
Abstract
Background
Anaplasma platys is a bacterium parasitic in the canine platelets, representing the causative agent of canine cyclic thrombocytopenia, with a worldwide distribution, mainly in tropical countries. The agent has zoonotic potential, being reported in several human clinical cases. The suspected vector is the tick Rhipicephalus sanguineus (sensu lato), widely distributed in animals and humans in the tropical regions of South America, Africa, Asia and Australia, but also in southern Europe. Only few scattered data concerning the epidemiology of A. platys in sub-Saharan Africa are available. The aim of our study was to evaluate the epidemiological status of A. platys in dogs and cats from rural communities in eastern and western Africa, where dogs and their ticks live in close contact with humans.
Methods
A total of 216 canine and 22 feline blood samples and ticks feeding on them were collected in 2013 and 2014 from eight localities in Ivory Coast and four localities in Kenya. PCR was performed using specific primers targeting a portion of the 16S rRNA gene, followed by sequencing.
Results
The main results highlight the high prevalence of A. platys infection in dogs in both Eastern and Western Africa and report it for the first time in Eastern Africa and in Rhipicephalus camicasi.
Conclusions
The presence of A. platys DNA in R. camicasi feeding on negative hosts together with the relatively high prevalence of A. platys, correlated with the absence of the probable vector Rhipicephalus sanguineus (s.l.) in Kenyan Island suggest the involvement of other tick species in the transmission of A. platys.
Similar content being viewed by others


Background
Anaplasma platys, the agent of infectious canine cyclic thrombocytopenia (ICCT), is an obligate intracellular rickettsial organism of the family Anaplasmataceae [1, 2]. It is commonly infecting platelets where it forms basophilic morulae [1, 3]. Anaplasma platys infection was initially considered a subclinical condition with few or no clinical signs [1, 4, 5]. However, in studies performed in experimentally and naturally infected dogs in Greece [6], France [7] and Israel [8], clinical signs have been described, suggesting the involvement of more virulent A. platys strains [9]. Moreover, the zoonotic potential was suggested in the last few years in Venezuela, USA and Grenada or South Africa [10–12].
The first A. platys infection was described by Harvey et al. [1] in dogs from Florida, USA. Since then, the pathogen was recorded in several countries from the New World: Venezuela [13], Brazil [14], Chile [15], Argentina [16], Panama [17] and French Guiana [18]. In Europe it was reported in southern and western countries and in several countries from eastern and central Europe [19–25]. Anaplasma platys infection was reported in the majority of the countries from Asia and also in Australia [26–37].
In sub-Saharan Africa, A. platys was identified in Rhipicephalus sanguineus sensu lato (s.l.) ticks collected from dogs in the Democratic Republic of Congo [38], in engorged ticks collected from domestic and wild ruminants in South Africa [39] and in dogs in Ivory Coast, Gabon and Nigeria [40, 41]. In North Africa, A. platys infection was also reported in ticks and dogs in Tunisia [42], Morocco [43] and Algeria [44].
Considering the scarce data regarding the epidemiology of A. platys in dogs in the sub-Saharan Africa, the almost ubiquitous presence of R. sanguineus (s.l.) on dogs and the zoonotic potential, the aim of this study is to evaluate the prevalence of A. platys and to extend the knowledge on its distribution in dogs from East and West Africa (Kenya and Ivory Coast), and to correlate its occurrence with ecological factors.
Methods
Study sites, blood sampling, tick collection and identification
Blood samples and ticks from dogs and cats were collected in 12 localities in Ivory Coast (April 2013) and Kenya (January 2014). A total of 216 free-roaming dogs and 22 cats were sampled. Demographic data, identification of animals (name, owner, age and sex), clinical signs and the presence of ectoparasites were recorded for each animal (Table 1). Blood was collected from the cephalic vein and then transferred into ethanol filled tubes for storage. Ticks were preserved in absolute ethanol. Morphological identification was done individually for each tick to the species level by using morphological keys and descriptions [45, 46].
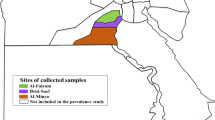
In Ivory Coast, the sampling sites (n = 8) were located in the southern part of the country (5.0–5.5 N/3.5–5.5 W°, Fig. 1, Additional file 1: Figure S1), while in Kenya the samples originated from four locations situated on Paté and Lamu Islands in Lamu Archipelago (2.0–3.35 S/40.8–41.15 E°, Fig. 2, Additional file 2: Figure S2). The collection sites from Ivory Coast belong to Eastern Guinean Forest ecoregion (Tropical and Subtropical Moist Broadleaf Forest), the dominant biome being semi-deciduous and moist evergreen forests [47]. The temperature ranges between 22 and 34 °C. The rainfall pattern can be divided into two wet and dry seasons, with a rainfall average between 1,400 and 2,500 mm per year [47].

Collection sites and A. platys-positive sites from Ivory Coast. ● collection sites; ▲ A. platys- positive sites

Collection sites and A. platys-positive sites from Kenya. ● collection sites;▲ A. platys-positive sites
Lamu and Paté Islands (Kenya) belong to the Eastern African Mangroves ecoregion. Annual rainfall averages range between 750 and 1,500 mm with two distinct rainy seasons, the temperatures are high, ranging between 23 and 33 °C [48].
DNA extraction
Genomic DNA was extracted from ethanol preserved blood clots and ticks using a modified phenol-chloroform method [49, 50]. Crushed ticks and 200 μl of clotted blood were dried at 56 °C for 30 min and then suspended in 1.5 ml lysis buffer (0.1 M NaCl, 0.05 M EDTA, 0.01 M Tris, 4.8 % SDS; pH 8). The digestion was performed in 20 μl proteinase K (Bioline, UK) at 56 °C for 90 min for blood and respectively overnight for ticks. After protein lysis, the mixture was extracted with an equal volume blend of phenol and chloroform, followed by one extraction with chloroform alone. Each extraction step included 1 min of shaking and 10 min centrifugation (13,000 × g). DNA was precipitated with 96 % ethanol for 15 min and then dried at room temperature in sterile medium for 12 h. The dried DNA pellet was re-suspended by adding 100 μl of PCR water. For each extraction procedure, negative controls consisting in reaction mixes without DNA (PCR water instead of blood) were used, in order to check for possible cross-contamination. Isolated genomic DNA from a representative number of samples was assessed quantitatively by Nanodrop ND-1000 spectrophotometer analyzer (NanoDrop Technologies, Inc., Wilmington, DE, USA).
Polymerase chain reaction (PCR) and agarose gel electrophoresis
PCR was initially performed using a group-specific set of primers that amplify a 345 bp fragment of the 16S rRNA gene. The primer sets EHR16SD (5′-GGTACCYACAGAAGAAGTCC-3′) and EHR16SR (5′-TAGCACTCATCGTTTACAGC-3′) amplify various species including Ehrlichia canis, E. chaffeensis, E. muris, Cowdria ruminantium, Anaplasma phagocytophilum, A. platys, A. marginale, A. centrale, Wolbachia pipientis, Neorickettsia sennetsu, N. risticii, and N. helminthoeca [51]. For all positive samples, a second PCR reaction was carried out using A. platys specific primers EPLAT5/EPLAT3 (forward primer: 5′-TTTGTCGTAGCTTGCTATGAT-3′ and a reverse primer: 5′-CTTCTGTGGGTACCGTC-3′), amplifying a 349-bp fragment of the 16S rRNA gene [52]. The amplification was performed as follows: 25 μl reaction mixture containing 4 μl aliquot of isolated DNA, 12.5 μl of 2× Green Master Mix (Rovalab GmBH) and 1 μl of each primer (0.01 mM). The PCR was carried out using the T100TM Thermal Cycler (Bio-Rad). The amplification profile of the first PCR consisted of 5 min of initial denaturation at 95 °C, followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 55 °C for 30 s and extension at 72 °C for 90 s, and a final extension at 72 °C for 5 min. The protocol used for the second PCR was: 5 min of initial denaturation at 95 °C, followed by 40 cycles of denaturation at 94 °C for 30 s, annealing at 58 °C for 30 s and extension at 72 °C for 45 s and a final extension at 72 °C for 5 min. In each PCR reaction set (48 samples), a positive and a negative control were included in order to assess the specificity of the reaction and the possible presence of contaminants. Positive controls consisted of DNA isolated from the blood of a dog naturally infected with A. platys in Israel. The reaction mix without DNA was used as a negative control. Amplicons were visualised by electrophoresis in a 1.5 % agarose gel (1 × TAE, pH 8.0) stained with SYBR® Safe DNA gel stain (Invitrogen) and their molecular weight was assessed by comparison to a molecular marker (O’GeneRuler™ 100 bp DNA Ladder, Thermo Fisher Scientific Inc., USA). All A. platys positive PCR samples were sequenced.
DNA sequencing
PCR products were purified from amplicons using the QIAquick PCR Purification Kit (QIAGEN). Sequencing analysis was performed (Macrogen Europe, Amsterdam) and the obtained sequences were compared with those available in GenBankTM by Basic Local Alignments Tool (BLAST) analysis.
Statistical analysis
Statistical analysis was performed using EpiInfoTM7 (CDC, USA) software. The total infection prevalence of A. platys (95 % CI), the infection prevalence based on age and sex of dogs, the prevalence of A. platys DNA presence in each ticks species, the infection prevalence in each location and the infection prevalence characterised by presence or absence of ticks on the animals and by ecoregions was assessed using the chi-square independence test. Maps showing the collection sites and the positive locations for A. platys were generated using QGIS 2.6 software.
Ethics statement
This study was approved by the USAMV CN Bioethics committee with the registration number 23/21–09–2015, following the EU 2010/63 and National directives Ord. 28/31–08–2011 and National Law 206/2004. Our projects developed in these countries followed the respective national laws, they were performed with the owners informed consent and were supervised by local Veterinary Authorities.
Results
A total of 238 blood samples from 216 dogs and 22 cats from Kenya and Ivory Coast were analysed for the presence of A. platys DNA using PCR. The overall prevalence in dogs was 12.5 % (27/216, 95 % CI: 8.4–17.66). All blood samples collected from cats were negative. In Kenya, the bacterium was identified in all four locations, while in Ivory Coast it was present in two out of eight locations. In the positive locations, the prevalence of infection varied between 10 and 30.4 %, with an average value of 21.26 % (27/127; 95 % CI: 14.5–29.4). The highest prevalence was recorded in Irobo village (Ivory Coast). When considering all collection sites, the differences in prevalence between localities were statistically significant (χ 2 = 25.11, df = 11, P < 0.01). However, no significant difference was observed when considering only the positive localities (χ 2 = 2.28, df = 5, P > 0.05). The prevalence of A. platys infection in each locality is shown in Table 1.
The overall prevalence by countries and respectively by ecoregions was significantly higher (χ 2 = 4.86, df = 1, P < 0.05) in Kenya compared to Ivory Coast (Table 1).
The prevalence of infection in young dogs (under 1 year) was 19.79 % (19/96, 95 % CI: 13.36–29.17) and the prevalence in adult dogs was 6.67 % (8/120, 95 % CI: 2.92–12.71); the young dogs were significantly more infected (χ 2 = 8.4, df = 1, P < 0.005). The prevalence of A. platys infection in females was 10.89 % (11/101, 95 % CI: 5.56–18.65) and in males 13.91 % (16/115, 95 % CI: 8.17–21.61), without significant differences between them.
Overall, 65.74 % (142/216, 95 % CI: 59–72.1) of the dogs were infested by ticks; of these, 16.9 % (24/142, 95 % CI: 11.14–24.10) were also positive for A. platys; only 4.05 % (3/74, 95 % CI: 0.84–11.39) of the dogs that were free of ticks were positive, the difference between these two categories being statistically significant (χ 2 = 7.3, df = 1, P < 0.05).
The prevalence of A. platys DNA presence in R. sanguineus (s.l.) was 4.38 % (16/365, 95 % CI: 2.61–7.17) in respectively 1.62 % (6/371, 95 % CI: 0.66–3.66) in R. camicasi. The other identified tick species shown in Table 1 were negative for A. platys DNA. Out of 16 A. platys-positive ticks, ten R. sanguineus (s.l.) derived from A. platys-positive dogs (6.49 %, 10/154, 95 % CI: 3.16–11.62) while six derived from negative dogs (2.84 %, 6/211, 95 % CI: 1.05–6.09) (Table 2). The positive R. camicasi were also collected from both positive and negative dogs, with a prevalence of A. platys DNA presence of 6.15 % (4/65, 95 % CI: 1.7–15.01) in ticks derived from positive dogs and 0.65 % (2/306, 95 % CI: 0.11–2.6) in ticks collected from negative dogs (Table 2).
The data regarding the structure of tick populations and the prevalence of each tick species are presented in Table 1. In Ivory Coast the presence of A. platys in dogs from one area can be correlated with the high prevalence (> 50 %) of R. sanguineus (s.l.) (Tabel 1, k = 1) and in Kenya, with the high prevalence (> 50 %) of R. camicasi (Table 1, k = 1).
From 49 A. platys-positive samples were obtained 37 sequences with a length varying between 320 and 340 bp. The BLAST analysis of all sequences showed 99 to 100 % similarity with various A. platys strains isolated from dogs originating from South America, Asia and Africa (accession numbers: KC989957, KF360842, JX112780, GQ395384,KC109446, KF576217, JX112781, AF478131 and KT357373). All sequences obtained from dogs and ticks were highly similar (99.7 %) amongst each other.
Discussion
This study reports the presence of A. platys in dogs from Ivory Coast and Kenya. This is the first report of this pathogen in dogs from East Africa and the first evidence of A. platys in R. camicasi ticks. Few reports are available regarding the presence of A. platys in Africa.
The overall prevalence of A. platys infection obtained in our study is higher than in other previous studies from Africa, but falls within the global average. The prevalence obtained in Kenya is similar to the highest prevalence rates, which were recorded in Brazil [53], Venezuela [54] and Japan [55]. The prevalence based on age groups (juveniles and adults) obtained in our study was higher in juvenile dogs (under 1 year-old). Similar results were obtained in the study of Brown et al. [56]. However, other studies showed no significant differences [41, 53]. Our results could be influenced by the tick burden, which is higher in young dogs [57, 58], including the ones sampled in this study (data not shown).
The different results regarding the prevalence in Ivory Coast and Kenya could be explained by the unequal sampling effort (140 versus 86), by the different collection time and by the presence and abundance of the tick vector. Besides molecular detection of A. platys in this tick species, the geographical distribution of A. platys infection seems to overlap with the distribution of the brown dog-tick R. sanguineus (s.l.), as described by Walker et al. [45]. Despite these facts, an experimental study failed to demonstrate the vectorial capacity of this tick species [59]. The employed microscopy method used for diagnosis showed a low sensitivity in experimental models [56]. However, R. sanguineus (s.l.) is still considered the most probable vector [38, 41].
The presence of R. sanguineus (s.l.) in Africa was described by Cumming [60] and Walker et al. [42]. In tropical and subtropical regions R. sanguineus s.l. is active throughout the year [61, 62] and apparently has no seasonality [62, 63]. However, a warmer temperature may contribute to increased tick abundance by a more rapid development resulting in lower mortality rates [64]. The abundance of the vector could influence the presence of A. platys because of the short bacteremia. In experimental models the average PCR positivity was 109 days after infection [65].
The prevalence of A. platys infection could be influenced also by the structure of the tick community, which was different in the collection sites, suggesting an influence of the environmental factors. The low prevalence or even absence of R. sanguineus (s.l.) in the sampled localities from the Kenyan islands and the presence of A. platys DNA in R. camicasi collected from negative hosts, suggest the possibility of this tick species acting as vector in this area. This hypothesis may be sustained by other studies that reported the presence of A. platys in other species of Rhipicephalus, such as unengorged R. turanicus in Israel and R. evertsi in South Africa [39, 66]. Moreover, Anaplasma strains closely related to A. platys were isolated from R. bursa in Turkey [67]. Further research on the topic is needed in order to confirm the vectorial role of other tick species.
The discrepancy between the high prevalence of A. platys infection in Kenyan dogs and the low prevalence in R. camicasi collected from negative hosts suggests the involvement of others factors such as the unequal number of the tested dogs and tick populations and the unknown previous hosts of R. camicasi during the immature stages [45].
Although in our study we did not find A. platys infections in the feline samples, there are two reports of infection in cats [3, 68] from Brazil and Thailand.
The zoonotic potential of A. platys is known, but its public health impact is still under debate. In the last few years, three human cases of multiple infections including A. platys, with clinical signs were reported [10, 11]. Also two clinical cases with single A. platys infection were recently confirmed in two women from Venezuela [12].
Conclusions
The results of this study highlight for the first time the presence of A. platys infection in dogs originating from East Africa. Moreover the detection of A. platys DNA in R. sanguineus (s.l.) ticks from Ivory Coast sustains its probable vectorial role. However presence of A. platys DNA in R. camicasi correlated with absence of the probable vector R. sanguineus (s.l.) in Kenyan Island suggest the involvement of other tick species in the transmission of A. platys.
References
Dumler JS, Barbet AF, Bekker CPJ, Dasch GA, Palmer GH, Ray SC, et al. Reorganization of genera in the families Rickettsiaceaeand Anaplasmataceaein the order Rickettsiales: unification of some species of Ehrlichia with Anaplasma, Cowdria with Ehrlichia, and Ehrlichia with Neorickettsia, descriptions of six new species combinations and designation of Ehrlichia equiand ‘HGE agent’ as subjective synonyms of Ehrlichia phagocytophila. Int J SystEvol Microbiol. 2001;51:2145–65.
Harvey JW, Simpson CF, Gaskin JM. Cyclic thrombocytopenia induced by a Rickettsia-like agent in dogs. J Infect Dis. 1978;137:182–8.
Salakij C, Lertwatcharasarakul P, Salakij J, Nunklang K, Rattanakunuprakarn J. Molecular characterization of Anaplasma platys in a domestic cat from Thailand. Comp Clin Path. 2012;21(3):345–8.
Hibler SC, Greene CE. Rickettsial infections in Dogs Part II: Ehrlichiosis and Infectious Cyclic Thrombocytopenia. Comp Cont EducPract Vet. 1986;8:106–14.
Woody BJ, Hoskins JD. Ehrlichial diseases of dogs. Vet Clin North Am Small AnimPract. 1991;21(1):75–98.
Kontos VI, Papadopoulos O, French TW. Natural and experimental canine infections with a Greek strain of Ehrlichia platys. Vet Clin Pathol. 1991;20:101–5.
Beaufils JP, Inokuma H, Martin-Granel J, Jumelle P, Barbault-Jumell M, Brouqui P. Anaplasma platys (Ehrlichia platys) infection in a dog in France: description of the case and characterization of the agent. Rev Med Vet. 2002;153:85–90.
Harrus S, Aroch I, Lavy E, Bark H. Clinical manifestations of infectious canine cyclic thrombocytopenia. Veterinary Record. 1997;141:247–50.
Agguire E, Tesouro MA, Ruiz L, Amusategui I, Sainz A. Genetic characterization of Anaplasma (Ehrlichia) platys in dogs in Spain. J Vet Med. 2006;53(4):197–200.
Maggi RG, Mascarelli PE, Havenga LN, Naidoo V, Breitschwerdt EB. Co-infection with Anaplasma platys, Bartonellahenselae and Candidatus Mycoplasma haematoparvum in a veterinarian. Parasit Vectors. 2013;6(1):103.
Breitschwerdt EB, Hegarty BC, Qurollo BA, Saito TB, et al. Intravascular persistence of Anaplasma platys, Ehrlichia chaffeensis, and Ehrlichia ewingii DNA in the blood of a dog and two family members. Parasit Vectors. 2014;7:298.
Arraga-Alvarado CM, Qurollo BA, Parra OC, Berrueta MA, Hegarty BC, Breitschwerdt EB. Molecular evidence of Anaplasma platys infection in two women from Venezuela. AmJ Trop Med Hyg. 2014;91(6):1161–5.
Arraga-Alvarado C, Parra O, Palmar M, Chango RE, Alvarado MC. Ehrlichia platys: preparación del antigeno y uso de la técnica de inmunofluorescenciaindirecta (IFI) en caninos y humanos. Rev Cient. 1997;FCV-LUZ 7(2):99–109.
Ferreira RF, de Mello Figueiredo Cerqueira A, Pereira AM, Guimaraes CM, de Sa AG, da Silva AF, et al. Anaplasma platys diagnosis in dogs: comparison between morphological and molecular tests. Int J Appl Res Vet Med. 2007;5(3):113.
Abarca K, López J, Perret C, Guerrero J, Godoy P, Veloz A, et al. Anaplasma platys in dogs. Chile Emerg Infect Dis. 2007;13:1392–5.
Eiras DF, Craviotto MB, Vezzani D, Eyal O, Baneth G. First description of natural Ehrlichia canis and Anaplasma platys infections in dogs from Argentina Comp. Immunol Microbiol Infect Dis. 2013;36(2):169–73.
Santamaria A, Calzada JE, Saldana A, Yabsley MJ, Gottdenker NL. Molecular diagnosis and species identification of Ehrlichia and Anaplasma infections in dogs from Panama, Central America. Vector Borne Zoonotic Dis. 2014;14(5):368–70.
Dahmani M, Marié JL, Mediannikov O, Raoult D, Davoust B. First identification of Anaplasma platys in the blood of dogs from French Guiana. Vector Borne Zoonotic Dis. 2015;15(2):170–2.
Beaufils JP. Rickettsioses du chien et du chat. In: EncyclopédieVétérinaire. french: Elsevier, Paris, médecine générale; 1998. p. 1100–9.
Glaser B, Gothe R. Imported arthropod-borne parasites and parasitic arthropods in dogs. Species spectrum and epidemiologic analysis of the cases diagnosed in 1995/96. TierarztlPraxAusg K Kleintiere/Heimtiere. 1998;26(1):40–6.
Sainz Á, Amusategui I, Tesouro MA. Ehrlichia platys infection and disease in dogs in Spain. J Vet Diagn Invest. 1999;11(4):382–4.
Sparagano OA, de Vos AP, Paoletti B, Camma C, de Santis P, Otranto D, et al. Molecular detection of Anaplasma platys in dogs using polymerase chain reaction and reverse line blot hybridization. J Vet Diagn Invest. 2003;15:527–34.
Cardoso L, Tuna J, Vieira L, Yisaschar-Mekuzas Y, Baneth G. Molecular detection of Anaplasma platys and Ehrlichia canis in dogs from the North ofPortugal. Vet J. 2010;183:232–3.
Dyachenko V, Pantchev N, Balzer HJ, Meyersen A, Straubinger RK. First case of Anaplasma platys infection in a dog from Croatia. Parasit Vectors. 2012;5(1):1–7.
Andersson M, Turcitu MA, Stefanache M, Tamba P, Barbuceanu F, Chitimia L. First evidence of Anaplasma platys and Hepatozoon canis co-infection in a dog from Romania–a case report. Ticks Tick Borne Dis. 2013;4(4):317–9.
Harrus S, Waner T, Eldor A, Zwang E, Bark H. Platelet dysfunction associated with experimental acute canine ehrlichiosis. Vet Rec. 1996;139(12):290–3.
Chang ACH, Chang WL, Lin CT, Pan MJ, Lee SC. Canine infectious cyclic thrombocytopenia found in Taiwan. J Vet Med Sci. 1996;58:473–6.
Hua P, Yuhai M, Slide T, Yang S, Bohai W, Xiangrui C. Canine ehrlichiosis caused simultaneously by Ehrlichia canisand Ehrlichia platys. Microbiol Immunol. 2000;44:737–9.
Inokuma H, Raoult D, Brouqui P. Detection of Ehrlichia platys DNA in brown dog ticks (Rhipicephalus sanguineus) in Okinawa Island Japan. J Clin Microbiol. 2000;38:4219–21.
Brown GK, Martin AR, Roberts TK, Aitken RJ. Detection of Ehrlichia platys in dogs in Australia. Aust Vet J. 2001;79(8):554–8.
Suksawat J, Pitulle C, Arraga-Alvarado C, Madrigal K, Hancock SI, Breitschwerdt EB. Coinfection with three Ehrlichia species in dogs from Thailand and Venezuela with emphasis on consideration of 16S ribosomal DNA secondary structure. J Clin Microbiol. 2001;39(1):90–3.
Karagenc T, Hosgor M, Bilgic HB, Pasa S, Kırlı G, Eren H. Detection of prevalence of E. canis, A. phagocytophila and A. platys in dogs around the Aegean coast of Turkey by using a nested PCR. Izmir: XIV National Parasitology Congress; 2005. p. 18–25.
Kim CM, Yi YH, Yu DH, Lee MJ, Cho MR, Desai AR, et al. Tick-borne rickettsial pathogens in ticks and small mammals in Korea. Appl Environ Microbiol. 2006;72:5766–76.
Kumar A, Varshney JP. Clinico-haematological and biochemical investigations in dogs concurrently infected with Anaplasma platys and other blood parasites. J Vet Parasitol. 2007;21:43–5.
Ybañez AP, Perez ZO, Gabotero SR, Yandug RT, Kotaro M, Inokuma H. First molecular detection of Ehrlichia canis and Anaplasma platys in ticks from Dogs in Cebu Philippines. Ticks Tick-Borne Dis. 2012;3(5):288–93.
Mokhtar AS, Lim SF, Tay ST. Research Note Molecular detection of Anaplasma platys and Babesia gibsoni in dogs in Malaysia. Trop Biomed. 2013;30(2):345–8.
Javkhlan G, Enkhtaivan B, Baigal B, Myagmarsuren P, Battur B, Battsetseg B. Natural Anaplasma phagocytophilum infection in ticks from a forest area of Selenge province, Mongolia. Western Pac Surveill Response J WPSAR. 2014;5(1):21.
Sanogo YO, Davoust B, Inokuma H, Camicas JL, Parola P, Brouqui P. First evidence of Anaplasma platys in Rhipicephalus sanguineus (Acari: Ixodida) collected from dogs in Africa. Onderstepoort J Vet Res. 2003;70:205–12.
Berggoetz M, Schmid M, Ston D, Wyss V, Chevillon C, Pretorius AM, et al. Protozoan and bacterial pathogens in tick salivary glands in wild and domestic animal environments in South Africa. Ticks Tick Borne Dis. 2014;5(2):176–85.
Marié JL, Shaw SE, Langton DA, Bourry O, Gomez J, Davoust B. Sub‐clinical infection of dogs from the Ivory Coast and Gabon with Ehrlichia, Anaplasma, Mycoplasma and Rickettsia species. Clin Microbiol Infect. 2009;15(s2):284–5.
Kamani J, Baneth G, Mumcuoglu KY, Waziri NE, Eyal O, Guthmann Y, et al. Molecular detection and characterization of tick-borne pathogens in dogs and ticks from Nigeria. PLoS Negl Trop Dis. 2013;7(3):e2108. doi:10.1371/journal.pntd.0002108.
Sarih MH, M’Ghirbi Y, Bouattour A, Gern L, Baranton G, Postic D. Detection and identification of Ehrlichia spp. in ticks collected in Tunisia and Morocco. J Clin Microbiol. 2005;43(3):1127–32.
Seng P, Sarih M, Socolovschi C, Boudebouch N, Hassar M, Parola P, et al. Detection of Anaplasmataceae in ticks collected in Morocco. Clin Microbiol Infect. 2009;15:86–7.
Dahmani M, Loudahi A, Mediannikov O, Fenollar F, Raoult D, Davoust B. Molecular detection of Anaplasma platys and Ehrlichia canis in dogs from Kabylie, Algeria. Ticks Tick Borne Dis. 2015;6(2):198–203.
Walker JB, Keirans JE, Horak IG. The GenusRhipicephalus (Acari, Ixodidae): A Guide to the Brown Ticks of the World. New York: Cambridge University Press; 2000.
Walker AR, Bouattour A, Camicas JL, Estrada-Peña A, Horak IG, Latif AA, et al. Ticks of Domestic Animals in Africa: a Guide to Identification of Species. Ed. Edinburgh: Bioscience Reports; 2003; 221–223
Fund W. Eastern Guinean forests. 2014. http://www.eoearth.org/view/article/151905. Accessed 15.09.2015.
Fund W. East African mangroves. 2014. http://www.eoearth.org/view/article/151864.Accessed 15.09.2015.
Maslov DA, Lukeš J, Jirků M, Simpson A. Phylogeny of trypanosomes as inferred from the small and large subunit rRNAs: implications for the evolution of parasitism in the trypanosomatid protozoa. Mol Biochem Parasitol. 1996;75:197–205.
Albrechtová K, Sedlák K, Petrželková KJ, Hlaváč J, Mihalca AD, Lesingirian A, et al. Occurrence of filarial in domestic dogs of Samburu pastoralists in Northern Kenya and its associations with canine distemper. Vet Parasitol. 2011;182:230–8.
Parola P, Roux V, Camicas JL, Baradji I, Brouqui P, Raoult D. Detection of Ehrlichiae in African ticks by PCR. Trans R Soc Trop Med Hyg. 2000;94:707–8.
Murphy GL, Ewing SA, Whitworth LC, Fox JC, Kocan AA. A molecular and serologic survey of Ehrlichia canis, E. chaffeensis, and E. ewingii in dogs and ticks from Oklahoma. Vet Parasitol. 1998;79:325–39.
Silva GCFD, Benitez ADN, Girotto A, Taroda A, Vidotto MC, Garcia JL. Occurrence of Ehrlichia canis and Anaplasma platys in household dogs from northern Parana. Rev Bras Parasitol Vet. 2012;21(4):379–85.
Huang H, Unver A, Perez MJ, Orellana NG, Rikihisa Y. Prevalence and molecular analysis of Anaplasma platys in dogs in Lara, Venezuela. Braz J Microbiol. 2005;36(3):211–6.
Motoi Y, Satoh H, Inokuma H, Kiyuuna T, Muramatsu Y, Ueno H, et al. First detection of Ehrlichia platys in dogs and ticks in Okinawa, Japan. Microbiol Immunol. 2001;45(1):89–91.
Brown GK, Canfield PJ, Dunstan RH, Roberts TK, Martin AR, Brown CS, et al. Detection of Anaplasma platys and Babesia canisvogeli and their impact on platelet numbers in free‐roaming dogs associated with remote Aboriginal communities in Australia. Aust Vet J. 2006;84(9):321–5.
Tinoco-Gracia L, Quiroz-Romero H, Quintero-Martínez MT, Rentería-Evangelista TB, González-Medina Y, Barreras-Serrano A, et al. Prevalence of Rhipicephalus sanguineus ticks on dogs in a region on the Mexico-USA border. Vet Rec. 2009;164:59–61.
Dantas-Torres F, Melo MF, Figueredo LA, Brandão-Filho SP. Ectoparasite infestation on rural dogs in the municipality of São Vicente Férrer, Pernambuco, Northeastern Brazil. Rev Bras Parasitol Vet. 2009;18:75–7.
Simpson RM, Gaunt SD, Hair JA, Kocan KM, Henk WG, Casey HW. Evaluation of Rhipicephalus sanguineus as a potential biologic vector of Ehrlichia platys. Am J Vet Res. 1991;52:1537–41.
Cumming GS. Host distributions do not limit the species range of most African ticks (Acari: Ixodida). Bull Entomol Res. 1999;89:303–27.
Jacobs PAH, Fourie LJ, Kok DJ, Horak IG. Diversity, seasonality and sites of attachment of adult Ixodid ticks on dogs in the central region of the Free State Province, South Africa. Onderstepoort J Vet Res. 2001;68:281–90.
Louly CCB, Fonseca IN, Oliveira VF, Borges LMF. Ocorrência de Rhipicephalus sanguineus emtrabalhadores de clínicasveterinárias e canis, no município de Goiânia, GO. Cienc Anim Bras. 2006;7:103–6.
D’Amico G, Dumitrache MO, Široký P, Albrechtová K, Sloboda M, Domșa C, et al. Altitudinal and seasonal differences of tick communitiesin dogs from pastoralist tribes of Northern Kenya. Vet Parasitol.2015; doi:10.1016/j.vetpar.2015.08.025
Gray J, Dantas-Torres F, Estrada-Peña A, Levin M. Systematics and ecology of the brown dog tick, Rhipicephalus sanguineus. Ticks Tick Borne Dis. 2013;4(3):171–80.
Gaunt S, Beall M, Stillman B, Lorentzen L, Diniz PPVP, Chandrashekar R, et al. Experimental infection and co-infection of dogs with Anaplasma platys and Ehrlichia canis: hematologic, serologic and molecular findings. Parasit Vectors. 2010;3(1):33.
Harrus S, Perlman‐Avrahami A, Mumcuoglu KY, Morick D, Eyal O, Baneth G. Molecular detection of Ehrlichia canis, Anaplasma bovis, Anaplasma platys, Candidatus Midichloria mitochondrii and Babesia canisvogeli in ticks from Israel. Clin Microbiol Infect. 2011;17(3):459–63.
Aktas M, Altay K, Dumanli N, Kalkan A. Molecular detection and identification of Ehrlichia and Anaplasma species in ixodid ticks. Parasitol Res. 2009;104(5):1243–8.
Lima MLF, Soares PT, Ramos CAN, Araújo FR, Ramos RAN, Souza IIF, et al. Molecular detection of Anaplasma platys in a naturally-infected cat in Brazil. Braz J Microbiol. 2010;41(2):381–5.
Acknowledgements
The research was supported from UEFISCDI grant TE298/2015. We are also grateful for the help of our collaborators in Ivory Coast during the field work in 2013: Dr. Stéphane Kouassi Kan and Dr. Kouassi Eugéne Koffi (Institut Pasteur de Côte d’Ivoire), Dr. Kouamé Bertin Akpatou and Ekoun Michael Konan (Université Félix Houphouet-Boigny, UFR Biosciences). Special thanks to the institutions in Ivory Coast who supported the research with the relevant permits and logistics: SODEFOR (Société de développement des Forêts), IPCI (Institut Pasteur de Côte d’Ivoire) and OIPR (Office Ivoirien des Parcs et Réserves). The field work in Kenya was done with the great support from Klára Kunčická, Martin Modrý, Ali Abdulrahman and other local field assistants. Special thanks to Dr. Alicia Rojas and Prof. Gad Baneth for providing the positive control. This paper was published under the frame of EurNegVec COST Action TD1303.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
Financial competing interests
The authors declare that they have no competing interests.
Non-financial competing interests
There are no non-financial competing interests.
Authors’ contributions
IAM wrote the manuscript and performed molecular diagnostic. GD performed ticks identification, AMI performed the DNA extraction. AAD performed molecular diagnosis. GD, CMG, PKY, ADS, ADM, AMI, MQ, PWNK and DM performed field work. DM and ADM coordinated the research teams. All authors read and approved the final manuscript.
Additional files
Additional file 1: Figure S1.
klm. Geographic coordinates of collection sites from Ivory Coast. (KML 4 kb)
Additional file 2: Figure S2.
klm.Geographic coordinates of collection sites from Kenya. (KML 1 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Matei, I.A., D’Amico, G., Yao, P.K. et al. Molecular detection of Anaplasma platys infection in free-roaming dogs and ticks from Kenya and Ivory Coast. Parasites Vectors 9, 157 (2016). https://doi.org/10.1186/s13071-016-1443-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-016-1443-3