trends Parasitol. 2021;37:263–4.
Otranto D, Lia RP, Buono V, Traversa D, Giangaspero A. Biology of Thelazia callipaeda (Spirurida, Thelaziidae) eyeworms in naturally infected definitive hosts. Parasitology. 2004;129:627–33.
Otranto D, Traversa D. Thelazia eyeworm: an original endo- and ecto-parasitic nematode. Trends Parasitol. 2005;21:1–4.
Otranto D, Lia RP, Cantacessi C, Testini G, Troccoli A, Shen JL, et al. Nematode biology and larval development of Thelazia callipaeda (Spirurida, Thelaziidae) in the drosophilid intermediate host in Europe and China. Parasitology. 2005;131:847–55.
Otranto D, Cantacessi C, Testini G, Lia RP. Phortica variegate as an intermediate host of Thelazia callipaeda under natural conditions: evidence for pathogen transmission by a male arthropod vector. Int J Parasitol. 2006;36:1167–73.
Otranto D, Deplazes P. Zoonotic nematodes of wild carnivores. Int J Parasitol Parasites Wildl. 2019;9:370–83.
Hodžić A, Latrofa MS, Annoscia G, Alić A, Beck R, Lia RP, et al. The spread of zoonotic Thelazia callipaeda in the Balkan area. Parasit Vectors. 2014;7:352.
Khrustalev AV, Shaitanov VM, Seredkin IV. New evidence for the spread of Thelazia callipaeda in the Far East. Med Parazitol (Mosck). 2015;1:46–8 (In Russian with English abstract).
Odoevskaya I, Khrustalev AV, Shaitanov VM, Seriodkin IV, Panayotova-Pencheva MS. Occurrence of the nematode Thelazia callipaeda Railliet and Henry, 1910 (Spirurida, Thelaziidae) in wild carnivores in the Russian Far East. Acata Zool Bulg. 2015;67:561–6.
Gama A, Pires I, Canado M, Coutinho T, Lopes AP, Latrofa MS, et al. First report of Thelazia callipaeda infection in wild European rabbits (Oryctolagus cuniculus) in Portugal. Parasit Vectors. 2016;9:236.
Mihalca AD, Ionică AM, D’Amico G, Daskalaki AA, Deak G, Matei IA, et al. Thelazia callipaeda in wild carnivores from Romania: new host and geographical records. Parasit Vectors. 2016;9:350.
Bezerra-Santos MA, Moroni B, Mendoza-Roldan JA, Perrucci S, Cavicchio P, Cordon R, et al. Wild carnivores and Thelazia callipaeda zoonotic eyeworm: a focus on wolves. Int J Parasitol Parasites Wildl. 2022;17:239–43.
Čabanová V, Miterpáková M, Oravec M, Hurníková Z, Jerg S, Nemčíková G, et al. Nematode Thelazia callipaeda is spreading across Europe. The first survey of red foxes from Slovakia. Acta Parasitol. 2018;63:160–6.
Papadopoulos E, Komnenou A, Karamanlidis AA, Bezerra-Santos MA, Otranto D. Zoonotic Thelazia callipaeda eyeworm in brown bears (Ursus arctos): a now host record in Europe. Transbound Emerg Dis. 2022;69:235–9.
Suzuki T, Uchida A, Itagaki H, Baba K. A case of canine thelaziasis in Kawasaki, Kanagawa. J Jpn Vet Med Assoc. 1978;31:458–61 (In Japanese with English abstract).
Kagei N, Hayashi S, Ishida T, Yamaguti T, Asami K, Takeuchi T. A case of Thelaziasis callipaeda in Tokyo, Japan. Jpn J Parasitol. 1981;30:337–44 (In Japanese with English abstract).
Fukumoto S, Yamada S, Konno K, Nishiwaki K, Ishida N, Furuya M, et al. Current situation of canine and feline Thelazia callipaeda infection in Japan. Jpn J Vet Parasitol. 2010;9:123 (In Japanese).
Kumase Y, Kato M, Takabatake M, Nakayama T, Ogihara N, Hasegawa H. A case of infestation of Thelazia callipaeda in the conjunctival sac with nine worms in one eye. Jpn J Clin Ophthalmol. 2010;64:1747–50 (In Japanese).
Okajima J, Sohmura Y, Iwaki T. Oriental eyeworm in dogs and cats in Tokyo. Jui Chikusan Shinpo. 2011;64:33–7 (In Japanese. The title was translated into English).
Ikeda T. Procyon lotor (Linnaeus, 1758). In: Ohdachi D, Ishibashi Y, Iwasa MA, Fukui D, Saitoh T, editors. The wild mammals of Japan. 2nd ed. Kyoto: Shoukadoh; 2015. p. 232–3.
Saeki M. Nyctereutes procyonoides (Gray, 1834). In: Ohdachi D, Ishibashi Y, Iwasa MA, Fukui D, Saitoh T, editors. The wild mammals of Japan. 2nd ed. Kyoto: Shoukadoh; 2015. p. 224–5.
Mitsuhashi I, Sako T, Teduka M, Koizumi R, Saito MU, Kaneko Y. Home range of raccoon dogs in an urban green area of Tokyo, Japan. J Mammal. 2018;99:732–40.
Doi K, Kato T, Hayama S. Infestation of introduced raccoons (Procyon lotor) with indigenous ixodid ticks on the Miura peninsula, Kanagawa Prefecture, Japan. Int J Parasitol Parasites Wildl. 2018;7:355–9.
Kass JM, Tingley MW, Tetsuya T, Koike F. Co-occurrence of invasive and native carnivorans affects occupancy patterns across environmental gradients. Biol Invasions. 2020;22:2251–66.
Uga S, Mizuno F, Matsumura T, Ito T, Shiomi M, Watanabe Y, et al. A helminthological survey of stray dogs in Hyogo Prefecture. Jpn J Parasitol. 1982;31:407–13 (In Japanese).
Abe G, Ikeda T, Tatsuzawa S. Differences in habitat use of the native raccoon dog (Nyctereutes procyonoides albus) and the invasive alien raccoon (Procyon lotor) in the Nopporo Natural Forest Park, Hokkaido, Japan. In: Koike F, Clour MN, Kawamichi M, De Poorter M, Iwatsuki K, editors. Assessment and control of biological invasion risks. Kyoto and Gland: Shoukadoh Book Sellar and IUCN; 2006. p. 116–21.
Montgomery GG. Tooth eruption in preweaned raccoons. J Wildl Manage. 1964;28:582–4.
Obara I. Age structure of Nyctereutes procyonoides viverrinus from the central and North region of Okayama Prefecture. J Mammal Soc Japan. 1983;9:204–7 (In Japanese with English abstract).
Kido N, Wada Y, TakahashiI M, Kamegaya C, Omiya T, Yamamoto Y. Prevalence of Dirofilaria immitis Infection in living raccoon dogs assessed by hematological examination. J Vet Med Sci. 2011;73:845–7.
Otranto D, Lia RP, Traversa D, Giannetto S. Thelazia callipaeda (Spirurida, Thelaziidae) of carnivores and humans: morphological study by light and scanning electron microscopy. Parassitologia. 2003;45:125–33.
Otranto D, Testini G, De Luca F, Hu M, Shamsi S, Gasser RB. Analysis of genetic variability within Thelazia callipaeda (Nematoda: Thelazioidea) from Europe and Asia by sequencing and mutation scanning of the mitochondrial cytochrome c oxidase subunit 1 gene. Mol Cell Probes. 2005;19:306–13.
Katoh K, Standley DM. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol Biol Evol. 2013;30:771–80.
Zhang X, Shi YL, Han LL, Xiong C, Yi SQ, Jiang P, et al. Population structure analysis of the neglected parasite Thelazia callipaeda revealed high genetic diversity in Eastern Asia isolates. PLos Negl Trop Dis. 2018;12:e0006165.
Bandelt H, Forster R, Röhl A. Median-joining networks for inferring intraspecific phylogenies. Mol Biol Evol. 1999;16:37–48.
Leigh JW, Bryant D. PopART: full-feature software for haplotype network construction. Methods Ecol Evol. 2015;6:1110–6.
Rozas J, Ferrer-Mata A, Sánchez-DelBarrio JC, Guirao-Rico S, Librado P, Ramos-Onsins SE, et al. DnaSP 6: DNA sequence polymorphism analysis of large datasets. Mol Biol Evol. 2017;34:3299–302.
Noda A. How do we make use of wastes from the zoo? Jpn J Zoo Wild Med. 2009;14:27–32 (In Japanese with English abstract).
Saito T, Morishige K, Tonugu Y. Parasites of pet dogs and cats in Fukuyama city. Jpn J Parasitol. 1995;44:149–53 (In Japanese with English abstract).
Otranto D, Ferroglio E, Lia RP, Traversa D, Rossi L. Current status and epidemiological observation of Thelazia callipaeda (Spirurida, Thelaziidae) in dogs, cats and foxes in Italy: a “coincidence” or a parasitic disease of the Old Continent? Vet Parasitol. 2003;116:315–25.
Dorchies P, Chaudieu G, Siméon LA, Cazalot G, Cantacessi C, Otranto D. First reports of autochthonous eyeworm infection by Thelazia callipaeda (Spirurida, Thelaziidae) in dogs and cat from France. Vet Parasitol. 2007;149:294–7.
Miró G, Montoya A, Hernández L, Dado D, Vázquez MV, Benito M, et al. Thelazia callipaeda: infection in dogs: a new parasite for Spain. Parasit Vectors. 2011;4:148.
Acknowledgements
We are grateful to the Wildlife Damage Control Support Center of Gunma Prefecture, the Agricultural Administration Department, Agricultural Policy Division, the City of Ota Municipality, the Greenery Environment Section, Natural Environment Division, Bureau of Environment, Tokyo Metropolitan Government, the Zephyrus Co., Ltd., the EGO Co., Ltd., Mr. Ryo Nomura (Kankyo Academy, Japan), and the Environmental Division, the Town of Hayama Municipality for trapping and providing animal carcasses. We also thank Dr. Hiroko Otsubo (Nippon Veterinary and Life Science University, Japan) and the members of the Laboratory of Wildlife Medicine, Nippon Veterinary and Life Science University for their cooperation to the sample collection.
Funding
This study was supported by JSPS KAKENHI grant no. JP22J01651 and the Wildlife Damage Control Support Center of the Gunma Prefecture, Japan.
Author information
Authors and Affiliations
Contributions
KD and TT conceived and designed the study, contributed with data analysis, drafted and revised the manuscript. MI, SC and TT performed the molecular characterization and TT identified morphologically the nematodes. KD, FY, TK and SH collected the samples. All authors read and approval the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Raccoon capturing procedures were conducted in accordance with the Management of Invasive Alien Species of the Japan Veterinary Medical Association (Japan Veterinary Medical Association, 2007). Japanese raccoon dog capturing procedures were conducted in accordance with the pest control measures for preventing agricultural damage and were killed by licensed hunters on behalf of the city of Takasaki and Ohta in Gunma Prefecture. All procedures followed the guidelines of The Mammal Society of Japan and approved by the Ethical Committee for the Care and Use of Animals at Nippon Veterinary and Life Science University (permit nos. 2020K-9 and 2021K-11).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Dataset S1.
Dataset of cox1 sequences of Thelazia callipaeda in text and fasta format.
Additional file 2: Table S2.
Geographical origin, host, GenBank/DDBJ/EMBL accession nos., and haplotype name of the Thelazia cox1 sequences.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Doi, K., Tokiwa, T., Imoto, M. et al. Molecular characterization of oriental eyeworm (Thelazia callipaeda) detected from raccoon (Procyon lotor) and Japanese raccoon dog (Nyctereutes viverrinus) in Kanto region, Japan. Parasites Vectors 16, 116 (2023). https://doi.org/10.1186/s13071-023-05736-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-023-05736-x