
Abstract
This article presents an overview of paratransgenesis as a strategy to control pathogen transmission by insect vectors. It first briefly summarises some of the disease-causing pathogens vectored by insects and emphasises the need for innovative control methods to counter the threat of resistance by both the vector insect to pesticides and the pathogens to therapeutic drugs. Subsequently, the state of art of paratransgenesis is described, which is a particularly ingenious method currently under development in many important vector insects that could provide an additional powerful tool for use in integrated pest control programmes. The requirements and recent advances of the paratransgenesis technique are detailed and an overview is given of the microorganisms selected for genetic modification, the effector molecules to be expressed and the environmental spread of the transgenic bacteria into wild insect populations. The results of experimental models of paratransgenesis developed with triatomines, mosquitoes, sandflies and tsetse flies are analysed. Finally, the regulatory and safety rules to be satisfied for the successful environmental release of the genetically engineered organisms produced in paratransgenesis are considered.
Graphical Abstract

Similar content being viewed by others
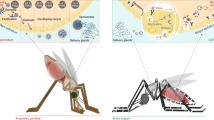

Background
When the transmission of pathogens by insect vectors is being considered, members of the orders Diptera and Hemiptera deserve particular attention. Many species belonging to these orders are of medical importance, transmitting a great variety of parasites causing diseases, including malaria, Chagas disease, leishmaniasis, sleeping sickness, filariasis, onchocerciasis and arboviruses.
Dipterans comprise approximately 150,000 known species catagorised in about 10,000 genera and 188 families. They include the mosquito (Culicidae), house fly (Muscidae), blow fly (Calliphoridae), robber fly (Asilidae), horse fly (Tabanidae), black fly (Simuliidae), sand fly (Phlebotominae), and gnat (e.g. Sciaridae) [1, 2]. Some species of mosquitoes, i.e. those belonging to the genera Anopheles, Aedes and Culex, act as vectors of many microorganisms that are etiologic agents of diseases, such as malaria, African typanosomiases, yellow fever, dengue, zika, chikungunya, West Nile fever and many others [3,4,5,6,7,8]. Mosquitoes alone are responsible for as many as 1 million deaths annually, including those from malaria for which high death rates have been occurring for many decades, with poor children aged < 5 years particularly affected [9]. Dengue fever transmitted by Aedes mosquitoes is the most common viral disease affecting 3.9 billion people annually in 129 countries, killing approximately 40,000 people every year [9, 10]. In addition, sand flies (genera Lutzomyia and Phlebotomus) are vectors of Leishmania and transmit leishmaniasis in Europe, northern Africa, the Middle East, Asia and parts of South America [11, 12]. Leishmaniasis, as well as onchcerciasis and filariasis transmitted by black flies and mosquito vectors, respectively, also cause permanent disfigurement in those infected. In Africa, several species of tsetse flies (Glossina spp.) are vectors of Trypanosoma brucei rhodesiense and T. b. gambiense, both of which cause sleeping sickness in humans, but these pathogens are not confined to humans since T. b. brucei, T. congolense, T. vivax, T. evansi, and T. equiperdum also result in African trypanosomiasis in cattle [13, 14]. The economic burden and human suffering caused by these diseases are enormous; for example, the direct costs (illness treatment and death) of malaria alone is estimated to be over 12 billion U.S. dollars per year [15].
Hemipterans have much lower impact than dipterans in terms of the numbers and disease burden of the human parasites vectored. The subfamily Triatominae includes Rhodnius prolixus and Triatoma infestans that transmit the flagellate protozoan Trypanosoma cruzi, which is the causative agent of Chagas disease, throughout South and Central America as well as the USA. The pathology of this disease is horrendous in patients with chronic inflammation of the heart, colon and nervous system. About 6 million people are estimated to be infected with T. cruzi in Latin America, of which one third will die from the disease [16]. The hemipterans also include the family Cimicidae, containing the bedbugs, such as Cimex lectularius, which also have the potential to transmit human diseases [17]. However, it should not be forgotten that the majority of hemipterans feed on plants and comprise the aphids, white flies and leaf hoppers that are significant vectors of viral diseases of crops [18].
The effect of global warming, the destruction of natural habitats and increases in international travel and trade have all served to increase both the spread of insect vector-borne parasitic diseases and the emergence of new microbial threats of pandemic proportions [19, 20]. For example, Aedes albopictus, the highly invasive Asian tiger mosquito, was probably introduced into Europe in 1990 via Italy in imported vehicle tyres. A favourable climate and global warming enhanced the mosquito’s spread and it has vectored outbreaks of chickungunya and dengue brought to Europe by international travellers [19]. The speed by which emerging pathogens can spread may be explosive, as illustrated with the Zika virus pandemic in the Americas; the virus was introduced into Brazil in 2015 and by 2016 it had infected approximately 211,700 people [21].
Challenges and ingenuity in controlling insect-borne diseases
Clearly, with the increased spread of many insect-vectored parasites and the arrival of newly emerging diseases, the need for effective control is absolutely vital. Control methods generally use several strategies and are focussed either on preventing the vector from feeding upon the host and transmitting the disease, or on treating infected individuals with drugs.
There are numerous vector-targeted control techniques, ranging from draining aquatic habitats and removing small domestic bodies of water to the use of mosquito nets, biological control agents, traps, spatial repellants, indoor residual spraying and anti-mosquito bands and creams [22,23,24,25]. These are often mediated through an integrated pest management scheme defined by the U.S. Department of Agriculture as “a sustainable, science-based, decision-making process that combines biological, cultural, physical, and chemical tools” [26].
One of the most successful twentieth century methods of controlling insect vectors, however, was the widespread use of insecticides. Among these, dichlorodiphenyltrichloroethane (DDT) was commonly used in the 1950s and 1960s to control mosquitoes and other pest species; however, it was banned in 1972 [27] despite probably saving hundreds of millions of lives [28]. In addition, insects have now developed resistance to the main chemicals used for pest control, namely the organochlorines, pyrethroids, organophosphates and carbamates [29, 30]. This resistance in African Anopheles mosquitoes has been described as “a worsening situation that needs urgent attention to maintain malaria control” [31]. Other vector control strategies have been utilised, such as insecticide-impregnated bed nets and microbial control agents [23], but both of these strategies have limitations. Bed nets are of limited use against Culex and Aedes spp. which bite more often outdoors in the daytime than at night [32]. In addition, the adoption of biological control agents, recognised as non-toxic alternatives to chemicals, may face resistance because of environmental concerns [33]. More recently, ingenious attempts to control insect vectors have turned to genetic modification so that the vector competence to transmit pathogens is reduced or, alternatively, the insect is engineered with a lethal transgene causing death during development [32]. There are, however, issues to be resolved before genetically modified vector insects can be widely released in the field to replace the wild populations, including concerns on the stability of the transgene, fitness of the transformed insects in the field and identification of genes driving favourable traits upon release, as well as ecological concerns over the release of genetically modified (GM) organisms [34, 35].
Once infection has occurred by the insect vector feeding on the human host, then tools are also available to suppress or kill the pathogen. These tools include vaccines and chemotherapy against the invasive parasites. Rapid progress is being made in both of these approaches, with researchers taking advantage, for example, of new information accruing from work on the immune interactions of host and parasite that has revealed key molecules as potential targets for vaccines and drugs [36]. Vaccines that are at present being tested in various phases against malaria [37] dengue [38], Zika [39], chikungunya [40] and leishmaniasis [40] will hopefully soon be available to prevent such infections. At present, the only vaccines for vector-borne diseases on the World Health Organisation-approved list without specific limitations are those against the yellow fever and Japanese encephalitis viruses [40]. In addition, a recent report has shown success rates of 74–77% with the malaria R21/MM vaccine in vaccinated children in Burkina Faso, even after 1 year [41]. The European Medicines Agency also recently accepted Japan’s leading drugmaker, Takeda, filing packages for its TAK-003 dengue vaccine candidate against any dengue virus serotype in people aged 4 to 60 years [42]. Many of these vaccines are therefore already showing favourable results although safety is still of some concern. At present, the prime method for treating protozoan parasites, including Plasmodium, Trypanosoma, Leishmania, Toxoplasma and Entameoba, is drug therapy, although resistance to these drugs is a growing problem [43,44,45]. One example of drug resistance is that to the sesquiterpine lactone, artemisinin, and its derivatives; these drugs are used against malaria as they act rapidly to clear parasites from the blood. Artemisinin is derived from the plant Artemisia annua and has been used for centuries in traditional Chinese Medicine to treat fevers; since 1980 it has saved the lives of millions of malaria patients [43]. Resistance to artemisinin-derived combinations was first detected in 2008 and has rapidly spread in Southeast Asia. The problem of drug resistance to one drug is exacerbated sometimes by the development of cross-resistance to other drugs. At present, Plasmodium falciparum, P. vivax and P. malariae are showing widespread resistance to a variety of drugs [43, 45]. Similar accounts of drug resistance are reported for leishmaniasis and African trypanosomiasis [43].
Drugs are also available for treating filariasis and onchocerciasis, and although these can be very effective in reducing worm loads, mass drug administration is required [46]. In addition, there is a lack of approved antivirals against the many present and emerging arboviruses [47] and other zoonotic viruses.
In summary, vaccines are still not widely available for preventing vector-borne pathogen infections, and disease vectors are becoming increasingly resistant to pesticides. Furthermore, parasites infecting patients are developing enhanced resistance against therapeutic drugs, and approved drugs are currently unavailable for treating arboviruses that are emerging more frequently and increasingly forming widespread epidemics. Therefore, new drugs and innovative strategies are urgently required for combating these vector-borne diseases [25, 36].
Definition and advantages of paratransgenesis
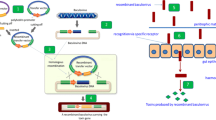
Paratransgenesis is a promising and particularly ingenious strategy currently being developed for controlling vector-transmitted diseases (Fig. 1). It utilises the genetically manipulated native microbiome (mutalistic symbiotic and commensal bacteria, fungi and viruses) [48] of the vector insect to inhibit or kill the disease pathogen. Native symbionts or commensals isolated from the vector are genetically transformed in vitro to produce anti-pathogen factors and then reintroduced to the insect to interrupt the life-cycle of the disease organism [32, 35, 49,50,51].

Summary of the analysis and selection of bacteria from vector microbiota for cultivation and genetic modification in vitro. The microorganism (A) is genetically modified by the insertion of an exogenous gene in a plasmid (B) or directly into the bacterial chromosome (C). The transgenic bacteria are offered to adult insects through an attractant bait. In the insect's digestive tract, the genetically modified microorganism expresses a peptide capable of interrupting the transmission of the parasite or a dsRNA that can silence genes in the parasite or the vector, if these are sensitive to RNA interference, thereby blocking parasite development. Abbreviations: dsRNA, Double-stranded RNA
The great advantage of this method over genetically transformed mosquitoes is that the transformed bacteria/fungi/viruses used may have the ability to colonise a range of different insect vector strains and even species. In contrast, with transgenic mosquitoes each strain or species may have to be transformed to prevent disease transmission. In addition, it is much easier to produce large numbers of transformed microbes than to generate sufficient numbers of transgenic mosquitoes [34]. Furthermore, the transformed microbe usually undergoes massive multiplication in the target insect vector [52] and may be passed horizontally as well as vertically from one generation to another [53].
A technique similar to—but not true—paratransgenesis involves the transfection of non-native, non-genetically transformed bacteria (or other microbes) to modify the microbiome in the insect gut, thereby altering host physiology and reducing vectoring ability. There are many examples of this process, such as Serratia marcescens-blocking Leishmania braziliensis, Trypanosoma cruzi and Plasmodium berghei in their vector insects [32, 54]. One example studied in some detail is Wolbachia pipientis (bacteria; family Rickettsiaceae) whose strains widely occur as intracellular pathogens in arthropods and nematodes and which can be inherited transovarially to spread rapidly through insect populations. Wolbachia infections in insects can cause incompatibility of the egg and sperm, leading to sterility, feminisation, parthenogenesis and male killing [50], and these outcomes can be used to control vectors. Thus, transfection of the Wolbachia wMelPop-CLA strain into Aedes aegypti reduces the life-span of the mosquito and its ability to vector chikungunya and dengue [51]. Inhibition of the parasite life-cycle can be achieved even when the mosquito strain is not naturally infected with Wolbachia. This inhibitory effect, however, is not universal since Aedes albopictus naturally infected with Wolbachia still effectively vectors chikungunya [55]. The difficulty in characterising the action of Wolbachia as paratransgenesis arises since the strains used may or may not be native to the vector insect, and only untransformed forms of Wolbachia are available to inhibit the life-cycles of parasites in different vector insects. Wolbachia is an intracellulatr endosymbiont, and to date genetic transformation has not been achieved and in vitro culture is only possible in a few cell lines [56,57,58]. Many recent articles and reviews on Wolbachia are available [59,60,61,62,63,64,65]; therefore, the present review is limited to paratransgenesis in which native microbes are transformed and transfected into vector insects.
Development, requirements and recent advances of the paratransgenesis technique
Development of paratransgenesis
Paratransgenesis was originally developed in the triatomine, Rhodnius prolixus, by Beard, Durvasula and colleagues [66,67,68,69,70,71,72,73] in an attempt to control the transmission of the protozoan parasite, Trypanosoma cruzi, the agent responsible for Chagas disease. This pioneering work provided a model for paratransgenesis research in other vector insects that transmit diseases not only to animals and humans but also to agricultural plants [74]. Briefly, the hindgut of R. prolixus contains very high concentrations of a Gram-positive, actinomycete bacterium, Rhodococcus rhodnii. Upon emergence of the R. prolixus nymphs, this organism is acquired in the first instars by coprophagy from the faeces of other members of the colony and is necessary for development of the insects to adults. The bacteria are well-placed to interact with T. cruzi as this parasite spends the last stages of its life-cycle in the hindgut of R. prolixus surrounded by R. rhodnii. The newly emerged nymphs are asymbiotic and can be maintained under sterile conditions and fed transformed or wild-type R. rhodnii in a blood meal using an artificial membrane feeder [70]. In the initial studies, the R. rhodnii were transformed with an Escherichia coli/R. rhodnii shuttle plasmid containing antibiotic resistance marker genes [66]; in later studies, however, more stable L1 mycobacteriophage integrative plasmids were used [75]. After the first instar R. prolixus are fed with transformed bacteria, they develop to sexual maturity at a rate similar to that of the controls fed with untransformed bacteria. In addition, transformed R. rhodnii could be detected in the gut for the 6.5 months of the experiment and also following successive moults [66, 70], thereby demonstrating that a transgenic symbiont could be introduced into a vector insect with no apparent cost to fitness or survival. Subsequently, a gene fragment for the trypanocidal, immune peptide, Cecropin A, from insects was inserted into the R. rhodnii symbionts and then aposymbiotic R. prolixus were colonised with these transformants or with wild-type control bacteria. Challenge of these two groups of R. prolixus fourth instars with T. cruzi resulted in 100% and 35% infection rates, respectively, for the control and experimental insects [69, 71]. The 35% of infected R. prolixus in the experimental group contained significantly reduced numbers of the final metacyclic forms of T. cruzi. These results provided both an innovative method for use in integrated pest management (IPM) programmes for controlling the triatomine vectors of Chagas disease in South and Central America, as well as a stimulus for developing this ingenious technique in other insect vector/parasite associations (Fig. 1).
The results of subsequent paratransgenesis research with other vector insects, including triatomines, are detailed in section Paratransgenesis in different groups of vectors of this review. However, some preliminary discussion on the requirements of and recent advances in the use of paratransgenesis are presented here to facilitate the successful employment of this technique.
Requirements for successful paratransgenesis
Requirements for successful paratransgenesis include:
-
i.
A culturable, symbiont or commensal bacterium (fungus or virus is occasionally used) should be present in the insect vector, be susceptible to genetic manipulation [76,77,78] and occupy the same body tissues as the pathogen in the host.
-
ii.
The microorganism ideally should colonise all instars during insect development throughout the life-cycle from first instars into adults. Most bacteria are lost during metamorphosis from larvae to adults, especially in mosquitoes, so that transstadial species such as Asaia in mosquitoes are ideal [79].
-
iii.
The microorganism should be non-pathogenic to humans and animals and capable of colonising a range of strains/species of mosquitoes or sand flies, etc. [34].
-
iv.
The ‘fitness’ of the genetically modified microorganism must not be compromised and its stability and normal functioning should be retained within the host vector [70].
-
v.
One or more effector molecules must be identified and then secreted by the recombinant microorganism to have the expected inhibitory effect on the parasite/insect vector interaction. The molecule must have no fitness cost to the insect vector [34, 80, 81].
-
vi.
There must be a way to facilitate the introduction and dispersal of the recombinant microorganism into wild vector insects under field conditions. Initial successes with semi-field trials have been reported in controlling triatomines with transformed R. rhodnii and mosquitoes with Asaia strains [53, 82].
-
vii.
Approval for use of the paratransgenesis technique from regulatory bodies and local populations must be sought. There are serious safety concerns about the release of genetically modified organisms in the field which will need to be addressed by environmental risk assessments. The risks of horizontal genetic transmission to the genomes of other organisms must also be minimised [34, 83].
Technical advances in the use of paratrangenesis
Most important innovative methods undergo improvements to increase efficiency and optimise the outcomes. This is most certainly true for paratransgenesis, with advances made in most stages, as outlined in the following sections, and shown in Table 1.
Analysis of microbiomes
Advances in molecular techniques beyond the 16S RNA gene method for the analysis of vector insect microbiomes have been made with, for example, high-throughput sequencing (HTS) commonly used for the complete analysis of all microbes in a sample [83]. The 16S RNA method uses just one gene for analysis while HTS fragments all the DNA in a sample, sequences these and then fits them together for analysis [84]. Thus, with HTS all groups of microorganisms in insect tissue samples can be identified; those selected for paratransgenesis must then be amenable to multiplication with traditional culture techniques. In addition, culturomics has recently been successfully introduced to identify previously unknown bacterial species in the vector gut microbiome. Basically, culturomics consists of multiple culture conditions combined with matrix-assisted laser desorption/ionisation–time of flight (MALDI-TOF) mass spectrometry or 16S ribosomal DNA (rDNA) amplification and sequencing [50, 85].
The choice of symbiotic microorganisms as recombinant candidates for the expression of effector molecules in paratransgenesis has also been extended from bacteria to include viruses and fungi, although the majority of studies have utilised bacterial symbionts [70, 83, 86]. Novel viruses and fungal genera have been identified in Culex pipiens by shotgun metagenomic sequencing, which is a HTS, PCR-independent technique, as well as by culture-dependent methods [87]. Wild mosquitoes are also commonly infected with insect-specific viruses belonging to several families, including the Densovirinae and Flaviviridae [86, 88], and these appear to suppress arbovirus infections in mosquitoes by superinfection suppression [51]. Work with mosquito densoviruses (MDVs) has demonstrated their potential use in paratransgenesis. MDVs are environmentally stable and colonise natural mosquito populations by vertical and horizontal transmission, and their host specificity is restricted to mosquitoes [89, 90]. They also have small genomes that are easily modified genetically to express foreign effector genes with potential for the transformation of target vector insects [90,91,92,93]. The recombinant viruses initially produced, however, were apparently replication defective and unable to undego secondary transmission due to the loss of viral capsid proteins essential for replication [90]. This problem has now been overcome using a microRNA (miRNA) expression system in which the recombinant MDVs are stable and self-replicating and induce silencing of mosquito genes [90]. In addition, the problem of large-scale production of the recombinant MDVs has been solved, allowing field testing experiments [94].
Fungi with long environmental survival times as spores and the ability to infect insects directly through the exoskleleton also have the potential for use in paratransgenesis. Mosquitoes, triatomines and sand flies have been shown to have extensive mycobiomes, although current knowledge of the interaction of these mycobiomes with vector insects and their infecting pathogen associations is very limited [83, 95,96,97]. Yeast species, such as Wickerhamomyces anomalus, have been isolated from both laboratory and wild colonies of Anopheles and are widespread in adult mosquito tissues, suggesting possible use in paratransgenesis [81, 98]. This potential has been confirmed with genetically modified yeast delivering double-stranded RNA (dsRNA) to a Drosophila sp. pest of soft fruits which, following ingestion, resulted in decreased locomotor activity and reduced egg-laying of the adult insects [99]. Another significant study was conducted on the entomopathogen, Metarhizium anisopliae, which was transformed to express anti-Plasmodium falciparum molecules, including various combinations of scorpion toxin (scorpine), artificial salivary gland and midgut (SMI) molecules and PfNPNA sporozoite-binding antibody. Transgenic Anopheles infected with M. anispliae expressing a combination of scorpine with SMI peptides resulted in > 98% inhibition of sporozoite levels in the salivary glands [100]. Subsequent to these studies, however, little progress seems to have been made in the use of fungi in paratransgenesis for the control of pathogen transmission by vector insects. This is surprising since entomopathogenic fungi can easily be mass produced and genetically transformed, do not infect vertebrates and have previously been used in the field to control insect pests. There would also be a synergistic effect since both the mosquito and the malarial parasite would be inhibited by the transformed fungus [101].
Transformation of symbionts
Once the symbionts have been selected, they are transformed to carry effector genes to inhibit the life-cycle of the pathogen in the vector insect or even in the vector itself. Various plasmid vector systems are usually used for this genetic transformation. For example, in studies of paratransgenesis in the vector insect, Rhodnius prolixus, L1 mycobacteriophage integrative plasmids inserted genes into the genome of the symbiotic bacterium, Rhodococcus rhodnii, to give highly stable constructs [77] One recent development has been the introduction of CRISPR-Cas (Clustered Regularly Interspaced Short Palindromic Repeats [CRISPR] with CRISPR-associated [Cas] endonuclease or enzyme) genome editing systems to transform the genomes of the insect gut microbiome. Not only can such systems transform bacteria by introducing one or more specific genes but it can also mediate gene silencing [102,103,104]. CRISPRs are derived from prokaryotes and include an endonuclease (Cas9) guided by a guide RNA (gRNA) to cut the chromosome at a specific site [32]. One advantage of this system is that the integration of the transgene is not only site-specific but also highly stable. The CRISPR technique has been applied successfully to edit an outer membrane gene (ompA) of a gut symbiont of Aedes aegypti to determine its role in biofilm formation in the vector gut [105], as well to transform several mosquito species to alter and drive specific genes into different generations [32, 106, 107]. The CRISPR system can transform microbes for paratransgenesis and also mediates gene silencing, resulting in more consistent and robust knockdowns with fewer off-target events than RNA interference (RNAi; see detailed comparative review on CRISPR and RNAi in [108]). RNAi knockdown techniques, however, have been invaluable in mosquitoes to investigate immune gene functions in relation to parasite interactions [83, 109]. In earlier studies, RNAi knockdown in Plasmodium was hampered by an apparent lack of appropriate RNAi machinery, but it is now possible by engineering two components, Argonaute 2 and a modified short hairpin RNA, into the parasite. These transgenic parasite lines, although not immediately transferable to the field, will be invaluable for studying Plasmodium gene function [110]. Thus, with both RNAi and CRISPR-Cas systems, it should be possible for multiple genes to be inhibited simultaneously and so prevent parasites developing resistance. The next step will be to inhibit parasite development within the insect vector using these techniques [83].
In addition, a modified form of paratransgenesis, termed RNAi-based paratransgenesis, has recently been introduced in which the transformed microbes deliver dsRNA instead of the usual effector proteins [74, 111,112,113,114,115,116] (Fig. 1). This method of delivery is ingenious as previously it was necessary to either inject or feed the target insects with the dsRNA, with both of these methods having significant disadvantages [117]. Injection is labour intensive, often kills many insects and induces immune/stress responses to potentially confuse interpretation of results, and the RNAi effect may be transient in long-lived species. Likewise, with feeding insects the dsRNA, the effect may be temporary and require repeating several times [74], although prolonged knockdown of the TsetseEP gene was achieved in Glossina by feeding dsRNA [118]. Using bacteria to express dsRNA has been successfully employed in the haematophagous triatomine, R. prolixus [74, 111], and the phytophagous crop pest, Frankliniella occidentalis [74]. In R. prolixus, an RNaseIII-deficient endosymbiont strain of Rhodococcus rhodnii had dsRNA expression cassettes stably incorporated into the chromosome and was used to successfully knock down Nitrophorin-1 and Nitrophorin-2. The knockdown phenotype produced colourless salivary glands in contrast to the cherry-red glands of the controls [74]. Likewise, a dsRNA expression cassette for Vitellin, responsible for producing approximately 80% of oocyte protein in R. prolixus, resulted in a significant 72.3% reduction in first instar-eclosed insects per adult insect per day. The transformed R. rhodnii not only persisted > 250 days in the R. prolixus gut with no apparent effects on insect fitness, but was also horizontally transmitted by coprophagy [74]. Also in R. prolixus, using E. coli expressing dsRNA, genes RHBP (Rhodnius heme binding protein) and CAT (catalase), involved in antioxidant activity and oocyte development were knocked down between 65 and 96%, respectively [111]. Finally, in the thrip, Frankliniella occidentalis, a similar strategy was adopted, except that alpha tubulin was targeted (tubulin alpha1), resulting in depletion of tubulin mRNA levels and significantly increased mortality of adults [74]. These studies indicate the real possibility of using symbiont-mediated RNAi to control both vectors of disease and agricultural pests. This technique has now been developed to mediate honey bee physiology and kill parasitic Varoa mites [116], and is being advanced to produce a mutant Asaia strain for RNAi-based paratransgemesis in Anopheles [115]. Future studies should be aware that using RNaseIII mutant bacteria has been shown to improve the delivery efficiency of dsRNA compared with normal transformed bacteria still producing RNAseIII [119].
Choice of effector molecules
The original inspirational studies on paratransgenisis in R. prolixus used the native endosymbiont, R. rhodnii, to deliver either a functional antibody fraction or Cecropin A, an insect antimicrobial peptide, as effector molecules against T. cruzi [66, 71]. Subsequently, numerous other effector proteins have been identified and used in paratransgenesis, with the majority associated with mosquitoes and against Plasmodium spp. (for more details see [81, 120,121,122,123] and section Mosquito microbiomes: native endosymbiotic bacteria in paratransgenesis of present article). In addition, a number of advances in use of effectors have been made. First, antimicrobial peptides are frequently used in paratransgenesis against various stages of Plasmodium; in order to facilitate this, Carter et al. [124] tested a range of 33 such molecules. These peptides were fed to Plasmodium-infected anopheline mosquitoes in the first 24 h of the sporogonic stage. Analysis identified seven peptides, mainly from bee and wasp venoms, that mediated significant killing of the parasites and had limited effects on mosquito fitness in terms of fecundity and longevity. It should be noted that this study involved feeding the peptides directly to infected insects rather than through secretion by transformed native symbionts. Second, studies have found that combinations of effector molecules are much more effective at killing parasites than single molecules (e.g. [75, 100, 120, 121, 124, 125]). Most of these studies, however, were with parasites mixed with effector proteins in vitro or with transgenic mosquitoes expressing combinations of proteins rather than by paratransgenesis with multiple effectors secreted by a single transformed symbiont. The outstanding paratransgenesis research of Fang et al. [100] and Wang et al. [120] with Anopheles gambiae and An. stephensi, respectively, however, describes the multiple simultaneous expressions of effector molecules by microbes. In An. gambiae, Metarhizium anisopliae was transformed to deliver scorpine as well as an [SM1]8:scorpine fusion protein, resulting in a 98% reduction of Plasmodium falciparum sporozoite counts [100]. Also, in An. stephensi, a Serratia strain of symbiotic bacteria (Serratia ASI) was discovered, capable of simultaneously expressing five anti-Plamodium effector proteins [120] (see also Pantoea agglomerans and Serratia in section Mosquito microbiomes: native endosymbiotic bacteria in paratransgenesis of present article). These combinations of proteins from transformed Serratia ASI or M. anisopliae were more effective at reducing parasite oocyst or sporozoite numbers, respectively, than those from symbionts producing just a single effector molecule [100, 120]. This is an important step forward since such combinations of anti-parasite effector molecules with various modes of action can be optimal for preventing the development of resistance by parasites. Third, one significant problem with transgenic symbionts released in the field is their potential loss of ability to compete with the microbiome already established in the gut of wild vectors [126]. For example, Asaia bogorensis colonises a range of vectors, including Ae. aegypti, Ae. albopictus and An. stephensi, and has been engineered to produce anti-Plasmodium effectors. Shane et al. [126] reasoned, however, that genetic modification of the bacteria may lead to a significant loss of fitness as a cost of the prodution of the effector protein, leading to lack of competiveness when released in the field. This problem was resolved by isolating blood meal-induced promotors (BMI) activated only during vector feeding on blood and exposure to nutrients [126]. Plasmids expressing the anti-Plasmodium protein scorpine under the control of the BMI promoters were constructed and transferred into Asaia sp. SF2.1 strain by electroporation. The Asaia BMI strains, in comparison to the constitutive scorpine-expressing control strain, had significantly increased maximum growth rates, enhamced ability to compete when co-cultured with wild-type Asaia and increased colonisation of mosquito midguts. The BMI strains also resulted in a significant reduction in oocyst numbers compared with the constitutive scorpine-producing control [126]. The authors hypthesised that for release in the field more than one effector protein should be expressed by the transformed symbiont to reduce chances of resistance. In addition, for the sake of stability, the effector genes should be inserted into the Asaia chromosome rather than carried on a plasmid (see CRISPR in section Transformation of symbionts of present article).
Tranfection into insect vectors
A significant problem for paratransgenetic control of disease vectors is the delivery of the transformed symbiont to a specific wild insect vector population under field conditions [127] (Fig. 2).

Spread of engineered microorganisms (EM) into wild mosquitoes. EM can be directly offered to winged adults through a baited trap or encapsulated and seeded or oviposited into water to contaminate aquatic juvenile forms. Females containing EM can also contaminate eggs laid on land, enabling vertical transmission. The selected bacterial species should preferably remain in the different stages of mosquito development or even be transmitted horizontally within this host. In this way, EM can remain permanently and cyclically in the environment
This process requires rapid spread of the foreign genes through the native population, without either fitness costs to the target vector or the occurrence of transfection of non-target insects. This technique could potentially be incorporated into IPM programmes. but problems associated with the release of GM organisms have yet to be fully resolved (see section Concluding remarks including safety and environmental concerns of present article). There have, however, been attempts to simulate natural conditions and investigate the potential of transfecting vectors in the wild. Some of these studies are more than 10 years old [73], with the major regulatory barriers to GM organisms presumably still not statisfied. The following are more recent pilot experiments more or less simulating natural conditions for transfection.
Mancini et al. [53] used large cages to study the horizontal and vertical transfection of Asaia sp.-transformed bacteria expressing green fluorescent protein (GFP), Asaiagfp, into laboratory-reared Anopheles stephensi and An. gambiae populations. Transfection occurred either by the release of paratransgenic male mosquitoes or from feeding on cotton pads soaked in sucrose plus 108 transformed Asaiagfp bacteria/ml. Transfection was monitored after 5, 12 and 20 days by fluorescence microscopy and PCR. The results showed the efficient horizontal spread of Asaiagfp into both An. stephensi and An. gambiae. For example, in An. stephensi, the release of paratransgenic males resulted in a 73% infection rate in 400 mosquitoes after 20 days. In addition, experiments on vertical and trans-stadial transmission in An. gambiae resulted in 78% of fourth instars and 44% of the newly emerged adults with Asaiagfp. In conclusion, this semi-field pilot study illustrates the feasibility of transfecting transformed bacteria into populations of mosquitoes [53].
Arora et al. [128] also used a simulated field study to address the problem of transfecting a pest insect, the glassy-winged sharpshooter, Homalodisca vitripennis (a hemipteran like the triatomines), with a transformed bacterium, Pantoea agglomerans, expressing a GFP. This insect is a vector of Xylella fastidiosa which is a bacterial pathogen of grapes and citrus fruits. The engineered P. agglomerans were microencapsulated in an alginate hydrogel and after ingestion by field-collected H. vitripennis, the bacteria colonised the foregut for up to 15 days. The bacteria were only released from the gel during the flow of plant sap into the foregut of feeding insects. More recently, it has also been shown that P. agglomerans can be transmitted horizontally between H. vitripennis and therefore may be self-sustaining [128].
Finally, Wang et al. [120] used laboratory cage experiments containing virgin female and male An. stephensi mosquitoes fed, respectively, with Serratia AS1−mCherry and AS1-gfp, to monitor how Serratia AS1 colonised and persisted in these mosquitoes. Subsequently, The results showed that all the offspring larvae and adults carried both fluorescent proteins so that the transformed Serratia AS1 spread through the whole mosquito life-cycle horizontally, vertically and transstadially. Additional experiments also showed that the bacteria persisted in multiple subsequent generations [120] (for more details, see section Mosquito microbiomes: bacteria of present article).
These studies are important as they describe the successful transfection and persistence of transformed bacteria into insect vector populations as well as the use of microencapsulation of engineered bacteria to limit their release and contamination of the environment (Fig. 2). These are significant steps in the evolution of paratransgenesis from the laboratory to the field [128, 129]. In addition, the results of semi-field trials provide information for formulating models of the efficacy of paratransgenesis and ways of improving the spread of transformed bacteria. For example, information on the distriburion of sugar baits could be used to help prevent a malaria outbreak and influence control policies [130]. On a cautionary note, however, the value of these semi-field pilot studies has been questioned since the insects tested are sometimes inbred laboratory strains and the doses of microbes used extremely high so the results obtained may not reflect the normal responses occurring in nature [131]. When cage trials are undertaken, it is also important to carefully assess the effects of genetic transformation on mosquito fitness in all developmental stages, including percentage of egg hatching. The above advances are summarised in Table 1.
Paratransgenesis in different groups of vectors
The majority of studies on paratransgenesis have been conducted using symbiotic or commensal bacteria for transformation (see section Analysis of microbiomes of present study). The composition of the bacteria in insects is highly dynamic and varies not only from vector species to species but also according to stage of development, sex, nutrition, habitat, geographical region of the insect and location in the insect [83, 97, 132,133,134,135,136,137,138,139,140,141]. The bacteria live in the intracellular or extracellular environment of the insect host and preferentially colonise the midgut and less frequently the salivary glands and reproductive organs [142, 143].
Mosquito microbiomes
There are over 3567 species of mosquitoes classified into 41 genera [144], but members of just three genera, Anopheles, Aedes and Culex, are responsible for the transmission of the majority of human diseases [9].
Studies on the microbiome composition of mosquitoes are most important since these have revealed the roles of the constituent microbes in the nutrition, physiology, immunity, metabolism, reproduction, longevity and even behavior of the host mosquitoes (e.g. [145,146,147,148,149]). In addition, the microbiome influences the relationship of the vector insect with infecting parasites and pathogens (e.g. [97, 120, 146, 149,150,151,152,153,154,155,156,157,158]). An excellent review on the interaction of the mosquito gut microbiota with the immune system and the pathogens is provided by Gabrieli et al. [159] who emphasises the importance of understanding this trilogy in order to maximise control strategies. Therefore, knowledge of the insect vector microbiome is vital for identifying microbes for use in paratransgenesis and for optimising the mass production for release of transgenic mosquitoes to control malarial parasites and arboviruses [34, 51, 83, 120, 160,161,162,163].
In the last decade, the realisation of the importance of the mosquito microbiome has resulted in over 300 publications on this topic [164]. This in turn has produced large amounts of data from many species in various physiological states and from different habitats, using alternative sampling and analysis techniques. This problem has made it difficult to compare the research results on mosquito microbiomes from different studies [51, 161, 164, 165]. Dada et al. [164] have therefore created a Mosquito Microbiome Consortium (www.mosquito-microbiome.org) as a repository for rationalisation of these data and to provide guidelines for conducting mosquito microbiome research to enhance collaboration. This Consortium focusses on four areas, namely: (i) sampling/experimental design; (ii) metadata collection; (iii) sample processing and controls; and (iv) data handling and analysis.
It is highly recommended to read this Consortium paper [164] as well as the publications of Romulo and Gendrin [157] and Rodríguez-Ruano et al. [165] for rationalisation of future research efforts and protocols. Consultation of the mosquito microbiome literature does, however, reveal some consistences in the the composition of bacterial species (see following subsections).
Mosquito microbiomes: bacteria
Bacteriomes of Aedes spp. and Anopheles spp. mosquitoes consist primarily of Gram-negative species, with as many as 98 genera described in anophelines [166]. Many of these bacteria are found in the midgut although the salivary glands and reproductive organs are also involved. In Anopheles culicifacies, the salivary glands are reported to contain more diverse microbial communities than the gut [142].
A detailed account of the bacteriome distribution in Aedes has been given by Scolari et al. [51]. Relatively few taxa, however, usually dominate, and these are often referred to as the core microbiota [156]. These may be highly variable depending upon the host stage and sex, the habitat, as well as whether the mosquitoes were laboratory-reared, field-caught and parasitised or not [149, 167,168,169],
The mosquito bacteriome is mainly composed of the Gram-negative phyla Proteobacteria and Bacteroidetes, but there are also representatives of the Gram-positive phyla Firmicutes and Actinobacteria. The Proteobacteria contain many species with great potential for paratransgeneisis experiments in mosquitoes and include the genera Asaia, Enterobacter, Pantoea, Serratia, Aerobacter, Aeromonas, Alicyclobacillus, Bacillus, Clostridia, Elizabethkingia, Escherichia, Flavobacterium, Geotrichum, Klebsiella, Lactobacillus, Micrococcus, Proteus, Pseudomonas, Shewanella, Spingomonas, Thorsellia and Wolbachia (e.g. [51, 83, 97, 132, 137, 140, 141, 149, 151, 160,161,162,163, 168,169,170,171,172,173,174,175,176,177,178,179]).
This list is incomplete, as shown by Tainchum et al. [179], who recorded five new genera in anophelines from Thailand, and by Nilsson et al. [180] who, working with Anopheles darlingi from the Amazon Basin, reported dominance of Escherichia/Shigella, Pseudomonas (all Proteobacteria) and Staphylococcus (phylum Firmicutes). Whether there are any differences in the microbiota of Aedes, Anopheles and Culex has also been the subject of study (e.g. [156]). It has been shown in a comparative study of the microbiome of field-collected Aedes, Anopheles and Culex carried out in the USA that there are similarities in the bacteria components in the gut [180]. Ecology seems to be important since different mosquito species from similar environments share core bacteria. The environment determines the nature of the food resources, such as the plants and nectar, as well as the composition of the microbiota at the breeding sites. The microbiota of the larvae will be acquired from the surrounding water, and a fraction will be retained by the adults following moulting (Fig. 2); the remainder will be modified following a blood or sugar meal or even parasitisation and co-occurrence and co-exclusion interactions in the microbiome with, for example, Wolbachia [135, 156, 175]. In fact, the mosquito gut has been described as a “selective eco-environment for its microbiome”, favouring enteric bacteria, such as the Enterobacteriaceae, with high redox capacities to manage the oxidative and nitrosative stresses from the digestion of the blood meal [181]. Finally, although genera of bacteria found in laboratory-reared and wild mosquitoes are similar, wild mosquitoes lose component microbiota within one generation of laboratory rearing [161].
Mosquito microbiomes: native endosymbiotic bacteria in paratransgenesis
Relatively few native bacterial species (Table 2) from the midgut of mosquitoes have been exploited for developing paratransgenesis for the control of disease and malaria in particular. These include Asaia, Pantoea, Serratia, Enterobacter, Escherichia, Chromobacterium and Pseudomonas.
Asaia
Asaia commonly infects mosquitoes via plant nectar and has been found in a large range of mosquitoes both from wild and laboratory strains. It is present in many Anopheles (Nyssorhynchus) species, including the important malaria vectors, An. stephensi, An. gambiae, An. fluviatilis and An. darling, as well as in Aedes aegypti, Ae. albopictus and the Culex pipiens complex [51, 163, 173, 182,183,184,185,186,187,188,189].
Asaia has great potential for development in paratransgenesis as not only is it widely distributed in mosquitoes but it also colonises the midgut, salivary glands and reproductive organs of both male and females. In addition, Asaia is horizontally and vertically transmitted, present in different stages of mosquito development and can be grown in culture and genetically manipulated (e.g. [138, 173, 190,191,192]). In addition to this potential of Asaia in paratransgenesis, wild Asaia strains can also inhibit the development of malarial parasites through the production of toxic proteins [183, 193], reduce the malarial parasite load by activation of the basic immune system of Anopheles after an infected blood meal [194] and inhibit competing Wolbachia infections [195]. Despite the potential of Asaia and other symbionts for the paratransgenesis control of malaria, there seems to be more emphasis on producing transgenic mosquitoes for release and control programmes rather than the potentially safer paratransgenesis alternative [10, 34, 122, 125].
Wang and Jacobs-Lorena [122] compiled a comprehensive table of possible anti-Plasmodium effector molecules, recognising four classes: (i) parasite killing; (ii) interaction with parasites; (iii) interaction with mosquito midgut or salivary gland epithelia; and (iv) manipulation of mosquito immune system. These molecules, together with those identified by Carter et al. [124], provide a useful choice for delivery by mosquito symbionts such as Asaia.
In order to determine more about the Asaia–mosquito interactions and test the suitability of Asaia for use in paratransgenesis with mosquitoes, Favia et al. [190] undertook a study on the kinetics of infection of Asaia in An. stephensi. Analysis of infected mosquitoes fed with GFP-tagged Asaia (Asaiagfp) showed that the bacteria colonised the female gut and salivary glands, the same compartments occupied by the malarial parasite during development. In addition, the larval gut and adult male reproductive system were massively invaded. Therefore, Asaia could potentially be orally and venerally transmitted, and, as reported previously, passed vertically from mother to offspring [190]. The bacteria remained in the adults throughout their lives and could be transmitted using simple sugar solutions. Capone et al. [196] also reported in An. stephensi that Plasmodium berghei genetically modified to express GFP (PbGFPcon) induced a host immune response but with no adverse effect against the midgut native population of Asaia. In fact, 2 days after PbGFPcon infection, the Asaia were enhanced by tenfold in mosquitoes. Other assays using Asaiagfp and AsaiaDsRedshowed the co-localisation of the bacteria with Plasmodium berghei (PbGFPcon) in the salivary glands and midgut, an optimal situation for reducing the vectorial capacity of the mosquito with transformed Asaia releasing anti-Plasmodium factors [196].
Subsequently, Mancini et al. [53] undertook a semi-field pilot study of the horizontal and vertical transfection of Asaiagfp into laboratory-reared Anopheles stephensi and An. gambiae populations and confirmed the efficient transmission of the bacteria in both species (see details in section Tranfection into insect vectors in present article). Thus, the feasibility of transfecting transformed Asaia into populations of mosquitoes was confirmed.
Successful experiments modifying Asaia to secrete heterologous proteins into the An. stephensi midgut and inhibit P. berghei were first reported by Bongio and Lampe [193]. Asaia bogorensis were genetically screened, and an efficiently secreted siderophore receptor protein was fused with the antiplasmodial gene scorpine or with an anti-Pbs21 scFv-Shiva1 immunotoxin. These Asaia strains were fed to mosquitoes that were then challenged with a P. berghei-infected blood meal; 2 weeks later mosquitoes were dissected and oocyst numbers on the midgut counted. Significant reductions in oocyst numbers occurred in both Asaia strains compared with the controls (P < 0.0001 and < 0.0006, respectively, for scorpine and the immunotoxin transformants) [193]. More recently, however, Shane et al. [126] reasoned that genetic modification of the bacteria may lead to a significant loss of fitness and competiveness in the field. These researchers therefore constructed Asaia BMI strains (bacteria with blood meal-induced promotors). These strains, in comparison to the constitutive scorpine-expressing controls, showed significant increases in maximum growth rates, in the ability to compete with wild-type Asaia, in the colonisation of mosquito midguts and in the inhibition of oocyst numbers [126] (see details in section Choice of effector molecules of present article).
Pantoea agglomerans (= Enterobacter agglomerans)
Pantoea agglomeratus was the most prevalent of 20 genera of symbiotic bacteria reported in wild-caught An. gambiae and An. funestus mosquitoes from Kenya and Mali [170]. There are also other reports of P. agglomeratus in anopheline, Aedes and other culicine mosquitoes from around the world [51, 197,198,199]. In addition, this bacterium is present in laboratory strains of Anopheles stephensi, An. gambiae and An. albimanus [200, 201]. Pantoea agglomeratus has been developed for potential paratransgenesis to prevent the transmission of malaria in mosquitoes [81, 114, 202], and of plant diseases and pests in agriculture crops [74, 129, 203] (see section Tranfection into insect vectors of present article). Pantoea agglomeratus has great potential for use in paratransgenesis as it naturally infects mosquitoes, resides in the insect midgut together with infecting malarial parasites, can be cultured and labelled with GFP for following the dynamics of infection, multipies 200-fold following ingestion and, most importantly, can be transformed to secrete anti-Plasmodium effector molecules [121].
The classic paper of Wang et al. [121] describes the use of the E. coli HlyA secretion system to separately transform P. agglomeratus strains with eight anti-Plasmodium effector molecules. The expression and secretion of each of the effector proteins by the recombinant P. agglomeratus were confirmed by Western blotting. To test the effects of the transformed bacteria on infection of An. gambiae and An. stephensi by Plasmodium falciparum or P. berghei, the bacteria were fed to the mosquitoes on cotton pads soaked with bacteria suspended in 5% sucrose. After 32 h, the mosquitoes were given an infected blood meal, and 8 days later numbers of oocysts formed were counted. Five of the effector proteins secreted by the transformed P. agglomeratus significantly inhibited parasite development by up to 98% for scorpine or (EPIP)4 (Plasmodium enolase–plasminogen interaction peptide). Combinations of two types of effectors were no more effective at parasite inhibition than individual proteins; this, however, does not consider possible enhanced resistance of P. agglomeratus protein combinations to parasite mutation and evolution. The importance of this study is that the engineered P. agglomeratus were equally effective at inhibiting malarial parasites in both An. gambiae and An. stephensi so that any reproductive or behavioural barriers that may exist between isolated vector populations in the wild will not affect paratransgenesis [121]. Progress in the widescale use of P. agglomerans for mosquito control has not advanced rapidly since the Wang et al. paper [121], probably as a result of the unresolved problem of driving the bacteria into wild mosquitoes [120] (see section Concluding remarks including safety and environmental concerns of present article) and the continued resistance to the release of engineered microorganisms into the environment. In addition, reports of P. agglomerans causing secondary human infections in bones and joints as well as pathogenic strains in some crops have to be considered [204, 205], although it is unlikely that these pathogenic bacteria are the same as those isolated from insects.
Serratia
Serratia spp. have been widely reported in the midguts and tissues of Anopheles, Aedes and Culex mosquitoes as well as in many non-vector insect orders [206, 207]. Interest in Serratia has previously been centred around the potential use of these bacteria for controlling the malarial parasite in the mosquito host (e.g. [208]). This Plasmodium-inhibitory activity of Serratia spp. has been shown to result from multiple mechanisms, including the upregulation of the mosquito immune system [209] by the direct production of anti-malaria factors by the bacteria themselves [157, 209,210,211], and by blocking ookinete penetration through the vector midgut epithelial cells [210]. The possibility therefore exists of transfecting mosquitoes with specific strains of Serratia to control malaria although much additional work is required and Wolbachia-based strategies have been given priority at the present time [62,63,64]. However, as mentioned earlier in this article, the use of Wolbachia in paratransgenesis has not been developed so far as the bacterium cannot be genetically transformed and is difficult to culture (it is an obligate intracellular symbiont) [56, 57, 212]. Reveillaud et al. [213], however, reported Wolbachia from four wild Culex pipiens mosquitoes carrying a plasmid (pWCP), indicating that future paratransgenesis utilising Wolbachia may be possible.
The potential use of Serratia for paratransgenesis has also been recognised [121, 122, 211, 214, 215]. Wang et al. [122] previously engineered natural symbiotic Pantoea agglomerans to secrete anti-Plasmodium effector molecules (see section Pantoea agglomerans (= Enterobacter agglomerans of present article) but failed to address the problem of infecting wild mosquito populations. These researchers then discovered, in Anopheles stephensi, a strain of Serratia called AS1 which has no fitness costs following engineering to produce anti-Plasmodium effectors in An. stephensi or An. gambiae [122]. Using fluorescent markers incorporated into the bacteria, the colonisation of the mosquitoes by Serratia AS1 was studied in laboratory cage experiments. In just one mosquito generation, AS1 was venerally transmitted horizontally from males to females during mating and then vertically to the offspring. It also survived larval metamorphosis to multiply in the mosquito midguts and other organs for multiple generations. The transformed Serratia AS1, producing multiple anti-Plasmodium effectors, were also fed to mosquitoes and inhibited the Plasmodium falciparum life-cycle [121]. Koosha et al. [215] also used Serratia AS1 labelled with mCherry fluorescent protein to study the acquisition of bacteria by arthropod vectors, including An. stephensi, Culex pipiens, C. quinquefaciatus and C. theileri. Subsequently, all adult mosquitoes took up the bacteria from the host skin during blood-feeding and from the water when larvae. The larvae then transferred them to the adults transstadially and these finally returned them back to the water during egg-laying (Fig. 2).
More recently, Huang et al. [134, 162] have addressed possible regulatory concerns about the release of engineered bacteria into the environment and any uncertain consequences that might occur. They have designed a self-limiting paratransgenesis using Serratia marcescens AS1 and An. stephensi. In this system, plasmids were used to transform Serratia AS1 bacteria, but these plasmids were lost in 130 generations so that the bacteria returned to wild type. Thereby, the plasmids were lost in three mosquito generations. Equally important, for satisfying regulators, there was no evidence, following feeding of plasmid-transformed AS1 to vector insects or their incubation in culture with high concentrations (1012) of E. coli or P. agglomerans, for horizontal transfer of plasmid genetic material to other bacteria (Table 1) [134, 162].
Enterobacter, Escherichia, Chromobacterium, Elizabethkingia and Pseudomonas
Apart from Asaia, Pantoea and Serratia described above, Enterobacter, Escherichia, Chromobacterium, Elizabethkingia and Pseudomonas are examples of other members of the phylum Proteobacteria with potential use in paratransgenesis but for which less published information is available. Enterobacter infections in Anopheles arabiensis and An. gambiae, without engineering, were shown to block Plasmodium falciparum parasites [216, 217]. Also, following an infected bloodmeal, Enterobacter cloacae rapidly colonised the midgut of An. stephensi and the bacteria were amenable to transformation but only weakly transferred from larvae to adults so of no use for multigeneration recycling [218]. Escherichia coli was transformed and shown to inhibit Plasmodium berghei in An. stephensi but the effect was suboptimal, the effector molecules stuck to the bacterial surface and the E. coli strain used survived poorly in the mosquito gut [81]. Chromobacterium isolated from the midgut of Aedes and Anopheles mosquitoes has been shown to have both anti-Plasmodium and anti-dengue virus activity in vitro and to kill Anopheles coluzzii after infective feeding [153, 219, 220]. These toxic properties are probably at least partially due to a secreted protease, suggesting that the bacteria could be engineered to produce this effector in the midgut of mosquitoes [220].
Elizabethkingia is also common in anopheline mosquito microbiomes from western Thailand [179], can be transmitted transstadially and has been transformed to re-infect Anopheles mosquitoes [221]. It is, however, a potential human pathogen with resistance to some antibiotics so caution would be required [222]. Similar pathogenic concerns exist for Pseudomonas isolated from the common Asian vector, Anopheles culicifacies [160] and from Culiseta longiareolata [223]. It is present in both larvae and adults and so may be transstadial and, depending upon the species, may be a possible candidate for paratransgenesis.
Mosquito microbiomes: viruses and fungi
The choice of symbiotic microorganisms for developing paratransgenesis in mosquitoes has also now been extended from bacteria to include viruses and fungi, although the majority of studies have utilised bacterial symbionts (e.g. [83, 86]). Details of potential viral and fungal candidates for paratransgenesis have been discussed in previous sections (see section Analysis of microbiomes of present article). Gurung et al. [224] believe that focussing attention too much on bacteria in the microbiome and ignoring the other microbial components, such as the fungi, viruses, archaea and protozoans, may hamper full understanding of the true impact of the microbiome on the insect pest. This is just as likely to apply to the effect of the microbiome on invading parasites.
Mosquito microbiomes: RNAi-based paratransgenesis
This is a relatively new technique in vector insects in which the transformed symbionts deliver dsRNA instead of the usual effector proteins to silence or knock down a specific host or even parasite genes (for more details, see section Transformation of symbionts of present article). In addition, the use of RNAi for the control of mosquitoes and malarial parasites is growing [115] although technical difficulties exist. For example, both Aedes aegypti and Ae. albopictus contain 10 dsRNases which would rapidly degrade any dsRNA in the gut lumen [225]. Subsequent dsRNA knockdown of two key dsRNases resulted in a high efficiency of gene knockdown by dsRNA targeting a cyan fluorescent protein (CFP) reporter gene given by feeding [225]. Another way to enhance the survival of the dsRNA in the insect would be to use symbiotic bacteria to both protect and produce the dsRNA rather by feeding or injecting naked dsRNA [74, 114, 226].
For a summary of this section, see Table 2.
Triatomine microbiomes
There are approximately 152 described species of triatomine bugs, of which 67 occur in Brazil [227]. About half of these species can carry Trypanosoma cruzi, the causative agent of Chagas disease. This disease also induces chronic inflammation of the heart, colon and nervous system, and the parasite DNA can undergo vertical transmission to the progeny of mammals [228].
There has been an increased interest in the microbiome of these insects since details of the roles of the component bacteriome in the host physiology and interactions with the flagellate parasite, T. cruzi, were revealed (e.g. [54, 229,230,231,232,233,234,235,236]). Resistance to conventional insecticides also stimulated research on the triatomine microbiome [237], resulting in the introduction of paratransgenesisis as an alternative control technique, for the first time in vector insects, in Rhodnius prolixus (e.g. [69]).
The expansion in this research area with triatomines was also mediated by the application of molecular techniques, including high-throughput 16S rRNA and, more recently, next-generation sequencing and bioinformatics, to identify most members of the microbiome (e.g. [233, 234, 238,239,240,241,242,243,244,245,246,247,248,249,250,251,252]). These studies have looked at the microbiomes of varying numbers of wild and laboratory-reared triatomine species with and without parasites. In addition, these insects were from different geographical regions and ecological niches, at various developmental stages, and involved different feeding regimes, sexes, physiological states and tissues, utilising alternative sampling and analytical techniques. Therefore, and similar to the situation in mosquitoes (see section Mosquito microbiomes in present article), generalisations have been difficult to make. Fortunately, Duarte Silva et al. [253] and Salcedo-Porras et al. [234] have recently analysed and rationalised the results of some of these studies in detail although many contradictions still exist. In addition, Brown et al. [250] designed their research to eliminate some of these variables by, for example, using five wild Triatoma species sampled from the nests of white-throated woodrats in which all five instars plus adults could be found occasionally, as well as other species, all feeding on the same blood source. Some basic but not universal conclusions that can be drawn from these papers on the triatomine microbiome are as follows:
-
i.
Most triatomines have a low diversity of bacterial genera in comparison with other insects, but variability exists between species even when they originate from identical environments, such as the same nest [245].
-
ii.
The triatomine microbiome, which shows similarities to other vector insects [246], contains members of the Gram-negative phylum Proteobacteria (e.g. Serratia, Enterobacter, Pantoea, Acinetobacter, Arsenophonus, Pseudomonas and Wolbachia) and the Gram-positive phylum Actinobacteria (including Rhodococcus, Nocardia, Dietzia, Gordonia, Corynebacterium and Mycobacterium), which together make up 20–50% of the microbiome. In addition, Gram-positive Firmicutes (20%; e.g., Enterococcus, Staphylococcus, Bacillus) and Gram-negative Bacteroidetes (e.g. Proteiniphilum; 5–7%) are also present [234]. However, in only three species of triatomines were mutualistic symbionts identified, all Actinobacteria [254].
-
iii.
Many Proteobacteria, but particularly the Enterobacteriales (e.g. Arsenophorus, Serratia and Enterobacter) and Corynebacteriales (e.g. Rhodococcus, Nocardia, Dietzia, Gordonia, Corynebacterium and Mycobacterium), are present in multiple triatomines.
-
iv.
Similar changes occur in the microbiome in wild triatomines throughout development and from one gut compartment to another. These involve a change from high microbiome diversity to low diversity from first instars to adults which are often dominated by a single bacterial genus, including Dietzia, Mycobacterium or Proteiniphilum [245].
-
v.
Wild insects naturally infected with T. cruzi have a more diverse microbiome than uninfected wild insects or infected or uninfected cultured insects [164, 241], but see [244, 247].
-
vi.
Rhodnius spp. and Triatoma infestans are the only triatomines in which Wolbachia has been reported in both wild and laboratory populations [234, 249, 252].
Some of the above bacteria, and many more reported in the papers cited previously, would be good candidates for paratransgenesis, assuming that they can be cultured, are non-pathogenic for humans or animals and can be genetically manipulated with no adverse effects on their stability or fitness or on the host vector. Serratia, Pantoea and Enterobacter have already been tested in mosquitoes (see section Mosquito microbiome: native endosymbiotic bacteria in paratransgenesis of present article) as have Corynebacterium, Escherichia and Rhodococcus in triatomines (see section Transformation of symbionts of present article). Another factor in choosing bacteria for paratransgenesis is to select a species with high GC-content since, in the triatomine gut, bacterial species with high GC-contents have been shown to outcompete those with low GC-content [242].
Triatomine microbiomes—native endosymbiotic bacteria in paratransgenesis
The pioneering steps in the development of paratransgenesis were made with the triatomine, Rhodnius prolixus, utilising the genetically transformed actinimycete bacterium, Rhodococcus rhodnii, to deliver the trypanolytic antimicrobial peptide (AMP), cecropin A [66,67,68,69,70,71, 77]. The target of this peptide was Trypanosoma cruzi (for details see section Development of paratransgenesis of present article). The use of cecropin A was a successful proof of concept study and led to further experiments with other effector molecules in order to improve both the efficiency of parasite killing and reduce the likelihood of resistance developing (Tables 1 and 3). Several other single AMPs tested in vitro killed T. cruzi, but when AMPs were combined, for example, apidaecin with cecropin, melittin or magainin, the results with all pair-wise combinations achieved 100% lethal concentration (LC100) levels, in contrast to the single AMPs [80]. Strains of R. rhodnii have been transformed in vitro to produce these AMPs although the results of in vivo experiments with T. cruzi have not appeared. Instead, single-chained antibodies and recombinant β-glucanase have been developed as effector molecules against T. cruzi [70, 72, 255, 256] (Table 3).
Durvasula et al. [72, 255] and Hurwitz et al. [82] have proven the feasibility in R. prolixus and Triatoma infestans of the expression and secretion by engineered symbionts of functional fragments of the murine three-domain antibody (rDB3) capable of recognising and binding to progesterone. For this, the genetically engineered symbionts R. rhodnii and Corynebacterium sp., respectively, for R. prolixus and T. infestans, expressed and secreted functional fragments of rDB3 into the insect gut. The recombinant strains of R. rhodnii and Corynebacterium sp. were added to the blood meal of aposymbiotic first instar nymphs and shown to synthesise and secrete rDB3 for 6 months of the study [72, 82, 255]. Subsequently, small antibody molecules were produced against the sialyl-Tn and sialyl-(le)a surface glycans of T. cruzi [257] and shown by confocal microscopy to specifically bind to fixed T. cruzi epimastigotes [76].
Regarding recombinant β-glucanase as an effector molecule in paratransgenesis, the surface of T. cruzi is covered in a layer of mucin-like glycoproteins that are probably essential for the in vivo development of the parasite by mediating its binding to the triatomine midgut and hindgut cells [258, 259]. Jose et al. [256] concluded that disruption of the surface glycocalyx of T. cruzi would therefore inhibit the development of the parasite. To prove this, R. rhodnii was transformed to express β-1,3-glucanase, as this protease was previously shown to efficiently promote cells lysis in T. cruzi [76, 256]. Jose et al. [256] inserted the complementary DNA (cDNA) encoding the Oerskovia xanthineolytica β-1,3-glucanase gene (i.e. Arthrobacter luteu strain 73–14) into plasmid pRrExpA used for manipulation of R. rhodnii. In vitro assays performed with T. cruzi incubated in culture medium together with the recombinant R. rhodnii showed a more than 80% inhibition of parasite growth. The results proved the efficiency of the recombinant bacteria-expressed β-1,3-glucanase in lysing T. cruzi cells. Therefore, recombinant β-1,3-glucanase represents a valuable additional effector molecule for paratransgenesis against T. cruzi in its triatomine hosts.
These studies show that it is potentially possible to produce effector molecules targeting a range of different sites in T. cruzi to reduce the likelihood of the parasite becoming resistant.
A significant problem for paratransgenetic control of diseases in insect vectors is the delivery of transformed symbionts to specific wild insect vector populations in the field [127]. In the case of the triatomine, R. prolixus, this problem is readily solved with coprophagy spreading the transformed symbionts naturally to the whole population. The newly emerging R. prolixus nymphs are aposymbiotic (devoid of gut symbionts) but soon become infected from the surrounding faeces produced by the whole colony. To study transgenesis in simulated field conditions, Durvasula et al. [73] used large cages containing local Guatemala dirt and thatch with panels impregnated with CRUZIGUARD, a paste containing transformed R. rhodnii suspended in sterile phosphate-buffered saline plus guar gum powder. Newly emerging first instar R. prolixus from eggs of field-caught insects were housed in the cages, and guts were sampled at the third and fifth instar and adult stages and tested for transformed R. rhodnii. Approximately 56% of the experimental insects contained the transformed bacteria to the exclusion of other competing bacteria in the environment [73]. In addition, when nymphs were allowed to develop for 9 months in the cages, approximately 50% of adults were shown to contain transformed R. rhodnii. This technique could potentially be used along with insecticides to prevent reinfestations of homes.
Triatomine microbiome: RNAi-based paratransgenesis
It is significant that it has been nearly 30 years and > 20 years, respectively, since the pioneering works of Beard et al. [66] and Durvasula et al. [71] were published on paratransgenesis in R. prolixus and, although significant advances have been made, approval for use in the field has yet to be obtained.
Recent work on the use of symbiotic bacteria to deliver dsRNA for knockdown of specific genes in triatomines represents a significant step forward (see section Transformation of symbionts in present article) [74, 111, 113, 260]. This technique has also been adapted for development in mosquitoes and may help to satisfy the regulatory process for the release of transgenic bacteria in the field (see section Concluding remarks including safety and environmental concerns of present article).
For a summary of the above, see Table 3.
Tsetse fly microbiomes
Tsetse flies (genus Glossina) are viviparous with 30–33 species and subspecies having been described [58]. These are usually divided into the Morsitans, Palpalis and Fusca groups containing various species and subspecies which are particularly important medically and economically due to transmission of African trypanosomes [58]. African trypanosomiasis affects both people and their livestock (e.g. [261, 262]). Cases of human sleeping sickness rapidly declined from 1997 to 2019, with many countries reporting no new cases for the last decade [14]; however, there is a constant risk of re-emegence from animal and human reservoirs. There are also no vaccines for sleeping sickness, and chemotherapy is both expensive and toxic, and the parasites are showing increasing resistance [262, 263].
A number of studies have been made of the tsetse microbiome [264,265,266,267,268,269,270,271,272,273,274,275,276,277,278,279]. The results indicate that tsetse flies host a large range of bacterial species, often including four maternally transmitted endosymbiotic bacteria present in both wild and laboratory-reared flies (e.g. [269, 274]), namely Wiggleworthia glossinidia, Sodalis glossinidius, Wolbachia and Spiroplasma. Wiggleworthia glossinidia occurs intracellularly in bacteriocytes in the anterior gut to produce supplements for tsetse nutrition and often dominates to form 34.5–99.8% of the microbiome [272]. Sodalis glossinidius is present in the midgut, muscle, fat body, salivary glands and milk glands, while Wolbachia is found in the ovaries, with both varying greatly in incidence (e.g. [264, 269, 274, 280]). Spiroplasma is a more recently discovered transovarially transmitted endosymbiont present in the Palpalis group and culturable in vitro [281,282,283]. In addition, the tsetse microbiome has a diversity of commensal bacteria from the environment, which usually account for < 1% of the bacteriome [269, 273]. The number and incidence of these bacterial species vary from one study to another depending both on the methodology used and the species of tsetse fly sampled (e.g. [272]). For example, of the 103 species of bacteria described in Glossina palpalis palpalis by Jacob et al. [272], Gram-negative bacteria predominated as did the phylum Proteobacteria (97% of isolates), with members of the phyla Bacteroidetes, Actinobacteria and Firmicutes also represented and the microbiome showing some resemblance to those of Anopheles and Aedes. In contrast, with the Glossina pallidipes microbiome, Malele et al. [269] reported that of 113 isolates, the descending order of prevalence was Firmicutes (86.6%), Actinobacteria (7.6%), Proteobacteria (5.5%) and Bacteroidetes (0.3%). Examples of bacteria identified in Glossina microbiomes include the genera Bacillus, Serratia, Pantoea, Acinetobacter, Arthrobacter, Enterobacter, Enterococcus, Providencia, Sphingobacterium, Chryseobacterim, Exiguobacterium, Lactococcus, Staphylococcus, Pseudomonas, Spiroplasm and Xylella (e.g. [268, 275, 278, 283, 284]).
Tsetse fly microbiomes: native endosymbiotic bacteria in paratransgenesis
The four maternally transmitted endosymbiotic bacteria, Wiggleworthia, Sodalis, Wolbachia and Spiroplasma, are the dominant symbionts in Glossina and are transmitted maternally so that these were the natural candidates for developing paratransgenesis in tsetse flies. Of these symbionts, however, Wigglesworthia cannot be cultured, Wolbachia is not genetically transformable or easily cultured and Spiroplasma is a recent discovery. Thus, to date, Sodalis, which can be both cultured and transformed, has been utilised for paratransgenesis-related experiments in Glossina [13, 261, 264, 270, 285,286,287,288,289,290,291,292,293,294,295]. There is also evidence that some Sodalis genotypes can favour the establishment of trypanosme infections in tsetse flies by inhibiting the trypanocidal activity of the Glossina midgut lectin (e.g. [269, 274, 296,297,298]), although this varies with species, location and study [271, 276].
As a first step in developing paratransgenesis in Glossina, Cheng and Aksoy [287] studied the transmission of S-symbionts (presumably Sodalis) to the vector progeny. For this, they injected transformed S-symbionts expressing GFP into the hemolymph via the thorax of mated female G. m. morsitans and collected the F1 and F2 progeny. The gut tissues of the progeny were sampled and analysed for S-symbionts by PCR amplification using GFP-specific primers. The progeny haemolymph was also cultured and tested for the presence of GFP-expressing symbionts. Both techniques detected the transformed S-symbionts in the F1 and F2 flies to confirm the vertical transmission from mother flies. The presence in the milk glands of fluorescent recombinant symbionts also indicated that the route of transmission was from the haemolymph to the intrauterine larvae via secretion of these glands [287]. This work indicated two important factors necessary for successful paratransgenesis in tsetse flies: (i) the vertical transmission of the symbiont from the mother to the progeny and (ii) the ability of the transformed symbiont to express the heterologous gene effectively and stably in the insect vector [287].
An additional study advancing paratransgenesis in Glossina is that by Aksoy et al. [288] who describe why Sodalis is well-suited for paratransgenesis in tsetse flies since it occurs in the gut together with the trypanosomes, can be cultured, is resistant to trypanocidal peptides and can be genetically transformed and transmitted to the progeny.
Transformation in Sodalis has previously been performed with plasmids [286, 289,290,291]; however, this may not be optimal for field experiments as plasmid maintenance may require constant selection. This possibility has been recognised by both Aksoy et al. [288] and De Vooght et al. [291] who have utilised alternative methods. Aksoy et al. [288] undertook transformation using a piece of non-replicating circular DNA, with a sequence homologous to the desired chromosomal loci, which allowed transgenic symbionts to be maintained without selection. De Vooght et al. [291] also used the chromosomal expression of a reporter gene under the control of a native or a heterologous constitutive promoter. More recently, it was also discovered in Sodalis glossinidius that conjugation can be used as a DNA delivery method to conduct forward and reverse genetic experiments [299].
In another fundamental study providing key background information for paratransgenesis in tsetse flies, reciprocal swopping of Sodalis populations between G. fuscipes fuscipes and G. morsitans morsitans flies had no dentrimental fitness effects compared to the wild-type flies in terms of fecundity and longevity [270]. For these experiments, newly emerged adult flies were fed with blood plus antibiotic to clear the bacteria and then injected with the Sodalis strain from the other Glossina species of the pair, i.e. reciprocal transinfection. In these flies, the bacteria were also successfully transmitted to their progeny. These results indicate that in the field it would be possible to simultaneously control African trypanosomatid transmission by different Glossina species with a single recombinant strain of Sodalis expressing anti-parasitic effectors [270].
The next step in paratransgenesis is to identify effector molecules produced by the transformant symbionts and capable of expression and killing trypanosomes in Glossina without loss of fitness of the symbiont or the host insect. Glossina attacin is one possibility as it can kill trypanosomes both in vitro and in vivo without affecting Sodalis [300]. The first report, however, of the successful use of an anti-trypanosome effector molecule expressed in vitro and in vivo in Glossina was of a single domain antibody (Nanobody® molecules), Nb_An33, targeting conserved epitopes of the variant surface glycoprotein (VSG) of Trypanosoma brucei [290,291,292, 301].
For this work, a FliCpelBNb46fliC plasmid-based Sodalis strain was initially produced that expressed the trypanolytic nanobodies. This system was shown to be highly stable in vitro after 27 generations; therefore, the ability of the recombinant Sodalis (recSodalis) to colonise the G. morsitans morsitans tissues after intrathoracic injection was assessed. In order for the recSodalis to succesfully colonise the flies, it was first necessary to remove the wild-type Sodalis present in the recipient Glossina with streptomycin [290]. The recSodalis also persisted at high densities in the thorax and gut tissues for up to 28 days without affecting the population of Wigglesworthia, an essential Glossina endosymbiont, or the fecundity of the flies. Furthermore, the recombinants were also transmitted to the F1 progeny, but at only very low levels. Finally, nanobody concentrations were quantified over time with a VSG-binding enzyme-linked immunosorbent assay; functional Nb_An33 was found to accumulate in the haemolymph and thorax, indicating the expression of the injected transgene [290]. It was calculated that the levels of nanobody produced would probably be sufficient to deal with the average parasitiaemia of 103 T. brucei in cattle.
The above results are very encouraging for the development of paratransgenesis in tsetse flies although the transmission levels to the F1 progeny were very low. This could be due, as mentioned above, to instability of the plasmid and the need for a more stable transformation system. De Vooght et al. [291], therefore used chromosomally GFP-tagged recSodalis to colonise various tissues of tsetse flies and follow their transmission to the F1 progeny using different infection procedures. Injecting adults intrathoracically resulted in high-density colonisation of the tissues but limited infection of the reproductive organs (milk glands, etc.) and no vertical transmission to the progeny. Oral feeding of Glossina with recSodalis also failed to infect either the adults or the offspring. Finally, injection of the third instars gave stably infected adults and subsequent vertical colonisation of the next generations of flies. Apparently, in the larvae, certain invasion and motility genes are upregulated, such as invC and fliC and motA, and these may be required for vertical transmission (e.g. [302]).
These studies are important steps forward in the development of paratransgenesis in tsetse flies. In the future, populations of the resistant recSodalis-infected tsetse flies might be driven into suscepible field populations utilising the cytoplasmic incompatibility induced in flies by Wolbachia infections [288]. This possibilty has been modelled by Gilbert et al. [293] with human African trypanosomes that could potentially be eliminated over a 25-year period if colonisation by Wolbachia had minimal fecundity or mortality impacts on tsetse flies. The chance of recombinant Sodalis vertical transmission was also > 99.9%. In addition, control of African trypanosomiasis coud be mediated by paratransgenesis in tsetse flies combined with the the sterile insect technique, as advocated by Demirbas-Uzel et al. [294].
Sand fly microbiomes
About 500 sand fly (phlebotomine) species are known, of which more than 90 transmit leishmaniasis. The main vectors of human leishmaniasis are species and subspecies of Phlebotomus in the Old World and Lutzomyia in the New World [12]. East Africa, Brazil and India are particularly affected by visceral (fatal) leishmaniasis. Sand flies also vector several pathogenic viruses, including phleboviruses causing encephalitis, meningitis and haemorrhagic fever [303, 304]. Like mosquitoes, the female sand fly needs blood for egg development and transmits the pathogens during feeding. There are > 20 species of Leishmania, with most infected people showing few symptoms. In 2019, 97 countries were endemic with > 1 billion people at risk of infection and almost 1 million new cases of cutaneous leishmaniasis occurring annually [12]. An inactivated/killed Leishmania major vaccine with Bacillus Calmette–Guérin was developed but failed to protect against the disease [305]. Recently, scientists have characterised a new strain of Leishmania for use in a human infection model and are seeking volunteers for an initial trial [306]. Medicines for treatment of leishmaniasis may be limited in poorer countries, with toxicity and emerging resistance problematic and complications arising from HIV co-infections [12]. In addition, pesticide resistance by sand flies has also been detected [307]. The conclusion, therefore, is that new tools are also required to control this disease [308].
The main requirements for the development of paratransgenesis in sand flies are the same as those in other vector insects (summarised in section Requirements for successful paratransgenesis of present article), and have been reviewed for phelbotamines by Wijerathna et al. [309]. The prime requirement is the identification of appropriate bacterial species in which to develop the technique. There have been many studies on the commensal bacteria in Lutzomyia (e.g. [310,311,312,313,314,315,316,317]), and Phlebotamus (e.g. [97, 318,319,320,321,322,323,324,325,326,327,328]). Fortunately, the extensive reviews of sand flies by Telleria et al. [315], Wijerathna et al. [309] and Omondi and Demir [329] provide summaries of many of these studies which, taken together, identify numerous species of bacteria.
These bacteria are dominated by Gram-negative members belonging to the phylum Proteobacteria, with the Gram-positive phyla Firmicutes and Actinobacteria also represented, and include Ochrobactrum, Serratia marcescens, Klebsiella, Enterobacter, Escherichia coli, Pseudomonas aeruginosa, Pantoea agglomerans, Acinetobacter baumannii, Methylobacterium, Wolbachia, Spiroplasma, Enterococcus faecalis, Staphylococcus aureus, Bacillus cereus, B. anthracis, B. subtilis and B. megaterium [309, 315, 319, 328]. Several of these species belong to the phylum Proteobacteria, with Serratia and Enterobacter in the family Enterobacteriales and Pseudomonas forming core taxa, as has been described above in some other vector insects. There is also divergence in the bacteriome between wild and laboratory-reared sand flies as well as a reduction in taxa from Leishmania-parasitised insects [314].
Sand fly microbiome: native commensal bacteria in paratransgenesis
Regarding the choice of bacteria for paratransgenesis in sand flies, S. marcescens, P. agglomeratus and Enterobacter cloacae have already been tested in mosquitoes while E. coli has been utilised in triatomines (see preceding sections on different vectors). These isolates have been reported in sand flies so that the appropriate technology could be tranferred for use. There are, however, reports of pathogenicity for strains of these bacteria in humans so that alternative species have been identified for developing paratransgenesis in sand flies. In fact, the first experiment on the transstadial passage of commensal bacteria in sand flies was successful but with a potentially major pathogen, Ochrobactrum sp. in Phlebotomus duboscqi [319]. Subsequently, Hillesland et al. [320] identified several particularly suitable non-pathogenic bacteria, including B. subtilis, B. megaterium and Brevibacterium linens, for developing paratransgenesis in Phlebotomus argentipes. All three species are sold as probiotics, with B. megaterium also having potential as a biofertiliser for spreading in the environment [320].
The first study of the possibility of genetically manipulating sand fly commensal bacteria capable of effectively colonising the insect and remaining permanent throughout the life-cycle was conducted by Hurtwitz et al. [330, 331] with B. subtilis, previously isolated from P. argentipes. For this work, B. subtilis expressing a GFP reporter gene was added in the diet and offered to fourth stage sand fly larvae; the insects were then dissected and the midgut homogenates analysed for colonisation by recombinant bacteria using PCR and colony-forming units. The Bacillus-gfp colonised the fourth stage larvae of P. argentipes effectively and stably and could be recovered throughout the different stages of insect development. The recombinant strain was isolated from all larvae and pupae and from 75% of adults. In addition, sand fly adult emergence over 18 days was similar in B. subtilis-treated larvae and controls [330]. There was also no apparent horizontal transfer of the plasmid used for transformation to other bacteria in the gut. Whether the bacteria affected the female sand fly fecundity or were transmitted to the progeny was not determined. The transformation of B. megaterium to express a single chain antibody has also been reported [320], as has the development of melittin and human histone 2B as anti-Leishmania effector molecules [332].
More recently, Abassi et al. [325] also studied the transformation of the commensal sand fly bacterium, Enterobacter cloacae subsp. dissolvens, to express a defensin and colonise Phlebotomos papatasi. This defensin is of plant origin and able to kill parasites but not bacteria [325]. The bacteria were transformed with a red fluorescent protein plus defensin plasmid. When first instar larvae were fed just once on a diet containing the transformed bacteria, the latter could be detected up to 36 days post-feeding but there was no transstadial transmission to adult sand flies. This may be due to the loss of the bacteria during pupation and/or the inability of bacteria to colonise the gut due to physiological changes, such as those of the pH gradients [333].
In conclusion, much additional work is required with sand flies before paratransgenesis can be fully instigated for field trials.
Concluding remarks including safety and environmental concerns
This overview describes progress in the development of paratransgenesis in vector insects and shows that the majority of the research, not surprisingly, is currently focussed on mosquitoes. As mentioned previously, the original technique was pioneered nearly 30 years ago and although significant technical advances have been made with regulatory laws in mind, approval for use in the field has yet to be obtained. Many of these advances are shown in Table 1, but reasons for failure to have paratransgenesis adopted into IPM programmes are manifold. Some of the problems to be addressed have been identified previously (e.g. [34, 69, 83, 334, 335]) and their partial resolutions are indicated in Table 1.
To gain support from regulatory bodies, there are important requisites, including proof that the released transformed bacteria are stable and non-pathogenic to other animals and humans, and that they do not infect harmless insects by horizontal gene transfer. In addition, it is necessary to have detailed studies of the tripartite interactions of vector microbiomes from different ecosystems with vector immunity and invading parasites in order to identify suitable candidate microbes for paratransgenesis [83, 157]. Furthermore, ideal properties of transgenic bacteria include ease of spread into wild populations, the ability to pass transstadially and the ability to persist in different generations of the vector without inducing resistance (e.g. [336, 337]).
Table 1 gives examples of studies satisfying many of these requirements. For example, horizontal gene transfer in vivo was shown by Huang et al. [134] not to occur in Anopheles stephensi infected with fluorescent Serratia. Additionally, Matthews et al. [133] modelled the chances of horizontal gene transfer in the gut of Rhodnius prolixus and predicted the frequency of this process occurring at less than 1.14 × 10–16 per 100,000 generations with a 99% certainty level. Other attempts to allay fears about the effects of transgenic bacteria released into the environment and prevent their excessive spread have also been reported using microencapsulation techniques (Table 1). For example, in semi-field trials with Rhodococcus rhodnii in R. prolixus, the bacteria were enclosed in guar gum to form CRUZIGARD [71] while with Pantoea agglomerans in Homalodisca vitripennis an alginate hydrogel was used [128]. The work of Huang et al. [134] on Serratia ASI in An. stephensi also specifically addresses the concerns of regulators about the effects of the release of transgenic bacteria on the environment if something goes wrong. These authors showed that the plasmids used for transforming Serratia were only transient and lost in vivo after three generations of mosquitoes, with the bacteria returning to the wild type [134] (Table 1). Also, concerns about the possible pathogenicity of any released transgenic bacteria towards animals and humans must be considered. For example, species/strains of insect endosymbionts used in paratransgenesis, such as P. agglomerans, Asaia and Serratia, have all been described as opportunistic pathogen in humans [203, 338, 339], although it is unlikely that these pathogens are the same as the bacteria isolated from insects. There are even reports of probiotics like Lactobacilli acting as human pathogens [340].
Nevertheless, it will be necessary to undertake risk assessment tests on the potential pathogenicity of transgenic bacteria of the sort described by Beard et al. [69], before their release into the environment [337]. Finally, the question of the target parasite potentially developing resistance to the effector molecules expressed by the transgenic bacteria has been addressed by Wang et al. [120], working with transformed Serratia AS1 in An. stephensi. The Serratia were engineered to produce, simultaneously, multiple effector genes with different targets in Plasmodium falciparum, significantly reducing the likelihood of possible resistance, and when the bacteria were fed to mosquitoes, 48 h before an infected blood meal, oocyst loads were reduced > 91% (Table 1).
While the above examples appear to answer many of the concerns of the regulatory bodies, they are insufficient by themselves to gain approval. First, semi-field experiments are limited in terms of number of insects utilised and subsequent risk assessments made of the effects on other insect species and the environment. Types of experimental data required by the European Food Safety Authority/European Commission (EFSA/EC) report on risk assessment [338] include: (i) demonstration of an exact understanding of the genetic modification of the GM organism; (ii) details of the release method and the receiving environment; (iii) any interactions (intended or unintended) between the GM organism and the recipient environment; and (iv) validated protocol details for monitoring and control of the GM organism following release [336]. Secondly, it is relatively early days in seeking approval for the use of paratransgenesis in the field and, as such, the structure of regulations governing the release of the engineered bacteria used may be insufficient.
The authors recommend consulting the EFSA document “Guidance on the environmental risk assessment of genetically modified animals” [337], which states clearly that “scientific activities in the area of GM animals indicate that future applications may include traits related to disease resistance” and “insects (e.g. mosquitoes, agricultural pests, bees)” are now part of the remit of this organisation. Other countries have their own bodies regulating the release of GM organisms; these include Brazil (the National Biosafety Technical Commission [CTNBio]) [341] and the USA (the Food and Drug Administration Center for Veterinary Medicine [FDA-CVM], the Centers for Disease Control and Prevention [CDC] and the Environmental Protection Agency [EPA]) [342]. Only now are some of these bodies developing regulations for the release of GM animals [341]. A search of the USA regulatory bodies (20 March 2021) identified only one reference to paratransgenesis, and this was for development of a paratransgenesis system to control Pierce’s disease of grapes (see EPA TSCA [343]). Contacting the relevant bodies above for guidance is highly recommended.
Anybody applying for approval for the release of transformed bacteria/vector insects in the environment should read the EPA documents submitted for field testing of genetically modified baculoviruses in the 1990s for the control of insect pests on plants [342]. Much of the work was concerned with the addition of scorpion toxin genes to enhance the kill rate of polyhedrosis viruses without a consequent change in host range of the viruses. These scorpion toxins modulate sodium channels but do not affect vertebrate activity, and testing with surrogate species and human cell lines also revealed no toxicity. It is, therefore, significant that scorpine has already been used successfully against Plasmodium berghei in Anopheles gambiae and An. stephensi mosquitoes in paratransgenisis experiments [e.g. 121]. However, regulations relevant to the 1990s would, no doubt, have been updated by now.
Apart from the need for the appropriate scientific experimental work and satisfying the regulatory laws, there are important additional considerations before the GM organism can be released (Fig. 3). These include the social and public health aspects [344, 345], which were dealt with in detail during the field releases of Oxitec’s GM transgenic Aedes aegypti mosquitoes in the Cayman Islands, Brazil and Mexico [344,345,346]. The public health dimension provides evidence to justify the intervention of the GM organisms in a particular health risk. The failure of present control strategies to contain dengue infections would provide the need for such new strategies. The people actually involved in this process would include those at risk from dengue, scientists and regulatory bodies [344]. The social aspect is also very important as it involves the local community in the project and is essential for nurturing trust and approval for the GM mosquito release process [344, 345, 347]. Details for formulating these scientific, regulatory, public health and social dimensions are given in the references cited above. The whole of the GM release process can take years of collaborative work due to lack of any pipeline created by previous successful projects. Even with these dimensions fulfilled, there has recently been strong opposition to GM mosquito release in Florida despite EPA and CDC approval [347, 348].

Some of the important considerations, apart from the laboratory and field experiments, which are vital for gaining approval prior to the release of genetically modified organisms, including mosquitoes, containing transgenic bacteria
Regarding the future of paratransgenesis, progress to approval is slow and the processes involved daunting. However, increases in pesticide and drug resistance and climate change have resulted in enhanced zoonoses and losses in food crops due to insect pests [349, 350], so that alternative strategies like paratransgenesis will be required in IPM schemes [351]. The development of paratransgenesis is one answer to the recent World Health Organisation’s “call for innovation” for “new malaria-fighting tools and approaches” [352]. The value of such new approaches will become self-evident once epidemiological results begin to show impacts on disease for the use of GM mosquitoes and paratransgenesis, and also indicate that the techniques could be cost-effective [346].
Availability of data and materials
All references cited are listed in the bibliography.
Abbreviations
- AgDNV:
-
Anopheles gambiae densonucleosis viruses
- AMP:
-
Antimicrobial peptide
- Asaia GFP :
-
Asaia strain labelled with green fluorescent protein
- BMI:
-
Blood meal-induced promoters
- CRISPRs:
-
Clustered regularly interspaced short palindromic repeats
- EFSA:
-
European Food Safety Authority
- EFSA/EC:
-
EFSA/European Commission
- EPA:
-
Environmental Protection Agency
- GFP:
-
Green fluorescent protein
- GM:
-
Genetically modified
- HTS:
-
High-throughput sequencing
- kanR:
-
Kanamycin resistant
- mCherry:
-
Red fluorescent protein
- MDVs:
-
Mosquito densoviruses
- PbGFPcon:
-
Plasmodium berghei engineered to express GFP
- pWCP:
-
Culex pipiens plasmid
- rDB3:
-
Murine antibody fragments
- recSodalis :
-
Recombinant Sodalis
- RHBP :
-
Rhodnius heme-binding protein
- SMI:
-
Salivary gland and midgut molecules
References
Gullan PJ, Cranston PS. The insects. An outline of entomology. 4th ed. Oxford: Wiley-Blackwell; 2010.
Baddi BK. Phylogeny and functional morphology of Diptera (flies). London: IntechOpen; 2020. https://doi.org/10.5772/intechopen.90421.
Snow RW, Guerra CA, Noor AM, Myint HY, Hay SI. The global distribution of clinical episodes of Plasmodium falciparum malaria. Nature. 2005;434:214–7.
Bhatt S, Gething PW, Brady OJ, Messina JP, Farlow AW, Moyes CL, et al. The global distribution and burden of dengue. Nature. 2013;496:504–7.
Chandy A, Thakur AS, Singh MP, Manigauha A. A review of neglected tropical diseases: filariasis. Asian Pac J Trop Med. 2011;4:581–6.
Petersen LR, Brault AC, Nasci RS. West Nile virus: review of the literature. JAMA. 2013;310:308–15.
Weaver SC, Lecuit M. Chikungunya virus and the global spread of a mosquito-borne disease. N Engl J Med. 2015;372:1231–9.
Weaver SC, Costa F, Garcia-Blanco MA, Ko AI, Ribeiro GS, Saade G, et al. Zika virus: history, emergence, biology, and prospects for control. Antivir Res. 2016;130:69–80.
World Health Organization (WHO). A global brief on vector-borne diseases. 2014. https://apps.who.int/iris/handle/10665/111008, Accessed 23 Jan 2021.
World Health Organization (WHO). Ethics and vector-borne diseases: WHO guidance. 2020. https://apps.who.int/iris/handle/10665/336075. Accessed 19 Jan 2021.
Torres-Guerrero E, Quintanilla-Cedillo MR, Ruiz-Esmenjaud J, Arenas R. Leishmaniasis: a review. F1000 Res. 2017;6:750.
World Health Organization (WHO). Leishmaniasis. 2019. https://www.who.int/leishmaniasis. Accessed 09 Nov 2020.
Aksoy S, Buscher P, Lehane M, Solano P, Abbeele JVD. Human African trypanosomiasis control: achievements and challenges. PLoS Negl Trop Dis. 2017;11: e0005454.
World Health Organization (WHO). Trypanosomiasis, human African (sleeping sickness). Updated February 2020. 2020. https://www.who.int/en/news-room/fact-sheets/detail/trypanosomiasis-human-african-(sleeping-sickness). Accessed 09 Nov 2020.
U.S. Center for Disease Control and Prevention (CDC). Malaria‘s impact worldwide. 25 Feb 2020. https://www.cdc.gov›malaria›impact. Accessed 09 Nov 2020.
Bern C. Chagas’ disease. N Engl J Med. 2015;373:456–66.
Lai O, Ho D, Glick S, Jagdeo J. Bed bugs and possible transmission of human pathogens: a systematic review. Arch Dermatol Res. 2016;308:531–8.
Perilla-Henao LM, Casteel CL. Vector-borne bacterial plant pathogens: interactions with hemipteran insects and plants. Front Plant Sci. 2016;7:1163.
Semenza JC, Suk JE. Vector-borne diseases and climate change: a European perspective. FEMS Microbiol Lett. 2018;365: fnx244.
Guzmán C, Calderón A, Mattar S, Tadeu-Figuereido L, Salazar-Bravo J, Alvis-Guzmán N, et al. Eco epidemiology of alphaviruses and flaviviruses. In: Ennaji MM, et al., editors. Emerging and reemerging viral pathogens. Cordoba: Moulay Ennaji; 2020. p. 101–25.
Pan American Health Organization/World Health Organization (PAHO/WHO). Response to the epidemic of Zika virus in the Americas, December 2015–2016. https://iris.paho.org/handle/10665.2/50608. Accessed 05 Dec 2020.
Organisation for Economic Co-operation and Development (OECD), Environment Directorate. Consensus document on the biology of mosquito Aedes aegypti, No. 65, ENV/JM/MONO(2018)23. 2018. www.oecd.org/science/biotrack/. Accessed 10 Dec 2020.
Achee NL, Grieco JP, Vatandoost H, Seixas G, Pinto J, Ching-NG L, et al. Alternative strategies for mosquito-borne arbovirus. PLoS Negl Trop Dis. 2019;13:e0007275.
Dahmana H, Mediannikov O. Mosquito-borne diseases emergence/resurgence and how to effectively control it biologically. Pathogens. 2020;9:310.
Wilson AL, Courtenay O, Kelly-Hope LA, Scott TW, Takken W, Torr SJ, et al. The importance of vector control for the control and elimination of vector-borne diseases. PloS Negl Trop Dis. 2020;14(1):0007831.
United States Department of Agriculture-Agricultural Research Service (USDA-ARS). A national road map for integrated pest management. 2018. https://www.ars.usda.gov/ARSUserFiles/OPMP/IPM%20Road%20Map%20FINAL.pdf. Accessed 14 Jan 2021.
Environmental Protection Agency (EPA). DDT ban takes effect. EPA press release on the ban of general use of the pesticide dichloro-diphenyl-trichloroethane (DDT). 31 Dec 1972. https://archive.epa.gov/epa/aboutepa/ddt-ban-takes-effect.html. Accessed 10 Jan 2020.
National Research Council. The life sciences: recent progress and application to human affairs. The world of biological research requirements for the future. Washington DC: The National Academies Press; 1970. https://doi.org/10.17226/9575.
Mougabure-Cueto G, Picollo MI. Insecticide resistance in vector Chagas disease: evolution, mechanisms and management. Acta Trop. 2015;149:70–85.
Riveron JM, Tchouakui M, Mugenzi L, Menze BD, Chiang M-C, Wondji CS. Insecticide resistance in malaria vectors: an update at a global scale. In: Manguin S, Dev V, editors. Towards malaria elimination—a leap forward. London: IntechOpen; 2018. p. 149–75. https://doi.org/10.5772/intechopen.78375.
Ranson H, Lissenden N. Insecticide resistance in African Anopheles mosquitoes: a worsening situation that needs urgent action to maintain malaria control. Trends Parasitol. 2016;32:187–96.
Shaw WR, Catterrucia F. Vector biology meets disease control: using basic research to fight vector-borne diseases. Nat Microbiol. 2019;4(1):20–34.
Barratt BIP, Moran VC, Bigler F. The status of biological control and recommendations for improving uptake for the future. Biocontrol. 2018;63:155–67.
Wilke ABB, Marrelli MT. Paratransgenesis: a promising new strategy for mosquito vector control. Parasit Vectors. 2015;8:342.
Evans BR, Kotsakiozi P, Costa-da-Silva AL, Ioshino RS, Garziera L, Pedrosa MC, et al. Transgenic Aedes aegypti mosquitoes transfer genes into a natural population. Sci Rep. 2019;9:13047.
Kassegne K, Zhou XN, Chen JH. Editorial: vectors and vector-borne parasitic diseases: infection, immunity, and evolution. Front Immunol. 2021;12:729415.
Duffy PE, Gorres JP. Malaria vaccines since 2000: progress, priorities, products. npj Vaccines. 2020;5:48. https://doi.org/10.1038/s41541-020-0196-3.
Biswal S, Reynales H, Saez-Lloren X, Lopez P, Borja-Tabora C, Kosalaraksa P, et al. Efficacy of a tetravalent dengue vaccine in healthy children and adolescents. N Engl J Med. 2019;381:2009–19.
Lecouturier V, Pavot V, Berry C. An optimized purified inactivated Zika vaccine—sustained immunogenicity and protection in cynomolgus macaques. npj Vaccines. 2020;5:19. https://doi.org/10.1038/s41541-020-0167-8.
Manning JE, Cantaert T. Time to micromanage the pathogen-host-vector interface: considerations for vaccine development. Vaccines. 2019;7(1):10.
Datoo MS, Natama MH, Somé A, Traoré O, Rouamba T, Bellamy D, et al. Efficacy of a low-dose candidate malaria vaccine, R21 in adjuvant Matrix-M, with seasonal administration to children in Burkina Faso: a randomised controlled trial. Lancet. 2021;397(10287):1809–18.
Balfour H. EMA accepts regulatory submission for Takeda’s dengue vaccine candidate. Eur Pharm Rev. 2021. https://www.europeanpharmaceuticalreview.com/news/.
Capela R, Moreira R, Lopes F. An overview of drug resistance in protozoal diseases. Int J Mol Sci. 2019;20(22):5748.
Pramanik PK, Alam MN, Chowdhury DR, Chakraborti T. Drug resistance in protozoan parasites: an incessant wrestle for survival. J Glob Antimicrob Resist. 2019;18:1–11.
Diakité SAS, Traoré K, Sanogo I, Clark TG, Campino S, Sangaré M, et al. A comprehensive analysis of drug resistance molecular markers and Plasmodium falciparum genetic diversity in two malaria endemic sites in Mali. Malar J. 2019;18:361.
Gyapong JO, Owusu IO, da Costa Vroom FB. Elimination of lymphatic filariasis: current perspectives on mass drug administration. Res Rep Trop Med. 2018;2018:25–33. https://doi.org/10.2147/RRTM.S125204.
Espinal MA, Andrus JK, Jauregui B, Waterman SH, Morens DM, Santos JI, et al. Emerging and reemerging Aedes-transmitted arbovirus infections in the region of the Americas: implications for health policy. Am J Public Health. 2019;109(3):387–92.
Marchesi JR, Ravel J. The vocabulary of microbiome research: a proposal. Microbiome. 2015;3:31.
Ricci I, Damiani C, Rossi P, Capone A, Scuppa P, Cappelli A, et al. Mosquito symbioses: from basic research to the paratransgenic control of mosquito-borne diseases. J Appl Entomol. 2011;135:487–93.
Rajashekhar M, Dharavath V, Savani AK, Raidu LC, Kalia V. Endosymbionts and paratransgenesis approach for pest management. J Progress Res Soc Sci. 2016;11(V):2822–6.
Scolari F, Casiraghi M, Bonizzoni M. Aedes spp. and their micribiota: a review. Front Microbiol. 2019;10:2036. https://doi.org/10.3389/fmicb.2019.02036.
Whitten MM, Shiao SH, Levashina EA. Mosquito midguts and malaria: cell biology, compartmentalization and immunology. Parasite Immunol. 2006;28(4):121–30.
Mancini MV, Spaccapelo R, Damiani C, Accoti A, Tallarita M, Petraglia E, et al. Paratransgenesis to control malaria vectors: a semi-field pilot study. Parasit Vectors. 2016;9:140.
Azambuja P, Feder D, Garcia ES. Isolation of Serratia marcescens in the midgut of Rhodnius prolixus: impact on the establishment of the parasite, Trypanosoma cruzi, in the vector. Exp Parasitol. 2004;107(1–2):89–96.
Ahmad NA, Vythilingam I, Lim YAL, Zabari NZAM, Lee HL. Detection of Wolbachia in Aedes albopictus and their effects on chikungunya virus. Am J Trop Med Hyg. 2017;96:148–56.
Dobson SL, Marsland EJ, Veneti Z, Bourtzis K, O’Neill SL. Characterization of Wolbachia host cell range via the in vitro establishment of infections. Appl Environ Microbiol. 2002;68(2):656–60.
Da Silva GD, Iturbe-Ormaetxe I, Martins-da-Silva A, Telleria EL, Rocha MN, Traub-Csekö YM, et al. Wolbachia introduction into Lutzomyia longipalpis (Diptera: Psychodidae) cell lines and its effects on immune-related gene expression and interaction with Leishmania infantum. Parasit Vectors. 2019;12:33.
Krafsur ES. Tsetse flies: genetics, evolution, and role as vectors. Infect Genet Evol. 2009;9:124–41.
Hofer U. New horizons for Wolbachia. Nat Rev Microbiol. 2017;15:66–7.
Kamtchum-Tatuene J, Makepeace BL, Benjamin L, Baylis M, Solomon T. The potential role of Wolbachia in controlling the transmission of emerging human arboviral infections. Curr Opin Infect Dis. 2017;30(1):108–16.
Niang EHA, Bassene H, Fenollar F, Mediannikov O. Biological control of mosquito-borne diseases: the potential of Wolbachia-based interventions in an IVM framework. J Trop Med. 2018;2018:1470459. https://doi.org/10.1155/2018/1470459.
Durovni B, Saraceni V, Eppinghaus A, Riback TIS, Moreira LA, Jewell NP, et al. The impact of large-scale deployment of Wolbachia mosquitoes on dengue and other Aedes-borne diseases in Rio de Janeiro and Niterói, Brazil: study protocol for a controlled interrupted time series analysis using routine disease surveillance data. F1000Research. 2020;8:1328.
Flores HA, Taneja de Bruyne J, O’Donnell TB, Nhu VT, Giang NT, Trang HTX, et al. Multiple Wolbachia strains provide comparative levels of protection against dengue virus infection in Aedes aegypti. PLoS Pathog. 2020;16(4): e1008433.
Ross PA, Callahan AG, Yang Q, Jasper M, Arif MAK, Afizah AN, et al. An elusive endosymbiont: does Wolbachia occur naturally in Aedes aegypti? Ecol Evol. 2020;10:1581–91.
Yen P-S, Failloux A-B. A review: Wolbachia-based population replacement for mosquito control shares common points with gentically modified control processes. Pathogens. 2020;9:404.
Beard CB, Mason PW, Aksoy S, Tesh RB, Richards FF. Transformation of an insect symbiont and expression of a foreign gene in the Chagas’ disease vector Rhodnius prolixus. Am J Trop Med Hyg. 1992;46(2):195–200.
Beard CB, O’Neill SL, Tesh RB, Richards FF, Aksoy S. Modification of arthropod vector competence via symbiotic bacteria. Parasitol Today. 1993;9(5):179–83.
Beard CB, Durvasula RV, Richards FF. Bacterial symbiosis in arthropods and the control of disease transmission. Emerg Infect Dis. 1998;4:581–91.
Beard CB, Dotson EM, Pennington PM, Eichler S, Cordon-Rosales C, Durvasula RV. Bacterial symbiosis and paratransgenic control of vector-borne Chagas disease. Int J Parasitol. 2001;31(5–6):621–7.
Beard CB, Cordon-Rosales C, Durvasula RV. Bacterial symbionts of the triatominae and their potential use in control of Chagas disease transmission. Annu Rev Entomol. 2002;47:123–41.
Durvasula RV, Gumbs A, Panackal A, Kruglov O, Aksoy S, Merrifield RB, et al. Prevention of insect-borne disease: an approach using transgenic symbiotic bacteria. Proc Natl Acad Sci USA. 1997;94(7):3274–8.
Durvasula RV, Gumbs A, Panackal A, Kruglov O, Taneja J, Kang AS, et al. Expression of a functional antibody fragment in the gut of Rhodnius prolixus via transgenic bacterial symbiont Rhodococcus rhodnii. Med Vet Entomol. 1999;13:115–9.
Durvasula RV, Kroger A, Goodwin M, Panackal A, Kruglov O, Taneja J, et al. Strategy for introduction of foreign genes into field populations of Chagas disease vectors. Ann Entomol Soc Am. 1999;92:937–43.
Whitten MM, Facey PD, Del Sol R, Fernandez-Martinez LT, Evans MC, Mitchell JJ, et al. Symbiont-mediated RNA interference in insects. Proc R Soc B. 2016;283:20160042.
Dotson EM, Plikaytis B, Shinnick TM. Transformation of Rhodococcus rhodnii, a symbiont of the Chagas disease vector Rhodnius prolixus, with integrative elements of the L1 mycobacteriophage. Infect Genet Evol. 2003;3(2):103–9.
Hurwitz I, Fieck A, Read A, Hillesland H, Klein N, Kang A, et al. Paratransgenic control of vector-borne diseases. Int J Biol Sci. 2011;7(9):1334–44.
Hurwitz I, Fieck A, Durvasula R. Antimicrobial peptide delivery strategies: use of recombinant antimicrobial peptides in paratransgenic control systems. Curr Drug Targets. 2012;13(9):1173–80. https://doi.org/10.2174/138945012802002366.
Sassera D, Epis S, Pajoro M, Bandi C. Microbial symbiosis and the control of vector-borne pathogens in tsetse flies, human lice, and triatomine bugs. Pathog Glob Health. 2013;107(6):285–92.
Raharimalala FN, Boukraa S, Bawin T, Boyer S, Francis F. Molecular detection of six (endo-) symbiotic bacteria in Belgian mosquitoes: first step towards the selection of appropriate paratransgenesis candidates. Parasitol Res. 2016;115(4):1391–9.
Fieck A, Hurwitz I, Kang AS, Durvasula R. Trypanosoma cruzi: synergistic cytotoxicity of multiple amphipathic anti-microbial peptides to T. cruzi and potential bacterial hosts. Exp Parasitol. 2010;125(4):342–7.
Wang S, Jacobs-Lorena M. Paratransgenesis applications: fighting malaria with engineered mosquito symbiotic bacteria. In: Wikel SK, Aksoy S, Dimopoulos G, editors. Vector microbiome and innate immunity of arthropods, vol. 1. Oxford: Elsevier; 2017. p. 219–34. https://doi.org/10.5772/intechopen.90421.
Hurwitz I, Fieck A, Klein N, Jose C, Kang A, Durvasula R. A paratransgenic strategy for the control of Chagas disease. Psyche. 2012;2012:178930. https://doi.org/10.1155/2012/178930.
Saldaña MA, Hegde S, Hughes GL. Microbial control of arthropod-borne disease. Mem Inst Oswaldo Cruz. 2017;112(2):81–93.
Allaband C, McDonald D, Vázquez-Baeza Y, Minich JJ, Tripathi A, Brenner DA, et al. Microbiome 101: studying, analyzing, and interpreting gut microbiome data for clinicians. Clin Gastroenterol Hepatol. 2019;17(2):218–30.
Lagier JC, Hugon P, Khelaifia S, Fournier PE, La Scola B, Raoult D. The rebirth of culture in microbiology through the example of culturomics to study human gut microbiota. Clin Microbiol Rev. 2015;28(1):237–64.
Kean J, Rainey SM, McFarlane M, Donald CL, Schnettler E, Kohl A, Pondeville E. Fighting arbovirus transmission: natural and engineered control of vector competence in Aedes mosquitoes. Insects. 2015;6(1):236–78.
Chandler JA, Liu RM, Bennett SN. RNA shotgun metagenomic sequencing of northern California (USA) mosquitoes uncovers viruses, bacteria, and fungi. Front Microbiol. 2015;6:185.
Nanfack MF, Vernick KD. A systematic review of the natural virome of Anopheles mosquitoes. Viruses. 2018;10(5):222.
Carlson J, Suchman E, Buchatsky L. Densoviruses for control and genetic manipulation of mosquitoes. Adv Virus Res. 2006;68:361–92.
Liu P, Li X, Gu J, Dong Y, Liu Y, Santhosh P, et al. Development of non-defective recombinant densovirus vectors for microRNA delivery in the invasive vector mosquito, Aedes albopictus. Sci Rep. 2016;6:20979.
Ren X, Hoiczyk E, Rasgon JL. Viral paratransgenesis in the malaria vector Anopheles gambiae. PLoS Pathog. 2008;4(8):e1000135.
Gu JB, Dong YQ, Peng HJ, Chen XG. A recombinant AeDNA containing the insect-specific toxin, BmK IT1, displayed an increasing pathogenicity on Aedes albopictus. Am J Trop Med Hyg. 2010;83:614–23.
Johnson RM, Rasgon JL. Densonucleosis viruses (‘densoviruses’) for mosquito and pathogen control. Curr Opin Insect Sci. 2018;28:90–7.
Sun Y, Dong Y, Li J, Lai Z, Hao Y, Liu P, et al. Development of large-scale mosquito densovirus production by in vivo methods. Parasit Vectors. 2019;12:255.
Moraes AML, Junqueira ACV, Celano V, Costa GL, Coura JR. Fungal flora of the digestive tract of Rhodnius prolixus, Rhodnius neglectus, Diptelanogaster maximus and Panstrongylus megistus, vectors of Trypanosoma cruzi, Chagas, 1909. Braz J Microbiol. 2004;35(4):288–91.
Akhoundi M, Bakhtiari R, Guillard T, Baghaei A, Tolouei R, Sereno D, et al. Diversity of the bacterial and fungal microflora from the midgut and cuticle of phlebotomine sand flies collected in North-Western Iran. PLoS ONE. 2012;7(11): e50259.
Jayakrishnan L, Sudhikumar AV, Aneesh EM. Role of gut inhabitants on vectorial capacity of mosquitoes. J Vector Borne Dis. 2018;55(2):69–78.
Cappelli A, Valzano M, Cecarini V, Bozic J, Rossi P, Mensah P, et al. Killer yeasts exert anti-plasmodial activities against the malaria parasite Plasmodium berghei in the vector mosquito Anopheles stephensi and in mice. Parasit Vectors. 2019;12:329.
Murphy KA, Tabuloc CA, Cervantes KR, Chiu JC. Ingestion of genetically modified yeast symbiont reduces fitness of an insect pest via RNA interference. Sci Rep. 2016;6:22587.
Fang W, Vega-Rodríguez J, Ghosh AK, Jacobs-Lorena M, Kang A, St Leger RJ. Development of transgenic fungi that kill human malaria parasites in mosquitoes. Science. 2011;331(6020):1074–7.
Rasgon JL. Using infections to fight infections: paratransgenic fungi can block malaria transmission in mosquitoes. Future Microbiol. 2011;6(8):851–3.
Dong Y, Maria L, Simões ML, Marois E, Dimopoulos G. CRISPR/Cas9-mediated gene knockout of Anopheles gambiae FREP1 suppresses malaria parasite infection. PLoS Pathog. 2018;14(3):e1006898.
Moreira D, Pereira AM, Lopes AL, Coimbra S. The best CRISPR/Cas9 versus RNA interference approaches for Arabinogalactan proteins’ study. Mol Biol Rep. 2020;47:2315–25.
Yang J, Schleicher TR, Dong Y, Park HB, Lan J, Cresswell P, et al. Disruption of mosGILT in Anopheles gambiae impairs ovarian development and Plasmodium infection. J Exp Med. 2020;217(1):e20190682.
Hegde S, Khanipov K, Albayrak L, Golovko G, Pimenova M, Saldaña MA, et al. Microbiome interaction networks and community structure from laboratory-reared and field-collected Aedes aegypti, Aedes albopictus, and Culex quinquefasciatus mosquito vectors. Front Microbiol. 2018;9:2160.
Burt A, Coulibaly M, Crisanti A, Diabate A, Kayondo JK. Gene drive to reduce malaria transmission in sub-Saharan Africa. J Responsible Innov. 2018;5(Suppl 1):66–80.
Wilke A, Beier J, Benelli G. Transgenic mosquitoes—fact or fiction? Trends Parasitol. 2018;34(6):456–65.
Boettcher M, McManus MT. Choosing the right tool for the job: RNAi, TALEN, or CRISPR. Mol Cell. 2015;58(4):575–85.
Xi Z, Ramirez JL, Dimopoulos G. The Aedes aegypi toll pathway controls dengue virus infection. PLoS Pathog. 2008;4(7):e1000098.
Hentzschel F, Mitesser V, Fraschka SAK, Krzikalla D, Carrillo EH, Berkhout B, et al. Gene knockdown in malaria parasites via non-canonical RNAi. Nucleic Acids Res. 2020;48(1):e2. https://doi.org/10.1093/nar/gkz927.
Taracena ML, Oliveira PL, Almendares O, Umaña C, Lowenberger C, Dotson EM, et al. Genetically modifying the insect gut microbiota to control Chagas disease vectors through systemic RNAi. PLoS Negl Trop Dis. 2015;9(2):e0003358.
Abrieux A, Chiu JC. Oral delivery of dsRNA by microbes: beyond pest control. Commun Integr Biol. 2016;9(6):e1236163.
Whitten M, Dyson P. Gene silencing in non-model insects: overcoming hurdles using symbiotic bacteria for trauma-free sustainable delivery of RNA interference. BioEssays. 2017;39:1600247.
Vogel E, Santos D, Mingels L, Verdonckt T-W, Broeck JV. RNA interference in insects: protecting beneficials and controlling pests. Front Physiol. 2019;10:1912.
Asgari M, Ilbeigikhamsehnejad M, Rismani E, Djadid ND, Raz A. Molecular characterization of RNase III protein of Asaia sp. for developing a robust RNAi-based paratransgensis tool to affect the sexual life-cycle of Plasmodium or Anopheles fitness. Parasit Vectors. 2020;13:42.
Leonard SP, Powell JE, Perutka J, Geng P, Heckmann LC, Horak RD, et al. Engineered symbionts activate honey bee immunity and limit pathogens. Science. 2020;367:573–6.
Araujo RN, Santos A, Pinto FS, Gontijo NF, Lehane MJ, Pereira MH. RNA interference of the salivary gland nitrophorin 2 in the triatomine bug Rhodnius prolixus (Hemiptera: Reduviidae) by dsRNA ingestion or injection. Insect Biochem Mol Biol. 2006;36(9):683–93.
Walshe DP, Lehane SM, Lehane MJ, Haines LR. Prolonged gene knockdown in the tsetse fly Glossina by feeding double stranded RNA. Insect Mol Biol. 2009;18:11–9.
Timmons L, Court DL, Fire A. Ingestion of bacterially expressed dsRNAs can produce specific and potent genetic interference in Caenorhabditis elegans. Gene. 2001;263:103–12.
Wang S, Santos ALA, Huang W, Liu KC, Oshaghi MA, Wei G, et al. Driving mosquito refractoriness to Plasmodium falciparum with engineered symbiotic bacteria. Science. 2017;357(6358):1399–402.
Wang S, Ghosh AK, Bongio N, Stebbings KA, Lampe DJ, Jacobs-Lorena M. Fighting malaria with engineered symbiotic bacteria from vector mosquitoes. Proc Natl Acad Sci USA. 2012;109(31):12734–9.
Wang S, Jacobs-Lorena M. Genetic approaches to interfere with malaria transmission by vector mosquitoes. Trends Biotechnol. 2013;31(3):185–93. https://doi.org/10.1016/j.tibtech.2013.01.001.
Sreenivasamurthy SK, Dey G, Ramu M, Kumar M, Gupta MK, Mohanty AK, et al. A compendium of molecules involved in vector-pathogen interactions pertaining to malaria. Malar J. 2013;12:216.
Carter V, Underhill A, Baber I, Sylla L, Baby M, Larget-Thiery I, et al. Killer bee molecules: antimicrobial peptides as effector molecules to target sporogonic stages of Plasmodium. PLoS Pathog. 2013;9(11): e1003790.
Dong Y, Simões ML, Dimopoulos G. Versatile transgenic multistage effector-gene combinations for Plasmodium falciparum suppression in Anopheles. Sci Adv. 2020;6: eaay5898.
Shane JL, Grogan CL, Cwalina C, Lampe DJ. Blood meal-induced inhibition of vector-borne disease by transgenic microbiota. Nat Commun. 2018;9(1):4127.
Berasategui A, Shukla S, Salem H, Kaltenpoth M. Potential applications of insect symbionts in biotechnology. Appl Microbiol Biotechnol. 2016;100:1567–77.
Arora AK, Forshaw A, Miller TA, Durvasula R. A delivery system for field application of paratransgenic control. BMC Biotechnol. 2015;15:59.
Arora AK, Miller TA, Durvasula R. Transmission of Pantoea agglomerans—a paratransgenic control agent—within a Homalodisca vitripennis population. J Appl Entomol. 2019;144:232–5.
Kotnis B, Kuri J. Evaluating the usefulness of paratransgenesis for malaria control. Math Biosci. 2016;277:117–25.
Yin C, Sun P, Yu X, Wang P, Chen G. Roles of symbiotic microorganisms in arboviral infection of arthropod vectors. Trends Parasitol. 2020;36:607–15.
Boissière A, Tchioffo MT, Bachar D, Abate L, Marie A, Nsango SE, et al. Midgut microbiota of the malaria mosquito vector Anopheles gambiae and interactions with Plasmodium falciparum infection. PLoS Pathog. 2012;8:e1002742.
Matthews S, Rao VS, Durvasula RV. Modeling horizontal gene transfer (HGT) in the gut of the Chagas disease vector Rhodnius prolixus. Parasit Vectors. 2011;4:77–85.
Huang W, Wang S, Jacobs-Lorena M. Self-limiting paratransgenesis. PLoS Negl Trop Dis. 2020;14(8):e0008542.
Hegde S, Nilyanimit P, Kozlova E, Anderson ER, Narra HP, Sahni SK, et al. CRISPR/Cas9-mediated gene deletion of the ompA gene in symbiotic Cedecea neteri impairs biofilm formation and reduces gut colonization of Aedes aegypti mosquitoes. PLoS Negl Trop Dis. 2019;13(12): e0007883.
Li J, Hanc M, Yu J. Simple paratransgenic mosquitoes models and their dynamics. Math Biosci. 2018;306:20–31.
Coon KL, Vogel KJ, Brown MR, Strand MR. Mosquitoes rely on their gut microbiota for development. Mol Ecol. 2014;23(11):2727–39.
Alfano N, Tagliapietra V, Rosso F, Manica M, Arnoldi D, Pindo M, Rizzoli A. Changes in microbiota across developmental stages of Aedes koreicus, an invasive mosquito vector in Europe: indications for microbiota-based control strategies. Front Microbiol. 2019;10:2832.
Bennett KL, Gómez-Martínez C, Chin Y, Saltonstall K, McMillan WO, Rovira JR, et al. Dynamics and diversity of bacteria associated with the disease vectors Aedes aegypti and Aedes albopictus. Sci Rep. 2019;9:12160.
Duguma D, Hall MW, Rugman-Jones P, Stouthamer R, Terenius O, Neufeld JD, et al. Developmental succession of the microbiome of Culex mosquitoes. BMC Microbiol. 2015;15:140.
Seabourn P, Spafford H, Yoneishi N, Medeiros M. The Aedes albopictus (Diptera: Culicidae) microbiome varies spatially and with Ascogregarine infection. PLoS Negl Trop Dis. 2020;14(8):e0008615.
Sharma P, Sharma S, Maurya RK, de Das T, Thomas T, Lata S, et al. Salivary glands harbor more diverse microbial communities than gut in Anopheles culicifacies. Parasit Vectors. 2014;7:235.
Tchioffo MT, Boissière A, Abate L, Nsango SE, Bayibéki AN, Awono-Ambéné PH, et al. Dynamics of bacterial community composition in the malaria mosquito’s epithelia. Front Microbiol. 2015;6:1500.
da Silva AF, Machado LC, de Paula MB, da Silva Pessoa Vieira CJ, de Morais Bronzoni RV, de Melo Santos MAV, et al. Culicidae evolutionary history focusing on the Culicinae subfamily based on mitochondrial phylogenomics. Sci Rep. 2020;10:18823. https://doi.org/10.1038/s41598-020-74883-3.
Engel P, Moran NA. The gut microbiota of insects—diversity in structure and function. FEMS Microbiol Rev. 2013;37:699–735.
Strand MR. Composition and functional roles of the gut microbiota in mosquitoes. Curr Opin Insect Sci. 2018;28:59–65.
Chen S, Zhang D, Augustinos A, Doudoumis V, Mokhtar NB, Maiga H, et al. Multiple factors determine the structure of bacterial communities associated with Aedes albopictus under artificial rearing conditions. Front Microbiol. 2020;11:605. https://doi.org/10.3389/fmicb.2020.00605.
Gupta A, Nair S. Dynamics of insect–microbiome interaction influence host and microbial symbiont. Front Microbiol. 2020;11:1357.
Sharma P, Ran J, Chauhan C, Kumari S, Tevatiya S, Tanwee DD, et al. Altered gut microbiota and immunity defines Plasmodium vivax survival in Anopheles stephensi. Front Immunol. 2020;11:609.
Gonzalez-Ceron L, Santillan F, Rodriguez MH, Mendez D, Hernandez-Avila JE. Bacteria in midguts of field collected Anopheles albimanus block Plasmodium vivax sporogonic development. J Med Entomol. 2003;40:371–4.
Dong Y, Manfredini F, Dimopoulos G. Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 2009;5:e1000423.
Moreira LA, Iturbe-Ormaetxe I, Jeffery JA, Lu G, Pyke AT, Hedges LM, et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell. 2009;139(7):1268–78.
Ramirez JL, Short SM, Bahia AC, Saraiva RG, Dong Y, Kang S, et al. Chromobacterium Csp_P reduces malaria and dengue infection in vector mosquitoes and has entomopathogenic and in vitro anti-pathogen activities. PLoS Pathog. 2014;10(10):e1004398.
Smith RC, Vega-Rodríguez J, Jacobs-Lorena M. The Plasmodium bottleneck: malaria parasite losses in the mosquito vector. Mem Inst Oswaldo Cruz. 2014;109(5):644–61.
Dieme C, Rotureau B, Mitri C. Microbial pre-exposure and vectorial competence of Anopheles mosquitoes. Front Cell Infect Microbiol. 2017;7:508.
Guégan M, Zouache K, Démichel C, Minard G, Van VT, Potier P, et al. The mosquito holobiont: fresh insight into mosquito–microbiota interactions. Microbiome. 2018;6(1):49.
Romoli O, Gendrin M. The tripartite interactions between the mosquito, its microbiota and Plasmodium. Parasit Vectors. 2018;11:200.
Caragata EP, Tikhe CV, Dimopoulos G. Curious entanglements: interactions between mosquitoes, their microbiota, and arboviruses. Curr Opin Virol. 2019;37:26–36.
Gabrieli P, Caccia S, Varotto-Boccazzi I, Arnoldi I, Barbieri G, Comandatore F, Epis S. Mosquito trilogy: microbiota, immunity and pathogens, and their implications for the control of disease transmission. Front Microbiol. 2021;12: 630438.
Chavshin AR, Oshaghi MA, Vatandoost H, Pourmand MR, Raeisi A, Terenius O. Isolation and identification of culturable bacteria from wild Anopheles culicifacies, a first step in a paratransgenesis approach. Parasit Vectors. 2014;7:419.
Dacey DP, Chain FJJ. The challenges of microbial control of mosquito-borne diseases due to the gut microbiome. Front Genet. 2020;11:504354. https://doi.org/10.3389/fgene.2020.504354.
Huang W, Wang S, Jacobs-Lorena M. Use of microbiota to fight mosquito-borne disease. Front Genet. 2020;11:196.
Oliveira TMP, Sanabani SS, Sallum MAM. Asaia (Rhodospirillales: Acetobacteraceae) and Serratia (Enterobacterales: Yersiniaceae) associated with Nyssorhynchus braziliensis and Nyssorhynchus darlingi (Diptera: Culicidae). Rev Bras Entomol. 2020;64(2):e20190010.
Dada N, Jupatanakul N, Minard G, Short SM, Akorli J, Villegas LM. Considerations for mosquito microbiome research from the Mosquito Microbiome Consortium. Microbiome. 2021;9:36.
Rodríguez-Ruano SM, Juhaňáková E, Vávra J, Nováková E. Methodological insight into mosquito microbiome studies. Front Cell Infect Microbiol. 2020;10:86.
Gendrin M, Christophides GK. The Anopheles mosquito microbiota and their impact on pathogen transmission. In: Manguin S, editor. Anopheles mosquitoes-new insights into malaria vectors. London: IntechOpen; 2013. p. 525–48.
Zouache K, Raharimalala FN, Raquin V, Tran-Van V, Raveloson LHR, Ravelonandro P, et al. Bacterial diversity of field-caught mosquitoes, Aedes albopictus and Aedes aegypti, from different geographic regions of Madagascar. FEMS Microbiol Ecol. 2011;75(3):377–89.
Osei-Poku J, Mbogo CM, Palmer WJ, Jiggins FM. Deep sequencing reveals extensive variation in the gut microbiota of wild mosquitoes from Kenya. Mol Ecol. 2012;21:5138–50.
David MR, dos Santos LMB, Vicente ACP, Maciel-de-Freitas R. Effects of environment, dietary regime and ageing on the dengue vector microbiota: evidence of a core microbiota throughout Aedes aegypti lifespan. Mem Inst Oswaldo Cruz. 2016;111(9):577–87.
Demaio J, Pumpuni CB, Kent M, Beier JC. The midgut bacterial flora of wild Aedes triseriatus, Culex pipiens, and Psorophora columbiae mosquitoes. Am J Trop Med Hyg. 1996;54(2):219–23.
Straif SC, Mbogo CN, Toure AM, Walker ED, Kaufman M, Toure YT, et al. Midgut bacteria in Anopheles gambiae and An. funestus (Diptera: Culicidae) from Kenya and Mali. J Med Entomol. 1998;35:222–6.
Pidiyar VJ, Jangid K, Patole MS, Shouche Y. Studies on cultured and uncultured microbiota of wild Culex quinquefasciatus mosquito midgut based on 16S ribosomal RNA gene analysis. Am J Trop Med Hyg. 2004;70:597–603.
Rami A, Raz A, Zakeri S, Djadid ND. Isolation and identification of Asaia sp. in Anopheles spp. mosquitoes collected from Iranian malaria settings: steps toward applying paratransgenic tools against malaria. Parasit Vectors. 2018;11(1):367.
Gusmão DS, Santos AV, Marini DC, Bacci M, Berbert-Molina MA, Lemos FJA. Culture-dependent and culture-independent characterization of microorganisms associated with Aedes aegypti (Diptera: Culicidae) (L.) and dynamics of bacterial colonization in the midgut. Acta Trop. 2010;115:275–81.
Gimonneau G, Tchioffo MT, Abate L, Boissière A, Awono-Ambene PH, Nsango SE, et al. Composition of Anopheles coluzzii and Anopheles gambiae microbiota from larval to adult stages. Infect Genet Evol. 2014;28:715–24.
Villegas LM, Pimenta PLP. Metagenomics, paratransgenesis and the Anopheles microbiome: portrait of the geographical distribution of the anopheline microbiota based on a meta-analysis of reported taxa. Mem Inst Oswaldo Cruz. 2014;109(5):672–84.
Segata N, Baldini F, Pompon J, Garrett WS, Truong DT, Dabiré RK, et al. The reproductive tracts of two malaria vectors are populated by a core microbiome and by gender-and swarm-enriched microbial biomarkers. Sci Rep. 2016;6:24207.
Kalappa DM, Subramani PA, Basavanna SK, Ghosh SK, Sundaramurthy V, Sreehari Uragayala S, et al. Influence of midgut microbiota in Anopheles stephensi on Plasmodium berghei infections. Malar J. 2018;17:385.
Tainchum K, Dupont C, Chareonviriyaphap T, Jumas-Bilak E, Bangs MJ, Manguin S. Bacterial microbiome in wild-caught Anopheles mosquitoes in Western Thailand. Front Microbiol. 2020;11:965.
Nilsson LKJ, de Oliveira MR, Marinotti O, Rocha EM, Håkansson S, Tadei WP, et al. Characterization of bacterial communities in breeding waters of Anopheles darlingi in Manaus in the Amazon Basin malaria-endemic area. Microb Ecol. 2019;78:781–91.
Muturi EJ, Ramirez JL, Rooney AP, Kim CH. Comparative analysis of gut microbiota of mosquito communities in central Illinois. PLoS Negl Trop Dis. 2017;11(2):e0005377.
Wang Y, Gilbreath TM III, Kukutla P, Yan G, Xu J. Dynamic gut microbiome across life history of the malaria mosquito Anopheles gambiae in Kenya. PLoS ONE. 2011;6(9):e24767.
Favia G, Ricci I, Damiani C, Raddadi N, Crotti E, Marzorati M, et al. Bacteria of the genus Asaia stably associate with Anopheles stephensi, an Asian malarial mosquito vector. Proc Natl Acad Sci USA. 2007;104:9047–51.
Crotti E, Damiani C, Pajoro M, Gonella E, Rizzi A, Ricci I, et al. Asaia, a versatile acetic acid bacterial symbiont, capable of cross-colonizing insects of phylogenetically distant genera and orders. Environ Microbiol. 2009;11(12):3252–64.
Chouaia B, Rossi P, Montagna M, Ricci I, Crotti E, Damiani C, et al. Molecular evidence for multiple infections as revealed by typing of Asaia bacterial symbionts of four mosquito species. Appl Environ Microbiol. 2010;76(22):7444–50.
Damiani C, Ricci I, Crotti E, Rossi P, Rizzi A, Scuppa P, et al. Mosquito-bacteria symbiosis: the case of Anopheles gambiae and Asaia. Microb Ecol. 2010;60:644–54.
De Freece C, Damiani C, Valzano M, D’amelio S, Cappelli A, Ricci I, et al. Detection and isolation of the α-proteobacterium Asaia in Culex mosquitoes. Med Vet Entomol. 2013;28(4):438–42.
Manguin S, Ngo CT, Tainchum K, Juntarajumnong W, Chareonviriyaphap T, Michon AL, et al. Bacterial biodiversity in midguts of Anopheles mosquitoes, malaria vectors in Southeast Asia. In: Manguin S, et al., editors. Anopheles mosquitoes: new insights into malaria vectors. IntechOpen: Croatia; 2013. p. 549–76.
Ramos-Nino ME, Fitzpatrick DM, Eckstrom KM, Tighe S, Hattaway LM, Hsueh AN, et al. Metagenomic analysis of Aedes aegypti and Culex quinquefasciatus mosquitoes from Grenada West Indies. PLoS ONE. 2020;15(4):e0231047.
Favia G, Ricci I, Marzorati M, Negri I, Alma A, Sacchi L, et al. Bacteria of the genus Asaia: a potential paratransgenic weapon against malaria. Adv Exp Med Biol. 2008;627:49–59.
Alonso DP, Mancini MV, Damiani C, Cappelli A, Ricci I, Alvarez MVN, et al. Genome reduction in the mosquito symbiont Asaia. Genome Biol Evol. 2019;11(1):1–10.
Mancini MV, Damiani C, Short SM, Cappelli A, Ulissi U, Capone A, et al. Inhibition of Asaia in adult mosquitoes causes male-specific mortality and diverse transcriptome changes. Pathogens. 2020;9(5):380.
Bongio NJ, Lampe DJ. Inhibition of Plasmodium berghei development in mosquitoes by effector proteins secreted from Asaia sp. bacteria using a novel native secretion signal. PLoS ONE. 2015;10(12):e0143541.
Cappelli A, Damiani C, Mancini MV, Valzano M, Rossi P, Serrao A, et al. Asaia activates immune genes in mosquito eliciting an anti-Plasmodium response: implications in malaria control. Front Genet. 2019;10:836.
Rossi P, Ricci I, Cappelli A, Damiani C, Ulissi U, Mancini MV, et al. Mutual exclusion of Asaia and Wolbachia in the reproductive organs of mosquito vectors. Parasit Vectors. 2015;8:278.
Capone A, Ricci I, Damiani C, Mosca M, Rossi P, Scuppa P. Interactions between Asaia, Plasmodium and Anopheles: new insights into mosquito symbiosis and implications in malaria symbiotic control. Parasit Vectors. 2013;6(1):182.
Terenius O, De Oliveira CD, Pinheiro WD, Tadei NP, James AA, Marinotti O. 16S rRNA gene sequences from bacteria associated with adult Anopheles darlingi (Diptera : Culicidae) mosquitoes. J Med Entomol. 2008;45:1725.
Dinparast Djadid N, Jazayeri H, Raz A, Favia G, Ricci I, Zakeri S. Identification of the midgut microbiota of An. stephensi and An. maculipennis for their application as a paratransgenic tool against malaria. PLoS ONE. 2011;6(12):e28484.
Correa LV. Estudo do Potencial Paratransgênico de Bactérias Cultiváveis Associadas ao Anopheles darlingi Root, 1926, Para Controle da Malária. PhD thesis. Manaus: PGSS–Biotecnologia e Recursos Naturais da Amazônia (Mestrado), Universidade do Estado do Amazonas; 2019. http://repositorioinstitucional.uea.edu.br//handle/riuea/2301.
Pumpuni CB, Demaio J, Kent M, Davis JR, Beier JC. Bacterial population dynamics in three anopheline species: the impact on Plasmodium sporogonic development. Am J Trop Med Hyg. 1996;54:214–8.
Riehle MA, Moreira CK, Lampe D, Lauzon C, Jacobs-Lorena M. Using bacteria to express and display anti-Plasmodium molecules in the mosquito midgut. Int J Parasitol. 2007;37(6):595–603.
Bisi DC, Lampe DJ. Secretion of anti-Plasmodium effector proteins from a natural Pantoea agglomerans isolate by using PelB and HlyA secretion signals. Appl Environ Microbiol. 2011;13:4669–75.
Mendiola SY, Civitello DJ, Gerardo NM. An integrative approach to symbiont-mediated vector control for agricultural pathogens. Curr Opin Insect Sci. 2020;39:57–62.
Dutkiewicz J, Mackiewicz B, Kinga Lemieszek M, Golec M, Milanowski J. Pantoea agglomerans: a mysterious bacterium of evil and good. Part III. Deleterious effects: infections of humans, animals and plants. Ann Agric Environ Med. 2016;23(2):197–205.
Büyükcam A, Tuncer Ö, Gür D, Sancak B, Ceyhan M, Cengiz AB, et al. Clinical and microbiological characteristics of Pantoea agglomerans infection in children. J Infect Public Health. 2018;11(3):304–9.
Kozlova EV, Hegde S, Roundy CM, Golovko G, Saldaña MA, Hart CE. Microbial interactions in the mosquito gut determine Serratia colonization and blood-feeding propensity. ISME J. 2021;15:93–108.
Rocha EM. Seleção de espécies bacterianas cultiváveis, simbiontes de Anopheles darlingi (Root, 1926), para o controle da malária por paratransgênese. 2020. PhD thesis. Manaus: Programa de Pós-Graduação em Biotecnologia da Universidade Federal do Amazonas (PPGBIOTEC-UFAM). https://tede.ufam.edu.br/bitstream/tede/7737/5/Tese_ElersonRocha_PPGBiotec.pdf.
Bahia AC, Dong Y, Blumberg BJ, Mlambo G, Tripathi A, BenMarzouk-Hidalgo OJ, et al. Exploring Anopheles gut bacteria for Plasmodium blocking activity. Environ Microbiol. 2014;16:2980–94.
Bai L, Wang L, Vega-Rodriguez J, Wang G, Wang S. A gut symbiotic bacterium Serratia marcescens renders mosquito resistance to Plasmodium infection through activation of mosquito immune responses. Front Microbiol. 2019;10:1580.
Bando H, Okado K, Guelbeogo WM, Badolo A, Aonuma H, Nelson B, Fukumoto S, et al. Intra-specific diversity of Serratia marcescens in Anopheles mosquito midgut defines Plasmodium transmission capacity. Sci Rep. 2013;3:1641.
Chen S, Blom J, Walker ED. Genomic, physiologic, and symbiotic characterization of Serratia marcescens strains isolated from the mosquito Anopheles stephensi. Front Microbiol. 2017;8:1483.
Krafsur AM, Ghosh A, Brelsfoard CL. Phenotypic response of Wolbachia pipientis in a cell-free medium. Microorganisms. 2020;8:1060.
Reveillaud J, Bordenstein SR, Cruaud C, Shaiber A, Esen OC, Mylène Weill M, et al. The Wolbachia mobilome in Culex pipiens includes a putative plasmid. Nat Commun. 2019;10:1051.
Koosha M, Vatandoost H, Karimian F, Choubdar N, Aba MRI, Oshaghi MA. Effect of Serratia AS1 (Enterobacteriaceae: Enterobacteriales) on the fitness of Culex pipiens (Diptera: Culicidae) for paratransgenic and RNAi approaches. J Med Entomol. 2019;56(2):553–9.
Koosha M, Vatandoost H, Karimian F, Choubdar N, Oshaghi MA. Delivery of a genetically marked Serratia AS1 to medically important arthropods for use in RNAi and paratransgenic control strategies. Microb Ecol. 2019;78(1):185–94.
Cirimotich CM, Dong Y, Clayton AM, Sandiford SL, Souza-Neto JA, Mulenga M, et al. Natural microbe-mediated refractoriness to Plasmodium infection in Anopheles gambiae. Science. 2011;332(6031):855–8.
Dennison NJ, Saraiva RG, Cirimotich CM, Mlambo G, Mongodin EF, Dimopoulos G. Functional genomic analyses of Enterobacter, Anopheles and Plasmodium reciprocal interactions that impact vector competence. Malar J. 2016;15(1):425.
Dehghan H, Oshaghi MA, Moosa-Kazemi SH, Yakhchali B, Vatandoost H, Maleki-Ravasan N, et al. Dynamics of transgenic Enterobacter cloacae expressing green fluorescent protein defensin (GFP-D) in Anopheles stephensi under laboratory condition. J Arthropod Borne Dis. 2017;11(4):515–32.
Gnambani EJ, Bilgo E, Sanou A, Dabiré RK, Diabaté A. Infection of highly insecticide-resistant malaria vector Anopheles coluzzii with entomopathogenic bacteria Chromobacterium violaceum reduces its survival, blood feeding propensity and fecundity. Malar J. 2020;19:352.
Saraiva RG, Fang J, Kang S, Anglero-Rodrıguez YI, Dong Y, Dimopoulos G. Aminopeptidase secreted by Chromobacterium sp. Panama inhibits dengue virus infection by degrading the E protein. PLoS Negl Trop Dis. 2018;12(4):e0006443.
Chen S, Bagdasarian M, Walker ED. Elizabethkingia anophelis: molecular manipulation and interactions with mosquito hosts. Appl Environ Microbiol. 2015;81(6):2233–43.
Frank T, Gody JC, Nguyen LB, Berthet N, Le Fleche-Mateos A, Bata P, et al. First case of Elizabethkingia anophelis meningitis in the Central African Republic. Lancet. 2013;381(9880):1876.
Khaligh FG, Vahedi M, Chavshin AR. Identification of symbiotic bacteria in the midgut of the medically important mosquito, Culiseta longiareolata (Diptera: Culicidae). BMC Res Notes. 2020;13:378.
Gurung K, Wertheim B, Salles JF. The microbiome of pest insects: it is not just bacteria. Entomol Experim Appl. 2019;167:156–70.
Giesbrecht D, Heschuk D, Wiens I, Boguski D, LaChance P, Whyard S. RNA interference is enhanced by knockdown of double-stranded RNases in the yellow fever mosquito Aedes aegypti. Insects. 2020;11:327.
Christiaens O, Niu J, Taning CNT. RNAi in insects: a revolution in fundamental research and pest control applications. Insects. 2020;11(7):415.
Gurgel-Gonçalves R, Komp E, Campbell LP, Khalighifar A, Mellenbruch J, Mendonc VJ, et al. Automated identification of insect vectors of Chagas disease in Brazil and Mexico: the Virtual Vector Lab. Peer J. 2017;5:e3040.
Teixeira ARL, Nascimento RJ, Sturm NR. Evolution and pathology in Chagas disease: a review. Mem Inst Oswaldo Cruz. 2006;101(5):463–91.
Azambuja P, Garcia ES, Ratcliffe NA. Gut microbiota and parasite transmission by insect vectors. Trends Parasitol. 2005;21(12):568–72.
Garcia ES, Castro DP, Figueiredo MB, Azambuja P. Immune homeostasis to microorganisms in the guts of triatomines (Reduviidae)—a review. Mem Inst Oswaldo Cruz. 2010;105:605–10.
Castro DP, Moraes CS, Gonzalez MS, Ratcliffe NA, Azambuja P, Garcia ES. Trypanosoma cruzi immune response modulation decreases microbiota in Rhodnius prolixus gut and is crucial for parasite survival and development. PLoS ONE. 2012;7:e36591.
Azambuja P, Garcia ES, Waniek PJ, Vieira CS, Figueiredo MB, Gonzalez MS, et al. Rhodnius prolixus: from physiology by Wigglesworth to recent studiesof immune system modulation by Trypanosoma cruzi and Trypanosoma rangeli. J Insect Physiol. 2017;97:45–65.
Rodríguez-Ruano SM, Škochová V, Rego ROM, Schmidt JO, Roachell W, Hypša V, et al. Microbiomes of North American triatominae: the grounds for Chagas disease epidemiology. Front Microbiol. 2018;9:1167.
Salcedo-Porras N, Umaña-Diaz C, Bitencourt ROB, Lowenberger C. The role of bacterial symbionts in triatomines: an evolutionary perspective. Microorganisms. 2020;8:1438.
Schaub GA. An update on the knowledge of parasite-vector interactions of Chagas disease. Res Rep Trop Med. 2021;12:63–76.
Guarneri AA, Schaub GA. Interaction of triatomines, trypanosomes and microbiota. In: Guarneri AA, Lorenzo MG, editors. Triatominae—the biology of Chagas disease vectors. New York: Springer Nature; 2021. p. 345–86.
Traverso L, Lavore A, Sierra I, Palacio V, Martinez-Barnetche J, Latorre-Estivalis JM, et al. Comparative and functional triatomine genomics reveals reductions and expansions in insecticide resistance-related gene families. PLoS Negl Trop Dis. 2017;11(2):e0005313.
Lima MS, Laport MS, Lorosa ES, Jurberg J, dos Santos KRN, da Silva Neto MAC, et al. Bacterial community composition in the salivary glands of triatomines (Hemiptera: Reduviidae). PLoS Negl Trop Dis. 2018;12(9):e0006739.
Da Mota FF, Marinho LP, Moreira CJ, Lima MM, Mello CB, Garcia ES, et al. Cultivation-independent methods reveal differences among bacterial gut microbiota in triatomine vectors of Chagas disease. PLoS Negl Trop Dis. 2012;6(5):e1631.
Gumiel M, da Mota FF, de Sousa RV, Sarquis O, Castro DP, Lima MM, et al. Characterization of the microbiota in the guts of Triatoma brasiliensis and Triatoma pseudomaculata infected by Trypanosoma cruzi in natural conditions using culture independent methods. Parasit Vectors. 2015;8:245.
Díaz S, Villavicencio B, Correia N, Costa J, Haag KL. Triatomine bugs, their microbiota and Trypanosoma cruzi: asymmetric responses of bacteria to an infected blood meal. Parasit Vectors. 2016;9:636.
Carels N, Gumiel M, da Mota FF, de Carvalho Moreira CJ, Azambuja P. A metagenomic analysis of bacterial microbiota in the digestive tract of triatomines. Bioinform Biol Insights. 2017;27(11):1177932217733422.
Lopez-Ordonez T, Flores-Lopez CA, Montejo-Lopez R, Cruz-Hernandez A, Conners EE. Cultivable bacterial diversity in the gut of the Chagas disease vector Triatoma dimidiata: identification of possible bacterial candidates for a paratransgenesis approach. Front Ecol Evol. 2018;5:174.
Oliveira JL, Cury JC, Gurgel-Goncalves R, Bahia AC, Monteiro FA. Field-collected Triatoma sordida from central Brazil display high microbiota diversity that varies with regard to developmental stage and intestinal segmentation. PLoS Negl Trop Dis. 2018;12(8):e0006709.
Orantes LC, Monroy C, Dorn PL, Stevens L, Rizzo DM, Morrissey L, et al. Uncovering vector, parasite, blood meal and microbiome patterns from mixed-DNA specimens of the Chagas disease vector Triatoma dimidiata. PLoS Negl Trop Dis. 2018;12(10):e0006730.
Dumonteil E, Ramirez-Sierra MJ, Pérez-Carrillo S, Teh-Poot C, Herrera C, Gourbière S, et al. Detailed ecological associations of triatomines revealed by metabarcoding and next-generation sequencing: implications for triatomine behavior and Trypanosoma cruzi transmission cycles. Sci Rep. 2018;8:4140.
Montoya-Porras LM, Omar T-C, Alzate JF, Moreno-Herrera CX, Cadavid-Restrepo GE. 16S rRNA gene amplicon sequencing reveals dominance of Actinobacteria in Rhodnius pallescens compared to Triatoma maculate midgut microbiota in natural populations of vector insects from Colombia. Acta Trop. 2018;178:327–32.
Waltmann A, Willcox AC, Balasubramanian S, Mayori KB, Guerrero SM, Sanchez RSS, et al. Hindgut microbiota in laboratory-reared and wild Triatoma infestans. PLoS Negl Trop Dis. 2019;13(5):e0007383.
Arias-Giraldo LM, Muñoz M, Hernández C, Herrera G, Velásquez-Ortiz N, Cantillo-Barraza O, et al. Species-dependent variation of the gut bacterial communities across Trypanosoma cruzi insect vectors. PLoS ONE. 2020;15(11):e0240916.
Brown JJ, Rodríguez-Ruano SM, Poosakkannu A, Batani G, Schmidt JO, Roachell W, et al. Ontogeny, species identity, and environment dominate microbiome dynamics in wild populations of kissing bugs (Triatominae). Microbiome. 2020;8:146.
Hu Y, Xie H, Gao M, Huang P, Zhou H, Ma Y, et al. Dynamic of composition and diversity of gut microbiome in Triatoma rubrofasciata in different developmental stages and environmental conditions. Front Cell Infect Microbiol. 2020;10: 587708.
Mann AE, Mitchell EA, Zhang Y, Curtis-Robles R, Thapa S, Hamer SA, et al. Comparison of the bacterial gut microbiome of North American Triatoma spp. with and without Trypanosoma cruzi. Front Microbiol. 2020;11:364.
Duarte-Silva E, Morais LH, Clarke G, Savino W, Peixoto C. Targeting the gut microbiota in Chagas disease: what do we know so far? Front Microbiol. 2020;11: 585857.
Schaub GA. Intestinal bacteria/mutualistic symbionts of triatomines—a review. Mitt Dtsch Ges Allg Angew Entomol. 2020;2020(22):191–4.
Durvasula RV, Sundaram RK, Kirsch P, Hurwitz I, Crawford CV, Dotson E, et al. Genetic transformation of a Corynebacterial symbiont from the Chagas disease vector Triatoma infestans. Exp Parasitol. 2008;119(1):94–8.
Jose C, Klein N, Wyss S, Fieck A, Hurwitz I, Durvasula R. Recombinant Arthrobacter β-1,3-glucanase as a potential effector molecule for paratransgenic control of Chagas disease. Parasit Vectors. 2013;6:65.
Markiv A, Anani B, Durvasula RV, Kang AS. Module based antibody engineering: a novel synthetic REDantibody. J Immunol Methods. 2011;364(1–2):40–9.
Gonzalez MS, Souza MS, Garcia ES, Nogueira NFS, Mello CB, Cánepa GE, et al. Trypanosoma cruzi TcSMUG L-surface mucins promote development and infectivity in the triatomine vector Rhodnius prolixus. PLoS Negl Trop Dis. 2013;7(11): e2552.
Giorgi ME, de Lederkremer RM. The glycan structure of T. cruzi mucins depends on the host. Insights on the chameleonic galactose. Molecules. 2020;25(17):3913.
Whitten MMA. Novel RNAi delivery systems in the control of medical and veterinary pests. Curr Opin Insect Sci. 2019;34:1–6.
Aksoy S. Control of tsetse flies and trypanosomes using molecular genetics. Vet Parasitol. 2003;115:125–45.
Holanda-Freitas IT, Cupertino MC, dos Santos EC, Oliveira L, Geller M, Siqueira-Batista R. Human African trypanosomiasis: current standing and challenges. J Trop Pathol. 2020;49(3):133–48.
Bateta R, Wang J, Wu Y, Weiss BL, Warren WC, Murilla GA, et al. Tsetse fly (Glossina pallidipes) midgut responses to Trypanosoma brucei challenge. Parasit Vectors. 2017;10:614.
Aksoy S. Tsetse—a haven for microorganisms. Parasitol Today. 2000;16(3):114–8.
Geiger A, Fardeau ML, Grebaut P, Vatunga G, Josénando T, Herder S, et al. First isolation of Enterobacter, Enterococcus, and Acinetobacter spp. as inhabitants of the tsetse fly (Glossina palpalis palpalis) midgut. Infect Gene Evol. 2009;9:1364–70. https://doi.org/10.1016/j.meegid.2009.09.013.
Geiger A, Fardeau ML, Njiokou F, Joseph M, Asonganyi T, Ollivier B, et al. Bacterial diversity associated with populations of Glossina spp. from Cameroon and distribution within the Campo sleeping sickness focus. Microb Ecol. 2011;62:632–43.
Geiger A, Fardeau ML, Njiokou F, Ollivier B. Glossina spp. gut bacterial flora and their putative role in fly-hosted trypanosome development. Front Cell Infect Microbiol. 2013;3:34.
Lindh JM, Lehane MJ. The tsetse fly Glossina fuscipes fuscipes (Diptera: Glossina) harbours a surprising diversity of bacteria other than symbionts. Antonie Van Leeuwenhoek. 2011;99:711–20.
Wang J, Weiss BL, Aksoy S. Tsetse fly microbiota: form and function. Front Cell Infect Microbiol. 2013;3:69.
Weiss BL, Mouchotte R, Rio RV, Wu YN, Wu Z, Heddi A, et al. Interspecific transfer of bacterial endosymbionts between tsetse fly species: infection establishment and effect on host fitness. Appl Environ Microbiol. 2006;72(11):7013–21.
Aksoy E, Telleria EL, Echodu R, Wu Y, Okedi LM, Weiss BL, et al. Analysis of multiple tsetse fly populations in Uganda reveals limited diversity and species-specifc gut microbiota. Appl Environ Microbiol. 2014;80:4301–12.
Jacob F, Melachio TT, Njitchouang GR, Gimonneau G, Njiokou F, Abate L, et al. Intestinal bacterial communities of trypanosome-infected and uninfected Glossina palpalis palpalis from three human African trypanosomiasis foci in Cameroon. Front Microbiol. 2017;8:1464.
Griffith BC, Weiss BL, Aksoy E, Mireji PO, Auma JE, Wamwiri FR, et al. Analysis of the gut-specific microbiome from field-captured tsetse flies, and its potential relevance to host trypanosome vector competence. BMC Microbiol. 2018;18(1):146.
Kame-Ngasse GI, Njiokou F, Melachio-Tanekou TT, Farikou O, Simo G, Geiger A. Prevalence of symbionts and trypanosome infections in tsetse flies of two villages of the “Faro and Déo” division of the Adamawa region of Cameroon. BMC Microbiol. 2018;18:159.
Malele I, Nyingilili H, Lyaruu E, Tauzin M, Ollivier BB, Cayol JL, et al. Bacterial diversity obtained by culturable approaches in the gut of Glossina pallidipes population from a non-sleeping sickness focus in Tanzania: preliminary results. BMC Microbiol. 2018;18(Suppl 1):164.
Odeniran PO, Macleod EW, Ademola IOA, Welburn SC. Endosymbionts interaction with trypanosomes in Palpalis group of Glossina captured in southwest Nigeria. Parasitol Int. 2019;70:64–9.
Ngoune JMT, Reveillaud J, Sempere G, Njiokou F, Melachio TT, Abate L, et al. The composition and abundance of bacterial communities residing in the gut of Glossina palpalis palpalis captured in two sites of southern Cameroon. Parasit Vectors. 2019;12(1):151.
Attardo GM, Scolari F, Malacrida A. Bacterial symbionts of tsetse flies: relationships and functional interactions between tsetse flies and their symbionts. In: Kloc M, editor. Symbiosis: cellular, molecular, medical and evolutionary aspects. Results and problems in cell differentiation. Cham: Springer; 2020. p. 69. https://doi.org/10.1007/978-3-030-51849-3_19.
Gaithuma A, Yamagishi J, Hayashida K, Kawai N, Namangala B, Sugimoto C. Blood meal sources and bacterial microbiome diversity in wild-caught tsetse flies. Sci Rep. 2020;10:5005.
Balmand S, Lohs C, Aksoy S, Heddi A. Tissue distribution and transmission routes for the tsetse fly endosymbionts. J Invertebr Pathol. 2013;112(Suppl):S116–22.
Doudoumis V, Blow F, Saridaki A, Augustinos A, Dyer NA, Goodhead I, et al. Challenging the Wigglesworthia, Sodalis, Wolbachia symbiosis dogma in tsetse flies: Spiroplasma is present in both laboratory and natural populations. Sci Rep. 2017;7:4699.
Masson F, Calderon Copete S, Schüpfer F, Garcia-Arraez G, Lemaitre B. In vitro culture of the insect endosymbiont Spiroplasma poulsonii highlights bacterial genes involved in host-symbiont interaction. MBio. 2018;9(2):e00024-18.
Schneider DI, Saarman N, Onyango MG, Hyseni C, Opiro R, Echodu R, et al. Spatial-temporal distribution of Spiroplasma infection in the tsetse fly (Glossina fuscipes fuscipes) in northern Uganda. PLoS Negl Trop Dis. 2019;13(8): e0007340.
Soumana IH, Simo G, Njiokou F, Tchicaya B, Abd-Alla AM, Cuny G, et al. The bacterial flora of tsetse fly midgut and its effect on trypanosome transmission. J Invertebr Pathol. 2013;112(Suppl):89–93.
Welburn SC, Maudlin I, Ellis DS. In vitro cultivation of rickettsia-like-organisms from Glossina spp. Ann Trop Med Parasitol. 1987;81(3):331–5.
Beard CB, O’Neill SL, Mason P, Mandelco L, Woese CR, Tesh RB, et al. Genetic transformation and phylogeny of bacterial symbionts from tsetse. Insect Mol Biol. 1993;1:123–31.
Cheng Q, Aksoy S. Tissue tropism, transmission and expression of foreign genes in vivo in midgut symbionts of tsetse flies. Insect Mol Biol. 1999;8:125–32.
Aksoy S, Weiss B, Attardo G. Paratransgenesis applied for control of tsetse transmitted sleeping sickness. Adv Exp Med Biol. 2008;627:35–48.
De Vooght L, Caljon G, Stijlemans B, De Baetselier P, Coosemans M, Van den Abbeele J. Expression and extracellular release of a functional anti-trypanosome Nanobody® in Sodalis glossinidius, a bacterial symbiont of the tsetse fly. Microb Cell Fact. 2012;11:23.
De Vooght L, Caljon G, De Ridder K, Van Den Abbeele J. Delivery of a functional anti-trypanosome Nanobody in different tsetse fly tissues via a bacterial symbiont, Sodalis glossinidius. Microb Cell Fact. 2014;13:156.
De Vooght L, Van Keer S, Van Den Abbeele J. Towards improving tsetse fly paratransgenesis: stable colonization of Glossina morsitans morsitans with genetically modified Sodalis. BMC Microbiol. 2018;18:165.
Medlock J, Atkins KE, Thomas DN, Aksoy S, Galvani AP. Evaluating paratransgenesis as a potential control strategy for African trypanosomiasis. PLoS Negl Trop Dis. 2013;7(8):e2374.
Gilbert JA, Medlock J, Townsend JP, Aksoy S, Ndeffo Mbah M, Galvani AP. Determinants of human African trypanosomiasis elimination via paratransgenesis. PLoS Negl Trop Dis. 2016;10(3):e0004465.
Demirbas-Uzel G, De Vooght L, Parker AG, Vreysen MJB, Mach RL, Van Den Abbeele J, et al. Combining paratransgenesis with SIT: impact of ionizing radiation on the DNA copy number of Sodalis glossinidius in tsetse flies. BMC Microbiol. 2018;18:160.
Hall RJ, Flanagan LA, Bottery MJ, Springthorpe V, Thorpe S, Darby AC, et al. A tale of three species: adaptation of Sodalis glossinidius to tsetse biology, Wigglesworthia metabolism, and host diet. MBio. 2019;10: e02106-18.
Dale C, Welburn SC. The endosymbionts of tsetse flies: manipulating host–parasite interactions. Int J Parasitol. 2001;31:628–31.
Farikou O, Njiokou F, Mbida Mbida JA, Njitchouang GR, Djeunga HN, Asonganyi T, et al. Tripartite interactions between tsetse flies, Sodalis glossinidius and trypanosomes an epidemiological approach in two historical human African trypanosomiasis foci in Cameroon. Infect Genet Evol. 2010;10:115–21.
Makhulu EE, Villinger J, Adunga VO, Jeneby MM, Kimathi EM, Mararo E, et al. Tsetse blood-meal sources, endosymbionts and trypanosome-associations in the Maasai Mara National Reserve, a wildlife-human-livestock interface. PLoS Negl Trop Dis. 2021;15(1):e0008267.
Kendra CG, Keller CM, Bruna RE, Pontes MH. Conjugal DNA transfer in Sodalis glossinidius, a maternally inherited symbiont of tsetse flies. mSphere. 2020;5(6):e00864-2.
Hu Y, Aksoy S. An antimicrobial peptide with trypanocidal activity characterized from Glossina morsitans morsitans. Insect Biochem Mol Biol. 2005;35:105–15.
Stijlemans B, Caljon G, Natesan SK, Saerens D, Conrath K, Pérez-Morga D, et al. High affinity nanobodies against the Trypanosome brucei VSG are potent trypanolytic agents that block endocytosis. PLoS Pathog. 2011;7(6): e1002072.
Dale C, Young SA, Haydon DT, Welburn SC. The insect endosymbiont S. glossinidius utilizes a type III secretion system for cell invasion. Proc Natl Acad Sci USA. 2001;98:1883–8.
Alkan C, Bichaud L, de Lamballerie X, Alten B, Gould EA, Charrel RN. Sandfly-borne phleboviruses of Eurasia and Africa: epidemiology, genetic diversity, geographic range, control measures. Antivir Res. 2013;100(1):54–74.
Ayhan N, Charrel RN. Sandfly-borne viruses of demonstrated/relevant medical importance. In: Vectors and vector-borne zoonotic diseases. London: IntechOpen; 2018. https://doi.org/10.5772/intechopen.81023.
Momeni AZ, Jalayer T, Emamjomeh M, Khamesipour A, Zicker F, Ghassemi RL, et al. A randomised, double-blind, controlled trial of a killed L. major vaccine plus BCG against zoonotic cutaneous leishmaniasis in Iran. Vaccine. 1999;17:466–72.
Ashwin H, Sadlova J, Vojtkova B, Becva T, Lypaczewski P, Schwartz E, et al. Characterization of a new Leishmania major strain for use in a controlled human infection model. Nat Commun. 2021;12:115.
Gomes B, Purkait B, Deb RM, Rama A, Singh RP, Foster GM, et al. Knockdown resistance mutations predict DDT resistance and pyrethroid tolerance in the visceral leishmaniasis vector Phlebotomus argentipes. PLoS Negl Trop Dis. 2017;11(4):e0005504.
Burza S, Croft SL, Boelaert M. Leishmaniasis. Lancet. 2018;392(10151):951–70.
Wijerathna T, Gunathunga S, Gunathilaka HN. Recent developments and future directions in the paratransgenesis based control of Leishmania transmission. Biol Control. 2020;145: 104260.
Gouveia C, Asensi MD, Zahner V, Rangel EF, de Oliveira SMP. Estudo sobre as bactérias da microbiota intestinal de diferentes populações brasileiras de Lutzomyia longipalpis (Lutz & Neiva) (Diptera: Psychodidae). Neotrop Entomol. 2008;37(5):597–601.
McCarthy CB, Diambra LA, Rivera Pomar RV. Metagenomic analysis of taxa associated with Lutzomyia longipalpis, vector of visceral leishmaniasis, using an unbiased high-throughput approach. PLoS Negl Trop Dis. 2011;5(9): e1304.
McCarthy CB, Santini MS, Pimenta PFP, Diambra LA. First comparative transcriptomic analysis of wild adult male and female Lutzomyia longipalpis, vector of visceral Leishmaniasis. PLoS ONE. 2013;8(3):e58645.
Sant’Anna MRV, Darby AC, Brazil RP, Montoya-Lerma J, Dillon VM, Bates PA, et al. Investigation of the bacterial communities associated with females of Lutzomyia sand fly species from South America. PLoS ONE. 2012;7(8):e42531.
Pires ACAM, Villegas LEM, Campolina TB, Orfanó AS, Pimenta PFP, Secundino NFC. Bacterial diversity of wild-caught Lutzomyia longipalpis (a vector of zoonotic visceral leishmaniasis in Brazil) under distinct physiological conditions by metagenomics analysis. Parasit Vectors. 2017;10:627.
Telleria EL, Martins-da-Silva A, Tempone AJ, Traub-Csekö YM. Leishmania, microbiota and sand fly immunity. Parasitology. 2018;145:1336–53.
Vivero RJ, Villegas-Plazas M, Cadavid-Restrepo GE, Herrera CXM, Uribe SI, Junca H. Wild specimens of sand fly phlebotomine Lutzomyia evansi, vector of leishmaniasis, show high abundance of Methylobacterium and natural carriage of Wolbachia and Cardinium types in the midgut microbiome. Sci Rep. 2019;9:17746.
Tabbabi A, Watanabe S, Mizushima D, Caceres AG, Gomez EA, Yamamoto DS, et al. Comparative analysis of bacterial communities in Lutzomyia ayacuchensis populations with different vector competence to Leishmania parasites in Ecuador and Peru. Microorganisms. 2020;9(1):68.
Dillon RJ, El Kordy E, Shehata M, Lane RP. The prevalence of a microbiota in the digestive tract of Phlebotomus papatasi. Ann Trop Med Parasitol. 1996;90(6):669–73.
Volf P, Kiewegová A, Nemec A. Bacterial colonisation in the gut of Phlebotomus duboseqi (Diptera: Psychodidae): transtadial passage and the role of female diet. Folia Parasitol. 2002;49(1):73–7.
Hillesland H, Read A, Subhadra B, Hurwitz I, McKelvey R, Ghosh K, et al. Identification of aerobic gut bacteria from the kala azar vector, Phlebotomus argentipes: a platform for potential paratransgenic manipulation of sand flies. Am J Trop Med Hyg. 2008;79:881–6.
Mukhopadhyay J, Braig HR, Rowton ED, Ghosh K. Naturally occurring culturable aerobic gut flora of adult Phlebotomus papatasi, vector of Leishmania major in the Old World. PLoS ONE. 2012;7(5):e35748.
Maleki-Ravasan N, Oshaghi MA, Afshar D, Arandian MH, Hajikhani S, Akhavan AA, et al. Aerobic bacterial flora of biotic and abiotic compartments of a hyperendemic zoonotic cutaneous leishmaniasis (ZCL) focus. Parasit Vectors. 2015;8:63.
Li K, Chen H, Jiang J, Li X, Xu J, Ma Y. Diversity of bacteriome associated with Phlebotomus chinensis (Diptera: Psychodidae) sand flies in two wild populations from China. Sci Rep. 2016;6:36406.
Fraihi W, Fares W, Perrin P, Dorkeld F, Sereno D, Barhoumi W, et al. An integrated overview of the midgut bacterial flora composition of Phlebotomus perniciosus, a vector of zoonotic visceral leishmaniasis in the Western Mediterranean Basin. PLoS Negl Trop Dis. 2017;11(3): e0005484.
Abbasi I, Nasereddin A, Warburg A. Development of a next generation DNA sequencing-based multi detection assay for detecting and identifying Leishmania parasites, blood sources, plant meals and intestinal microbiome in phlebotomine sand flies. Acta Trop. 2019;199: 105101.
Karimian F, Vatandoost H, Rassi Y, Maleki-Ravasan N, Mohebali M, Shirazi MH, et al. Aerobic midgut microbiota of sand fly vectors of zoonotic visceral leishmaniasis from northern Iran, a step toward finding potential paratransgenic candidates. Parasit Vectors. 2019;12(1):10.
Gunathilaka N, Perera H, Wijerathna T, Rodrigo W, Wijegunawardana NDAD. The diversity of midgut bacteria among wild-caught Phlebotomus argentipes (Psychodidae: Phlebotominae), the vector of leishmaniasis in Sri Lanka. Biomed Res Int. 2020;2020:5458063.
Papadopoulos C, Karas PA, Vasileiadis S, Ligda P, Saratsis A, Sotiraki S, et al. Host species determines the composition of the prokaryotic microbiota in Phlebotomus sandflies. Pathogens. 2020;9:428.
Omondi ZN, Demir S. Bacteria composition and diversity in the gut of sand fly: impact on Leishmania and sand fly development. Int J Trop Insect Sci. 2021;41:25–32.
Hurwitz I, Hillesland H, Fieck A, Das P, Durvasula R. The paratransgenic sand fly: a platform for control of Leishmania transmission. Parasit Vectors. 2011;4:82.
Pereira AV, de Barros G, Pinto EG, Tempone AG, Orsi Rde O, Dos Santos LD, et al. Melittin induces in vitro death of Leishmania (Leishmania) infantum by triggering the cellular innate immune response. J Venom Anim Toxins Incl Trop Dis. 2016;22:1.
Soto M, Requena JM, Quijada L, Perez MJ, Nieto CG, Guzman F, Patarroyo ME, Alonso C. Antigenicity of the Leishmania infantum histones H2B and H4 during canine viscerocutaneous leishmaniasis. Clin Exp Immunol. 1999;115(2):342–9.
Heerman M, Weng J-L, Hurwitz I, Durvasula R, Ramalho-Ortigao M. Bacterial infection and immune responses in Lutzomyia longipalpis sand fly larvae midgut. PLoS Negl Trop Dis. 2015;9(7):e0003923.
Coutinho-Abreu IV, Zhu KY, Ramalho-Ortigao M. Transgenesis and paratransgenesis to control insect-borne diseases: current status and future challenges. Parasitol Int. 2010;59(1):1–8.
Ogaugwu CE, Durvasula RV. Developing the arsenal against pest and vector dipterans: inputs of transgenic and paratransgenic biotechnologies. In: Shields VDC, editor. Biological control of pest and vector insects. London: InTechOpen; 2017. p. 325–47. https://doi.org/10.5772/66440.
Aguilera J, Gomes AR, Nielsen KM. Genetically modified microbial symbionts as arthropod pest controllers: risk assessment through the European legislations. J Appl Entomol. 2011;135:494–502.
European Food Safety Authority (EFSA). Guidance on the environmental risk assessment of genetically modified animals. EFSA J. 2013;11(5):3200.
Kawai M, Higashiura N, Hayasaki K, Okamoto N, Takami A, Hirakawa H, et al. Complete genome and gene expression analyses of Asaia bogorensis reveal unique responses to culture with mammalian cells as a potential opportunistic human pathogen. DNA Res. 2015;22(5):357–66.
Abreo E, Altier N. Pangenome of Serratia marcescens strains from nosocomial and environmental origins reveals different populations and the links between them. Sci Rep. 2019;9:46.
Sherid M, Samo S, Sulaiman S, Husein H, Sifuentes H, Sridhar S. Liver abscess and bacteremia caused by lactobacillus: role of probiotics? Case report and review of the literature. BMC Gastroenterol. 2016;16:138.
Araújo HRC, Carvalho DO, Ioshino RS, Costa-da-Silva AL, Capurro ML. Aedes aegypti control strategies in Brazil: incorporation of new technologies to overcome the persistence of dengue epidemics. Insects. 2015;6:576–94.
Wozniak CA, McClung G, Gagliardi J, Segal M, Matthews K. Chapter 4, regulation of genetically engineered microorganisms under FIFRA, FFDCA and TSCA. In: Wozniak CA, McHughen A, editors. Regulation of agricultural biotechnology: the United States and Canada. Dordrecht: Springer; 2013. p. 57–94. https://doi.org/10.1007/978-94-007-2156-2_4.
Environmental Protection Agency (EPA). TSCA environmental release application (TERA) for Alcaligenes xylosoxidans subspecies denitrificans strain AL6.1, R04-03, submitted by the University of California, Riverside. 2004. https://www.epa.gov/regulation-biotechnology-under-tsca-and-fifra/tsca-environmental-release-application-tera-4.
Ramsey JM, Bond JG, Macotela ME, Facchinelli L, Valerio L, Brown DM, et al. A regulatory structure for working with genetically modified mosquitoes: lessons from Mexico. PLoS Negl Trop Dis. 2014;8(3):e2623.
Meghani Z, Boëte C. Genetically engineered mosquitoes, Zika and other arboviruses, community engagement, costs, and patents: ethical issues. PLoS Negl Trop Dis. 2018;12(7):e0006501.
Flores HA, O’Neill SL. Controlling vector-borne diseases by releasing modified mosquitoes. Nat Rev Microbiol. 2018;16(8):508–18.
Singh JA. Informed consent and community engagement in open field research: lessons for gene drive science. BMC Med Ethics. 2019;20:54.
LaMotte S. Genetically engineered mosquitoes get EPA approval for Florida release despite objections from environmental groups. CNN News. 18 June 2020. https://edition.cnn.com/2020/06/18/health/genetically-engineered-mosquitoes-gmo-florida-scn-wellness/index.html.
Deutsch CA, Tewksbury JJ, Tigchelaarm M, Battisti DS, Merrill SC, Huey RB, et al. Increase in crop losses to insect pests in a warming climate. Science. 2018;361(6405):916–9.
Caminade C, McIntyre KM, Jones AE. Impact of recent and future climate change on vector-borne diseases. Ann N Y Acad Sci. 2019;1436(1):157–73.
Bourtzis K, Lees RS, Hendrichs J, Vreysen MJ. More than one rabbit out of the hat: Radiation, transgenic and symbiont-based approaches for sustainable management of mosquito and tsetse fly populations. Acta Trop. 2016;157:115–30.
World Health Organization (WHO). World malaria report 2020. www.who.int/teams/global-malaria-programme. Accessed 6 Jan 2021.
Acknowledgements
We gratefully thank PROPPi-UFF, Universidade Federal Fluminense, for the support provided for the development of this research.
Funding
This work was supported by CAPES processo No. 88881.068062/2014-01.
Author information
Authors and Affiliations
Contributions
NAR and JPFP wrote the main text which was subsequently edited and enlarged by the other co-authors who also provided the Figures. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that there is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ratcliffe, N.A., Furtado Pacheco, J.P., Dyson, P. et al. Overview of paratransgenesis as a strategy to control pathogen transmission by insect vectors. Parasites Vectors 15, 112 (2022). https://doi.org/10.1186/s13071-021-05132-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-021-05132-3