
Abstract
Background
Scrub typhus is a lethal infectious disease vectored by larval trombiculid mites (i.e. chiggers) infected with Orientia tsutsugamushi (OT) and recent decades have witnessed an emergence of scrub typhus in several countries. Identification of chigger species and their vertebrate hosts is fundamental for the assessment of human risks to scrub typhus under environmental changes, but intensive and extensive survey of chiggers and their hosts is still lacking in Taiwan.
Methods
Chiggers were collected from shrews and rodents in nine counties of Taiwan and were assayed for OT infections with nested polymerase chain reaction (PCR). PCR products were further sequenced to reveal probable OT strains. Rodents were assessed for OT exposure by immunofluorescent antibody assay. Lastly, incidence rate of scrub typhus in each county was associated with loads and prevalence of chigger infestations, seropositivity rate in rodents, and OT positivity rate in chiggers.
Results
Rattus losea was the most abundant (48.7 % of 1,285 individuals) and widespread (occurred in nine counties) small mammal species and hosted the majority of chiggers (76.4 % of 128,520 chiggers). Leptotrombidium deliense was the most common (64.9 % of all identified chiggers) and widespread (occurred in seven counties) chigger species but was replaced by Leptotrombidium pallidum or Leptotrombidium scutellare during the cold seasons in two counties (Matsu and Kinmen) where winter temperatures were lower than other study sites. Seropositivity rate for OT exposure in 876 assayed rodents was 43.0 % and OT positivity rate in 347 pools of chiggers was 55.9 %, with 15 OT strains identified in the 107 successfully sequenced samples. Incidence rate of scrub typhus was positively correlated with chigger loads, prevalence of chigger infestations, seropositivity rate but not OT positivity rate in chiggers.
Conclusions
Our study reveals R. losea as the primary host for chiggers and there exists a geographical and seasonal variation in chigger species in Taiwan. It also emphasizes the importance of recognition of chigger vectors and their vertebrate hosts for a better prediction of human risks to scrub typhus under rapid environmental changes.
Similar content being viewed by others


Background
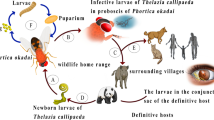
Scrub typhus is a potentially severe febrile disease transmitted by trombiculid mites infected with rickettsia Orientia tsutsugamushi (OT). Prevalent mainly along the western Pacific, about one million human cases occur each year and an estimated one billion people are at risk of this disease [1–3]. Except for parts of Asia, Australia, and Oceania where scrub typhus is traditionally endemic, suspected human cases have also been reported in Africa (Cameroon) and South America (Chile) [4, 5]. In addition, Orientia-like bacteria have been detected in rodents in France and Senegal [6]. In recent decades, several countries have experienced a great increase in the incidence of scrub typhus, including South Korea and China [7, 8]. In fact, scrub typhus is one of the most severely neglected diseases around the world [9, 10].
Among the different life stages of trombiculid mites, only larvae are parasitic, with rodents as the primary hosts and humans as the accidental hosts; nymphs and adults are free living in the soil, feeding mainly on arthropods [11]. Larval trombiculid mites (so called chiggers) are thus of significant medical importance because of their potential for transmitting scrub typhus to humans. Identification of chigger species is particularly fundamental for the evaluation of human risks because only a subset of chigger species, (mostly the genus Leptotrombidium [12]) are responsible for transmitting scrub typhus. Furthermore, even among the genus Leptotrombidium, chigger species vary in the seasonal occurrence and likely also virulence in the OT strains they carry. For instance, in Japan, Leptotrombidium akamushi occurs mainly in summer while Leptotrombidium pallidum and Leptotrombidium scutellare appear primarily from fall to early spring [11]. Besides, OT strains transmitted by L. akamushi and L. pallidum could be severe; in comparison, L. scutellare-borne OT might be relatively mild, although chigger-OT strain relationship is not fully understood [13, 14]. Recognition of chigger species is also essential for the elucidation of mechanisms underlying a potential surge in human incidence after global warming: such heighted risks could be due to an increase in chigger abundance or alternatively, the consequence of a change in chigger species. Similarly, when investigating environmental associations with spatial variation in incidence of scrub typhus (e.g. [15, 16]), what is actually modeled is how environments influence abundance of chiggers, which in turn determine human risks to scrub typhus. An implicit assumption for such investigation is that chiggers at different locations respond to environments identically, which might not be true when the primary vector species vary geographically. This can be exemplified, as mentioned above, by the difference in seasonal occurrence of L. akamushi versus L. pallidum or L. scutellare, which is more abundant at higher and lower temperatures, respectively [11].
Rodents are the main hosts of chiggers (as food resources) although their role in sustaining OT transmission (i.e. as a reservoir) requires further clarification [9, 11]. Rodents are thus indispensable for the survival of chiggers and play a key role in the transmission of scrub typhus. Because intensity of chigger infestations, even feeding success of chiggers, could vary considerably with rodent species [17, 18], it is imperative to recognize the primary host species. This will help predict whether a change in land use might enhance the risks to scrub typhus by favouring the survival of main rodent host species. For example, a transition to agricultural cultivation might increase the number of chiggers if their principal rodent hosts also flourish in agricultural fields. A good knowledge on the relative importance of rodent species might also assist the forecast of human health outcomes after an invasion of exotic rodent species [19].
In Taiwan, studies on vectors of scrub typhus are limited to a few places and no systematic survey has ever been implemented. Leptotrombidium deliense was found to be the primary chigger species in Penghu (Pescadores Islands, [20]) and Leptotrombidium imphalum was the most common species in Hualien [17]. In Kinmen Islands, occurrence of chigger species varies with seasons, with L. deliense as the main species in summer and L. scutellare the principal species in winter [21]. These sporadic studies suggest a geographical variation in chigger species and call for a more extensive survey of chiggers in Taiwan. Similarly, primary rodent hosts of chiggers have only been reported in a few localities in Taiwan, including Penghu, Kinmen, and Hualien [17, 21, 22]. Hosts of chiggers were surveyed in 15 counties in Taiwan but only an overall status was briefly reported; significance of local host species were only documented in one county [23]. Recently, hosts of chiggers were studied in six counties in Taiwan but only prevalence of chigger infestation (i.e. whether or not a host was infested with chiggers) was reported [24]; knowledge on the intensity of chigger infestation (i.e. the number of chiggers infested in each host) was still lacked for most regions of Taiwan.
In this study, chiggers and their associated hosts were systematically surveyed in different parts of Taiwan. In addition, we reported variation in seropositivity rate of OT infections among host species and geographical regions. Positivity rate of OT infections in chiggers and the probable OT strains were also studied. Lastly, we investigated whether geographic variation in human incidence of scrub typhus was correlated with prevalence and intensity of chigger infestations, OT seropositivity rate in rodent hosts, and OT positivity rate in chiggers. To our knowledge, this is the most extensive and comprehensive study on hosts and vectors of scrub typhus in Taiwan.
Methods
Study sites and small mammal trapping
From 2006 to 2010, small mammals (rodents and shrews) were trapped in different parts of Taiwan, including eastern (Yilan, Hualien, Taitung), western (Taoyuan, Taichung, Kaoping), and main islets near Taiwan (Matsu, Kinmen, Penghu) (Fig. 1). These nine counties vary profoundly in the incidence rate of scrub typhus (2001–2005, 0.2 to 125.2 cases per 100,000 people per year; on-line data, Taiwan National Infectious Disease Statistics System, http://nidss.cdc.gov.tw/). Mean temperature for the hottest month (typically in July in Taiwan) is similar among the nine study sites (27.1 °C–29.3 °C, Taiwan Central Weather Bureau) while mean temperature for the coldest month (typically in January) is more varied (8.9 °C–19.5 °C), with lower January temperature occurring in Matsu (8.9 °C) and Kinmen (12.8 °C) (Fig. 1a). Because scrub typhus prevails mainly in rural areas and occurs outside house [12], trappings were limited to the field but not implemented inside or close to human buildings. At each county, 80 Sherman traps (26.5 × 10 × 8.5 cm) and 80 Taiwan made rodent traps (27 × 16 × 13 cm) were deployed and baited with sweet potato covered with peanut butter. Each county was surveyed for four consecutive nights and surveyed at least twice (Yilan: March and August 2009; Hualien: May and November 2006, April 2009; Taitung: September 2006, June and September 2009; Taoyuan: March and October 2010; Taichung: March 2009, July 2010; Kaoping: February and May 2010; Matsu: March and July 2007; Kinmen: June 2006, January 2007, March 2008; Penghu: May and September 2007). Each night, traps were set up at different locations within the same county to maximize trapping coverage. Trapped small mammals (shrews and rodents) were euthanized with an overdose of Zoletil 50 (Virbac SA, Carros, France), and blood was collected by cardiac puncture. Chiggers recovered from small mammals were preserved in 70 % ethanol and stored at−70 °C until subsequent molecular detection of OT.

Study sites for the collection and identification of Orientia tsutsugamushi (OT) strains in chiggers from small mammal hosts in Taiwan during 2006–2010. a mean temperature for the coldest and hottest months (shown in parenthesis); b OT strains and genotypes (in parenthesis) identified in chiggers
Species identification of chiggers
Chiggers were soaked in deionized water 2–3 times (30 min each), and then slide-mounted in Berlese fluid (Asco Laboratories, Manchester, UK). Chiggers were examined under a light microscope and identified with published keys to full species level based on a combination of characteristics of scutum, gnathosoma, and ventral and dorsal seta following [25, 26].
Immunofluorescent antibody assay (IFA)
Seroprevalence of OT exposure in rodents was assessed following [17]. Briefly, each rodent serum sample was diluted 1:40 in phosphate-buffered saline (PBS), applied to slides coated with OT antigens (Gu-Yuan Biotech. Ltd., Taiwan), and mixed by pipetting with fluorescein isothiocyanate–goat anti-mouse immunoglobulin G (IgG) + A + M (H + L; Zymed Laboratories, Inc., San Francisco, CA) diluted 1:40 in PBS. Shrew serum was not assessed to avoid false negativity due to our application of anti-mouse but not anti-shrew secondary antibodies. We chose 1:40 as the cutoff titer because this is the same criterion used for confirmation of human cases of scrub typhus in Taiwan by the Taiwan Centers for Disease Control. OT antigen slides allowed simultaneous screening of three strains (Kato, Karp, and Gilliam). Serum samples were scored negative for OT when all three strains were negative; if any strains yielded positive results, the sample was recorded as positive.
Detection of OT in chiggers with nested PCR
Pools of chiggers (largely with 100 chiggers for each pool to retrieve enough DNA) were detected for OT infections with nested polymerase chain reaction (PCR) following [27]. This method targeted well conserved DNA corresponding to a 56-kDa type specific antigen located on the OT outer membrane. Primers for the 1st stage PCR were 56 kDa-1 F: 5’-AGAATCTGCTCGCTTGGATCCA-3’ and 56 kDa-1R: 5’-ACCCTATAGTCAATACCAGCACAA-3’ and for the 2nd stage: 56 kDa-2 F: 5’-GAGCAGAGATAGGTGTTATGTA-3’ and 56 kDa-2R: 5’-TATTCATTATAGTAGGCTGA-3’. Laboratory Karp and Gilliam strains and deionized water were used as positive and negative controls, respectively. The PCR products were separated by electrophoresis in 3 % NuSieve and 1.0 % agarose gels, stained with ethidium bromide, and identified under UV fluorescence. The PCR products were purified with a QIA quick Gel Extraction Kit (Qiagen GmbH, Hilden, Germany) and then sequenced twice in each direction. DNA nucleotide sequences were assessed with the Basic Local Alignment Search Tool (www.ncbi.nlm.nih.gov) for any resemblance to known OT strains. Because of the minute size of chiggers, identification of chiggers to full species requires mounting specimens with the use of Berlese fluids, which destroys DNA material and renders the detection of OT unlikely. Chigger pools were thus selected randomly irrespective of species identity.
Incidence rate of scrub typhus
Information on human cases of scrub typhus, a notifiable diseases in Taiwan, was retrieved from the Taiwan National Infectious Disease Statistics System (http://nidss.cdc.gov.tw/) maintained by Taiwan Centers for Disease Control. Details on presumptive location of infections and the date for the onset of diseases were available from the database. We enumerated the number of human cases in the towns (a county includes several towns) where small mammals were trapped and included only cases reported during the period when trapping was implemented. Due to the relatively few human cases of scrub typhus in most of the towns, the period during which human cases were tallied included not only the month when small mammals were trapped but also its preceding and following month (totaling three months; this will increase human cases and thus statistical power). Population size for each town when trapping was implemented was obtained from the Department of Statistics of the Taiwan Ministry of the Interior. Towns were aggregated into counties for the calculation of incidence rate, which was defined as number of human cases per 100000 residents per month.
Statistical analysis
Correlation analyses were applied when studying association between loads and prevalence of chigger infestations, OT seropositivity rate in rodents, OT positivity rate in chiggers, and human incidence rate of scrub typhus. Because normality could not be achieved for most of the analyses even after data transformation, non-parametric Spearman rank correlations were used for all of the correlational studies.
Ethical considerations
This study was implemented with the permission of Taiwan Centers for Disease Control and all trapping and handling procedures met Taiwanese legal requirements.
Results
Small mammal trapping
A total of 1,285 small mammals were trapped, including one species of shrew (Suncus murinus) and 10 species of rodents (Table 1). Among all species, Rattus losea was the most abundant (48.7 % of total captures), followed by S. murinus (21.2 %) and Mus caroli (11.2 %). R. losea was also the most widely distributed species, occurring in all nine counties. On the contrary, Rattus tanezumi were trapped only in Lanyu Island (Orchid Island) in Taitung county, Rattus exulans only in Hualien county, and the squirrel Callosciurus erythraeus only in Yilan county (Table 1).
Prevalence and loads of chiggers among small mammal species
Prevalence of chigger infestations among host species varied from 9.1 % to 100 %, disregarding the very few captures of C. erythraeus and Niviventer coninga (Table 1). All R. tanezumi trapped in Lanyu Island were infested with chiggers (100 % prevalence). R. losea (61.3 %), Apodemus agrarius (54.2 %), and Bandicota indica (45.8 %) were also frequently infested (Table 1).
Mean number of chiggers per host varied profoundly among small mammal species, too. R. tanezumi had the highest chigger loads (394 ± 28.5, mean ± SE), followed by R. losea (156.9 ± 11.2). In comparison, chigger loads were much lower in M. caroli (1.9 ± 0.5) and M. musculus (2.3 ± 1.1) (Table 1). There was a significant correlation between prevalence and loads of chigger infestations in host species (Spearman rank correlation, r s = 0.84, d.f. = 10, P < 0.005).
A total of 128,520 chiggers were collected from small mammal hosts. 76.4 % of these chiggers were hosted by R. losea. This was followed by R. tanezumi (14.7 %). Each of the other host species supported <5 % of chiggers (Table 1).
Chigger species composition among small mammal species
Among the 2,860 identified chiggers this included eight Leptotrombidium species, three Walchia species, and one Eutrombicula species (Table 2). L. deliense was the most common species (64.9 % of all identified chiggers). L. imphalum (10.8 %), Walchia pacifica (7.0 %), Walchia xishaensis (4.1 %), L. pallidum (4.0 %), and L. scutellare (3.7 %) were uncommon while the other six chigger species were rarely observed (Table 2).
L. deliense was the most dominant chigger species in small mammal hosts except in A. agrarius and R. exulans, which harboured more L. imphalum than L. deliense (M. caroli excluded for its small number of samples). A moderate proportion of chiggers (32.3 %) in B. indica also comprised of W. pacifica (Table 2).
Seroprevalence among small mammal species
Seropositivity rate for the 876 rodents assayed for OT exposure was 43.0 %. Positivity rate differed significantly among rodent species, ranging from 15.4 % to 91.7 % when C. erythraeus and N. coninga were excluded (both with small sample size of one). There was a tendency for rodent species with higher chigger loads to have higher seropositivity rate (r s = 0.64, d.f. = 7, P = 0.08) except for M. musculus, which had low chigger load (2.3 chiggers per mouse) but with moderate seropositivity rate (56.5 %) (r s = 0.89, d.f. = 6, P < 0.01 after M. musculus excluded from analysis). Accordingly, seropositivity rate was higher in R. tanezumi (91.7 %), R. losea (46.2 %) and A. agrarius (57.9 %) and was lower in M. caroli (16.1 %) and R. exulans (15.4 %) (Table 1).
PCR positivity rate and OT strains identified in chiggers
A total of 347 pools of chiggers were detected for 56 kDa-PCR positivity, with a positivity rate of 55.9 % (Table 1). Both R. tanezumi and R. losea that had high chigger loads and seropositivity rates also harboured chiggers with higher OT positivity rate (95.8 % and 54.8 %, respectively). In comparison, R. exulans carried chiggers with moderate positivity rate (66.7 %) even with low chigger loads and seropositivity rate while A. agrarius had moderate chigger loads and seropositivity rate but with very low positivity rate in chiggers (0 %) (Table 1). There was no significant correlation between chigger loads and OT positivity rate in chiggers (r s = 0.27, d.f. = 6, P > 0.05); likewise, there was no significant correlation between seropositivity rate and OT positivity rate (r s = 0.17, d.f. = 5, P > 0.05).
A total of 15 OT strains belonging to five genotypes were identified in the 107 successfully sequenced samples (Table 3). TP0607a was the most common strain (33.6 % of samples), followed by KM0606a (15.9 %), TP0708a (11.2 %), KM0605a (10.3 %), and MZ01-2 (6.5 %). Each of the remaining strains comprised <5 % of all sequenced samples (Table 3). The five genotypes, starting from the commonest, were Karp (66.4 %), Kawasaki (24.3 %), Kato (7.5 %), Gilliam (0.9 %), and TA678 (0.9 %).
Geographical variation in incidence rate of scrub typhus, chigger infestations, and OT occurrence
Because small mammal species varied in the extent of chigger infestations (Table 1), geographical variation would ideally be compared based on a single host species to control for any spatial variation in the composition of local small mammal species. R. losea was thus selected for comparison because of its abundance in all study sites.
Incidence rate of scrub typhus (cases per 100000 residents per month) varies significantly among the nine counties, ranging from zero (Kaoping, Taichung, and Yilan) to 12.13 (Penghu) (Table 4). There was also a profound geographical variation, with R. losea as the proxy, in the chigger loads, prevalence of chigger infestations, seropositivity rate, and OT positivity rate in chiggers (Table 4). Counties with higher incidence rate also harboured R. losea with higher chigger loads (r s = 0.75, d.f. = 8, P < 0.05), higher prevalence of chigger infestations (r s = 0.73, d.f. = 8, P < 0.05), and higher seropositivity rate (r s = 0.80, d.f. = 8, P < 0.05). There was nevertheless no correlation between incidence rate and OT positivity rate in chiggers (r s = −0.23, d.f. = 6, P > 0.05).
Geographical variation in chigger species composition
L. deliense was the dominant chigger species (>70 % of total chiggers) in most study sites (disregarding Kaoping and Taoyuan for collecting no or very few chiggers). However, L. imphalum and L. pallidum were more common in Hualien and Matsu, respectively (Table 5). Besides, there was a clear seasonal variation in the occurrence of chigger species in Matsu. L. deliense and Leptotrombidium kawamurai were only collected in the summer (July) while L. pallidum, L. scutellare, and Leptotrombidium yui were only observed in the early spring (March). Similarly, in Kinmen, L. deliense was found exclusively in the summer (June) and L. scutellare solely in the winter (January).
Geographical variation in OT strains and genotypes
TP0607a and TP0708a were the most widely distributed OT strain, followed by KM0605a (Table 3). Among the five OT genotypes, Karp was identified in all study sites (excluding Taoyuan and Kaoping where no chiggers were detected for OT), and Kawasaki was also not uncommon, especially in eastern Taiwan (Fig. 1b). Eastern Taiwan also harboured the most diverse OT strains and genotypes, especially Hualien and Taitung (Fig. 1b).
Discussion
In this study, R. losea was found to be the most abundant and widespread rodent species in Taiwan; it also hosted the highest proportion of chiggers among all small mammal species. L. deliense was identified as the dominant chigger species in Taiwan but its occurrence was limited to certain seasons (mainly summer) in some study sites (Kinmen and Matsu).
R. losea commonly occurs in agricultural fields and grasslands in lowland Taiwan, including most islets near Taiwan ([28]; this study, data not shown). In Kinmen, this species was sometimes classified as Rattus flavipectus (e.g. [21, 24]), but it was recently confirmed to be R. losea based on molecular evidence [29]. We have found that R. losea hosted the majority of chiggers (76.4 %) and OT positivity rate in these chiggers was higher than those collected from most other host species. Moreover, our previous study revealed a higher feeding success of chiggers on R. losea than on the other two main host species in eastern Taiwan [18]. R. losea thus deserves special attention when rodent control is envisaged as a viable strategy for lowering the burden of scrub typhus. Because R. losea rarely occurs in forests (including secondary forests), which is instead inhabited by N. coninga [30, 31], preserving lowland forests in Taiwan could potentially mitigate the risks to scrub typhus through a reduction in the occurrence of R. losea. This will also depend on the relative importance of N. coninga as hosts of chiggers, which to our best knowledge, has never been intensively investigated before. But the low population density of N. coninga in Taiwan [31] suggests that R. losea might be a better host of chiggers than N. coninga in lowland Taiwan.
Among all vertebrate hosts, R. tanezumi had the highest loads and prevalence of chigger infestations, OT seropositivity rate in rodents, and OT positivity rate in chiggers. This rodent species was recognized as Rattus rattus before (e.g. [24]); however, R. rattus in Taiwan should instead be R. tanezumi [32]. R. tanezumi commonly occurs near human dwellings in Taiwan, but in this study, it was trapped only in Lanyu Island. This is due to the fact that we deployed traps in the field but not inside or close to human buildings and Lanyu Island is unusual in that R. tanezumi can be easily found in grassland and forest in Lanyu [33]. High chigger infestation and OT positivity rate in R. tanezumi should reflect a high risk to scrub typhus in Lanyu. Indeed, seropositivity rate for scrub typhus was extremely high (100 %) when children in Lanyu reached 7 years old [34]. Why Lanyu Island shelters abundant chiggers remains unclear and should warrant further investigation.
L. deliense is the most dominant and widespread vector of scrub typhus in the tropics of Asia and southwest Pacific islands, including Malaysia, New Guinea, Philippine, Thailand and many other countries [11]. In this study, L. deliense was also found to be the most common and widely distributed chigger species in Taiwan. Because of the high infectivity rate of OT in L. deliense and L. deliense can readily bite humans [26], the commonness of this chigger species in Taiwan indicates that people in Taiwan could be at high risk of scrub typhus infection. However, in Matsu and Kinmen, two offshore islands with lower winter temperature than the other study sites (Fig. 1a), L. deliense occurred only in the summer. In Kinmen, this species was replaced by L. scutellare in the winter. Likewise, L. pallidum, L. scutellare, and L. yui appeared in the cooler season in Matsu. Both L. pallidum and L. scutellare have been found in Japan and South Korea and occur mainly from fall to early spring [11, 35, 36]. In Yunnan of China, L. deliense is distributed at lower latitudes and L. scutellare at higher latitudes [37]. In South Korea, L. scutellare occurs primarily in the southern part of the country while L. pallidum in found more in northern regions [36]. The studies in China, Japan, and South Korea indicate that relative to L. deliense, L. pallidum and L. scutellare live in cooler environments, and L. pallidum can survive at further lower temperatures than L. scutellare. This agrees with our observation that L. pallidum and L. scutellare occurred only during cooler seasons and L. pallidum was distributed in Matsu rather than Kinmen, where winter temperature is lower in Matsu (8.9 °C) than in Kinmen (12.8 °C).
For the past 10 years (2005–2014), a total of 689 cases of scrub typhus were recorded in Kinmen (on-line data, accessed 26th July 2015, Taiwan National Infectious Disease Statistics System, http://nidss.cdc.gov.tw/). Most of these human cases (671 cases, or 97.4 % of all cases) occurred from late spring (May) to mid-fall (October). Likewise, 92.4 % of the 144 cases of scrub typhus from 2005 to 2014 in Matsu happened between June and October. Seasonal occurrence of human cases in Kinmen and Matsu roughly matched the seasonal appearance of L. deliense, suggesting that L. deliense was the main vector in both islands; our previous study in Kinmen [21] arrived at a similar conclusion. It is unexpected, however, that there were very few human cases from late fall to early spring in Kinmen and Matsu when L. pallidum or L. scutellare appeared because both species have been proven to be vectors of scrub typhus in Japan and South Korea [11, 35, 36]. The causes for such a lack of cases during the winter are unknown, and could be due to less human outdoor activity during the winter or other not yet recognized reasons. However, the number of cases peaking during the warm season implies that global warming might increase incidence of scrub typhus or lengthen the period during which scrub typhus prevails in Kinmen and Matsu. Monitoring abundance of main chigger vectors and their association with meteorological factors and human incidence is thus needed.
Besides incidence rate of scrub typhus, there also existed a profound geographical variation in loads and prevalence of chigger infestations in rodents (Table 4). The reasons for such spatial variation have previously been explored but conclusive remarks can only be made after moving beyond simple correlational studies. Incidence rate of scrub typhus on the main island of Taiwan was elevated in districts with a higher proportion of dry-field farmers in the population, a higher normalized difference vegetation index and lower mean annual temperature [15]. It was further revealed that incidence rate also increased with the proportion of land that contained mosaics of cropland and natural vegetation, which may be because a mixture of crop and vegetation habitats are ideal for rodents by providing easy access to abundant food from nearby vegetative shelters [16]. Chiggers could also proliferate in this kind of favourable habitat with abundant rodent hosts and natural vegetation that helps provide moist terrain essential for chigger survival [27]. Indeed, lower chigger infestations occur in highly populated western Taiwan where less natural vegetation survives intensive human exploitation.
With R. losea as a surrogate, we found that incidence rate of scrub typhus was positively correlated with chigger loads, prevalence of chigger infestations and seropositivity rate but not OT positivity rate in chiggers. In addition, more than half (>0.732 = 0.533) of the variation in incidence rate can be accounted for by chigger loads, chigger prevalence, or seropositivity rate. This is despite the fact that human infection to scrub typhus is a product of several factors, at least including number of questing chiggers (which may or may not be correlated with loads of chiggers in vertebrate hosts), infectivity rate of OT in questing chiggers, and intensity of human activity in infested areas. Such significant association is useful because in our experience, surveying questing chiggers, including the method such as black plates, could be very inefficient particularly when few chiggers exist. Our study indicates that instead, by examining hosts, each of these indices (loads and prevalence of chigger infestations on hosts or seropositivity rate of hosts) can be a useful proxy for relative human risks to scrub typhus, as has been found in South Korea where incidence rate increased with the load of chiggers on rodents [38].
Conclusions
Scrub typhus is emerging in many regions and whether the rapid increase in incidence is associated with global warming or alteration of habitat is still unknown. Our study reveals R. losea as the primary host for chiggers and there exists a geographical and seasonal variation in chigger species in Taiwan. We argue that only with a solid knowledge on chigger vectors and their vertebrate hosts can we start to predict the consequence of environmental change for human risks to scrub typhus.
References
Rosenberg R. Drug-resistant scrub typhus: paradigm and paradox. Parasitol Today. 1997;13:131–2.
Watt G, Parola P. Scrub typhus and tropical rickettsioses. Curr Opin Infect Dis. 2003;16:429–36.
Kelly DJ, Fuerst PA, Ching WM, Richards AL. Scrub typhus: the geographic distribution of phenotypic and genotypic variants of Orientia tsutsugamushi. Clin Infect Dis. 2009;48 Suppl 3:S203–230.
Ghorbani RP, Ghorbani AJ, Jain MK, Walker DH. A case of scrub typhus probably acquired in Africa. Clin Infect Dis. 1997;25:1473–4.
Balcells ME, Ricardo R, Patricia G, Helena P, David O, Marcela C, et al. Endemic scrub typhus-like illness, Chile. Emerg Infec Dis. 2011;17:1659–63.
Cosson J, Galan M, Bard E, Razzauti M, Bernard M, Morand S, et al. Detection of Orientia sp. DNA in rodents from Asia, West Africa and Europe. Parasit Vectors. 2015;8:172.
Wei Y, Huang Y, Luo L, Xiao X, Liu L, Yang Z. Rapid increase of scrub typhus: an epidemiology and spatial-temporal cluster analysis in Guangzhou City, Southern China, 2006–2012. PLoS One. 2014;9:e101976.
Lee HW, Cho P, Moon SU, Na BK, Kang YJ, Sohn Y, et al. Current situation of scrub typhus in South Korea from 2001–2013. Parasit Vectors. 2015;8:238.
Paris DH, Shelite TR, Day NP, Walker DH. Unresolved problems related to scrub typhus: a seriously neglected life-threatening disease. Am J Trop Med Hyg. 2013;89:301–7.
Chikeka I, Dumler JS. Neglected bacterial zoonoses. Clin Microbiol Infect. 2015;21:404–15.
Takamura A, Tanaka H, Kawamura A. Tsutsugamushi disease: an overview, Tanaka H. Tokyo, Japan: University of Tokyo Press; 1995.
Traub R, Wisseman Jr CL. The ecology of chigger-borne rickettsiosis (scrub typhus). J Med Entomol. 1974;11:237–303.
Ohashi N, Koyama Y, Urakami H, Fukuhara M, Tamura A, Kawamori F, et al. Demonstration of antigenic and genotypic variation in Orientia tsutsugamushi which were isolated in Japan, and their classification into type and subtype. Microbiol Immunol. 1996;40:627–38.
Nakayama K, Kurokawa K, Fukuhara M, Urakami H, Yamamoto S, Yamazaki K, et al. Genome comparison and phylogenetic analysis of Orientia tsutsugamushi strains. DNA Res. 2010;17:281–91.
Kuo CC, Huang JL, Ko CY, Lee PF, Wang HC. Spatial analysis of scrub typhus infection and its association with environmental and socioeconomic factors in Taiwan. Acta Trop. 2011;120:52–8.
Wardrop NA, Kuo CC, Wang HC, Clements AC, Lee PF, Atkinson PM. Bayesian spatial modelling and the significance of agricultural land use to scrub typhus infection in Taiwan. Geospat Health. 2013;8:229–39.
Kuo CC, Huang CL, Wang HC. Identification of potential hosts and vectors of scrub typhus and tick-borne spotted fever group rickettsiae in eastern Taiwan. Med Vet Entomol. 2011;25:169–77.
Kuo CC, Wang HC, Huang CL. Variation within and among host species in engorgement of larval trombiculid mites. Parasitology. 2011;138:344–53.
Kuo CC, Wang HC, Huang CL. The potential effect of exotic pacific rats Rattus exulans on vectors of scrub typhus. J Appl Ecol. 2011;48:192–8.
Olson J, Bourgeois A, Fang R. Population indices of chiggers (Leptotrombidium deliense) and incidence of scrub typhus in Chinese military personnel, Pescadores Islands of Taiwan, 1976–1977. Trans R Soc Trop Med Hyg. 1982;76:85–8.
Wang HC, Chung CL, Lin TH, Wang CH, Wu WJ. Studies on the vectors and pathogens of scrub typhus on murine-like animals in Kinmen County, Taiwan. Formosan Entomol. 2004;24:185-200. In Chinese, English Abstract.
Cooper W, Lien J, Hsu S, Chen W. Scrub typhus in the Pescadores Islands: an epidemiologic and clinical study. Am J Trop Med Hyg. 1964;13:833–8.
Lin TH. Ecological distribution of the scrub typhus vector in Taiwan. Chin J Entomol Spec Publ. 1999;12:171-178. (In Chinese, English Abstract).
Lin PR, Tsai HP, Weng MH, Lin HC, Chen KC, Kuo MD, et al. Field assessment of Orientia tsutsugamushi infection in small mammals and its association with the occurrence of human scrub typhus in Taiwan. Acta Trop. 2014;131:117–23.
Wang DQ, Yu ZZ. Chigger mites of the genus Leptotrombidium: key to species and their distribution in China. Med Vet Entomol. 1992;6:389–95.
Li J, Wang D, Chen X. Trombiculid mites of China: studies on vector and pathogen of tsutsugamushi disease. Guangzhou, China: Guangdong Science and Technology Publishing; 1997 [in Chinese].
Kuo CC, Huang JL, Shu PY, Lee PL, Kelt DA, Wang HC. Cascading effect of economic globalization on human risks of scrub typhus and tick-borne rickettsial diseases. Ecol Appl. 2012;22:1803–16.
Adler GH. Habitat relations within lowland grassland rodent communities in Taiwan. J Zool. 1995;237:563–76.
Jeng WS. Geographic variations and phylogeographic studies of Rattus losea and Rattus tanezumi in Taiwan. 2007. Master Thesis. National Chiayi University: Department of Biological Resources; 2007 (In Chinese, English Abstract).
Wu HY, Yu HT. Population dynamics of the spiny country rat (Niviventer coxingi) in a subtropical montane forest in central Taiwan. J Zool. 2000;250:339–46.
Yu HT. Distribution and abundance of small mammals along a subtropical elevational gradient in central Taiwan. J Zool. 1994;234:577–600.
Musser GG, Carleton MD. Superfamily Muroidea. Pp. 894–1531. In: Reeder DM, Wilson DE, editors. Mammal species of the world: a taxonomic and geographic reference. Baltimore: Johns Hopkins University Press; 2005.
Shih CD. Forest resource use by Asian house rat (Rattus tanezumi) in Orchid Island. Master thesis. National Dong Hwa University: Institute of Natural Resources Management; 2006. In Chinese, English Abstract.
Wu BH. An epidemiological study on scrub typhus in kindergarten children in Lanyu. Taiwan Epidemiol Bull. 1993;9:25–30 (In Chinese).
Ogawa M, Ono T. Epidemiological characteristics of tsutsugamushi disease in Oita prefecture, Japan: yearly and monthly occurrences of its infections and serotypes of its causative agent, Orientia tsutsugamushi, during 1984–2005. Microbiol Immunol. 2008;52:135–43.
Roh JY, Song BG, Park WI, Shin EH, Park C, Park M-Y, et al. Coincidence between geographical distribution of Leptotrombidium scutellare and scrub typhus Incidence in South Korea. PLoS One. 2014;9:e113193.
Peng PY, Guo XG, Ren TG, Song WY. Faunal analysis of chigger mites (Acari: Prostigmata) on small mammals in Yunnan province, southwest China. Parasitol Res. 2015;114:2815–33.
Lee IY, Kim HC, Lee YS, Seo JH, Lim JW, Yong TS, et al. Geographical distribution and relative abundance of vectors of scrub typhus in the Republic of Korea. Korean J Parasitol. 2009;47:381–6.
Acknowledgements
This study was financially supported by Taiwan Centers for Disease Control (DOH95-DC-2022, DOH96-DC-2020, DOH97-DC-2004, DOH98-DC-2012, DOH99-DC-2028).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
CCK and HCW designed the study and wrote the manuscript. PLL, CHC and HCW collected the data. HCW carried out the chigger identification. PLL and CHC participated in the immunoassays and molecular studies. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Kuo, CC., Lee, PL., Chen, CH. et al. Surveillance of potential hosts and vectors of scrub typhus in Taiwan. Parasites Vectors 8, 611 (2015). https://doi.org/10.1186/s13071-015-1221-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13071-015-1221-7