
Abstract
Background
Immune checkpoint inhibition and in particular anti-PD-1 immunotherapy have revolutionized the treatment of advanced melanoma. In this regard, higher tumoral PD-L1 protein (gene name: CD274) expression is associated with better clinical response and increased survival to anti-PD-1 therapy. Moreover, there is increasing evidence that tumor suppressor proteins are involved in immune regulation and are capable of modulating the expression of immune checkpoint proteins. Here, we determined the role of p53 protein (gene name: TP53) in the regulation of PD-L1 expression in melanoma.
Methods
We analyzed publicly available mRNA and protein expression data from the cancer genome/proteome atlas and performed immunohistochemistry on tumors with known TP53 status. Constitutive and IFN-ɣ-induced PD-L1 expression upon p53 knockdown in wildtype, TP53-mutated or JAK2-overexpressing melanoma cells or in cells, in which p53 was rendered transcriptionally inactive by CRISPR/Cas9, was determined by immunoblot or flow cytometry. Similarly, PD-L1 expression was investigated after overexpression of a transcriptionally-impaired p53 (L22Q, W23S) in TP53-wt or a TP53-knockout melanoma cell line. Immunoblot was applied to analyze the IFN-ɣ signaling pathway.
Results
For TP53-mutated tumors, an increased CD274 mRNA expression and a higher frequency of PD-L1 positivity was observed. Interestingly, positive correlations of IFNG mRNA and PD-L1 protein in both TP53-wt and -mutated samples and of p53 and PD-L1 protein suggest a non-transcriptional mode of action of p53. Indeed, cell line experiments revealed a diminished IFN-ɣ-induced PD-L1 expression upon p53 knockdown in both wildtype and TP53-mutated melanoma cells, which was not the case when p53 wildtype protein was rendered transcriptionally inactive or by ectopic expression of p53L22Q,W23S, a transcriptionally-impaired variant, in TP53-wt cells. Accordingly, expression of p53L22Q,W23S in a TP53-knockout melanoma cell line boosted IFN-ɣ-induced PD-L1 expression. The impaired PD-L1-inducibility after p53 knockdown was associated with a reduced JAK2 expression in the cells and was almost abrogated by JAK2 overexpression.
Conclusions
While having only a small impact on basal PD-L1 expression, both wildtype and mutated p53 play an important positive role for IFN-ɣ-induced PD-L1 expression in melanoma cells by supporting JAK2 expression. Future studies should address, whether p53 expression levels might influence response to anti-PD-1 immunotherapy.
Similar content being viewed by others

Background
Antibodies directed against the cytotoxic T-lymphocyte-associated antigen 4 (CTLA-4) receptor or programmed cell death-1 (PD-1) receptor have revolutionized the systemic therapy of advanced melanoma [1]. Indeed, with these immunotherapeutic approaches durable responses in the treatment of metastatic melanoma have been achieved for the first time [2,3,4]. In the first-line setting, response rates to CTLA-4 or PD-1 blockade range between 10 and 19% or 40–45%, respectively [5, 6]. Moreover, when both antibodies are combined, response rates increase up to 61% [5]. Since these therapies, especially in case of combination, may be accompanied by major, possibly even life-threatening side effects, much effort is being spent on identifying predictive biomarkers. So far, the most commonly used predictor of therapeutic response to PD-1 blockade is the expression of programmed-death-ligand 1 (PD-L1), a ligand of PD-1, on tumor cells. PD-L1 (also denoted as B7-H1) is encoded by the CD274 gene and is one of the two known ligands for PD-1, a costimulatory molecule that negatively regulates T-cell immune responses [7]. Notably, PD-L1 positivity (> 5% or > 1% of tumor cells positive for PD-L1 staining) is associated with a better overall response rate (ORR), progression free survival (PFS) and overall survival (OS) to anti-PD-1 immunotherapies [3, 4, 6, 8,9,10].
PD-L1 expression is inducible in many cell types, and increased expression has been observed in several tumor entities including melanoma, non-small cell lung cancer (NSCLC) and renal cell carcinoma. The interaction of cancer cell PD-L1 with PD-1 on cytotoxic T-lymphocytes (CTL) results in diminished T-cell killing [11, 12]. Possible mechanisms include suppressed T-cell proliferation and T-cell activation [13], induction of T-cell apoptosis [14] and also differentiation of CD4+ T-cells into FOXP3+ regulatory T-cells [15].
Various mechanisms have already been described that regulate PD-L1 expression in an often cell type-dependent manner [16]. Upregulation in tumor cells can be due to constitutively active oncogenic signaling pathways (referred to as innate immune resistance); although for melanoma cell lines PD-L1 expression levels were variable and independent from any driver mutation in MAPK or PI3K pathway [11, 17]. In addition, PD-L1 expression may occur in response to tumor-targeting immune cells that release interferon (IFN)-ɣ upon recognition of their cognate antigen expressed by cancer cells. PD-L1 expression on cancer cells subsequently inhibits PD-1-positive T-cells, a process known as adaptive immune resistance [11, 12]. IFN-ɣ signals through the canonical type II interferon receptor pathway [12, 18]. When IFN-ɣ binds to the IFN-ɣ receptor, JAK2 is activated by autophosphorylation of two tyrosine residues (Tyr 1007/Tyr 1008), and then transphosphorylates JAK1 on Tyr1022/Tyr1023. This leads to phosphorylation of tyrosine 440 in the IFN-ɣ receptor 1 by JAK1, which serves as recruitment site for STAT1 then allowing its phosphorylation on Y701 by most likely JAK2 [18, 19]. Subsequently, activated STAT1 dimers accumulate in the nucleus and act as transcription factors binding to the GAS (interferon-gamma activated site) elements of IFN-ɣ-inducible genes. The most important of these genes is interferon regulatory factor 1 (IRF1), which itself acts as a transcription factor during its ligation to IRF1-binding site-containing promoters like the PD-L1 promoter [18, 19]. Further transcriptional factors involved in PD-L1 regulation in melanoma include MYC, hypoxia-inducible factor-1α and 2α (HIF-1α/2α), STAT3 and NF-κB [16].
Post-transcriptionally, PD-L1 expression can be negatively regulated by various microRNAs (miRNAs, miR) such as miR-17-5p, miR-138-5p, miR-197, miR-200, miR-424, miR-513 and miR-570 [16, 20,21,22,23,24,25,26]. In addition, Cortez et al. recently demonstrated that p53 transcriptional activity leads to elevated miR-34a expression, which in turn reduced PD-L1 expression in NSCLC [27].
The main function of the tumor suppressor p53 is the regulation of cell proliferation and the induction of death in cells, which harbor genomic abnormalities [28, 29]. The molecular structure of p53 protein encompasses six domains (given residues are based on [30], but vary between studies): the transactivation domain (TAD) (residues 1–67), which can further be subdivided in two TADs; the proline-rich region (residues 68–98); the DNA-binding domain (DBD, residues 98–292); the hinge domain (HD, residues 293–325); the oligomerization domain (OD, residues 326–353); and the carboxy-terminal regulatory domain (CTD, residues 353–393). Most TP53 mutations occur in the DBD, and by impaired binding to target gene DNA its tumor suppressor ability is often reduced [28]. In addition of losing its tumor-suppressive properties, stabilized mutant p53 may gain novel functions (referred to as gain-of-function, GOF) [28, 29]. Those GOF are able to promote tumor progression or produce resistance to antitumor therapies.
Since in melanoma TP53 is frequently wildtype, we hypothesized that p53 might play a key role in repressing PD-L1 expression in melanoma and therefore investigated the role of p53 in PD-L1 regulation in melanoma.
Indeed, by conducting knockdown experiments of p53 in melanoma cell lines as well as immunohistochemistry of PD-L1 in melanoma tissue and analyzing the cancer genome atlas (TCGA) database, we found evidence for p53 being involved in the regulation of PD-L1 expression. Furthermore, we observed that induction of PD-L1 by IFN-ɣ is reduced after p53 knockdown. This is partly due to a reduction of JAK2, since ectopic JAK2 expression can largely rescue the effect of p53 knockdown on IFN-ɣ induced PD-L1 expression.
Material and methods
Cell lines, reagents and treatment regimens
We studied well-characterized melanoma cells from the NCI-60 panel that were TP53-wildtype (wt), i.e. LOX-IMVI, M19-MEL, MALME-3 M, SK-MEL-5, UACC-62, UACC-257, or TP53-mutated, i.e. M14, MDA-MB-435, SK-MEL-2 and SK-MEL-28. Furthermore, we used self-established TP53-mutated melanoma cell lines MS149 (p53 Q199*) and MS186 (p53 E154K). The two NCI-60 TP53-wt NSCLC cell lines A549 and H460 served as controls. All melanoma NCI-60 cell lines were obtained from ATCC. The 1205Lu melanoma cell line was originally obtained from the Wistar Institute, and the TP53-knockout variant was generated by Prof. Veit Hornung, Bonn/Munich, Germany, and kindly provided by co-author Markus Heppt, Munich.
Cell lines were cultured in RPMI-1640 with 10% fetal bovine serum, 100 U/ml penicillin and 0.1 mg/ml streptomycin (all from Sigma-Aldrich, Darmstadt, Germany) at 37 °C with 5% CO2. For IFN-ɣ stimulation, we used a concentration of 200 IU/ml (ImmunoTools, Friesoythe, Germany) for up to 48 h. Cell lines were regularly tested for mycoplasma contamination.
Lentiviral transduction and generation of small hairpin RNAs
For knockdown experiments we transduced cells with an antibiotic-selectable (zeocin or blasticidin), doxycycline (Dox)-inducible lentiviral small hairpin (sh) RNA vector. These vectors are based on a previously described system and the sequence is deposited (Accession number MH749464) [31]. The TP53 targeting sequences used were 5′-GAC TCC AGT GGT AAT CTA CT-3′ or 5′-CAC CAT CCA CTA CAA CTA CAT-3′ (in the confirmation experiments). A scrambled shRNA sequence (scr) served as control. Lentiviral transduction was performed as described before [32] and knockdown efficacy was determined by immunoblot. Cells containing doxycycline-inducible p53 or scr shRNA were incubated with doxycyline (1 μg/ml) for 6 days.
CRISPR/Cas mediated p53 inactivation
CRISPR/Cas technology was used to render wt p53 transcriptionally inactive by introducing deletions in the DBD/HD region. To achieve p53 inactivation, cells were transducted with a lentiviral system consisting of Dox-inducible Cas9 (pCW-Cas9 was a gift from Eric Lander & David Sabatini, Addgene plasmid # 50661) and lentiguide for gRNA expression (lentiGuide-Puro was a gift from Feng Zhang, Addgene plasmid # 52963). Used guides were 5′-CAT GTG TAA CAG TTC CTG CA-3′ (exon 7) and 5′-GTG AAA TAT TCT CCA TCC AG-3′ (exon 9) for LOX-IMVI and 5′-AGA TTA CCA CTA CTC AGG AT-3′ (exon 8) and 5′-GGA GAG GAG CTG GTG TTG TT-3′ (exon 9) for UACC-62, respectively. Exon 7 and 8 encode amino acids belonging to the DBD, while exon 9 contributes amino acids to the HD. These two cell lines contained a GFP based p53 reporter construct to measure p53 activity, as previously described [33].
Ectopic JAK2 and p53L22Q,W23S expression
JAK2 expression plasmid (pUNO1-hJAK2, Invivogen, San Diego, CA, USA) was kindly provided by Prof. Annette Paschen (Dept. of Dermatology, University Hospital Essen, Germany) and cloned into a pCDH-based lentiviral vector. Two melanoma cell lines (M19-MEL, UACC-62), already containing the zeocin-selectable, Dox-inducible p53 or scr shRNA, were transduced with this vector. JAK2 expression was confirmed by immunoblotting.
For inducible ectopic expression of an p53 variant severely compromised for transactivation we mutagenized a pCW-based vector encoding a Dox-inducible flag-HA-tagged TP53-wt at positions 22 and 23 to generate p53L22Q,W23S using the QuickChange Lightning Site-directed Mutagenesis kit (Agilent, Frankfurt, Germany) [34]. Two TP53-wt melanoma cell lines (M19-MEL, UACC-62) as well as a TP53-knockout melanoma cell line 1205Lu were transduced with this lentiviral vector.
Immunoblotting
Total cellular proteins were extracted at 4 °C using erythrocytes lysis buffer (ELB) containing protease inhibitors (Roche, Basel, Switzerland). Proteins (10–20 μg) were resolved on 8–12% SDS–polyacrylamide gels and transferred to Amersham™ Protran™ Premium 0.45 μm NC (GE Health Care Europe, Freiburg, Germany).
Immunoblots were probed with antibodies against PD-L1 (E1L3N), JAK1 (6G4), phospho-JAK1 (Tyr1022/1023; D7N4Z), JAK2 (D2E12), phospo-JAK2 (Tyr1008; D4A8), phospho-STAT1 (Tyr701; 58D6; all by Cell Signaling Technology, Boston, MA, USA), STAT1 (10C4B40), phospho-STAT1 (S727; A15158B), IRF-1 (13H3A44; all by BioLegend, San Diego, CA, USA), monoclonal p53 (DO-1; Santa Cruz Biotechnology, Dallas, TX, USA) or polyclonal p53 (#9282; Cell Signaling Technology). ß-tubulin (TUB2.1) or ß-actin (AC-15; both Sigma-Aldrich) served as loading control.
Flow cytometry
Adherent cells were detached using 0.02% ethylenediaminetetraacetic acid (EDTA) in phosphate buffered saline (PBS). After washing them with 0.1% bovine serum albumin (BSA; all from Sigma-Aldrich) in PBS, cells were incubated with PD-L1 APC antibody (29E.2A3, 1:20; BioLegend) or with HLA-ABC (MHC class I) APC antibody (W6/32; 1:50; ImmunoTools) for 20 min on ice. After washing twice with 0.1% BSA in PBS cells were analyzed with the BD FACS Canto. Non-viable cells were excluded using 7-Amino Actinomycin D (7-AAD; BD Biosciences, Franklin Lakes, NJ, USA).
Immunohistochemistry
All analyzed samples were collected from patients who received treatment at the Department of Dermatology at the University Hospital Würzburg between November 2014 and July 2016. Written informed consent was obtained from each patient.
TP53-mutation status was determined by next generation sequencing (for detailed information on library preparation and sequencing, please refer to Appenzeller et al. [35]).
Four μm-sections of paraffin-embedded primary and metastatic tumors were dried at 75 °C for 20 min and then treated twice with xylol for 5 min at room temperature. Subsequently, sections were washed twice with absolute ethanol and once with 70% ethanol followed by one rinse with bi-distilled water. For antigen retrieval, sections were incubated with Tris/EDTA-buffer pH 9.0 for 40 min at 90 °C and then cooled down for 20 min. After washing in TBS buffer for 5 min, slides were incubated with monoclonal antibody against PD-L1 (E1L3N, CST, 1:200) for 40 min. Washing in TBS buffer for 5 min was followed by incubation with secondary antibodies (REAL Biotinylated Secondary Antibodies (AB2), Dako) for 30 min. Samples were briefly washed with TBS buffer again, then incubated in peroxidase-blocking solution for 5 min, streptavidin peroxidase for 25 min, CHROM AEC/H2O2 substrate solution for 15 min, automation hematoxylin histological staining reagent (all from Dako) for 5 min, and cleansed with bi-distilled water. Between every incubation step, slides were washed with TBS buffer.
Analysis of the cancer genome atlas (TCGA)
TCGA data (http://cancergenome.nih.gov/) were retrieved and analyzed by software R with different packages [36]. In this regard, RNAseq and miRNA data for the cases of patients diagnosed with cutaneous melanoma, were downloaded and prepared with the package “TCGAbiolinks” [37]. Mutational data of TP53 (i.e. missense, truncated, frameshift, splice mutations and homozygous deletions) was downloaded from cBioPortal and prepared by the package “maftools” [38]. Reverse phase protein array (RPPA) was obtained from the cancer proteome atlas (TCPA) [39]. For mRNA expression the transcripts per million (TPM) values, for miRNA the reads per million (RPM) and for the RPPA data the level 4 data generated by replicate-based method processing was extracted. Each data set was log2-transformed after adding the value 1 for TPM and RPM and 5.99 for RPPA. The mRNA and miRNA data were available for 447, the mutational data for 347 and the RPPA for 354 individual cases. All data were available for a cohort of 262 cases. For correlation analysis, the mRNA and miRNA datasets extracted from the RNAseq data available from the TCGA database was first restricted to those genes, which had at least 1 TPM (for correlation with genes) or at least 1 RPM (for correlation with miRNA) in 20% of the 447 cases using the R-package edgeR [40]. Subsequently, CD274 mRNA expression was correlated to either the expression of the remaining genes or miRNA using the built in spearman correlation and loop function of R.
Real-time PCR for quantification of TP53, CD274 and JAK2 mRNA expression
mRNA isolation, cDNA transcription and RT-qPCR with RPLP0 as endogeneous control was performed as described before [41]. Cells were treated for 6 days with doxycycline in the absence or presence of IFN-ɣ for the last 2 days. Primers used to detect the expression of the respective gene of interest by SYBR green assay were the following: TP53_fw: GAG GTT GGC TCT GAC TGT ACC; TP53_rv: TCC GTC CCA GTA GAT TAC CAC; JAK2_fw: CAG GCA ACA GGA ACA AGA TG; JAK2_rv: CCA TTC CCA TGC AGA GTC TT; CD274_fw: CAT CTT ATT ATG CCT TGG TGT AGC A; CD274_rv: GGA TTA CGT CTC CTC CAA ATG TG; RPLP0_fw: CCA TCA GCA CCA CAG CCT TC; RPLP0_rv: GGC GAC CTG GAA GTC CAA CT. Relative expression was calculated by the ΔΔCq method with the cells treated without doxycycline and IFN-ɣ serving as calibrator [42].
Statistics
Statistical analysis was performed with R. Graphs were prepared with “ggplot2” [43]. Expression data between groups were compared by the Wilcoxon–Mann–Whitney test. Relations between two genes were calculated by linear regression. Correlations of genes, miRNA and protein expressions was calculated by Spearman correlation on filtered data (> 1 TPM or > 0 RPM, respectively, in at least 20% of cases). The factors with the best correlation estimate values were then depicted as heatmap generated with the package “ComplexHeatmap” [44] using Spearman as clustering distance and ward. D as clustering method for the factors. A p value < 0.05 was regarded significant. Gene enrichment analysis was performed using PANTHER overrepresentation test (version 13.1, released 2018.02.03) and Fisher test statististics [45, 46]. FCM data and relative mRNA expression (log2 transformed) were depicted and analyzed by Prism 7 (Graphpad) using a paired T-test.
Results
TP53-mutated melanoma present higher CD274 mRNA expression levels
To allow for an easier discrimination between mRNA and protein in our manuscript, we refer to the official gene names for mRNA (TP53 and CD274) and to the widely-used molecule names p53 and PD-L1 (CD274) for protein expression.
Cortez et al. [27] analyzed NSCLC data from TCGA and reported significantly higher CD274 mRNA levels in TP53-mutated NSCLC than in wt counterparts. Furthermore, they revealed a statistically inverse correlation between TP53 and CD274 mRNA for the total cohort of lung adenocarcinoma cases. To test a possible correlation between TP53 and CD274 in melanoma, we analyzed TCGA skin cutaneous melanoma (SKCM) data. To this end, also for melanoma CD274 mRNA levels were significantly higher in TP53-mutated than in TP53-wt samples (Fig. 1a; p = 0.0181; Mann-Whitney).

TP53-mutated melanoma show higher CD274 mRNA expression levels, and are immunohistochemically more often PD-L1 positive. a Analysis of TCGA melanoma samples (n = 347) for differences in CD274 mRNA expression. Wilcoxon-Mann-Whitney test was used to compare CD274 expression between TP53-wt and -mutant samples. b PD-L1 immunohistochemistry of primary and metastatic melanoma samples (n = 81). Next-generation sequencing of the TP53 gene was performed previously. Samples harboring more than > 1% PD-L1 positive melanoma cells were regarded as PD-L1-positive. A PD-L1 negative, and a positive metastasis are depicted. Magnification, each 400x (c, d) Samples (n = 262) were stratified by TP53-mutation status and publicly available PD-L1 expression determined by reverse phase protein array was compared with the Wilcoxon-Mann-Whitney test (c) and by linear regression association analysis with p53 (d). mRNA and protein data are presented on a logarithmic scale. p < 0.05 is regarded as statistically significant. TPM, transcripts per million; wt, wildtype; mut, mutation
In a next step, we performed immunohistochemistry for PD-L1 on 81 primary and metastatic melanoma samples with known TP53-mutation status, which has been determined by next generation sequencing before [34]. Regarding PD-L1 staining, it is common practice to use a threshold of either 1% or 5% of stained tumor cells to classify tumor samples [3, 4, 6, 8, 9]. Here we scored samples with > 1% stained tumor cells as PD-L1-positive. Noteworthy, 4/7 (57%) of TP53-mutated samples but only 14/74 (19%) of TP53-wt samples (p = 0.0401) were PD-L1-positive (Fig. 1b; for clinical information on this patient cohort see Additional file 1: Figure S1a).
In order to expand our investigation on PD-L1 expression in melanoma, we analyzed data obtained from the cancer proteome atlas (TCPA) project, which collects results from reverse phase protein array (RPPA) for different entities including melanoma [39]. These data did not reveal a statistically significant difference in overall PD-L1 expression levels between TP53-wt or -mutated melanoma samples (Fig. 1c). We did, however, detect a positive correlation between PD-L1 and p53 protein expression, which reached statistically significance only for the TP53-wt cohort (p = 4 × 10− 13; R2 = 0.211; Fig. 1d).
miR-34a is not the key regulator of PD-L1 in TCGA skin cutaneous melanoma
For NSCLC, p53-driven miR-34a expression was demonstrated to be a key regulator of PD-L1 expression [27]. Since at least at the mRNA level the observed dependency of CD274 on the TP53-mutational status was similar for melanoma as for NSCLC, we analyzed the melanoma TCGA data for miRNA and CD274 expression focusing first on miR-34a. Again, similar as for NSCLC, miR-34a expression was significantly higher in TP53-wt than in TP53-mutated tumors (p = 0.0181; Additional file 2: Figure S2a), even with a small, but significant negative association of TP53 mRNA and miR-34a expression (p = 0.00057; R2 = 0.0395; data not shown). However, no correlations between miR-34a and CD274 mRNA (data not shown) nor between miR-34a and PD-L1 protein (Additional file 2: Figure S2b) were detected. Accordingly, neither miR-34a nor most of the other known miR negatively affecting PD-L1 were among the top 24 miRs correlating with CD274 mRNA expression (Additional file 2: Figure S2c). The best correlation was observed for CD274 mRNA and miR-7702 (Additional file 2: Figure S2d; p = 2 × 10− 16; R2 = 0.353), whose expression, however, was not significantly different between TP53-wt and -mutated tumors (Additional file 2: Figure S2e). Indeed, a positive correlation between CD274 mRNA and miR-7702 was evident in both TP53 genotypes (Additional file 2: Figure S2f).
Members of immune-related pathways are enriched among the genes correlating best with CD274 mRNA expression in TCGA dataset
So far, our results revealed an increased CD274 mRNA expression in the TCGA data and a higher percentage of PD-L1 positivity in TP53-mutated tumors in our cohort of 81 samples, while overall PD-L1 expression levels were not significantly different. We thus analyzed 6 TP53-wt and 6 TP53-mutated melanoma cell lines for PD-L1 expression. Our immunoblot analysis revealed that - as expected [47] - p53 expression levels were mostly higher in mutated cell lines, but no consistent difference in basal PD-L1 expression was evident when stratified by TP53 status (Fig. 2a).

CD274 expression correlates with genes of immune-related pathways while PD-L1 levels lack correlation with p53’s transcriptional activity. a Immunoblot for PD-L1 and p53 expression of each six untreated TP53-wildtype and -mutated melanoma cell lines. Before lysis, cells were cultured for at least three days. Blot is representative of two individual experiments. b Correlation of CD274 with other genes was calculated using Spearman correlation (n = 447). Only genes with > 1 TPM in a least 20% of cases were included. The 24 best correlating genes are presented in a “heatmap”. Expression values are presented in a spectrum of blue (small) to red (high). c Gene enrichment analysis of the 24 best correlating genes with PANTHER overrepresentation test and Fisher test statistics depicting overrepresented pathways. Total number (#) of genes ascribed to the respective pathway, number of genes from the top 24 genes belonging to the pathway, expected frequency, fold enrichment, overpresentation indicated by “+”, raw p-value and false discovery rate (FDR) are given. p < 0.05 is regarded as statistically significant. TPM, transcripts per million
Hence, to identify factors, which modulate CD274 mRNA expression, we searched for the 24 genes with the best correlation to CD274 mRNA expression (Fig. 2b). In order to determine pathways that were overrepresented among these 24 genes, we analyzed them with PANTHER [45, 46]. This analysis demonstrated that the only pathways overrepresented by the genes are immune response-related (Fig. 2c), indicating that an ongoing immune response with IFN-ɣ secretion and activation of the JAK/STAT pathway might impact PD-L1 expression.
These observations prompted us to perform in vitro experiments in order to determine the role of p53 in the regulation of PD-L1 expression.
p53 knockdown increases basal, but negatively affects IFN-ɣ-induced PD-L1 expression in melanoma cells
In order to directly test the impact of p53 on PD-L1 expression, we generated several cell lines with inducible shRNA targeting TP53. Two TP53-wt NSCLC cell lines, i.e. A549 and H460, which in another study demonstrated increased PD-L1 expression upon p53 knockdown or miR-34a transfection, served as positive controls [27]. Indeed, H460 cells displayed a slightly increased expression of PD-L1 upon p53 knockdown while expression levels were unaffected in scr control cells (Fig. 3a). Among the 6 p53 wt melanoma cells tested (LOX-IMVI, M19-MEL, MALME-3 M, SK-MEL-5, UACC-62, UACC-257), a modest increase of PD-L1 upon p53 knockdown was only evident for LOX-IMVI and MALME-3 M. For SK-MEL-5 there was no difference, and because for all others melanoma lines basal PD-L1 expression levels were below the detection limit of the immunoblot, we analyzed PD-L1 expression also by flow cytometry. This analysis demonstrated that in all analyzed cell lines a slight increase of PD-L1 expression was measurable upon p53 knockdown which, however, was not significant in any cell line (Additional file 3: Figure S3a).

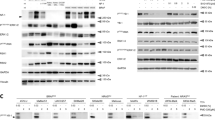
Presence of p53 protein, but not its transcriptional activity determines extent of IFN-ɣ induced PD-L1 expression in melanoma. a Immunoblot for PD-L1 and p53. NSCLC cell lines A549 and H460 served as control; all other are melanoma cell lines. IFN-ɣ treatment was for 48 h. b p53 was rendered transcriptionally inactive by introducing deletions using CRISPR/Cas9 technology and gRNA targeting exons 7 and 9 (LOX-IMVI) or exons 8 and 9 (UACC-62), respectively. Loss of transcriptional activity was determined by expression of GFP-based p53 reporter (left histograms; red: parental cells; blue: cells after CRISPR/Cas9 genome editing). Protein expression of p53 and PD-L1 in the absence or presence of IFN-ɣ for 48 h was determined by immunoblot. c Immunoblot for PD-L1 and p53 in TP53-mutated melanoma cells upon shRNA-mediated p53 knockdown. Cells were treated with IFN-ɣ as their TP53-wildtyp counterparts described in (a). p53 knockdown was achieved by culturing cells in doxycycline for 6 days. d, e Two TP53-wt melanoma cell lines (d) or a TP53-knockout cell line (e) were transduced with doxycycline-inducible p53L22Q,W23S expression construct. Cells were incubated with doxycycline and treated with IFN-ɣ for 48 h, as described before. Expression of indicated proteins was determined by immunoblot. Arrow (d) indicates ectopic p53 expression. Please note, that for M19-MEL (d) the ectopic p53 expression was so much stronger than the endogenous that, in those samples without doxycycline, the signal for endogenous p53 was too low to be detected. ß-tubulin (a-d) or actin (e) served as loading controls. DOX, doxycycline. All blots are representative of two individual experiments
It is known from the literature as well as apparent from our TCGA data analysis that PD-L1 expression is modified by immune responses leading to IFN-ɣ secretion [11, 12]. We therefore analyzed the effect of p53 knockdown on IFN-ɣ-induced PD-L1 expression. To this end, after culture for 4 days in the absence or presence of doxycycline to induce knockdown of p53, cells were additionally treated for 48 h with IFN-ɣ, and PD-L1 expression was determined by immunoblot or flow cytometry. As expected, IFN-ɣ did increase PD-L1 expression in all cell lines (Fig. 3a, Additional file 3: Figure S3a). This increase was much more pronounced than the increases observed upon p53 knockdown. Only for SK-MEL-5 the increase upon IFN-ɣ treatment was marginal. In the two NSCLC cell lines, IFN-ɣ-induced PD-L1 upregulation was similar or slightly increased upon additional p53 knockdown (Fig. 3a, Additional file 3: Figure S3a). Surprisingly, however, in the five melanoma cell lines (LOX-IMVI, M19-MEL, MALME-3 M, UACC-62, UACC-257) demonstrating a distinct IFN-ɣ-induced PD-L1 upregulation in our immunoblot analyses, a reduction of p53 protein levels was accompanied by a decreased IFN-ɣ-inducible PD-L1 expression. Indeed, the most prominent decrease of PD-L1-inducibilty was observed in those cell lines that presented with the strongest PD-L1 induction upon IFN-ɣ treatment (M19-MEL, UACC-62 and UACC-257, Fig. 3a, Additional file 3: Figure S3a).
Since shRNA might have off-target effects, we repeated these experiments in four melanoma cell lines with a second TP53-targeting shRNA. Although knockdown efficiency of this shRNA was not as good, we could still confirm our observation that upon p53 knockdown IFN-ɣ treatment was less effective in inducing PD-L1 expression (Additional file 3: Figure S3b).
Altogether, p53 knockdown in TP53-wildtype melanoma cells resulted in only a very modest increase of basal PD-L1 expression, but clearly reduced IFN-ɣ induced expression.
The extent of IFN-ɣ-inducible PD-L1 expression is dependent on the presence of p53 protein, but not on its transcriptional activity
Our analyses of publicly available data had revealed that there is a positive correlation between PD-L1 and p53 expression. Further analyses demonstrated that IFNG mRNA levels were not different between tumors with TP53-wt or TP53-mutant status (Additional file 4: Figure S4a). Moreover, while IFNG levels did not correlate with p53 expression, there was a positive correlation with PD-L1 expression for both TP53 genotypes (Additional file 4: Figure S4b, c). Taken together, these observations suggest that while p53 presence augments IFN-ɣ-induced PD-L1 expression, this might not dependent on its transcriptional activity. In order to test this hypothesis, we rendered p53 transcriptionally inactive in originally TP53-wt melanoma cells by introducing deletions in the DBD/HD using CRISPR/Cas9 technology. These two generated melanoma cell lines expressed truncated p53 which resulted in diminished p53 reporter activity. The extent of PD-L1 induction following IFN-ɣ treatment was, however, not affected (Fig. 3b).
Furthermore, we analyzed whether the effect observed for p53 knockdown in the wildtype TP53 melanoma cell lines could be reproduced in three TP53-mutant cell lines (M14, SK-MEL-2, SK-MEL-28). Similar to TP53-wt melanoma cell lines, the reduction of p53 protein in these cells led to an impaired induction of PD-L1 by IFN-ɣ (Fig. 3c).
The ability to activate gene transcription is among the best-characterized properties of p53. This function is ascribed to the two transactivation domains (TADs), and introducing mutations at the amino acids 22 and 23 generates a p53 protein (p53L22Q,W23S) with clearly reduced transactivation-potential [34]. Hence, to further evaluate the role of transcriptional activity for IFN-ɣ-induced PD-L1 expression, we transduced TP53-wt melanoma cell lines or a TP53-knockout cell line with a vector allowing inducible expression of p53L22Q,W23S. When expressed in TP53-wt melanoma cell lines, PD-L1 inducibility by IFN-ɣ was hardly affected (Fig. 3d). Importantly, however, expression in a TP53-knockout melanoma cell line was already associated with an increased basal PD-L1 expression, and boosted PD-L1 expression upon IFN-ɣ stimulation (Fig. 3e). Thus, even when transactivation activity is impaired, the presence of p53 can augment IFN-ɣ-induced PD-L1 expression.
p53 knockdown leads to a reduction of JAK2, which is associated with a delayed JAK2 and a diminished STAT1 phosphorylation by IFN-ɣ
As mentioned before, IFN-ɣ signals through the JAK-STAT-IRF1 axis to regulate PD-L1 [18]. Notably, among the top 24 genes whose mRNA correlated with CD274 mRNA in the TCGA SKCM data set, the best correlation was for STAT1 (p = 2 × 10− 16; R2 = 0.584, Figs. 2c, 4a), which translated also in a positive correlation of STAT1 mRNA and PD-L1 (p = 2 × 10− 16; R2 = 0.271, Fig. 4b). Importantly, STAT1 activation by genotoxic agents has been demonstrated to depend on p53 protein but not on its transcriptional activity [48]. In this regard, STAT1 possesses two phosphorylation sites (Y701 and S727), both of which are functionally important for efficient signaling through the type II interferon receptor pathway [19]. STAT1 Y701 phosphorylation depends directly on activated JAK1/2, whereas STAT1 S727 phosphorylation is fundamental for maximal ability to activate transcription of target genes, and can be modulated by different cellular influences [19, 49]. We first examined the effect of short term IFN-ɣ treatment, i.e. after 5, 15, 30 or 60 min, on the JAK/STAT pathway in two melanoma cell lines. These analyses revealed that total JAK2 and to a lesser extent STAT1 were reduced in p53 knockdown cells. Shortly after addition of IFN-ɣ, JAK2 became phosphorylated at Y1008 and STAT1 at Y701 while the phosphorylation of STAT1 at S727 was unchanged. In cells with reduced p53 levels, absolute JAK2 Y1008 phosphorylation levels were at least at 5 and 15 min lower; although the ratio of phosphorylated to total JAK2 was even higher due to decreased total JAK2 levels. Similarly, although more of STAT1 is phosphorylated at Y701, the total amount is slightly reduced due to accompanied decreased total-STAT1 levels (Fig. 4c). Of note, treatment of cells with IFN-ɣ for 48 h generally led to a decrease of JAK2 in melanoma cells, which was even more evident in cells with p53 knockdown (Fig. 5a). At this time point, STAT1 S727 phosphorylation was reduced in p53 knockdown cells.

Correlation of STAT1 and CD274/PD-L1 in melanoma, and disturbed IFN-ɣ signaling upon p53 knockdown in melanoma cell lines. a, b Linear regression analyses of STAT1 mRNA with CD274 mRNA (n = 347) (a) or PD-L1 protein (n = 262) (b). c LOX-IMVI and M19-MEL were incubated in six well plates for 6 days with doxycycline for p53 knockdown and subsequently treated with IFN-ɣ for 5, 15, 30 or 60 min. Effect of treatment on the IFN-ɣ signaling pathway was analyzed by immunoblot with the indicated antibodies. ß-tubulin served as a loading control. p < 0.05 is regarded as statistically significant. DOX, doxycycline; TPM, transcripts per million

Restoration of p53 knockdown-associated JAK2 downregulation improves IFN-ɣ-inducible PD-L1 expression. a Immunoblot of three melanoma cell lines containing either an inducible TP53-targeting or a scr shRNA expression vector. IFN-ɣ treatment was for 48 h. b, c Linear regression analyses of JAK2 mRNA with CD274 mRNA (b; n = 347) or PD-L1 protein (c; n = 262). d M19-MEL and UACC-62 cells containing an inducible TP53-targeting shRNA vector were treated for 6 days with doxycycline with the last 2 days either in absence or presence of IFN-ɣ. After RNA isolation and cDNA generation, real-time quantitative PCR was performed for determination of TP53, JAK2 and CD274 mRNA expression. Relative expressions were calculated by the ΔΔCq method to the respective cell line sample without doxycycline and IFN-ɣ treatment. After log2 transformation, the means + SD of three independent experiments are depicted. Significant differences are indicated by stars (* < 0.05; ** < 0.01; paired t-test). e Two melanoma cell lines containing Doxycycline-inducible p53 shRNA were transduced with a JAK2 expression construct. Control cells and JAK2-overexpressing cells were incubated with doxycycline and treated with IFN-ɣ for 48 h as described before. Expression of indicated proteins was determined by immunoblot. ß-tubulin served as a loading control. p < 0.05 is regarded as statistically significant. DOX, doxycycline; scr, scramble; TPM, transcripts per million, ctrl, control. All blots are representative of two individual experiments
Ectopic JAK2 can largely restore reduced IFN-ɣ-inducible PD-L1 expression after p53 knockdown
Besides a reduced JAK2 expression upon p53 knockdown, we also detected within the TCGA-SKCM data a positive correlation between CD274 mRNA and JAK2 mRNA (p = 2.2 × 10− 16; R2 = 0.459) as well as between PD-L1 and JAK2 mRNA (p = 7.6 × 10− 10; R2 = 0.136; Figs. 5b and c). To test, if reduction of IFN-ɣ-induced JAK2 and PD-L1 protein levels after p53 knockdown is accompanied with a decrease of corresponding mRNAs, we performed real-time PCR. These analyses revealed that p53 knockdown had no dramatic impact on basal JAK2 or CD274 mRNA expression levels in the 2 melanoma cell lines M19-MEL and UACC-62. Upon stimulation with IFN-ɣ, cells demonstrated enhanced JAK2 and CD274 mRNA expression. When combined with p53 knockdown, these increases were significantly reduced (Fig. 5d).
Based on our observations, we hypothesized that reduced JAK2 levels upon p53 knockdown are at least partially responsible for the decreased IFN-ɣ-induced PD-L1 expression. Therefore, we analyzed p53 knockdown in two melanoma cell lines ectopically expressing JAK2. In those cells, JAK2 levels were higher than endogenous levels in control cells. While IFN-ɣ-induced expression of interferon regulatory factor 1 (IRF-1) was reduced by p53 downregulation in control cells, ectopic expression of JAK2 led to levels similar to that in control cells without p53 downregulation. Importantly, JAK2 overexpression largely restored IFN-ɣ-induced PD-L1 expression in p53 knockdown cells (Fig. 5e).
Discussion
p53 is a central tumor suppressor protein, which is stabilized and activated following different cellular stresses including DNA damage and replication stress provoked by deregulated oncogenes [29]. Once activated this transcription factor can promote cell cycle arrest, DNA repair or apoptosis. Importantly, the specific p53 triggered response depends on the cellular context, which includes cell type, epigenetic state, tissue microenvironment and activating signals [29, 50]. In particular, it has been proposed that melanocytes (and accordingly melanocyte-derived tumor cells) may respond differently to p53 activation because these cells are adopted to survive even with p53 induction by highly mutagenic UV light and by the oxidative stress of melanin production [51].
Besides the response to DNA damage, p53 controls many further distinct processes and plays e.g. an important role in inflammation and immune responses [52]. Indeed, p53 is directly involved in the upregulation of antigen presentation via the major histocompatibility complex (MHC) I pathway [53]. Reported mechanisms include induction of members of the antigen processing machinery like TAP1 or ERAP1 by wildtype p53 [54, 55]. As a result, wildtype p53 can improve MHC class I expression and thereby promote tumor cell killing by cytotoxic T-lymphocytes (CTL) [53,54,55]. Therefore, besides several other aspects of p53 biology, also immune modulation may contribute to the impact that TP53 mutation status has on prognosis and even more on immunotherapy response. Indeed, TP53 mutation has been demonstrated to be associated with poorer outcome in melanoma patients receiving anti-CTLA-4 treatment [56]. In another study with melanoma patients treated by different immune checkpoint blockade therapies, TP53 mutation was one of the factors associated with inferior outcome [57]. In melanoma, the best known predictor for response to anti-PD-1 therapy is the expression of PD-L1 on tumor cells [3, 4, 6, 8,9,10]. In this regard, in NSCLC it has been demonstrated that p53 downregulates PD-L1 via miR-34a and thereby enhances CTL activity [27]. We thus analyzed publically available mRNA and protein expression data as well as own samples to investigate whether TP53 might influence PD-L1 expression in melanoma. For CD274 mRNA we obtained similar results to those described for NSCLC, with a higher expression level for TP53-mutated tumors and an inverse correlation between TP53 and CD274 mRNA for TP53 wt melanoma tumors (Additional file 1: Figure S1b). This correlation, however, was quite weak, and the underlying mechanism of regulation appears to be different from that reported for NSCLC, since we could not observe a correlation for miR-34a and CD274 mRNA indicating that specific p53 activity is context-dependent [29, 50]. Moreover, we detected a positive correlation of p53 and PD-L1 at the protein level. When stratified by TP53 status, PD-L1 expression levels were not different although p53 expression was higher in TP53-mutated tumors (data not shown). Since RPPA protein expression data cannot distinguish between expression on tumor or stroma cells, we analyzed a series of melanoma samples with known TP53 status by immunohistochemistry revealing that PD-L1 tumor cell positivity was more frequent in TP53-mutated tumors. This observation certainly has to be confirmed in a larger cohort. Nevertheless, this may match the situation of primary pulmonary lymphoepithelioma-like carcinoma, where immunohistochemically p53-positive samples, which - based on their staining pattern - were regarded as TP53-mutated, and were also significantly more often PD-L1-positive [58]. In contrast, the TP53 mutation status did not correlate with PD-L1 expression in colon cancer, suggesting again diverging mechanisms of PD-L1 regulation in different tumor types [59]. Given that our immunoblot results did not reveal a clear correlation between TP53-mutation status and PD-L1 baseline expression in melanoma cell lines (Fig. 2a), and p53 knockdown only slightly influenced PD-L1 baseline expression, it is unlikely that p53 has a major intrinsic effect on PD-L1 expression in melanoma.
Accordingly, TCGA data analyses demonstrated that, among the mRNAs best correlating with CD274 expression, only immune response-related genes were enriched. This indicates that an ongoing immune response may determine PD-L1 expression. Indeed, immunohistochemistry of many cancers revealed PD-L1 expression typically in T-cell–rich areas of tumors, particularly at the invasive margin, illustrating IFN-ɣ-inducible PD-L1 expression to be more common than constitutive expression [12, 60, 61].
This in vivo observation can be reenacted by exposing tumor cells to IFN-ɣ, thereby leading to a marked increase of PD-L1 expression (Knol et al. [62] and Fig. 3a). Surprisingly, this increase was diminished upon p53 knockdown in melanoma cells. Importantly, decreased IFN-ɣ-induced PD-L1 expression upon p53 knockdown was also evident in TP53-mutated melanoma cell lines, while rendering p53 inactive by CRISPR/Cas9 had no effect on PD-L1 inducibility. Furthermore, a histopathological study of desmoplastic melanoma, which frequently harbor TP53 mutations, identified a significant positive correlation between PD-L1 and p53 expression [63, 64]. Hence, presence of p53 but not its transcriptional activity appears to be required for full IFN-ɣ-induced PD-L1 expression in melanoma. Consistently, expression of p53L22Q,W23S, a p53 protein with impaired transactivation activity, in the 1205Lu TP53-knockout− melanoma cell line led to a prominent increase of IFN-ɣ-induced PD-L1 expression. In this context, it certainly would also be interesting to analyze the impact of p53 with GOF mutations on IFN-ɣ-induced PD-L1 expression, an aspect we did not yet address.
The many functions of p53 can be divided into transcription-dependent and -independent activities. Indeed, besides in the nucleus p53 can act also in the cytosol or at the mitochondria [65]. Regarding apoptosis promotion, it has been demonstrated that the pro-apoptotic protein Bax can be activated by certain transcription-deficient mutant p53 proteins [66]. As another example, genotoxic drugs can induce STAT1 activation, a process that depends on p53 protein but not on its transcriptional activity. This has been demonstrated by restoring drug-induced STAT1 Y701 phosphorylation by expression of transcriptional-inactive p53 mutants in p53-null cell lines [48]. Of note, it has previously been demonstrated that genotoxic stress-induced upregulation of PD-L1 is also p53-dependent in a breast carcinoma cell line [67].
In our experiments, reduced total-STAT1, but also STAT1 S727 phosphorylation was evident after 48 h of IFN-ɣ-stimulation in p53 knockdown cells. Importantly, phosphorylation of this STAT1 site is induced by various stimuli (e.g. LPS, PDGF) besides IFN-ɣ signaling, and is essential for maximal transcription of target genes [19, 49]. Therefore, decreased STAT1 S727 phosphorylation through molecules beyond JAK-STAT-signaling, could have contributed to the diminished IFN-ɣ induced PD-L1 expression after p53 knockdown.
Furthermore, it has been shown that inducible but not constitutive PD-L1 expression depends on NF-κB activation in melanoma cells [68]. Interestingly, while NF-κB and p53 often have opposing effects in cancer cells, in human monocytes and macrophages both co-regulate induction of pro-inflammatory genes [69].
Our real-time PCR results indicate that IFN-ɣ-induced upregulation of JAK2 and PD-L1 is already affected at mRNA level (Fig. 5d). This sustains our hypothesis that p53 knockdown impairs IFN-ɣ-induced CD274 transcription mainly through interference with the JAK-STAT signaling pathway. Consistently, we revealed that p53 knockdown was associated with a reduction of JAK2 protein levels (Figs. 4c, 5a and d). Luo et al. recently demonstrated that JAK2 knockdown in various melanoma cell lines only diminished PD-L1, but did not substantially change IFN-ɣ-induced MHC-I expression [70]. In contrast, in an NSCLC cancer cell line p53 cooperated with IFN-ɣ to enhance the expression of surface MHC-I [54]. Naturally, we also analyzed IFN-ɣ induced MHC-I surface expression upon p53 knockdown and noted that induction was only slightly affected (Additional file 3: Figure S3c). This observation is in accordance with the results of Luo et al. and again indicates differences between melanoma and NSCLC [70].
Conclusions
In summary, there is a tendency of higher PD-L1 expression in TP53-mutated melanoma cells. One contributing factor might be an increased p53 expression level in these tumors. Indeed, while our analyses demonstrate that p53 has only a minor influence on constitutive PD-L1 expression, its presence is important for IFN-ɣ-induced PD-L1 expression through the JAK-STAT-signaling pathway in melanoma cell lines. Moreover, it does not seem to be important whether p53 is transcriptionally active. Interestingly, short-term JAK2 inhibition in a preclinical melanoma model did not affect immunotherapeutic responses, while for melanoma patients inactivating JAK2 mutations have already been associated with a diminished response to anti-PD-1 directed immunotherapy [70,71,72]. Accordingly, future studies should analyze, whether p53 expression levels correlate with JAK2 expression in melanoma, and how they affect response to anti-PD-1-based immunotherapy.
Availability of data and materials
The data presented is partially obtained by analyzing already publicly available data. All other original data generated or analyzed during this study are included in the manuscript or can be accessed through its supplementary files.
Abbreviations
- CD274 :
-
gene coding for PD-L1 protein
- Dox:
-
Doxycycline
- mut:
-
mutated
- RPM:
-
Reads per million
- TP53 :
-
gene coding for p53 protein
- TPM:
-
Transcripts per millions
- wt:
-
wildtype
References
Redman JM, Gibney GT, Atkins MB. Advances in immunotherapy for melanoma. BMC Med. 2016;14:20.
Robert C, Thomas L, Bondarenko I, O’Day S, Weber J, Garbe C, et al. Ipilimumab plus dacarbazine for previously untreated metastatic melanoma. N Engl J Med. 2011;364:2517–26.
Robert C, Long GV, Brady B, Dutriaux C, Maio M, Mortier L, et al. Nivolumab in previously untreated melanoma without BRAF mutation. N Engl J Med. 2015;372:320–30.
Robert C, Schachter J, Long GV, Arance A, Grob JJ, Mortier L, et al. Pembrolizumab versus Ipilimumab in advanced melanoma. N Engl J Med. 2015;372:2521–32.
Postow MA, Chesney J, Pavlick AC, Robert C, Grossmann K, McDermott D, et al. Nivolumab and ipilimumab versus ipilimumab in untreated melanoma. N Engl J Med. 2015;372:2006–17.
Larkin J, Chiarion-Sileni V, Gonzalez R, Grob JJ, Cowey CL, Lao CD, et al. Combined Nivolumab and Ipilimumab or monotherapy in untreated melanoma. N Engl J Med. 2015;373:23–34.
Sharpe AH, Wherry EJ, Ahmed R, Freeman GJ. The function of programmed cell death 1 and its ligands in regulating autoimmunity and infection. Nat Immunol. 2007;8:239–45.
Daud AI, Wolchok JD, Robert C, Hwu WJ, Weber JS, Ribas A, et al. Programmed death-ligand 1 expression and response to the anti-programmed death 1 antibody Pembrolizumab in melanoma. J Clin Oncol. 2016;34:4102–9.
Ribas A, Puzanov I, Dummer R, Schadendorf D, Hamid O, Robert C, et al. Pembrolizumab versus investigator-choice chemotherapy for ipilimumab-refractory melanoma (KEYNOTE-002): a randomised, controlled, phase 2 trial. Lancet Oncol. 2015;16:908–18.
Weng YM, Peng M, Hu MX, Yao Y, Song QB. Clinical and molecular characteristics associated with the efficacy of PD-1/PD-L1 inhibitors for solid tumors: a meta-analysis. Onco Targets Ther. 2018;11:7529–42.
Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer. 2012;12:252–64.
Ribas A. Adaptive immune resistance: how Cancer protects from immune attack. Cancer Discov. 2015;5:915–9.
Tseng SY, Otsuji M, Gorski K, Huang X, Slansky JE, Pai SI, et al. B7-DC, a new dendritic cell molecule with potent costimulatory properties for T cells. J Exp Med. 2001;193:839–46.
Dong H, Strome SE, Salomao DR, Tamura H, Hirano F, Flies DB, et al. Tumor-associated B7-H1 promotes T-cell apoptosis: a potential mechanism of immune evasion. Nat Med. 2002;8:793–800.
Wang L, Pino-Lagos K, de Vries VC, Guleria I, Sayegh MH, Noelle RJ. Programmed death 1 ligand signaling regulates the generation of adaptive Foxp3+CD4+ regulatory T cells. Proc Natl Acad Sci U S A. 2008;105:9331–6.
Sun C, Mezzadra R, Schumacher TN. Regulation and function of the PD-L1 checkpoint. Immunity. 2018;48:434–52.
Atefi M, Avramis E, Lassen A, Wong DJ, Robert L, Foulad D, et al. Effects of MAPK and PI3K pathways on PD-L1 expression in melanoma. Clin Cancer Res. 2014;20:3446–57.
Garcia-Diaz A, Shin DS, Moreno BH, Saco J, Escuin-Ordinas H, Rodriguez GA, et al. Interferon receptor signaling pathways regulating PD-L1 and PD-L2 expression. Cell Rep. 2017;19:1189–201.
Schroder K, Hertzog PJ, Ravasi T, Hume DA. Interferon-gamma: an overview of signals, mechanisms and functions. J Leukoc Biol. 2004;75:163–89.
Audrito V, Serra S, Stingi A, Orso F, Gaudino F, Bologna C, et al. PD-L1 up-regulation in melanoma increases disease aggressiveness and is mediated through miR-17-5p. Oncotarget. 2017;8:15894–911.
Chen L, Gibbons DL, Goswami S, Cortez MA, Ahn YH, Byers LA, et al. Metastasis is regulated via microRNA-200/ZEB1 axis control of tumour cell PD-L1 expression and intratumoral immunosuppression. Nat Commun. 2014;5:5241.
Fujita Y, Yagishita S, Hagiwara K, Yoshioka Y, Kosaka N, Takeshita F, et al. The clinical relevance of the miR-197/CKS1B/STAT3-mediated PD-L1 network in chemoresistant non-small-cell lung cancer. Mol Ther. 2015;23:717–27.
Gong AY, Zhou R, Hu G, Li X, Splinter PL, O’Hara SP, et al. MicroRNA-513 regulates B7-H1 translation and is involved in IFN-gamma-induced B7-H1 expression in cholangiocytes. J Immunol. 2009;182:1325–33.
Wang W, Sun J, Li F, Li R, Gu Y, Liu C, et al. A frequent somatic mutation in CD274 3′-UTR leads to protein over-expression in gastric cancer by disrupting miR-570 binding. Hum Mutat. 2012;33:480–4.
Xu S, Tao Z, Hai B, Liang H, Shi Y, Wang T, et al. miR-424(322) reverses chemoresistance via T-cell immune response activation by blocking the PD-L1 immune checkpoint. Nat Commun. 2016;7:11406.
Zhao L, Yu H, Yi S, Peng X, Su P, Xiao Z, et al. The tumor suppressor miR-138-5p targets PD-L1 in colorectal cancer. Oncotarget. 2016;7:45370–84.
Cortez MA, Ivan C, Valdecanas D, Wang X, Peltier HJ, Ye Y, et al. PDL1 regulation by p53 via miR-34. J Natl Cancer Inst. 2016;108:1–9.
Kim MP, Lozano G. Mutant p53 partners in crime. Cell Death Differ. 2018;25:161–8.
Kastenhuber ER, Lowe SW. Putting p53 in context. Cell. 2017;170:1062–78.
Vieler M, Sanyal S. p53 Isoforms and Their Implications in Cancer. Cancers (Basel). 2018;10:E288.
Herold MJ, van den Brandt J, Seibler J, Reichardt HM. Inducible and reversible gene silencing by stable integration of an shRNA-encoding lentivirus in transgenic rats. Proc Natl Acad Sci U S A. 2008;105:18507–12.
Houben R, Adam C, Baeurle A, Hesbacher S, Grimm J, Angermeyer S, et al. An intact retinoblastoma protein-binding site in Merkel cell polyomavirus large T antigen is required for promoting growth of Merkel cell carcinoma cells. Int J Cancer. 2012;130:847–56.
Houben R, Hesbacher S, Schmid CP, Kauczok CS, Flohr U, Haferkamp S, et al. High-level expression of wild-type p53 in melanoma cells is frequently associated with inactivity in p53 reporter gene assays. PLoS One. 2011;6:e22096.
Lin J, Chen J, Elenbaas B, Levine AJ. Several hydrophobic amino acids in the p53 amino-terminal domain are required for transcriptional activation, binding to mdm-2 and the adenovirus 5 E1B 55-kD protein. Genes Dev. 1994;8:1235–46.
Appenzeller S, Gesierich A, Thiem A, Hufnagel A, Jessen C, Kneitz H, et al. The identification of patient-specific mutations reveals dual pathway activation in most patients with melanoma and activated receptor tyrosine kinases in BRAF/NRAS wild-type melanomas. Cancer. 2019;125:586–600.
Team RDC. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2008. Available online at http://www.R-project.org
Colaprico A, Silva TC, Olsen C, Garofano L, Cava C, Garolini D, et al. TCGAbiolinks: an R/Bioconductor package for integrative analysis of TCGA data. Nucleic Acids Res. 2016;44:e71.
Mayakonda A, Koeffler HP. Maftools: Efficient analysis, visualization and summarization of MAF files from large-scale cohort based cancer studies: bioRxiv. 2016. https://www.biorxiv.org/content/10.1101/052662v1.article-info.
Li J, Lu Y, Akbani R, Ju Z, Roebuck PL, Liu W, et al. TCPA: a resource for cancer functional proteomics data. Nat Methods. 2013;10:1046–7.
Robinson MD, McCarthy DJ, Smyth GK. edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics. 2010;26:139–40.
Ritter C, Fan K, Paschen A, Reker Hardrup S, Ferrone S, Nghiem P, et al. Epigenetic priming restores the HLA class-I antigen processing machinery expression in Merkel cell carcinoma. Sci Rep. 2017;7:2290.
Schmittgen TD, Livak KJ. Analyzing real-time PCR data by the comparative C(T) method. Nat Protoc. 2008;3:1101–8.
Wickham H. ggplot2: Elegant Graphics for Data Analysis. New York: Springer-Verlag; 2009.
Gu Z, Eils R, Schlesner M. Complex heatmaps reveal patterns and correlations in multidimensional genomic data. Bioinformatics. 2016;32:2847–9.
Mi H, Huang X, Muruganujan A, Tang H, Mills C, Kang D, et al. PANTHER version 11: expanded annotation data from gene ontology and Reactome pathways, and data analysis tool enhancements. Nucleic Acids Res. 2017;45:D183–D89.
Mi H, Thomas P. PANTHER pathway: an ontology-based pathway database coupled with data analysis tools. Methods Mol Biol. 2009;563:123–40.
Fischer M. Census and evaluation of p53 target genes. Oncogene. 2017;36:3943–56.
Youlyouz-Marfak I, Gachard N, Le Clorennec C, Najjar I, Baran-Marszak F, Reminieras L, et al. Identification of a novel p53-dependent activation pathway of STAT1 by antitumour genotoxic agents. Cell Death Differ. 2008;15:376–85.
Decker T, Kovarik P. Serine phosphorylation of STATs. Oncogene. 2000;19:2628–37.
Tanikawa C, Zhang YZ, Yamamoto R, Tsuda Y, Tanaka M, Funauchi Y, et al. The transcriptional landscape of p53 Signalling pathway. EBioMedicine. 2017;20:109–19.
Box NF, Vukmer TO, Terzian T. Targeting p53 in melanoma. Pigment Cell Melanoma Res. 2014;27:8–10.
Munoz-Fontela C, Mandinova A, Aaronson SA, Lee SW. Emerging roles of p53 and other tumour-suppressor genes in immune regulation. Nat Rev Immunol. 2016;16:741–50.
Braun MW, Iwakuma T. Regulation of cytotoxic T-cell responses by p53 in cancer. Transl Cancer Res. 2016;5:692–7.
Zhu K, Wang J, Zhu J, Jiang J, Shou J, Chen X. p53 induces TAP1 and enhances the transport of MHC class I peptides. Oncogene. 1999;18:7740–7.
Wang B, Niu D, Lai L, Ren EC. p53 increases MHC class I expression by upregulating the endoplasmic reticulum aminopeptidase ERAP1. Nat Commun. 2013;4:2359.
Xiao W, Du N, Huang T, Guo J, Mo X, Yuan T, et al. TP53 mutation as potential negative predictor for response of anti-CTLA-4 therapy in metastatic melanoma. EBioMedicine. 2018;32:119–24.
DeLeon T, Kipp B, Langlais B, Flotte T, Winters J, Kosiorek HE, et al. Assessment of treatment response to immunotherapy in melanoma patients with pathogenic mutations of NRAS, BRAF, CDKN2A and P53. J Clin Oncol. 2017;35:e21057–e57.
Yu XY, Zhang XW, Wang F, Lin YB, Wang WD, Chen YQ, et al. Correlation and prognostic significance of PD-L1 and P53 expression in resected primary pulmonary lymphoepithelioma-like carcinoma. J Thorac Dis. 2018;10:1891–902.
Agersborg S, Jiang S, Chen W, Ma W, Albitar M. PD-L1 expression correlation with TP53 gene mutation status in lung cancer but not in colorectal cancer. J Clin Oncol. 2016;34:11557.
Taube JM, Anders RA, Young GD, Xu H, Sharma R, McMiller TL, et al. Colocalization of inflammatory response with B7-h1 expression in human melanocytic lesions supports an adaptive resistance mechanism of immune escape. Sci Transl Med. 2012;4:127ra37.
Tumeh PC, Harview CL, Yearley JH, Shintaku IP, Taylor EJ, Robert L, et al. PD-1 blockade induces responses by inhibiting adaptive immune resistance. Nature. 2014;515:568–71.
Knol AC, Nguyen JM, Pandolfino MC, Denis MG, Khammari A, Dreno B. PD-L1 expression by tumor cell lines: a predictive marker in melanoma. Exp Dermatol. 2018;27:647–55.
Wiesner T, Kiuru M, Scott SN, Arcila M, Halpern AC, Hollmann T, et al. NF1 mutations are common in desmoplastic melanoma. Am J Surg Pathol. 2015;39:1357–62.
Kraft S, Fernandez-Figueras MT, Richarz NA, Flaherty KT, Hoang MP. PDL1 expression in desmoplastic melanoma is associated with tumor aggressiveness and progression. J Am Acad Dermatol. 2017;77:534–42.
Speidel D. Transcription-independent p53 apoptosis: an alternative route to death. Trends Cell Biol. 2010;20:14–24.
Speidel D, Helmbold H, Deppert W. Dissection of transcriptional and non-transcriptional p53 activities in the response to genotoxic stress. Oncogene. 2006;25:940–53.
Yoon KW, Byun S, Kwon E, Hwang SY, Chu K, Hiraki M, et al. Control of signaling-mediated clearance of apoptotic cells by the tumor suppressor p53. Science. 2015;349:1261669.
Gowrishankar K, Gunatilake D, Gallagher SJ, Tiffen J, Rizos H, Hersey P. Inducible but not constitutive expression of PD-L1 in human melanoma cells is dependent on activation of NF-kappaB. PLoS One. 2015;10:e0123410.
Lowe JM, Menendez D, Bushel PR, Shatz M, Kirk EL, Troester MA, et al. p53 and NF-kappaB coregulate proinflammatory gene responses in human macrophages. Cancer Res. 2014;74:2182–92.
Luo N, Formisano L, Gonzalez-Ericsson PI, Sanchez V, Dean PT, Opalenik SR, et al. Melanoma response to anti-PD-L1 immunotherapy requires JAK1 signaling, but not JAK2. Oncoimmunology. 2018;7:e1438106.
Shin DS, Zaretsky JM, Escuin-Ordinas H, Garcia-Diaz A, Hu-Lieskovan S, Kalbasi A, et al. Primary resistance to PD-1 blockade mediated by JAK1/2 mutations. Cancer Discov. 2017;7:188–201.
Zaretsky JM, Garcia-Diaz A, Shin DS, Escuin-Ordinas H, Hugo W, Hu-Lieskovan S, et al. Mutations associated with acquired resistance to PD-1 blockade in melanoma. N Engl J Med. 2016;375:819–29.
Acknowledgments
The authors thank Eva Maria Sarosi, Claudia Siedel, Lena Neuß and Ellina Schulz for excellent technical assistance. Furthermore, we greatly appreciate the support of Prof. Annette Paschen, who provided the JAK2 expression plasmid.
Funding
This work was made possible by a Clinician Scientist scholarship that was awarded to Alexander Thiem by the Comprehensive Cancer Center Mainfranken and furthermore supported by the Interdisziplinäres Zentrum für Klinische Forschung (IZKF) of the Medical Faculty, University of Würzburg (IZKF B-323).
This publication was funded by the German Research Foundation (DFG) and the University of Wuerzburg in the funding programme Open Access Publishing.
Author information
Authors and Affiliations
Contributions
AT designed and performed experiments, analyzed data and wrote the manuscript. SH, HK, TdP performed experiments. MH provided the 1205Lu TP53-knockout cell line and helped preparing resubmission. HH assisted with analysis of JAK-STAT-signaling. MG discussed experiments and provided conceptual input. SM had the initial idea of this project and provided cell lines. RH designed experiments and analyzed data. DS designed and performed experiments, analyzed data and wrote the manuscript. All authors critically reviewed the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Written informed consent was obtained from each patient prior to obtaining the sample to use for research purposes. Analyses of tumor samples was approved by the Ethics Committee of the Medical Faculty, University of Würzburg (study reference: 241/2014).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Additional file 1: Figure S1.
Clinical characteristics of patients included in immunohistochemistry study and weak negative correlation of TP53 and CD274 in TP53-wildtype TCGA samples. (a) A total of 81 samples were included in this analysis. ALM indicates acral lentiginous melanoma; LMM, lentigo maligna melanoma, NM, nodular melanoma; SSM, superficial spreading melanoma; mut, mutated; wt, wildtype. (b) Linear regression analysis of TP53 mRNA with CD274 mRNA (n = 347) was conducted separately for TP53-wt and -mutant samples. (JPG 1844 kb)
Additional file 2: Figure S2.
miR-34a is lower expressed in TP53-mutated tumors but does not correlate with PD-L1 protein in melanoma. (a) Wilcoxon-Mann-Whitney test was applied to compare miR34a expression between TP53-wt and -mutant samples. (b) Linear regression analysis was used to analyze the relation between miR34a and PD-L1 protein. (c) Correlation of CD274 with miRs was calculated using spearman correlation. Only miRs with > 1 RPM in a least 20% of cases were included. The 24 best correlating miRs are presented in a “heatmap”. Expression values are presented in a spectrum of blue (small values) to red (high values). (d) Linear regression analysis of miR7702 and CD274. (e) Wilcoxon-Mann-Whitney test was applied to compare miR7702 expression between TP53-wt and -mutant samples. (f) Linear regression analysis of miR7702 with CD274 was conducted separately for TP53-wt and -mutant samples. p < 0.05 is regarded as statistically significant. Mut, mutated; TPM, transcripts per millions; RPM, reads per million; wt, wildtype. (JPG 1937 kb)
Additional file 3: Figure S3.
IFN-ɣ induced PD-L1 expression is reduced after p53 knockdown in melanoma while HLA expression is hardly affected. (a) Flow cytometry for PD-L1 expression. Presented is the PD-L1 mean normalized to the mean of control cells cultured without DOX or IFN-ɣ for 48 h. The data are presented as the mean + s.e.m. of three independent experiments. *p < 0.05, ns = not significant. (b) The effect of a second TP53-targeting shRNA on IFN-ɣ-induced PD-L1 expression. Those melanoma cell lines, which had revealed the most distinctive reduction of IFN-ɣ-induced PD-L1 expression upon p53 knockdown by the first shRNA, were transduced by another inducible shRNA. IFN-ɣ treatment was for 48 h. PD-L1 and p53 expression was measured by immunoblot. ß-tubulin served as a loading control. Blot is representative of two individual experiments. (c) LOX-IMVI and UACC-257, transduced with the first inducible TP53-targeting shRNA, were treated with IFN-ɣ for 48 h. MHC-I (grey) and PD-L1 (black) expression was measured by flow cytometry. Depicted is the fluorescence mean normalized to the mean of control cells treated neither with doxycycline nor IFN-ɣ. The data is presented as the mean + s.e.m. of three independent experiments. *p < 0.05, **p < 0.01, ns = not significant. p53 knockdown was achieved by culturing cells in doxycycline for 6 days in all experiments. (JPG 1296 kb)
Additional file 4: Figure S4.
No differential expression of IFN-ɣ between TP53-genotypes, but positive correlation of IFN-ɣ and PD-L1 expression for both genotypes. (a) Wilcoxon-Mann-Whitney test was applied to compare IFN-ɣ expression between TP53-wt and -mutant samples. (b, c) Linear regression analysis of IFN-ɣ with p53 protein (b) or with PD-L1 protein (c) was conducted separately for TP53-wt and -mutant samples. p < 0.05 is regarded as statistically significant. Mut, mutated; TPM, transcripts per millions; wt, wildtype. (JPG 591 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Cite this article
Thiem, A., Hesbacher, S., Kneitz, H. et al. IFN-gamma-induced PD-L1 expression in melanoma depends on p53 expression. J Exp Clin Cancer Res 38, 397 (2019). https://doi.org/10.1186/s13046-019-1403-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13046-019-1403-9