
Abstract
Background
Tubeimoside-I (TBM), a plant-derived bioactive compound, shows antitumor activity in different tumors and can enhance the efficacy of chemotherapeutic agents. However, the detail mechanism underlying remains to be elucidated.
Methods
The cytotoxic potential of TBM towards CRC cells was examined by CCK8 assay, colony formation, LDH release assay, flow cytometry method and Western blots. The ROS levels, autophagy, apoptosis, chemosensitivity to 5-FU or DOX, etc. were determined between control and TBM-treated CRC cells.
Results
In this study, we found that TBM could inhibit proliferation and induce apoptosis in colorectal cancer (CRC) cells. Intriguingly, TBM treatment could either promote autophagy initiation by ROS-induced AMPK activation, or block autophagy flux through inhibiting lysosomal hydrolytic enzymes, which leaded to massive impaired autophagylysosomes accumulation. Administration of autophagy initiation inhibitor (3-MA or selective ablation of autophagy related proteins) relieves TBM-induced CRC suppression, while combination use of autophagy flux inhibitor chloroquine (CQ) slightly augments TBM-induced cell death, suggesting that impaired autophagylysosomes accumulation contributes to TBM-induced growth inhibition in CRC cells. Notably, as an autophagy flux inhibitor, TBM works synergistically with 5-fluorouracil (5-FU) or doxorubicin (DOX) in CRC suppression.
Conclusion
Together, our study provides new insights regarding the anti-tumor activity of TBM against CRC, and established potential applications of TBM for CRC combination therapies in clinic.
Similar content being viewed by others


Background
Although the dissemination of colonoscopy with polypectomy has led to a decline in the incidence and mortality rates, colorectal cancer (CRC) still is the third most diagnosed cancers and the second leading causes of cancer death among both men and women worldwide, with over 1.8 million new colorectal cancer cases and 881,000 deaths in 2018 [1, 2]. If CRC patients can be diagnosed and treated at localized stage, 5-year survival rate of which is about 90%, that will decline to 71 and 14% for patients diagnosed with regional and distant-stage disease [1, 3]. Adjuvant chemotherapy is the main treatment strategy for advanced CRC, however, a subset of patients eventually become relapsed because of acquired drug resistance to the current commonly used chemotherapeutic drugs and the effectiveness of chemotherapy has often been limited [4, 5]. Hence at present, development of new drugs for colorectal cancer is an urgent need.
The stem tuber of Bolbostemma paniculatum (Maxim.) Franquet (Cucurbitaceae) is a traditional Chinese medicine named as Rhizoma Bolbostemmatis (Chinese name“Tu Bei Mu”) [6, 7]. It was listed in the Supplement to the Compendium of Materia Medica in Qing Dynasty for treating breast cancer, acute mastitis, inflammation and snake venoms [6, 7]. Tubeimoside-I (TBM), a triterpenoid saponin, is isolated from Rhizoma Bolbostemmatis and shows antitumor activity in different tumor such as promyelocytic leukemia, lung cancer, cervical cancer, nasopharyngeal carcinoma, esophageal carcinoma with low toxicity [6, 7]. TBM could trigger a mitochondrial-related apoptotic pathway and cell cycle arrest in cervical carcinoma, ovarian cancer, choriocarcinoma and glioma [8,9,10,11]. TBM can also inhibit the growth and invasion of CRC cells [12]. More interestingly, tubeimosides are effective in combination therapies, particularly at targeting drug-resistant cancerous cells [13]. However, the underlying mechanisms of its anti-tumor activity have not been fully clarified so far, especially in CRC.
Autophagy is a highly conserved catabolic process during which de novo-formed membrane-enclosed vesicles engulf damaged or senescent organelles and transport to lysosomes for degradation and recycling in response to nutrient starvation or metabolic stress [14, 15]. Autophagy plays an important role in the regulation of cancer progression and in determining the response of tumor cells to stress induced by chemotherapy [14, 15]. Four functional forms of autophagy induced by chemotherapy have been described to date: the cytoprotective autophagy which facilitates the resistance of cancer cells to chemotherapeutic drugs, cytotoxic autophagy which promotes cell death, cytostatic autophagy which prolongs growth inhibition as well as reduced clonogenic survival, and nonprotective autophagy which don’t affect drug or radiation sensitivity [16]. Considering the crucial roles during chemotherapy, targeting the autophagy process has recently attracted considerable attention to develop novel antitumor therapeutics via pharmacological modulation of autophagy. Tubeimoside-I can induce cytoprotective autophagy in human breast cancer cells in vitro, while promote cytotoxic autophagy in cervical cancer cells [17, 18]. However, the role of autophagy in TBM-treated CRC cells remains largely unexplored, let alone the underlying mechanisms.
In this study, we found that TBM inhibited the growth of CRC cells by stimulating apoptosis. Interestingly, TBM elicits autophagy by ROS-induced activation of AMPK and blocks autophagic flux by impairing the degradation of the autophagolysosome, which contributes to TBM-induced anti-cancer activity. Notably, as an autophagy modulator, TBM synergistically suppresses CRC cell growth with 5-FU or DOX. These findings provide the evidences for the use of TBM as a new therapeutic agent against CRC, especially in combination chemotherapy.
Material and methods
Cell culture
The SW480 and SW620 cell lines were purchased from the American Type Culture Collection (ATCC), HCT116 and RKO cell lines were purchased from Shanghai cell bank. All cell lines were cultured according to the guidelines and were maintained in DMEM (Gibco) supplemented with 100 U/mL penicillin (Sigma), 10 mg/L streptomycin (Sigma), and 10% serum (Hyclone) in a humidified incubator at 37 °C under 5% CO2 atmosphere.
Reagents and antibodies
The following primary antibodies were used: Lamp1 (Santa Cruz Biotechnology), LC3 (MBL International Corporation), p62 (MBL International Corporation), PARP 1(Cell Signaling Technology), PRKAA/AMPK (Cell Signaling Technology), phosphor-PRKAA/ AMPK(Cell Signaling Technology), Beclin1 (Cell Signaling Technology), ATG5 (Cell Signaling Technology), CASP9 (Cell Signaling Technology), CASP3 (Cell Signaling Technology) and Cleaved-CASP3 (Cell Signaling Technology). TBM (BP1415) was purchased from Phytopurify Biotechnology. 3-methyladenine (3-MA), chloroquine (CQ) and N-acetyl-L-cysteine (NAC) were purchased from Sigma-Aldrich. Doxorubicin (DOX, HY-15142A) and 5-fluorouracil (5-FU, HY-90006) were from MedChem Express.
Transfection
All siRNAs were designed using BLOCK-iT™ RNAi Designer (Invitrogen) and synthesized by GenePharma (Shanghai, China). The sequences of the siRNAs used are listed in Additional file 1: Table S1. Cells were transfected with siRNAs using Lipofectamine RNAiMax (Invitrogen) according to the manufacturer’s instruction.
Dominant-negative AMPK (DN-PRKAA1/DN-AMPKα1) plasmid was obtained as described previously [19]. The plasmids were transfected into the indicated cells using Lipofectamine 2000 (Invitrogen) according to the manufacturer’s instruction.
Immunofluorescence
Cells grown on coverslips that treated with TBM for 24 h were fixed with 4% paraformaldehyde (Sigma) for 30 min and then washed three times with PBS. Fixed cells were permeabilized with 0.4% Triton 100 and 5% BSA for 1 h at 37 °C. For staining, cells were incubated with primary antibodies for 12 h at 4 °C, followed by incubation with secondary antibodies (Thermos Fisher Scientific) for 1 h at 37 °C. Finally, Nuclei were stained with DAPI for 10 min. For autophagy flux studies, cells were transfected with GFP-RFP-LC3 for 24 h and then treated with TBM for another 24 h. Images were captured using a confocal microscopy (Carl Zess Micromaging).
Cell viability and proliferation assays
As described previously [20], CCK8 assay was performed to determine cell viability. Briefly, cells were seeded in 96-well plates at a density of 5000 cells. After treatment, CCK8 (Dojindo, Kamimashiki-gun, Kumamoto, Japan; CK04) were added and incubated for 40 min. The absorbance value was then determined at 450 nm.
The long-term effects of TBM on CRC cell proliferation were analyzed with a colony formation assay. After treatment, 1500–2000 cells were seeded in six-well plates and the medium was changed every 3 days. After two weeks, cells were fixed with 4% paraformaldehyde (Sigma) for 30 min and stained with Crystal Violet for another 30 min, then the colonies were washed three times and taken photos.
Immunoblot and immunoprecipitation
Cells were lysed with RIPA buffer (50 mM Tris base, 1.0 mM EDTA, 150 mM NaCl, 0.1% SDS, 1% Triton X-100, 1% sodium deoxycholate, 1 mM PMSF) supplemented with protease inhibitor cocktail (Sigma) and then protein lysates were centrifuged and boiled with loading buffer. For immunoprecipitation, Whole cell lysates were prepared in RIPA buffer (40 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.5% Nonidet P-40, cocktail, 5% glycerol, 10 mM NaF) and incubated with 1 μg of antibody overnight at 4 °C. Next day, Sepharose protein A/protein G beads were added for 2 h. The immune-complexes were then centrifuged and washed 3 times using RIPA buffer. All lysates were quantified by the BCA Protein Assay (Thermo Fisher Scientific) and analysed by SDS–PAGE.
Transmission electron microscopy
Transmission electron microscopy assay was used to visualize autophagic vesicles. After 24-h TBM treatment, SW480 and HCT116 cells were fixed in glutaraldehyde (Sigma) and ultrathin sections were prepared using a sorvall MT5000 microtome. Then, the sections were stained by lead citrate and /or 1% uranyl acetate and visualized by Philips EM420 electron microscopy.
Flow cytometry
As described previously [21], the indicated cells treated with TBM for 24 h were harvested and washed three times with PBS, then resuspended and incubated with PI-ANXA5 solution (KeyGEN Biotech). Apoptosis was analyzed with a FACSCalibur flow cytometer (Becton Dickinson, San Jose, CA USA).
Reactive oxygen species (ROS) measurement
Intracellular ROS level was detected by staining cells with 2′, 7′-dichlorofluorescein diacetate (DCFH-DA) (GENMED, GMS10016.2) according to the manufacturer’s instructions. The DCFH-DA signal was measured with a FACSCalibur flow cytometer or a Molecular Devices SPECTRAMAX M5 fluorimeter (490 nm excitation and 530 nm emission).
Data analysis and statistics
Data were expressed as means ± s.d. All experiments were performed at least three times. Statistical analysis was performed with GraphPad Prism 6.0 software. Statistical differences between groups were determined using two-tailed Student’s t-test. Significance was designated as follows: *, P < 0.05, **, P < 0.01, ***, P < 0.001.
Results
TBM inhibits proliferation and induces apoptosis in CRC cell
To validate whether TBM exhibited antitumor effect against CRC, CCK8 assay was performed to assess the cell viability in response to TBM treatment in different human CRC cell lines. As shown in Fig. 1a, TBM treatment for 24 h markedly decreased the cell viability of various CRC cell lines (SW480, HCT116, SW620 and RKO) in a dose-dependent manner. Consistently, the proliferation of CRC cells was significantly inhibited under TBM treatment, as evidenced by reduced colony formation (Fig. 1b). To further verify its anti-CRC activity, lactate dehydrogenase (LDH) release assay was performed and the results showed that TBM treatment exhibited marked cytotoxicity in SW480 and HCT116 cells (Fig. 1c). The initiation of apoptosis is largely involved in the antitumor activity of chemotherapeutic drugs, thus TUNEL assay and Annexin-V/PI double staining flow cytometry method were used to evaluate whether apoptosis is involved in TBM-induced cytotoxicity in CRC. As shown as Fig. 1d and Additional file 1 :Figure S1A, TBM could induce a prominent apoptotic effect in CRC cells with a dose-dependent manner. The pro-apoptotic effect of TBM was further supported by increased levels of cleaved PARP1, CASP9 and CASP3, the specific and sensitive markers of apoptosis, in TBM-treated cells (Fig. 1e and Additional file 1 :Figure S1B). Taken together, these results indicated that TBM exhibited a significant antitumor effect in CRC cells.

TBM inhibits proliferation and induces apoptosis in CRC cells. a. SW480, HCT116, SW620 and RKO cells were treated with the indicated concentration of TBM for 24 h and cell viability was determined by CCK8 kit. b. Cell proliferation rate was analyzed by clone formation assay. SW480, HCT116, SW620 and RKO cells were treated with the indicated concentration of TBM for 24 h, after treatment, cells were seeded into 6-well plates for two weeks and colony numbers were quantified. c. Analysis of dehydrogenase (LDH) release in SW480 and HCT116 cells subjected to indicated concentration of TBM. Cell lysates serve as positive control. d. SW480 and HCT116 cells treated with 10 μM TBM for 24 h were fixed, stained with Annexin V/PI, and then analyzed by flow cytometry. e. Immunoblot analysis of PARP1 in HCT116 and SW480 cells. Data are means ± s.d. and are representative of 3 independent experiments. *, P < 0.05; **, P < 0.01; ***, p < 0.001
TBM activates autophagy in CRC cell
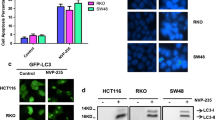
Accumulated evidence has shown that autophagy was usually activated by chemotherapeutic drugs and contributed to antitumor effect [14, 15]. To test whether autophagy is regulated by TBM in CRC cells, we first examined the processing of LC3-I to its PE-conjugated LC3-II, which is one of the most widely accepted methods for monitoring autophagy [22]. As indicated in Fig. 2a and b, TBM treatment promoted the turnover of LC3-I to lipidated LC3-II in a dose- and time-dependent manner in CRC cells. In line with this, accumulation of GFP-LC3 puncta was dramatically increased in CRC cells treated with TBM by fluorescence images (Fig. 2c). Furthermore, transmission electron microscopy experiment also showed that the appearance of double-membraned autophagosomes was frequently observed in CRC cancer cells treated with TBM (Fig. 2d).

TBM induces autophagy in CRC cells. a. Immunoblot analysis of LC3 and p62 expression in SW480 and HCT116 cells treated with indicated concentrations of TBM for 24 h. b. Time course analysis of LC3 and p62 expression in SW480 and HCT116 cells treated with TBM (10 μM) for 6, 12, 24, 48 h by immunoblotting. c. SW480 and HCT116 cells were transfected with GFP-LC3 for 48 h, and then treated with 10 μM TBM for another 24 h. GFP-LC3 puncta was shown and quantitated by immunofluorescence analysis. Scale bar, 10 μm. d. Autophagic vesicles were detected by transmission electron microscope in SW480 and HCT116 cells treated as in (C). Scale bar, 2.5 or 1 μm. Data are means ± s.d. and are representative of 3 independent experiments. *, P < 0.05; **, P < 0.01; ***, p < 0.001
Next, we examined the expression levels of Atg5 and Beclin 1, two autophagy-related proteins, which is correlated with the extent of autophagosome formation. Results showed that TBM induced the expression of Atg5 and Beclin 1 in a dose-dependent manner in CRC cells (Fig. 3a-b). While silencing the expression of Beclin 1 or Atg5 using siRNA partially blocked LC3 lipidation in TBM-treated cells (Fig. 3c-d). Coadministration of 3-MA, a class III PI3K inhibitor, with TBM failed to induce autophagy (Fig. 3e-f). In addition, it has been reported that the diminished interaction of Beclin 1 with Bcl-2 are key events in the initiation process of autophagy [22]. As shown in coimmunoprecipitation assay, TBM treatment decreased the binding of Beclin 1 with Bcl-2 (Fig. 3g). Collectively, our data suggest that TBM induce autophagy initiation and autophagosome accumulation in CRC cells.

TBM induces autophagy initiation in CRC cells. a. Immunoblot analysis of Atg5 expression in SW480 and HCT116 cells treated with indicated concentrations of TBM for 24 h. b. Immunoblot analysis of Beclin 1 expression in HCT116 and SW480 cells treated with indicated concentrations of TBM for 24 h. c. Immunoblot analysis of LC3 and Atg5 expression in CRC cells transfected with siNC or siAtg5 for 24 h, followed by treatment with or without 10 μM TBM for another 24 h. d. Immunoblot analysis of LC3 and Beclin 1 expression in HCT116 and SW480 cells transfected with siNC or siBeclin 1 for 24 h, followed by treatment with or without 10 μM TBM for another 24 h. e. SW480 and HCT116 cells were treated with vehicle, TBM (10 μM), 3-MA (5 mM), or in combination for 24 h. Immunoblot analysis was used to detect the expression of LC3. f. SW480 and HCT116 cells were transfected with GFP-LC3 for 48 h, and then were treated as in (E). The LC3 puncta by immunofluorescence analysis was shown and quantitated. Scale bar, 10 μm. g. Co-immunoprecipitation analysis of the interaction between Beclin1 and Bcl-2 in SW480 and HCT116 cells treated with or without 10 μM TBM for 24 h. ***, p < 0.001
ROS/AMPK axis contributes to TBM-induced autophagy initiation
Previous studies have shown that TBM promote cell death mainly by triggering a mitochondrial-related apoptotic pathway [13], which may decrease the production of energy and then activate AMPK signaling pathway to initiate autophagy [23]. To ascertain this hypothesis, we detected the phosphorylation status of AMPK to validate whether AMPK was involved in TBM-induced autophagy. As shown in Fig. 4a, TBM treatment resulted in activation of AMPK as evidenced by increased phosphorylation levels of AMPK in a dose-dependent manner in TBM-treated cells. To determine whether the activation of AMPK was involved in TBM-induced autophagy, we transfected a siRNA target AMPK to inhibit AMPK expression or domain negative AMPK (DNAMPK) to inhibit AMPK activation. As shown in Fig. 4b-c, inhibition of AMPK prominently inhibited elevation of LC3 lipidation in TBM-treated cells. Dysfunction of mitochondria-induced activation was mainly involved in ROS generation, thus, we determined whether TBM could increase cellular ROS levels. As shown as Fig. 4d, treatment of TBM dramatically increased cellular ROS levels in SW480 and HCT116 cells. Accumulating evidence has shown ROS in response to cancer treatment can promote autophagy by activating various signaling pathways including AMPK signaling. Next, we would like to identify the role of ROS in TBM-induced autophagy initiation. We found that combinatorial treatment of TBM with ROS scavenger N-Acetyl-cysteine (NAC) resulted in AMPK inhibition, reduced LC3-II turnover and accumulation of GFP-LC3 punta, decreased expression of Beclin1 and Atg5 (Fig. 4e and f). Collectively, these results suggest that activation of ROS/AMPK signaling pathway is required for TBM-induced autophagy in CRC cells.

TBM-induced autophagy is dependent on ROS/AMPK activation in CRC cells. a. SW480 and HCT116 cells were treated with indicated concentrations of TBM. The total and phosphorylation levels of AMPK were examined by immunoblotting. b. Immunoblot analysis of LC3, AMPK and p-AMPK expression in SW480 and HCT116 cells transfected with siNC or siAMPK for 24 h, followed by treatment with or without 10 μM TBM for another 24 h. c. SW480 and HCT116 cells were transfected with Vector or DN-AMPK for 24 h, followed by treatment with or without 10 μM TBM for another 24 h. Immunoblot analysis of LC3, AMPK and p-AMPK expression in CRC cells. d-e. SW480 and HCT116 cells were treated with vehicle, TBM (10 μM), NAC (5 mM), or in combination for 24 h. ROS was detected by flow cytometric analysis (D), and the levels of cleaved-PARP1, p-AMPK, AMPK, Beclin1, Atg5 and LC3 were examined by immunoblotting (E). F. SW480 and HCT116 cells were transfected with GFP-LC3 for 48 h, and then treated as (D-E). GFP-LC3 puncta was shown and quantitated by immunofluorescence analysis. Scale bar, 10 μm. ***, p < 0.001
TBM impairs autophagy flux in CRC cell by inhibiting proteolytic activity of lysosome
To determine whether TBM can also induce complete autophagy flux in CRC cells, we firstly examined the protein levels of p62, a well-known autophagic substrate. However, the results showed that endogenous p62 protein expression were notably increased dose- and time-dependently after TBM stimulation (Fig. 2a-b), suggesting that autophagic flux may be inhibited by TBM. To further confirm this result, we measured autophagy flux by monitoring the conversion of LC3-I to LC3-II in the presence of chloroquine (CQ), a lysosomal inhibitor. The result showed that combinatorial treatment of TBM with CQ did not further significantly enhanced LC3-II turnover in CRC cells (Fig. 5a-c). Using a tandem monomeric RFP-GFP–tagged LC3, we also found that TBM treatment induced notable formation of yellow fluorescent autophagosomes and slight increase of red fluorescent autophagolysosomes (Fig. 5d-e). These results demonstrated that TBM impairs autophagy flux in CRC cells.

TBM inhibits autophagy flux in CRC cells. a. SW480 and HCT116 cells were treated with vehicle, TBM (10 μM), CQ (5 μM), or in combination for 24 h. Immunoblot analysis was used to detect the expression of LC3. b. The accumulation of LC3 puncta was examined by fluorescent analysis of cells transfected with GFP-LC3 plasmids and treated with CQ (5 μM) in the presence or absence of TBM (10 μM) for 24 h. Scale bar, 10 μm. c. The number of LC3 puncta was quantitated. d. Fluorescence analysis of cells transiently transfected with tandem mRFP-GFP-tagged LC3 and treated with vehicle or TBM (10 μM). Scale bar, 10 μm. e. The ratio of red puncta indicating autophagolysosome (GFP−/RFP+) versus yellow puncta indicating autophagosome (GFP+/RFP+) was quantitated. f. SW480 and HCT116 cells were treated with indicated concentrations of TBM. The expression of LAMP1 was examined by immunoblotting. g. Immunofluorescence analysis of the co-localization of endogenous LC3 and LAMP1 in SW480 and HCT116 cells treated with vehicle or TBM (10 μM) for 24 h. Scale bar, 10 μm. h. Immunoblot of ubiquitin in SW480 and HCT116 cells treated with TBM in indicated concentrations for 24 h. ***, p < 0.001
Autophagy flux signified autophagosomes fuse with lysosomes for further degradation, thus dysfunction of lysosomes may play an important role in inhibition of TBM-induced autophagy flux. We firstly detected the expression of lysosome- associated membrane protein (LAMP) 1, a marker for mature lysosome. As shown in Fig. 5f, TBM significantly up-regulated of LAMP1 in a dose dependent manner, indicating that TBM promote lysosomal mature. We subsequently examined whether autophagosome-lysosome fusion was affected by TBM by assessing the colocalization of endogenous LC3 with LAMP1. However, our results also showed that TBM treatment induced obvious colocalization of LC3 with LAMP1, suggesting the fusion of autophagosome with lysosome (Fig. 5g). Finally, we compared the lysosomal hydrolytic enzymes after TBM treatment by examining the level of ubiquitinated proteins. Interestingly, ubiquitinated protein conjugates was increased following TBM treatment (Fig. 5h). These results suggested that TBM impairs autophagy flux by inhibiting proteolytic activity of lysosome.
ROS-induced impaired autophagolysosomes accumulation contributes to TBM-induced cell death
Impaired autophagolysosomes accumulation is harmful to cancer cells and may lead to cell death. To evaluate whether autophagy was involved in the anti-CRC effect of TBM, we transfected CRC cells with Beclin1 or ATG5 siRNA, followed by TBM treatment. Cell viability was measured with CCK8 assay and the pro-apoptotic effect was examined by Annexin V/PI staining measured with flow cytometry. These data demonstrated that reduction in ATG5 or Beclin1 levels rescued the cell viability and inhibited the cell apoptosis caused by TBM (Fig. 6a and Additional file 1 :Figure S2A). Furthermore, similar results were obtained by using the autophagic inhibitors, 3-MA (Fig. 6b and Additional file 1 :FigureS2B). However, combination use of TBM with CQ slightly exacerbated TBM-induced cell death (Fig. 6b and Additional file 1 :FigureS2B). Next, we also elucidated the role of ROS accumulation in TBM-induced growth suppression. As shown in Fig. 6c-d, combinatorial treatment of TBM with ROS scavenger (NAC) inhibited TBM-induced growth suppression. In addition, treatment of NAC also reduced the pro-apoptotic effect of TBM, as evidenced by flow cytometry and levels of cleaved PARP1 (Fig. 4e and Additional file 1 :FigureS2C). These data indicated that TBM-induced autophagolysosome accumulation by ROS contributes to cell death in CRC cells.

ROS-induced impaired autophagolysosomes accumulation contributes to TBM-induced cell death. a. Cells were transfected with siScramble, siATG5 or siBeclin1 for 24 h, and then treated with 10 μM TBM for another 24 h. Cell viability was detected by CCK8 assay. b. Cells were treated with 10 μM TBM in the absence or presence of 3-MA (5 mM) or CQ (5 μM). Cell viability was determined by CCK8 assay. c. Cells were treated with 10 μM TBM in the absence or presence of NAC (5 mM). Cell viability was determined by CCK8 assay. d. Cells were treated as in (C) and ROS level was analyzed by DCFH-DA staining via a fluorescence microplate Reader (Thermos scientific). ***, p < 0.001
TBM enhances the chemotherapeutic response of CRC cells response to 5-FU and DOX
Previously studies have found that pharmacological modulation of autophagy can merit exploration as a therapeutic strategy for cancer treatment and can increase sensitivity of cancer cells to other chemotherapeutic agents which usually induces protective autophagy [14,15,16]. As a specific autophagy flux inhibitor, we wonder if TBM can also enhance the chemosensitivity of CRC cells to 5-FU or DOX. Firstly, we determine the antitumor effect of 5-FU and DOX to CRC cell lines with a variety of concentration. As shown as Fig. 7a-b, the cell growth of CRC cells can be inhibited by 5-FU and DOX, however the effect is not obvious as the concentration increased. To obtain the best result of TBM in combination 5-FU and DOX, CRC cells were combinational treated of TBM with 5-FU or DOX in different concentration. The result reveals that 2 μM 5-FU combined with 0.2 μM or 0.5 μM TBM exhibits cost-effective anticancer effect in SW480 cells and HCT116 cells, respectively, while 0.2 μM DOX combined with 1 μM TBM markedly decreased the cell viability in both SW480 cells and HCT116 cells. Consistently, significantly lower cell growth and clone formation are observed in the combination arm with this treatment (Fig. 7c-d). Together, these results indicated that TBM effectively sensitizes CRC cells to 5-FU and DOX treatment.

TBM enhances the chemotherapeutic response of CRC cells response to 5-FU and DOX. a-b. SW480 (A) and HCT116 (B) cells were treated with indicated concentrations of 5-fluorouracil (5-FU) or doxorubicin (DOX) in the absence or presence of indicated concentrations TBM for 24 h and CCK8 assays were performed to assess cell viability. c-d. Cell proliferation rate was analyzed by clone formation assay. SW480 and HCT116 cells were treated with 5-fluorouracil (5-FU, 2 μM) (C) or doxorubicin (DOX, 0.4 μM) (D) in the absence or presence of indicated concentrations TBM (10 μM) for 24 h, after treatment, cells were seeded into 6-well plates for two weeks and colony numbers were quantified. ***, P < 0.001
Discussion
Recently, TBM has been found to have considerable potential to inhibit various types of cancer [13]. Multiple mechanisms of TBM have been uncovered recently, including the modulation of multiple enzyme activities and signaling pathways [13]. It is of great interest to reveal novel activities and mechanisms of TBM in cancer treatment. In the present study, we demonstrated that TBM exhibited potent antitumor effect on CRC. TBM inhibited the growth of CRC via inhibition of cell proliferation and induction of apoptosis. Interestingly, we also found that TBM was a potent autophagy modulator in CRC cells. TBM initiates autophagy by activating ROS-AMPK signaling and impairs autophagy flux through inhibiting lysosomal proteolysis activity, leading to the rough accumulation of autophagolysosomes in CRC cells. Meanwhile, our findings indicated that accumulation of immature autophagolysosomes by TBM triggered apoptotic cell death. Our results also provided the support that low concentration of TBM synergizing with 5-FU and DOX shows preferential cytotoxicity compared with treatment alone.
Autophagy is a tightly-regulated catabolic process for clearing damaged or long-lived proteins and organelles, and plays important roles in response to various stresses including cancer treatment [14,15,16]. The role of autophagy in response to chemotherapy is multifaceted [14,15,16]. On one hand, autophagy usually play a supportive role to hamper the anti-cancer effectiveness and induce drug resistance by preserving the organelle function and eliminating cellular waste products [14,15,16]. On the other hand, autophagy also can be pro-death or inhibit proliferation to benefit the anticancer outcome [14,15,16]. In this study, we found that TBM could significantly induce autophagy initiation and promote the fusion of autophagosome with lysosome, while impairs the degradation ability of autophagolysosome, leading to the robust accumulation of autophagolysosomes and autophagic cargo proteins (p62). The inhibition of autophagic flux is harmful to cancer cells in chemotherapy and has potential for clinical benefit. Indeed, inhibition of autophagy initiation could counteract TBM-induced cytotoxicity, while impeding autophagy flux by CQ would further promote cell death slightly in CRC cells, suggesting autophagy account for the anti-cancer activity of TBM in CRC cells.
Accumulating evidence has shown that autophagy can give cancer cells resistant ability to cancer treatment including chemotherapy and radiotherapy, whereas inhibition of autophagy could enhance the efficacy of anticancer agents [14, 24, 25]. Targeting the autophagy process has been regarded as a novel therapeutic approach, and autophagy inhibitors including chloroquine (CQ) and its derivative hydroxychloroquine (HCQ) have been applied in clinic [14, 24, 25]. Thus, development of novel autophagy modulator is worthwhile to be exploited. The classical chemotherapeutic drug 5-FU and DOX have exhibited compromised treatment outcome in CRC treatment due to protective autophagy initiation [26,27,28]. Thus, several efforts have been made to explore new combinational therapies with autophagy inhibitor [26,27,28]. TBM has been reported to enhance the efficacy of many chemotherapeutic agents in cancer cells. For example, TBM has the capability to increase the sensitivity of cisplatin (CDDP)-resistant human ovarian cancer A2780/DDP cells and Hela cells toward CDDP-induced cell death [18, 29], while the combination of TBM with fuzi extracts (Radix Aconiti Lateralis Preparata) exerted synergistic effects on breast cancer MDA-MB-231 and SKBR3 cells proliferation and migration [30]. Therefore, we determined the possibility that TBM addition might sensitive CRC cells to 5-FU or DOX treatment. The present study shows that low-dose TBM in combination 5-FU or DOX exhibit good synthetic lethal effect in CRC cells, supporting that combinational treatment of TBM with conventional anticancer therapies may warrant consideration for clinical trials by modulation of autophagy.
Reactive oxygen species (ROS) is a collective term that encompasses one or more unpaired electrons of oxygen, including superoxide (O2−), hydrogen peroxide (H2O2) and the hydroxyl radical (HO•) [31, 32]. Excessive production of ROS cause biomolecules damage and even cell demise, whereas moderate levels of ROS can act as a second-messenger to regulate pro-survival pathways [31, 32]. ROS have been widely reported to play a pivotal role in apoptosis and autophagy upon cancer treatment [33]. Major sources of cellular ROS are generated from mitochondrial respiration, NADPH oxidases (NOXs), peroxisomal β-oxidation and endoplasmic reticulum (ER) [31, 32]. TBM has been shown to induce mitochondrial damage and ROS generation in several cancer cells including hepatoma HepG2 cells, prostate cancer DU145 cells, lung cancer A549 and PC9 cells, as well as glioma U251 cells, which was involved in TBM-induced cell death [10, 13, 34,35,36]. In this study, we also found that TBM can promote ROS production, which in turn induces autophagy initiation by activating AMPK.
AMPK has been well characterized to play an essential role in the induction of autophagy response to cancer treatment and could be regulated by redox modification under oxidative stress [19, 37, 38]. AMPK can trigger autophagy through regulation of early autophagic events, such as activating the proautophagic VPS34 complex by phosphorylating S91/S94 in beclin1, releasing ULK1 from mTORC1 or directly activating ULK1 by phosphorylation [19, 37, 38]. AMPK is a metabolic checkpoint and regulated by energy supply, and mitochondria are the most important organelles for energy production [19, 37, 38]. TBM induced apoptosis mainly via mitochondrial pathway which will damage the mitochondria and decrease the production of energy. These may lead to AMPK activation and subsequently autophagy initiation to protect cells [13]. Our study reveals that TBM strongly increases Thr172 phosphorylation of AMPK in a dose-dependent manner, which is crucial for the initiation of autophagy in TBM-treated CRC cells.
Conclusions
In summary, we demonstrate that TBM exerts strong cytotoxicity by inducing apoptosis in this study. Moreover, we revealed that TBM could promote impaired autophagolysosomes accumulation by both enhancing autophagy initiation and disrupting the autophagy flux, which is contributed to the anticancer activity of TBM. In addition, as a specific autophagy modulator, TBM could act as one attractive drug in combining with other chemotherapy agents to treat CRC cells for overcoming drug resistance inducing by protective autophagy. Our findings provide a bold therapeutic strategy to potential use of TBM as an adjuvant for further cancer treatment, especially in CRC.
Availability of data and materials
All data and materials can be provided upon request (email: leiyunlong@126.com).
Abbreviations
- 3-MA:
-
3-methyladenine
- 5-FU:
-
5-fluorouracil
- AMPK:
-
Adenosine monophosphate-activated protein kinase
- ATG:
-
Autophagy-related
- Bcl-2:
-
B-cell lymphoma-2
- CDDP:
-
Cisplatin
- CQ:
-
chloroquine
- CRC:
-
Colorectal cancer
- DOX:
-
Doxorubicin
- GFP:
-
Green fluorescent protein
- HCQ:
-
Hydroxychloroquine
- LAMP:
-
Lysosomal-associated membrane protein
- LC3:
-
Microtubule-associated protein 1 light chain 3
- LDH:
-
Lactic dehydrogenase
- NAC:
-
N-acetyl-L-cysteine
- p62:
-
Sequestosome 1
- PARP1:
-
Poly (ADP-ribose) polymerase 1
- RFP:
-
Red fluorescent protein
- ROS:
-
Reactive oxygen species
- siRNA:
-
Small interfering RNA
- TBM:
-
Tubeimoside I
References
Siegel RL, Miller KD, Fedewa SA, Ahnen DJ, Meester RGS, Barzi A, et al. Colorectal cancer statistics, 2017. CA-Cancer J Clin. 2017;67(3):177–93.
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA-Cancer J Clin. 2018;68(6):394–424.
Brenner H, Kloor M, Pox CP. Colorectal cancer. Lancet. 2014;383(9927):1490–502.
Miller KD, Siegel RL, Lin CC, Mariotto AB, Kramer JL, Rowland JH, et al. Cancer treatment and survivorship statistics, 2016. CA-Cancer J Clin. 2016;66(4):271–89.
Schimanski CC, Galle PR, Moehler MH. Is sorafenib effective in colorectal cancer? Definition of resistance indicators and impact of potential comedications. J Clin Oncol. 2011;29(15):1.
Yu L, Ma R, Wang Y, Nishino H. Potent anti-tumor activity and low toxicity of tubeimoside 1 isolated from Bolbostemma paniculatum. Planta Med. 1994;60(3):204–8.
Xu Y, Chiu LF, He QY, Chen F. Tubeimoside-1 exerts cytotoxicity in HeLa cells through mitochondrial dysfunction and endoplasmic reticulum stress pathways. J Proteome Res. 2009;8(3):1585–93.
Chen WJ, Yu C, Yang Z, He JL, Yin J, Liu HZ, et al. Tubeimoside-1 induces G2/M phase arrest and apoptosis in SKOV-3 cells through increase of intracellular Ca2+ and caspase-dependent signaling pathways. Int J Oncol. 2012;40(2):535–43.
Wang F, Ma RD, Yu LJ. Role of mitochondria and mitochondrial cytochrome c in tubeimoside I-mediated apoptosis of human cervical carcinoma HeLa cell line. Cancer Chemother Pharmacol. 2006;57(3):389–99.
Jia G, Wang Q, Wang R, Deng DN, Xue L, Shao NY, et al. Tubeimoside-1 induces glioma apoptosis through regulation of Bax/Bcl-2 and the ROS/cytochrome C/Caspase-3 pathway. OncoTargets Ther. 2015;8:303–9.
Huang P, Yu C, Liu XQ, Ding YB, Wang YX, He JL. Cytotoxicity of tubeimoside I in human choriocarcinoma JEG-3 cells by induction of cytochrome c release and apoptosis via the mitochondrial-related signaling pathway. Int J Mol Med. 2011;28(4):579–87.
Bian Q, Liu PF, Gu JP, Song BJ. Tubeimoside-1 inhibits the growth and invasion of colorectal cancer cells through the Wnt/beta-catenin signaling pathway. Int J Clin Exp Pathol. 2015;8(10):12517–24.
Islam MS, Wang CY, Zheng JY, Paudyal N, Zhu YL, Sun HX. The potential role of tubeimosides in cancer prevention and treatment. Eur J Med Chem. 2019;162:109–21.
Levy JMM, Towers CG, Thorburn A. Targeting autophagy in cancer. Nat Rev Cancer. 2017;17(9):528–42.
White E. The role for autophagy in cancer. J Clin Invest. 2015;125(1):42–6.
Gewirtz DA. The four faces of autophagy: implications for Cancer therapy. Cancer Res. 2014;74(3):647–51.
Jiang S-L, Y-d G, Chen X-S, Ge P, Wang X-L, Lao Y-Z, et al. Tubeimoside-1, a triterpenoid saponin, induces cytoprotective autophagy in human breast cancer cells in vitro via Akt-mediated pathway. Acta Pharmacol Sin. 2018.
Feng X, Zhou J, Li J, Hou X, Li L, Chen Y, et al. Tubeimoside I induces accumulation of impaired autophagolysosome against cervical cancer cells by both initiating autophagy and inhibiting lysosomal function. Cell Death Dis. 2018;9(11):1117.
Xie N, Yuan KF, Zhou L, Wang K, Chen HN, Lei YL, et al. PRKAA/AMPK restricts HBV replication through promotion of autophagic degradation. Autophagy. 2016;12(9):1507–20.
Zhou J, Liu H, Zhang L, Liu X, Zhang C, Wang Y, et al. DJ-1 promotes colorectal cancer progression through activating PLAGL2/Wnt/BMP4 axis. Cell Death Dis. 2018;9(9):865.
Wang S, Wang K, Zhang C, Zhang W, Xu Q, Wang Y, et al. Overaccumulation of p53-mediated autophagy protects against betulinic acid-induced apoptotic cell death in colorectal cancer cells. Cell Death Dis. 2017;8(10):e3087.
Klionsky DJ, Abdelmohsen K, Abe A, Abedin MJ, Abeliovich H, Arozena AA, et al. Guidelines for the use and interpretation of assays for monitoring autophagy (3rd edition). Autophagy 2016, 12(1): 1–222.
Hardie DG. AMP-activated protein kinase-an energy sensor that regulates all aspects of cell function. Genes Dev. 2011;25(18):1895–908.
Sui X, Chen R, Wang Z, Huang Z, Kong N, Zhang M, et al. Autophagy and chemotherapy resistance: a promising therapeutic target for cancer treatment. Cell Death Dis. 2013;4:12.
Lei YY, Zhang D, Yu J, Dong H, Zhang JW, Yang SM. Targeting autophagy in cancer stem cells as an anticancer therapy. Cancer Lett. 2017;393:33–9.
Sasaki K, Tsuno NH, Sunami E, Tsurita G, Kawai K, Okaji Y, et al. Chloroquine potentiates the anti-cancer effect of 5-fluorouracil on colon cancer cells. BMC Cancer. 2010;10:11.
Li J, Hou N, Faried A, Tsutsumi S, Kuwano H. Inhibition of autophagy augments 5-fluorouracil chemotherapy in human colon cancer in vitro and in vivo model. Eur J Cancer. 2010;46(10):1900–9.
Fang LM, Li B, Guan JJ, Xu HD, Shen GH, Gao QG, et al. Transcription factor EB is involved in autophagy-mediated chemoresistance to doxorubicin in human cancer cells. Acta Pharmacol Sin. 2017;38(9):1305–16.
Liu HZ, Yu C, Yang Z, He JL, Chen WJ, Yin J, et al. Tubeimoside I sensitizes cisplatin in cisplatin-resistant human ovarian cancer cells (A2780/DDP) through down-regulation of ERK and up-regulation of p38 signaling pathways. Mol Med Rep. 2011;4(5):985–92.
Chen D, Cao R, He JH, Guo Y, Wang LP, Ji W, et al. Synergetic effects of aqueous extracts of Fuzi (Radix Aconiti Lateralis Preparata) and Tubeimu (Rhizoma Bolbostemmatis) on MDA-MB-231 and SKBR3 cells. J Tradit Chin Med. 2016;36(1):113–24.
Lei YL, Wang K, Deng LF, Chen Y, Nice EC, Huang CH. Redox regulation of inflammation: old elements, a new story. Med Res Rev. 2015;35(2):306–40.
Wang K, Jiang JW, Lei YL, Zhou ST, Wei YQ, Huang CH. Targeting metabolic-redox circuits for Cancer therapy. Trends BiochemSci. 2019;44(5):401–14.
Zhang L, Wang K, Lei YL, Li QF, Nice EC, Huang CH. Redox signaling: potential arbitrator of autophagy and apoptosis in therapeutic response. Free Radic Biol Med. 2015;89:452–65.
Yang JB, Khan M, He YY, Yao M, Li YM, Gao HW, et al. Tubeimoside-1 induces oxidative stress-mediated apoptosis and G (0)/G (1) phase arrest in human prostate carcinoma cells in vitro. Acta Pharmacol Sin. 2016;37(7):950–62.
Hao WL, Wang S, Zhou ZL. Tubeimoside-1 (TBMS1) inhibits lung cancer cell growth and induces cells apoptosis through activation of MAPK-JNK pathway. Int J Clin Exp Pathol. 2015;8(10):12075–83.
Xu Y, Ching YP, Zhou Y, Chiu JF, Chen F, He QY. Multiple pathways were involved in tubeimoside-1-induced cytotoxicity of HeLa cells. J Proteome. 2011;75(2):491–501.
Zhao B, Qiang L, Joseph J, Kalyanaraman B, Viollet B, He Y-Y. Mitochondrial dysfunction activates the AMPK signaling and autophagy to promote cell survival. Genes & diseases. 2016;3(1):82–7.
Kim J, Kundu M, Viollet B, Guan KL. AMPK and mTOR regulate autophagy through direct phosphorylation of Ulk1. Nat Cell Biol. 2011;13(2):132–U171.
Acknowledgements
Not applicable.
Funding
This work was supported by grants from the Chinese NSFC (31400999, 81872014), and Scientific and Technological Research Program of Chongqing Municipal Education Commission (Grant No. KJQN201800417, KJQN201800429).
Author information
Authors and Affiliations
Contributions
Conception and design: YLL, JHY; Acquisition of data: JHY, XYD, JZ, YFX, LM and LHL; Analysis and interpretation of data (e.g., statistical analysis, biostatistics, computational analysis): JHY and JZ; Writing, review, and/or revision of the manuscript: YLL, LHL and JHY; Study supervision: YLL. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
This article does not contain any studies with human subjects and animals performed by any of the authors.
Consent for publication
The authors agree for publication.
Competing interests
Jianghong Yan, Xiaoyun Dou, Jing Zhou, Yuanfeng Xiong, Ling Mo, Longhao Li and Yunlong Lei declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional file
Additional file 1:
Figure S1. TBM promotes apoptosis of CRC cells. Figure S2. ROS-induced impaired autophagolysosomes accumulation contributes to TBM-induced apoptosis. Table S1. List of Small Interference RNA Sequences (PDF 1173 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Yan, J., Dou, X., Zhou, J. et al. Tubeimoside-I sensitizes colorectal cancer cells to chemotherapy by inducing ROS-mediated impaired autophagolysosomes accumulation. J Exp Clin Cancer Res 38, 353 (2019). https://doi.org/10.1186/s13046-019-1355-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13046-019-1355-0