
Abstract
Background
Translocation of native species and introduction of non-native species are potentially harmful to the existing biota by introducing e.g. diseases, parasites and organisms that may negatively affect the native species. The enemy release hypothesis states that parasite species will be lost from host populations when the host is introduced into new environments.
Methods
We tested the enemy release hypothesis by comparing 14 native and 29 introduced minnow (Phoxinus phoxinus) populations in Norway with regard to the ectoparasitic Gyrodactylus species community and load (on caudal fin). Here, we used a nominal logistic regression on presence/absence of Gyrodactylus spp. and a generalized linear model on the summed number of Gyrodactylus spp. on infected populations, with individual minnow heterozygosity (based on 11 microsatellites) as a covariate. In addition, a sample-based rarefaction analysis was used to test if the Gyrodactylus-species specific load differed between native and introduced minnow populations. An analysis of molecular variance was performed to test for hierarchical population structure between the two groups and to test for signals of population bottlenecks the two-phase model in the Wilcoxon signed-rank test was used. To test for demographic population expansion events in the introduced minnow population, we used the kg-test under a stepwise mutation model.
Results
The native and introduced minnow populations had similar species compositions of Gyrodactylus, lending no support to the enemy release hypothesis. The two minnow groups did not differ in the likelihood of being infected with Gyrodactylus spp. Considering only infected minnow populations it was evident that native populations had a significantly higher mean abundance of Gyrodactylus spp. than introduced populations. The results showed that homozygotic minnows had a higher Gyrodactylus spp. infection than more heterozygotic hosts. Using only infected individuals, the two minnow groups did not differ in their mean number of Gyrodactylus spp. However, a similar negative association between heterozygosity and abundance was observed in the native and introduced group. There was no evidence for demographic bottlenecks in the minnow populations, implying that introduced populations retained a high degree of genetic variation, indicating that the number of introduced minnows may have been large or that introductions have been happening repeatedly. This could partly explain the similar species composition of Gyrodactylus in the native and introduced minnow populations.
Conclusions
In this study it was observed that native and introduced minnow populations did not differ in their species community of Gyrodactylus spp., lending no support to the enemy release hypothesis. A negative association between individual minnow host heterozygosity and the number of Gyrodactylus spp. was detected. Our results suggest that the enemy release hypothesis does not necessarily limit fish parasite dispersal, further emphasizing the importance of invasive fish species dispersal control.
Similar content being viewed by others

Background
Anthropogenic translocation of species between ecosystems occurs worldwide at an increasing rate [1]. Some species that are introduced into new environments become invasive, imposing major negative ecological effects on the native biota [2] with concomitant economic costs for the society [3–6]. Introduced species may also act as vectors for new parasites and diseases that may infect native hosts [7]. Transmission of non-native parasite species may lead to large population-dynamic effects [8]. Noteworthy examples are the introduction of the monogenean Gyrodactylus salaris Malmberg, 1957 [9] into Norwegian rivers with subsequent dramatic decline in the native Atlantic salmon (Salmo salar L.) populations [10] and the crayfish plague (Aphanomyces astaci Schikora, 1906), a parasite that has been introduced to Norwegian watercourses and causes mass-mortalities in European crayfish (Astacus astacus L.) populations [11]. In these cases, strong and visible effects are evident, but often the effects of introduced parasites are difficult to observe [12].
Freshwater fish are commonly transported outside their native distribution area; this potentially leads to a loss of native fish species and that the fish communities become more similar (sometimes called species homogenization)[13, 14]. The minnow (Phoxinus phoxinus L.) is distributed from Urals in the east to Europe in the west. In Norway, the minnow's natural distribution is limited to the northern and southeastern parts [15]. During the last decades, new minnow populations have been established due to human activities such as fishing with live bait, stocking (intentional and non-intentional), and reorganization of waterways [16]. Genetic studies suggest that both short- and long-distance translocations of minnows have occurred between Norwegian watercourses [17, 18]. Several species of ectoparasites of the genera Gyrodactylus have been reported from minnow in Europe [9, 19, 20]. The diversity of Gyrodactylus spp. on Norwegian minnow is not well known, but earlier studies indicate that up to five Gyrodactylus species can be found [21]. The Gyrodactylus fauna in Norway is depauperate compared to the rest of Europe [22]. Gyrodactylus spp. often exhibit a high degree of host-specificity and a direct life cycle where transmission typically takes place after direct contact with a new host [23]. These traits make Gyrodactylus spp. particularly tractable for parasite studies, as it is not necessary to take infracommunities from intermediate hosts into consideration [24]. The effects of Gyrodactylus spp. on minnow hosts are not known, but based on other Gyrodactylus - host systems it is reasonable to assume they impose negative fitness impacts [25].
The “enemy release hypothesis” (ERH) states that introduced species lose some of their natural enemies such as pathogens and parasites in the new environment [26, 27]. This will provide a fitness advantage as less energy is used to respond to the parasites and more can be allocated to growth and reproduction. Comparing native and introduced plant species, Mitchell and Power [28] found that introduced plants harboured less fungi and virus species than plants in their native habitats. Further, Torchin et al. [29] compared 26 host taxa (molluscs, crustaceans, fishes, birds, mammals, amphibians and reptiles) and showed that introduced species had half the number of species of parasites compared to the native species. A comparison of 176 different studies addressing the enemy release hypothesis found almost as many studies in support (36 %) as questioning the hypothesis (43 %) [30]. There is also some support for the ERH-hypothesis in freshwater fish [31–34]. However, few studies have addressed this topic at the population level. Halvorsen [35] hypothesized that local movement of fish between neighbouring water bodies would similarly disseminate their parasites, assuming that ecological conditions were similar. Under this hypothesis, the prediction would be that transport of individuals between closely located water bodies would lead to a more similar parasite fauna in native and introduced hosts than if hosts are introduced to more geographically distant locations. Alternatively, the number of parasites might increase in introduced populations compared to native populations by acquiring new parasites in the new environments from resident hosts [36, 37].
There are several factors that can affect the abundance of and resistance to parasites in host species, both in native and introduced populations. One such factor is heterozygosity, measured by neutral genetic markers, which is hypothesized to be associated with fitness (review by Chapman et al. [38]), and fitness-related traits such as resistance to parasite infections [39]. The assumption is that a genetically diverse host has a more robust immune system to handle parasite infections as it holds a larger diversity of anti-parasite specific genes [40, 41]. Several studies on fish species have documented that selection acts on MHC genes related to e.g. monogenean infection (reviewed by Alvarez-Pellitero, [42]). Also, demographic bottlenecks during introduction to the new environments may result in reduced genetic diversity in host and/or parasite population, putatively affecting persistence and fitness in the new environment [43].
The main aim of this study was to test the enemy release hypothesis using a dataset on native and introduced minnow populations in Norway. First we tested if the diversity of Gyrodactylus species differed between native and introduced minnow populations. Then we tested for variation in prevalence (i.e. presence and absence) and intensity of Gyrodactylus (species pooled) between native and introduced minnow. By estimating multilocus heterozygosity of the minnow hosts using a set of eleven neutral microsatellites we tested for association between individual heterozygosity and intensity. We also tested if the transplanted minnow populations had gone through demographic bottlenecks and subsequent population expansions, which potentially could explain some of our results.
Methods
Study area and sampling
The sampling sites were selected to cover most of the distribution of native [15] and introduced [17, 44] minnow populations in Norway (Fig. 1, Table 1). The minnow populations, both native and introduced, are localized geographically far apart, usually in different watersheds. It is thus highly unlikely that there is or has been natural dispersal between the different minnow populations. The native minnow populations were all found below the upper marine limit, this limit is regarded as limiting the dispersal of minnow. The introduced minnow populations were mainly located in mountainous regions in southern Norway, and all above the upper marine limit [16]. Potentially, non-native minnow could have been introduced into native populations, leading to native and non-native minnow living in sympatry. However, a detailed study of the genetic population structure of minnow in many of these lakes indicated that this was not the case [17]. The Norwegian freshwater fish fauna is depauperate and in most populations only one or two species are present. Details on the composition of fish species at the different sampling locations is presented in Additional file 1. Brown trout (Salmo trutta) was the only species present at all sampling sites.

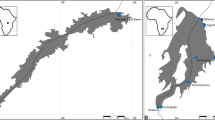
Sampling locations in Norway. The 43 minnow sampling locations in Norway (see Table 1). The 11 native minnow populations were collected from the grey part of the map
The water temperature regimes may differ between native minnow populations located at lower altitudes below the upper marine limit compared with introduced minnow usually found in high mountain lakes. Based on recent genetic studies, the newly founded minnow populations seem to have a diverse origin, some originating from fish transported at local scales and some transported at regional (European) scales [17, 18].
A total of 1278 minnow were randomly sampled from 43 populations (14 native; 29 introduced) during the years of 1997–2003, mainly during August and September (Fig. 1, Table 1). The introduced populations were identified based on their elevation above sea level (i.e. being situated outside the main natural distribution area of the minnow in Norway) as well as not having prior records of minnow occurrence [15, 44, 45]. However, the original sources of the introduced minnow populations are unknown, precluding a direct comparison between introduced and source populations.
Minnows were captured in small rivers/streams and in lakes close to the shoreline, using backpack electric fishing equipment. After capture, all minnows were immediately euthanized and stored on 96 % ethanol for later analysis. However, in some cases only the caudal fin was preserved in ethanol for later analysis (see description below). There is no information available on the density of minnow in the different lakes.
Gyrodactylus spp. identification
In the laboratory, the caudal fin was excised from each fish that had been preserved as a total individual. These excised caudal fins and the caudal fins that were excised and conserved in the field were then investigated in detail. The caudal fin was chosen in order to utilize the total material of minnow samples. Thus we here assume that parasite counts on the caudal fin are representative for the total Gyrodactylus spp. community. To assess this approach previously unpublished data (Pettersen, R.A.) were used to compare the number of G. aphyae, G. magnificus, and G. macronychus on the caudal fin with the number of specimen on the total body (using protocol developed by Buchmann, [46]). The results showed that the correlation was strong (Spearman ρ = 0.871, p = 0.0001. Nfish = 73).
Each fin was inspected using a stereomicroscope (40X) to count the number of Gyrodactylus spp. individuals. All Gyrodactylus spp. individuals were carefully removed for preparation for species identification using light microscopy. However, three individuals were lost during the preparation procedures.
Each Gyrodactylus individual was identified to species based on the haptoral hard parts [47]. The haptoral hard parts were digested with Proteinase K (1 % in buffer) until all the tissue was removed, and were mounted on a microscope slide in formaldehyde-glycerine (15:85) fixative. A Leica DM 4000 B microscope with a Heine phase contrast condenser, and a 100X/1.25 oil immersion objective (no. 506159) was used. This equipment was linked to a Leica DFC 320 digital camera and archiving system, and the Leica software LAS© to take pictures. These pictures were used to identify the different Gyrodactylus species. To establish the identity of the different Gyrodactylus species we used information from the first description and follow-up descriptive literature [9, 48]. We also used an additional visual identification as recommended by Shinn et al. [49] comparing our samples with pictures from the database GyroDb [22]. Five species of Gyrodactylus have previously been reported on minnow from Norway: G. laevis, Malmberg 1957, G. magnificus, Malmberg 1957, G. phoxini, Malmberg 1957, G. macronychus, Malmberg 1957, G. aphyae, Malmberg 1957 [50]. A further nine Gyrodactylus species are reported on minnow elsewhere in Eurasia [22]. One of the authors (Pettersen, R. A.) first identified all the Gyrodactylus species and counted all individuals, while another (Mo, T. A.) checked and confirmed all the species identifications.
Genetic diversity in the minnow hosts
Neutral multilocus genetic diversity for the minnow hosts was assessed by genotyping 11 microsatellites for a total of 1278 minnows. The 11 microsatellites were selected from a set of 36 microsatellite markers developed for cyprinids (the markers codes are: Z7634, Ca1, Ca3, Ca5, Ca6, Ca12, MFW1, MFW17, GF11, Z15751, Z9692) [17, 51]. Here, individual and population level heterozygosity were estimated using GenAlEx 6.5 [52], while gene diversity and allelic richness was estimated for populations using Fstat 2.9.3.2. [53]. Population estimates of these three measures of genetic diversity are given in Table 1. We used estimated hetereozygosity as a covariate in the statistical analyses, as Chapman et al., [38] suggests that this is a robust measure of genetic diversity. It is also assumed that heterozygosity is positively associated with parasite resistance (e.g. Blanchet et al., [39]).
To test for hierarchical population structure between native and introduced minnows, as well as for a geographical pattern of population subdivision, analysis of molecular variance (MANOVA, [54]) in the program GenAlex 6.5 was used [55]. Genetic variance (based on ϕPT) was partitioned among minnow individuals within populations, among populations, and among the population types (native and introduced minnows) using 9999 permutations.
If heterozygosity is important for individual parasite resistance in minnow hosts, demographic population bottlenecks associated with transfer of minnow to new locations may be important. To test for heterozygosity excess, being a signal of a population bottleneck, the Bottleneck 1.2.0.2 software was used [56]. Here, results were evaluated based on the two-phase model (TPM) in the Wilcoxon sign-rank test (1000 iterations), and the "mode - shift" indicator, which discriminates bottlenecked populations from demographically stable populations [57].
After introduction of minnows to new locations a demographic population expansion may occur, which can potentially be linked to persistence against parasites. To test for a demographic population expansion event in the introduced minnow population, we used the kg-test under a stepwise mutation model (SMM) [58]. Here, the k-test compares intra-locus allelic distributions between expanding and stable populations, while the interlocus g-test compares variance in number of repeats between expanding and stable population [58, 59]. A significant number of negative k-values indicate a signature of a demographic population expansion. The g-test significance level was compared to the recommended cut-off value in Table 1 (p. 455) in Reich et al., [58].
Statistical analyses
In a number of populations we did not find any individual minnows infected with Gyrodactylus spp. To test if the probability of being infected with Gyrodactylus spp. differed between native and introduced minnow populations, we used a nominal logistic regression (binomial distribution, logit link, the distribution of Gyrodactylus spp. is given in Online Resource 1). Presence or absence of Gyrodactylus spp. was used as the response variable, population group (native or introduced) as factor, and average heterozygosity as a covariate. Here, all Gyrodactylus species were pooled (43 minnow populations; 1278 fish individuals).
Further, Gyrodactylus species accumulation curves were calculated for each of the native and introduced minnow groups using EstimateS 8.2.0 [60]. This is a sample-based rarefaction curve that gives species accumulation as a function of occurrence, presenting associated 95 % confidence intervals. These curves allow species richness comparisons among test groups, while accounting for differences between locations in the number of individual minnows sampled [61, 62]. Here, only 10 native and 11 introduced populations out of the original 44 populations (Table 1) were used since the sample size of Gyrodactylus specimens in each population must be larger than the total number of Gyrodactylus species observed in the total dataset [60]. Thus, in all the 21 selected minnow populations more than four Gyrodactylus specimens was observed.
The mean number of Gyrodactylus specimens per host was estimated for native and introduced minnow populations, using the minnow populations where Gyrodactylus spp. were observed (29 populations, 880 individuals). We used a generalized linear model with the number of Gyrodactylus specimens as a response variable (using a Poisson error distribution and a log-link, using a maximum likelihood estimation method), and native or introduced minnow populations, population identity nested under the two groups, heterozygosity, and interaction between heterozygosity and native or introduced population groups as factors (interaction was not significant and thus removed from further analyses).
The same statistical test was applied but only using minnow hosts that were infected comprising 21 populations and 253 minnow hosts. Here, again the interaction between heterozygosity and the native or introduced population groups as factors was not significant and subsequently removed from the analyses.
All the statistical analyses, except the rarefaction analyses, were implemented in JMP 9.0 (SAS, 2012) [63].
Results
Gyrodactylus species occurrence
A total of 515 Gyrodactylus specimens were examined and identified. Four species were found: G. magnificus, G. phoxini, G. macronychus, and G. aphyae (Table 2). G. aphyae was the most common species being present in all the 21 infected minnow populations. G. magnificus was found in 12 populations, G. macronychus in 10 populations. G. phoxini was only found in 3 minnow populations. Most commonly, only one species of Gyrodactylus was found on each minnow, rarely two species of Gyrodactylus were found on the same tail fin. The combination of G. aphyae and G. magnificus was the most common, being detected on 11 minnows (from 10 populations). The combination of G. aphyae and G. phoxini was detected on 3 minnows (from 3 populations), and one minnow had the combination of G. magnificus, and G. macronychus.
Genetic variation and demographic tests in minnow populations
A total of 1278 minnows were genotyped for 11 microsatellites, and the observed individual level of heterozygosity ranged between 0.091 and 1.0, with the overall mean 0.534 for all fish. The population level heterozygosity (N = 43 populations) ranged from 0.381 (Møsvatn) to 0.632 (Birisjøen). Gene diversity ranged from 0.421 to 0.675, and allelic richness ranged from 1.422 to 1.673 (Table 1). Based on the MANOVA analysis the genetic variance was partitioned with 77.7 % among individuals within the population level, 21.7 % at the population level and 0.6 % at the among group level (native and introduced) (Table 3). None of the populations showed a significant signal of a bottleneck event (The results are given in Additional file 2). Based on the population expansion analyses (k and g-tests), there were no significant signals of demographic expansion in any of the sampled populations (Additional file 2).
Gyrodactylus species diversity and prevalence
Out of the 43 surveyed populations, 15 minnow populations were not infected with Gyrodactylus spp. (3 native and 12 introduced), while 28 minnow populations had at least one or more hosts infected by one or more Gyrodactylus spp. specimens. A total of 253 individuals in the samples were infected with at least one Gyrodactylus sp. The overall prevalence (i.e. the proportion of individual minnows infected with one or more Gyrodactylus spp. in a given population) ranged between 0 and 100 %. The prevalence did not differ between native and introduced minnows (χ2 1 = 4.252, P = 0.119), and there was no effect of average heterozygosity (χ2 42 = 2.518, P = 0.113).
The number of Gyrodactylus species did not differ on minnows classified as belonging to either native or introduced populations (tested using EstimateS; χ2 1 = 0.029, P = 0.865). The diversity varied from 1 to 4 Gyrodactylus species per populations, and the predicted mean number of Gyrodactylus species was 4 species both in the native (95 % confidence interval: 3.9-4.1) and the introduced populations (3.8-4.2).
The intensity of Gyrodactylus spp. per individual minnow host ranged between 1 and 19 (see Additional file 3). When using all the 29 infected populations (880 minnows in total) it was found that the mean number of Gyrodactylus individuals (all species) per host differed significantly between the native (0.64 ± 1.34, mean ± SD) and introduced (0.35 ± 1.58) minnow groups (whole model: χ2 28 = 553.4, P < 0.0001), where the introduced group had a lower infection (Table 4). The abundance was significantly negatively associated with individual heterozygosity, and there was no interaction effect (Table 4).
Discussion
The “enemy release hypothesis” suggests that introduced species should harbour fewer parasite species than native species. The observed results do not support the enemy release hypothesis as similar numbers of Gyrodactylus species were observed on native and introduced minnow hosts, and they had the same likelihood of being infected with Gyrodactylus spp. In support of the observed results, Daverdin [64] compared some native minnow populations to an introduced minnow population (one of the lakes in our study) and observed no difference in the internal parasite fauna showing that the same fauna was established in the new environment. The same result was found in a study of the common carp (Cyprinus carpio L.) where there were no differences in helminth communities between native and introduced populations [65]. Several other studies have found some support for the enemy release hypothesis spanning a range of organisms including freshwater fish [29, 31, 33, 66, 67]. However, the success of parasite species introductions will likely depend on the complexity of the parasite lifecycle. Parasites with a lifecycle that requires more than one host will likely have a lower probability of introduction and establishment in a new environment [68] than parasites with a direct life-cycle (no intermediate hosts).
An alternative to the enemy release hypothesis is that the number of Gyrodactylus species increase in the introduced populations due to transmission of new Gyrodactylus species from other fish species already present in the new environment. However, this seems unlikely in the case of the minnow as most Gyrodactylus species seem to be host specific [22]. Further, it is possible that abiotic environmental conditions are also important factors determining species numbers [35]. Thus, if the environmental conditions differ strongly also the parasite fauna may differ. Environmental conditions are usually more similar for geographically close locations. Also, multiple introductions from the same source population would likely ensure that the whole parasite species fauna would be found in both environments. Thaulow et al. [18] have shown that multiple introductions of minnows have occurred from different sources into one of the same river systems studied here (River Skiensvassdraget). Thus, it is possible that multiple introduction events of minnows into lakes in this study could partly explain the similarity of the Gyrodactylus species fauna we observed in native and introduced minnows.
The abundance of Gyrodactylus was observed to be lower in introduced minnow compared to native populations in this study. This seems at odds with the enemy-release hypothesis. However, the enemy-release-hypothesis may be imprecise as it usually only considers presence or absence of parasite species. The parasite species-specific abundance of hosts is not taken into account. It is reasonable to assume that the more diverse parasite species infection a host has, as well as the abundance of each species, the higher challenge will be imposed on the immune system of the host [69]. Torchin et al. [29], who studied introduced and native populations of a set of diverse organisms observed that the mean number of parasite individuals within parasite species were lower in introduced than in native populations, similar to our observation of Gyrodactylus on the minnows.
Most of the studies that test the enemy release hypothesis do not have data on individual heterozygosity. In this study, we found no significant association between mean heterozygosity and the probability of being infected with Gyrodactylus (absence versus presence of infection) when using the whole dataset. However, when using only infected minnow there was a significant negative association between individual heterozygosity and mean number of Gyrodactylus. Here, the association of a higher Gyrodactylus infection level in more homozygotic minnow hosts was found for both native and introduced populations. This may indicate that more diverse hosts are better able to combat the infections. In two studies on the rostrum dace (Leuciscus leuciscus L.), Blanchet et al., [39, 70] tested if heterozygosity was associated with the mean number of the harmful fin-feeder ectoparasite Tracheliastes polycolpus Nordmann, 1832. They observed that parasite burdens were highest in hosts being moderately heterozygous, while extremely homozygous and heterozygous hosts had a lower parasite burden. This result seems in conflict with our observations for the minnow-Gyrodactylus system. However, this apparent conflict may be caused by different ranges of genetic variation among hosts in the various studies.
It is likely that the number of founder populations differ between native and introduced minnow populations. Also, the degree of heterozygosity may be associated with the success of founder populations [71]. However, the lack of significant bottleneck signals in the minnow populations suggested that no drastic decrease in genetic diversity occurred during colonization events (although the power of the test may be weak; see [57, 72]). This is also supported by the results from the AMOVA analysis, showing that only 0.6 % of the genetic variation was partitioned between the native and introduced populations. Thus, the relatively high level of genetic variation in the introduced populations could help explain that the two groups had the same number of Gyrodactylus species observed (from species accumulation curves). This evaluation is valid under the assumption that similar levels of heterozygosity reflects similar abilities to withstand negative impacts from Gyrodactylus spp. infection, and that heterozygosity based on neutral microsatellites is correlated to genetic variation in e.g. adaptive immunocompetence genes (MHC) associated to parasite resistance [40, 73].
Geographical distribution of Gyrodactylus spp. on minnows
In order to place the findings of this study in a biogeographical framework we here report on the distribution of Gyrodactylus spp. in Norway and other parts of Europe. A total of fourteen species of Gyrodactylus have been described on minnow on a global scale [22]. In this study, a total of four out of five previously reported Gyrodactylus species in Norway [50] were observed in the 43 minnow populations. To the southeast, in Sweden, two more Gyrodactylus species have been found on minnow [9]. The most plausible explanation for why Norway has a lower number of Gyrodactylus species than the rest of Europe is Norway`s relatively recent deglaciation (<10 000 years ago) and location to the west on the Scandinavian peninsula, with relatively long colonization routes from assumed glacial refugia. For minnows these refugia are probably situated in south central Europe (based on species determination of dated bones from the Eem interglacial (ca .150 000 years before present) [74], and likely also somewhere in Russia. Indeed, hosts at the geographical limits of their distribution often have fewer parasites in general or lack species-specific parasites (see [75]). In this study, a maximum of four Gyrodactylus species was observed in a minnow population. This finding is not too different from other European studies, in which a maximum of six Gyrodactylus species have been found in a single minnow population [19, 20]. In the current study, G. aphyae was found in all populations, while the three other species were more or less rare. G. phoxini was found in only three locations, detected on a few hosts only. The two most common species (G. aphyae and G. macronychus) in this study were also the most common species reported in the literature on Gyrodactylus on European minnow [19, 20]. Most commonly, only one Gyrodactylus species, rarely two, was observed on the individual hosts. If two species co-occurred, the combination of G. aphyae and G. magnificus was the most commonly observed.
The environmental conditions in a given lake may likely affect the establishment of the hosts in the new environments, as well as being important for survival and demographics of Gyrodactylus on hosts [76–79]. In this study, this could influence the mean number of Gyrodactylus as lakes are situated at different altitudes and thus comprise a range of environmental regimes for Gyrodactylus spp. Further, introduced minnow populations may need to be of a certain size in order to uphold a viable population of Gyrodactylus [80]. Also the population density and behaviour of minnows in a new location may be important with regard to horizontal transmission and population dynamics of Gyrodactylus [81].
Physio-chemical conditions of the lake environment could affect the success of establishment of minnows and its parasite fauna during introduction to new environments. Such factors could be e.g. pH and water temperature as these factors have been shown to be associated with Gyrodactylus spp. development and survival [77–79]. In our study, this could influence the results as lakes are situated at different altitudes and thus comprise differential temperature regimes for Gyrodactylus spp. However, the wide geographic range covered by both native and non-native populations suggests that such abiotic drivers do not significantly bias the results.
Conclusions
In this study it was observed that native and introduced minnow populations did not differ in their species community of Gyrodactylus spp., which lends no support to the enemy release hypothesis. However, the average number of parasites per host was higher in the native than in the introduced minnow. Interestingly, a negative association between individual minnow host heterozygosity and abundance was detected, being evident in both the native and introduced minnow populations. Our results suggest that the enemy release hypothesis does not necessarily limit fish parasite dispersal, further emphasizing the importance of invasive fish species dispersal control.
References
Vitousek PM, Mooney HA, Lubchenco J, Melillo JM. Human domination of Earth's ecosystems. Science. 1997;277(5325):494–9.
Rahel FJ, Olden JD. Assessing the effects of climate change on aquatic invasive species. Conserv Biol. 2008;22(3):521–33.
Gherardi F, Britton JR, Mavuti KM, Pacini N, Grey J, Tricarico E, et al. A review of allodiversity in Lake Naivasha, Kenya: Developing conservation actions to protect East African lakes from the negative impacts of alien species. Biol Conserv. 2011;144(11):2585–96.
Hulme PE, Pysek P, Nentwig W, Vila M. Will Threat of Biological Invasions Unite the European Union? Science. 2009;324(5923):40–1.
Perrings C, Dehnen-Schmutz K, Touza J, Williamson M. How to manage biological invasions under globalization. Trends Ecol Evol. 2005;20(5):212–5.
Pimentel D, Zuniga R, Morrison D. Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol Econ. 2005;52(3):273–88.
Peeler EJ, Oidtmann BC, Midtlyng PJ, Miossec L, Gozlan RE. Non-native aquatic animals introductions have driven disease emergence in Europe. Biol Invasions. 2011;13(6):1291–303.
Parker IM, Simberloff D, Lonsdale WM, Goodell K, Wonham M, Kareiva PM, et al. Impact: toward a framework for understanding the ecological effects of invaders. Biol Invasions. 1999;1(1):3–19.
Malmberg G. Om förekomsten av Gyrodactylus på svenska fiskar. Skrifter utgivna av Söndre Sverges fiskeriförening. 1957;1956:19–76.
Johnsen BO, Jensen AJ. The Gyrodactylus story in Norway. Aquaculture. 1991;98(1–3):289–302.
Vralstad T, Strand DA, Grandjean F, Kvellestad A, Hastein T, Knutsen AK, et al. Molecular detection and genotyping of Aphanomyces astaci directly from preserved crayfish samples uncovers the Norwegian crayfish plague disease history. Vet Microbiol. 2014;173(1–2):66–75.
Pegg J, Williams CF, Cucherousset J, Britton JR. What are the consequences of infection by the introduced parasite Philometroides sanguineus for threatened crucian carp Carassius carassius populations in England? Ecol Freshw Fish. 2011;20(4):598–604.
Rahel FJ. Homogenization of fish faunas across the United States. Science. 2000;288(5467):854–6.
Rahel FJ. Biogeographic barriers, connectivity and homogenization of freshwater faunas: it's a small world after all. Freshw Biol. 2007;52(4):696–710.
Huitfeldt-Kaas. Ferskvandsfiskenes utbredelse og indvandring i Norge, med et tillæg om krebsen. in Norwegian (Distribution and post-glacial colonisation of freshwater fishes in Norway, including the cray fish.). In: Centraltrykkeriet, Kristiania (Oslo). 1918.
Museth J, Hesthagen T, Sandlund OT, Thorstad EB, Ugedal O. The history of the minnow Phoxinus phoxinus (L.) in Norway: from harmless species to pest. J Fish Biol. 2007;71:184–95.
Holmen J: The Eurasian minnow: post-glacial dispersal history and recent invasion patterns in Norway. Ph.D. thesis: University of Oslo; 2013.
Thaulow J, Borgstrøm R, Heun M. Genetic analyses of European minnow, Phoxinus phoxinus, in a river system outside its native range indicate multiple invasions from different sources. Fisheries Manag Ecol. 2014;21(1):75–81.
Dorovskikh GN, Stepanov VG. Change in the structure of component parasite communities with host age. Russ J Ecol. 2008;39(3):215–20.
Matejusova I, Morand S, Gelnar M. Nestedness in assemblages of gyrodactylids (Monogenea : Gyrodactylidea) parasitising two species of cyprinid - with reference to generalists and specialists. Int J Parasit. 2000;30(11):1153–8.
Sterud E: Parasites of Norwegian fresh-water fish. vol. 7. Oslo: Norsk zoologisk forening; 1999: 22 s.
Harris PD, Shinn AP, Cable J, Bakke TA, Bron J. GyroDb: gyrodactylid monogeneans on the web. Trends Parasitol. 2008;24(3):109–11.
Bakke TA, Cable J, Harris PD. The biology of gyrodactylid monogeneans: The "Russian-doll killers". Adv Parasitol. 2007;64:161–330.
Poulin R: Evolutionary ecology of parasites. Second edition. Princeton University: Princeton University Press; 2007.
Karvonen A, Seehausen O. The Role of Parasitism in Adaptive Radiations-When Might Parasites Promote and When Might They Constrain Ecological Speciation? Int J Ecology. 2012;280169:280161–20.
Elton CS. The ecology of invasions by animals and plants. London: Methuen; 1958.
Wilson F. The biological control of weeds. In: Report of the Sixth Commonwealth Entomological Congress, London. 1954. p. 95–8.
Mitchell CE, Power AG. Release of invasive plants from fungal and viral pathogens. Nature. 2003;421(6923):625–7.
Torchin ME, Lafferty KD, Dobson AP, McKenzie VJ, Kuris AM. Introduced species and their missing parasites. Nature. 2003;421(6923):628–30.
Heger T, Jeschke JM. The enemy release hypothesis as a hierarchy of hypotheses. Oikos. 2014;123(6):741–50.
Roche DG, Leung B, Franco EFM, Torchin ME. Higher parasite richness, abundance and impact in native versus introduced cichlid fishes. Int J Parasit. 2010;40(13):1525–30.
Ondračková M, Šimková A, Civáňová K, Vyskočilová M, Jurajda P. Parasite diversity and microsatellite variability in native and introduced populations of four Neogobius species (Gobiidae). Parasitology. 2012;139(11):1493–505.
Lacerda ACF, Takemoto RM, Tavares-Dias M, Poulin R, Pavanelli GC. Comparative parasitism of the fish Plagioscion squamosissimus in native and invaded river basins. J Parasitol. 2012;98(4):713–7.
Gendron AD, Marcogliese DJ, Thomas M. Invasive species are less parasitized than native competitors, but for how long? The case of the round goby in the Great Lakes-St. Lawrence Basin Biol Invasions. 2012;14(2):367–84.
Halvorsen O. Studies of the helminth fauna of Norway XVII: On the composition of the parasite fauna of coarse fish in the River Glomma South Eastern Norway. Nor J Zoology. 1971;19(2):181–92.
Cornell HV, Hawkins BA. Accumulation of Native Parasitod Species on Introduced Herbivores: A Comparison of Hosts as Native and Hosts as Invaders. Am Nat. 1993;141(6):847–65.
Poulin R, Mouillot D. Host introductions and the geography of parasite taxonomic diversity. J Biogeogr. 2003;30(6):837–45.
Chapman JR, Nakagawa S, Coltman DW, Slate J, Sheldon BC. A quantitative review of heterozygosity-fitness correlations in animal populations. (vol 18, pg 2746. Mol Ecol 2011. 2009;20(12):2655.
Blanchet S, Rey O, Berthier P, Lek S, Loot G. Evidence of parasite-mediated disruptive selection on genetic diversity in a wild fish population. Mol Ecol. 2009;18(6):1112–23.
Collin H, Burri R, Comtesse F, Fumagalli L. Combining molecular evolution and environmental genomics to unravel adaptive processes of MHC class IIB diversity in European minnows (Phoxinus phoxinus). Ecology and Evolution. 2013;3(8):2568–85.
Voegeli B, Saladin V, Wegmann M, Richner H. Parasites as mediators of heterozygosity-fitness correlations in the Great Tit (Parus major). J Evol Biol. 2012;25(3):584–90.
Alvarez-Pellitero P. Fish immunity and parasite infections: from innate immunity to immunoprophylactic prospects. Vet Immunol Immunopathol. 2008;126(3–4):171–98.
Moran EV, Alexander JM. Evolutionary responses to global change: lessons from invasive species. Ecol Lett. 2014;17(5):637–49.
Hesthagen T, Sandlund OT. Fish distribution in a mountain area in south-eastern Norway: human introductions overrule natural immigration. Hydrobiologia. 2004;521(1–3):49–59.
Hesthagen T, Sandlund OT. Chanfes in the distribution of European minnow, Phoxinus phoxinus, in Norway: causes and effects, vol. 013. Norwegian Institute for Nature Researc: Trondheim; 1997.
Buchmann K, Uldal A. Gyrodactylus derjavini infections in four salmonids: Comparative host susceptibility and site selection of parasites. Dis Aquat Org. 1997;28(3):201–9.
Mo TA, Appleby C. A special technique for studyng haptoral sclerites of monogeneans. Syst Parasitol. 1990;17(2):103–8.
Ziętara MS, Lumme J. The crossroads of molecular, typological and biological species concepts: two new species of Gyrodactylus Nordmann, 1832 (Monogenea : Gyrodactylidae). Syst Parasitol. 2003;55(1):39–52.
Shinn AP, Collins C, Garcia-Vasquez A, Snow M, Matejusova I, Paladini G, et al. Multi-centre testing and validation of current protocols for the identification of Gyrodactylus salaris (Monogenea). Int J Parasit. 2010;40(12):1455–67.
Sterud E. Parasitter hos norske ferskvannsfisk. Norsk Zoologisk Forening. 1999;7:1–22.
Holmen J, Vøllestad LA, Jakobsen KS, Primmer CR. Cross-species amplification of zebrafish and central stoneroller microsatellite loci in six other cyprinids. J Fish Biol. 2005;66(3):851–9.
Peakall R, Smouse PE. GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes. 2006;6(1):288–95.
Goudet J. FSTAT (Version 1.2): A computer program to calculate F-statistics. J Hered. 1995;86(6):485–6.
Excoffier L, Smouse PE, Quattro JM. Analyse of Molecular Variance Innferred From Metric Distances Among DNA haplotypes: Application to Human Mitochondrial DNA Restriction Data. Genetics. 1992;131(2):479–91.
Peakall R, Smouse PE. GenAlEx 6.5: genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics. 2012;28(19):2537–9.
Piry S, Luikart G, Cornuet JM. BOTTLENECK: A computer program for detecting recent reductions in the effective population size using allele frequency data. J Hered. 1999;90(4):502–3.
Cornuet JM, Luikart G. Description and power analysis of two tests for detecting recent population bottlenecks from allele frequency data. Genetics. 1996;144(4):2001–14.
Reich DE, Feldman MW, Goldstein DB. Statistical properties of two tests that use multilocus data sets to detect population expansions. Mol Biol Evol. 1999;16(4):453–66.
Reich DE, Goldstein DB. Genetic evidence for a Paleolithic human population expansion in Africa. Proc Natl Acad Sci U S A. 1998;95(14):8119–23.
Colwell R, K.: EstimateS 8.2.0: Statistical estimation of species richness and shared species from samples. User’s Guide and application published at: http://viceroy.eeb.uconn.edu/estimates/.
Gotelli NJ, Colwell RK. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett. 2001;4(4):379–91.
Longino JT, Coddington J, Colwell RK. The ant fauna of a tropical rain forest: Estimating species richness three different ways. Ecology. 2002;83(3):689–702.
Sall J, Creighton L, Lehman A. JMP start statistics: a guide to statistics and data analysis using JMP. Cary, N.C.: SAS Pub; 2007.
Daverdin RH. Dissemination of parasites through introduction of freshwater fish species: introduction of minnow (Phoxinus phoxinus) parasites to high altitude lakes with brown trout (Salmo trutta), vol. 466. Trondheim: Norwegian Institute for Nature Research; 1997.
Kennedy CR, Pojmanska T. Richness and diversity of helminth parasite communities in the common carp and in three more recently introduced carp species. J Fish Biol. 1996;48(1):89–100.
Kennedy CR, Hartvigsen R, Halvorsen O. The importance of fish stocking in the dissemination of parasites throughout a group of reservoirs. J Fish Biol. 1991;38(4):541–52.
Ondračková M, Matějusová I, Grabowska J. Introduction of Gyrodactylus perccotti (Monogenea) into Europe on its invasive fish host, Amur sleeper (Perccottus glenii, Dybowski 1877). Helminthologia. 2012;49(1):21–6.
Bauer ON. Spread of parasites and diseases of aquatic organisms by acclimation - ashort review. J Fish Biol. 1991;39(5):679–86.
Zuk M, Stoehr AM. Immune defense and host life history. Am Nat. 2002;160:S9–S22.
Blanchet S, Rey O, Loot G. Evidence for host variation in parasite tolerance in a wild fish population. Evol Ecol. 2010;24(5):1129–39.
Forsman A. Effects of genotypic and phenotypic variation on establishment are important for conservation, invasion, and infection biology. Proc Natl Acad Sci U S A. 2014;111(1):302–7.
Peery MZ, Kirby R, Reid BN, Stoelting R, Doucet-Bëer E, Robinson S, et al. Reliability of genetic bottleneck tests for detecting recent population declines. Mol Ecol. 2012;21(14):3403–18.
Monzon-Arguello C, de Leaniz CG, Gajardo G, Consuegra S. Eco-immunology of fish invasions: the role of MHC variation. Immunogenetics. 2014;66(6):393–402.
Movius HL. Radiocarbon Dates and Upper Palaeolithic Archaeology in Central and Western Europe. Curr Anthropol. 1960;1(5–6):355–91.
Dogiel VA: General parasitology. 3rd Edit., revd. Polyanski, Yu. I. & Khaeisin, E. M., transl. Kabata, Z. (Oliver & Boyd). Edinburgh 1964.
Kennedy CR. Parasite fauna of resident char (Salvelinus alpinus) from Arctic islands, with special reference to Bear Island. J Fish Biol. 1978;13(4):457–66.
Marcogliese DJ, Cone DK. On the distribution and abundance of eel parasites in Nova Scotia: Influence of pH. J Parasitologi. 1996;82(3):389–99.
Pettersen RA, Vøllestad LA, Flodmark LEW, Poléo ABS. Effects of aqueous aluminium on four fish ectoparasites. Sci Total Environ. 2006;369(1–3):129–38.
Poléo ABS, Schjolden J, Hansen H, Bakke TA, Mo TA, Rosseland BO, et al. The effect of various metals on Gyrodactylus salaris (Platyhelminthes, Monogenea) infections in Atlantic salmon (Salmo salar). Parasitology. 2004;128:169–77.
Lyles AM, Dobson AP. Infectious disease and intensive management population dynamics, threatened hosts, and their parasites. J Zoo Wildl Med. 1993;24(3):315–26.
Bernhardt B, Lampert KP, Leese F, Mayer C, Tollrian R. Are shoals of minnow Phoxinus phoxinus formed by close kin? J Fish Biol. 2012;80(3):713–21.
Acknowledgements
First of all, we want to thank all the people who sampled minnow for us. We further thank Nanna Winger Steen and Emelita Rivera Nerli for technical assistance in the laboratory. The collection of fish and genotyping was financially supported by grants from the Norwegian Research Council, the Norwegian Environment Agency and the Finnish Academy.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors' contributions
RAP, KØ, JH, LAV and TAM, designed the study. JH collected the minnow. RAP and JH performed data collection. RAP, KØ, JH and LAV analysed the results. RAP, KØ, JH, LAV and TAM co-drafted the manuscript together. All authors read and approved the final version of the manuscript.
Additional files
Additional file 1: Table S1.
A table of other fish species in the sampled localities. All the Norwegian minnow (Phoxinus phoxinus) populations used in the study and the other fish species in the same locality. The species of fish are: AC = Arctic charr (Salvelinus alpinus) BL = bleak (Alburnus alburnus), BR = bream (Abramis brama), BT = brown trout (Salmo trutta), BU = burbot (Lota lota), CC = crucian carp (Carassius carassius), CH = chub (Leuciscus cephalus), EE = eel (Anguilla anguilla), GR = grayling (Thymallus thymallus), PE = perch (Perca fluviatilis), PI = pike (Esox lucius), RU = ruffe (Gymnocephalus cernuus), SS = Siberian sculpin (Cottus poecilopus), VE = vendace (Coregonus albula), WB = white bream (Blicca bjoerkna), WF = European whitefish, SM = smelt (Osmerus eperlanus), ID = ide (Leuciscus idus), DA = dace (Leuciscus leuciscus), 9-SB = nine-spined stickleback (Pungitius pungitius) and 3-SB = tree-spined stickleback (Gasterosteus aculeatus). (DOC 192 kb)
Additional file 2:
A table of tests result of demographic bottleneck. All the Norwegian minnow (Phoxinus phoxinus) populations used in the study. Information from tests of demographic bottleneck using Bottleneck 1.2.0.2. (Piry et al. 1999) with the two-phase model (TPM) and the Wilcoxon sign-rank test, and demographic expansion (where the intra-locus k-test identify signals of recent population expansion, while the inter-locus g-test identify signals of more ancient population expansion) using Kg-test (Reich et al. 1999). Footnotes: *Samples used in the rarefaction analysis using EstimateS 8.2.0 (Colwell 2011) where more than 5 Gyrodactylus spp. individuals were observed in the population. This number of observed Gyrodactylus spp. individuals is needed to calculate rarefaction in EstimateS 8.2.0. # None of tests were significant. (DOC 116 kb)
Additional file 3:
Figure of distribution of Gyrodactylus spp. individuals. Distribution of Gyrodactylus spp. individuals in the total dataset of 1278 individual minnows (Phoxinus phoxinus) hosts from 43 populations in Norway. Here the 14 native and 23 stocked populations have been pooled. On the left axis is the probability of occurrence and on right axis is given counts. (DOCX 76 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Pettersen, R.A., Østbye, K., Holmen, J. et al. Gyrodactylus spp. diversity in native and introduced minnow (Phoxinus phoxinus) populations: no support for “the enemy release” hypothesis. Parasites Vectors 9, 51 (2016). https://doi.org/10.1186/s13071-016-1306-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-016-1306-y