
Abstract
Background
Non-progressive cerebellar ataxia with mental retardation (CANPMR, OMIM 614756) and chromosome 1p32-p31 deletion syndrome (OMIM 613735) are two very rare inherited disorders, which are caused by mono-allelic deficiency (haplo-insufficiency) of calmodulin-binding transcription activator 1 (CAMTA1) and, respectively, nuclear factor 1 A (NFIA) genes. The yet reported patients affected by mono-allelic CAMTA1 dysfunction presented with neonatal hypotonia, delayed and ataxic gait, cerebellar atrophy, psychological delay and speech impairment, while individuals carrying a disrupted NFIA allele suffered from agenesis/hypoplasia of the corpus callosum, ventriculomegaly, developmental delay and urinary tract abnormalities. Both disorders were not seen in one patient together before.
Results
In this study two related individuals affected by a complex clinical syndrome, characterized by cognitive, neurological and nephrological features were studied for the underlying genetic disorder(s) by molecular cytogenetics. The two individuals present dysmorphic facies, macrocephaly, generalized ataxia, mild tremor, strabismus, mild mental retardation and kidney hypoplasia. Moreover, neuro-radiological studies showed hypoplasia of corpus callosum. Genetic investigations revealed a paracentric inversion in the short arm of one chromosome 1 with breakpoints within CAMTA1 and NFIA coding sequences.
Conclusions
To the best of our knowledge, this is the first report of two patients harboring the simultaneous mono-allelic disruptions and consequent haplo-insufficiencies of two genes due to an inversion event. Disruption of CAMTA1 and NFIA genes led to neuro-psychological and nephrological dysfunctions, which comprised clinical features of CANPMR syndrome as well as chromosome 1p32-p31 deletion syndrome.
Similar content being viewed by others


Background
Non-progressive cerebellar ataxia with mental retardation (CANPMR, OMIM 614756) is a very rare neuro-developmental disorder, belonging to the heterogeneous family of genetically determined cerebellar ataxias [1, 2] with recessive [3] and dominant [4] pattern of inheritance. The affected patients present with ataxic gait, dysmetries, variable mental retardation, cerebellar abnormalities and dysmorphic facies with heterogeneous penetrance. To date few genes/loci have been associated to autosomal recessive forms of cerebellar ataxias: ATCAY [5], chromosome 20q11-q13 locus [6], VLDLR [7], ZNF592 [8], SPTBN2 [9], CWF19L1 [10], PMPCA [11]. Calmodulin-binding transcription activator 1 (CAMTA1) maps on chromosome 1p36, carries 23 exons and encodes 2 protein isoforms in mammalians. The brain-specific transcription factor CAMTA1 functions as homodimeric complex binding to gene promoters’ CGCG box thorough CG-1 domain, supporting the assembly of other transcription factors (e.g. Nkx2-5) and enhancing transcription of effector genes (e.g. Fbxl4) [12–14]. CAMTA1 dysfunction has been associated to human pathology by Thevenon et al. [15], who reported five patients affected by CAMTA1 haploinsufficiency due to deletions or duplications in the gene region coding for CG-1 domain, which plays a pivotal role in the whole CAMTA1 function. As mentioned above, the reported patients suffered from ataxia, broad-based gait, tremor, intellectual impairment and speech delay, cerebellar abnormalities (atrophy of lobes and/or vermis) and facial dysmorphisms (strabismus, large forehead, wide and broad nose, small ears).
The clinical features of chromosome 1p32-p31 deletion syndrome (OMIM 613735) were firstly described by Campbell et al. [16]. Some years later, Lu et al. proposed the causal association between this malformation syndrome and Nuclear Factor 1 A (NFIA) haplo-insufficiency [17]. NFIA maps on 1p31.2, carries 11 exons and produces at least 9 different protein isoforms [18–20]. The protein is functionally divided in two sections: a 200 amino acid long N-terminal DNA binding and dimerization domain, mainly encoded by exon 2, and C-terminal transactivation and repression domains, mainly encoded by exons 3 to 11. The first one binds to the nucleotide consensus sequence TTGGC(N)5GCCAA within the promoter region of several genes. The latter ones operate by directly activating basal transcription factors at transcription start sites, by displacing repressive histones from target genes, by interacting with other co-activator proteins [20]. All five individuals reported by Lu et al. (three of which previously described by Campbell et al.) presented with hypoplastic or absent corpus callosum, ventriculomegaly with or without relevant hydrocephalus and development delay; some of them carried urinary tract abnormalities (3 patients), epileptic seizures (3 patients), tethered spinal cord (4 patients) and Chiari malformation (3 patients). Although the chromosomal abnormalities [a translocation t(1;20), a translocation t(1;2), a 2.2 Mb deletion in 1p31-p32, a 12 Mb deletion in 1p31-p32] differed and the deleted regions comprised different genes among the five patients, only NFIA gene was either disrupted or fully deleted in all five patients, thus underpinning the association between NFIA haplo-insufficiency and the common CNS abnormalities (hypoplasia of corpus callosum, ventriculomegaly and hydrocephalus). A strong confirmation on their pathophysiological hypothesis was given by the clinical and histopathological findings of Nfia -/- mouse model [17, 21]. A detailed study of the genome expression profile in murine Nfia -/- brain at embryonic and post-natal stages showed a very strong imbalance in time-related expression of several genes playing a pivotal role in oligodendrocyte differentiation (e.g. Mag, Mal, Mobp, Mog, Sox2, Sox4, Sox11, Dio2, Myef2) as well as in axonal growth/guidance (e.g. Clusterin, aFGF, Ndrg2, EphrinB2, Crmp1) [22]. The essential role of transcription factor Nfib and Nfic in brain, tooth and lung development was already described in the corresponding mouse models [23, 24]. Few further reports described heterogeneous clinical findings associated with deletions mapping on chromosome 1p31 and 1p32 and encompassing several other genes [25, 26].
We describe the first family (Fig. 1) with two related individuals (II.1 and III.1) carrying simultaneous disruptions and consequent haplo-insufficiencies of NFIA and CAMTA genes due to a paracentric inversion in the short arm of chromosome 1 and presenting with clinical signs of CANPMR syndrome as well as chromosome 1p32-p31 deletion syndrome.

Pedigree of the affected family. The index-patient (III.1) and her mother (II.1) presents with the same karyotype
Results and discussion
Patient 1
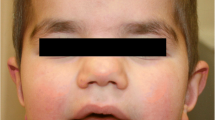
Since the first months of life the index-patient (III.1) presented with generalized hypotonia, reduced muscular strength, particularly evident at trunk und pelvis, normal tendon reflexes, large head (> P90), prominent forehead, strabismus divergens and light bilateral ptosis. The patient could sit at 11 months and walk at 23 months of age. Gowers’ sign was through-out positive. Light dyskinesias and ataxia appeared at 3 years and, respectively, 6 years of age. She could speak the first words at about 30 months of age.
She slowly improved her motor, cognitive and language skills. Now, at the age of 7 years she can walk unassisted, climb stairs and speak short sentences. Her daily skills are compromised and she attends a special care kindergarten. Her intelligence quotient (IQ) score of 51 (Snijders-Oomen Non-Verbal test, SON-R) indicates a severe intellectual impairment (Table 1).
The electroencephalogram (EEG) did not show epileptic discharges. The brain MR investigations at 2 and 6 ½ years of age revealed hypoplasia of corpus callosum, while the cerebellum was structurally normal (Fig. 2). Organ ultrasound did not show any structural abnormalities of kidneys and urinary tract, although the patient suffered from recurrent urine infections starting from 18 months of age.

Brain MR investigation of index-patient (III.1, pictures a, b, c) and her mother (II.1, pictures d, e, f). Hypoplasia of the corpus callosum is revealed in both patients (red arrows). Otherwise no other structural anomalies were observed
Patient 2 and other individuals of pedigree
The mother (II.1) of the index patient (28 years) presents with a similar clinical constellation of III.1, whereby she suffered from neonatal hypotonia, but sat unassisted at 8 months, walked at 14 months and spoke the first words at 12 months of age. She presents with a moderate ataxia, gait instability and dysmetria, which does not strongly impair the daily skills. Her IQ score of 65 (SON-R test) revealed a mental retardation. She has not completed any job training course and is currently unemployed. The brain MR scan, performed at 28 years of age, revealed hypoplasia of corpus callosum (Fig. 2).
Since the early childhood, she suffered from recurrent urinary tract infections due to vesicoureteral reflux (VUR) and from hypoplasia of the right kidney. The father (II.2) of the index patient carries a normal male karyotype and is a healthy individual. The maternal grandparents (I.1 and I.2) are healthy individuals; nevertheless their karyotype could not be analyzed due to missing compliance.
Cytogenetic investigations
The index-patient III.1 harbors a paracentric inversion in the short arm of chromosome 1 with breakpoints within CAMTA1 (1p36.31) and NFIA (1p31.3) genes (Fig. 3).

Karyotype of index-patient (III.1). Karyotyping (GTG-banding) of lymphocytes of peripheral blood revealed an inversion in the short arm of chromosome 1. Breakpoints in 1p31.3 and 1p36.31 (arrows) correspond to two deletions, which have been revealed by SNP array and depicted in Fig. 4
In correspondence of both breakpoints, two deletions occurred affecting CAMTA1 exon 5 (deletion length 211 Kb; position 6,936,272 - 7,146,519 Mb, GRCh37/hg19) and NFIA exons 3-4 (deletion length 217 Kb; position 61,591,640 - 61,807,789 Mb, GRCh37/hg19) in patients II.1 and III1, which could only be resolved by SNP array (Fig. 4).

SNP Array on DNA from peripheral blood leukocytes. a Two deletions are revealed in the short arm of chromosome 1 in both the index-patient (III.1) and her mother (II.1). The distal deletion maps in band 1p36.31 and spans over 211 Kb (b) and the proximal deletion maps in band 1p31.3 and spans over 271 Kb (c). (GRC37h/hg19 genome build)
The resulting genotype is described as [46,XX,inv(1)(p36.31p31.3).arr 1p36.31(6,936,272-7,146,519)x1mat, 1p31.3(61,591,640-61,807,789)x1mat]. Due to the chromosomal inversion, the remaining coding regions of the affected alleles are restricted to exons 1 to 4 for CAMTA1 and, respectively, exons 1 to 2 for NFIA. For CAMTA1, C-terminal 2/3 of CG-1 domain is deleted (about 80 out of 120 amino acids) and the other 3 C-terminal domains (TIG, IQ motif and Ankirin) are fully deleted. For NFIA, the N-terminal DNA binding domain (encoded by exon 2) is not affected by the deletion, but the C-terminal transactivation and repression domains (including also the proline-rich domain) are absent from the remaining coding sequence. The second affected individual II.1, mother of the index-patient, carries the same chromosomal aberration as her daughter, III.1. The individual II.2, father of III.1, does present a normal male karyotype. We were not able to test the karyotype of I.1 and I.2, grandparents of the index patient, due to their missing compliance; nevertheless, they are healthy individuals without any clinical findings resembling the syndromic features of II.1 and III.1.
Given the similar clinical and genetic findings of both patients the chromosomal aberration occurred de novo in patient II.1 at embryonic stage and was transmitted to patient III.1.
Conclusions
To our knowledge, our related patients are the unique two described individuals with combined features from two independent syndromes. CANPMR syndrome (OMIM 614756) is a rare genetic disorder of the nervous system presenting with cerebellar ataxia and mild to severe mental retardation, whose genetic cause was recently described to be CAMTA1 haploinsufficiency [15]. Chromosome 1p32-p31 deletion syndrome (OMIM 613735) presents with hypoplasia/aplasia of the corpus callosum, one of the most common congenital abnormalities of the CNS [27], accompanied by ventriculomegaly with or without hydrocephalus and developmental delay. The haploinsufficiency of NFIA was implied by Lu et al [17] to be the molecular cause of this syndrome.
According to the last insights on the genetic patho-physiology of these two syndromes [15, 17], the complex disorder affecting our two related patients is very likely caused by the simultaneous haplo-insufficiencies of NFIA and CAMTA1, which are the breakpoints of a paracentric inversion in the short arm of chromosome 1. The inversion-associated disruption of NFIA’s and CAMTA1’s reading frames causes very likely the decay of the remaining mRNA strains (corresponding to NFIA exons 1–2 and, respectively CAMTA1 exons 1–4), which are transcribed from the affected alleles of both genes.
If a truncated NFIA protein is synthesized, the C-terminal transactivation and repression domains would completely lack and therefore the truncated NFIA protein would be unable to exert his effector function. The absence of seizures in our two patients may exclude that NFIA haplo-insufficiency causes epileptic seizures, which were described in 3 out of 5 patients reported by Lu et al. On the contrary, the urinary tract abnormalities and VUR in 3 out of 5 patients from Lu et al and in our 2 patients (carrying mono-allelic NFIA disruption) seem to reinforce the view that NFIA plays a major role in the human urinary tract development. Remarkably, our patients carry mono-allelic disruptions of only two genes in comparison with the patients reported by Lu et al. [17] and Koehler et al. [25], which harbored larger deletions encompassing several other genes apart from NFIA (Fig. 5). This fortuitous genetic condition strengthens the casual association between NFIA haplo-insufficiency and some clinical findings described in our study.

Schematic representation of the two affected genes (CAMTA1 and NFIA) in our two related patients (II.1 and III.1) and in other previously reported families and patients. The blue bars (for CAMTA1) and the orange bars and lines (for NFIA) represent deletions, duplications and translation breakpoints affecting the two genes in other reported families (F) and single patients (P). The red bars indicate the two simultaneous deletions disrupting CAMTA1 (1p36.31) and NFIA (1p31.3), which affect our two patients (II.1 and III.1) (scheme based on UCSC genomic bioinformatics, GRC37/hg19)
The pathogenic insights on CAMTA1 haploinsufficiency are enforced by the clinical findings of our two patients. Ataxic and delayed gait, dysmetric movements, instability, macrocephaly and ventriculomegaly, cerebellar and cortical abnormalities at MR scans strengthen the clinical stigmata of CANPMR syndrome. While all former patients reported by Thevenon et al carried intragenic deletions and duplications disrupting the N-terminal CG-1 domain (short deletion of exon 4 in familiy 1, duplication of exons 3 to 5 in family 2, short deletion of exon 3 in familiy 3), the chromosomal inversion harbored by our 2 patients represents the first report on mono-allelic deletion of C-terminal domains TIG, IQ motif and Ankirin from CAMTA1 reading frame and shows the patho-physiological consequence of this genetic event. Among 11 patients from 3 families reported by Thevenon et al, 9 out of 11 patients are reported to have a pathologic intelligence quotient (IQ) between 40 and 67 and a mild to severe intellectual disability (ID), like our 2 patients.
Although an inversion-dependent imbalance on the expression of neighboring genes, located nearby to inversion breakpoints, cannot be theoretically excluded as co-factor responsible for the clinical phenotype, such positional effect seems to be very unlikely in our cases. In fact, no gene mapping in the proximity of deletion 1p36.31 (e.g. DNAJC11, THAP3, PHF13, VAMP3, PER3, UTS2) or in the proximity of the deletion 1p31.3 (e.g. CYP2J2, HOOK1, FGGY, TM2D1, INADL, KANK) was associated with brain and urinary tract abnormalities to date.
Taken together, in our two patients the study of the mono-allelic and simultaneous disruption of genes CAMTA1 and NFIA, whose haploinsufficiencies were already associated to two independent pathological phenotypes [15, 17], sheds further light on the clinical and genetic features of the rare developmental disorders CANPMR syndrome and chromosome 1p32-p31 deletion syndrome.
Methods
Upon extraction of DNA from total peripheral blood leucocytes, conventional karyotyping was performed using G-banding techniques on stimulated blood lymphocytes with standard cytogenetic methods and analyzed at 500–550 band resolution. Karyotypes were described according to the International System for Human Cytogenetic Nomenclature (ISCN 2013).
SNP array was performed using an Infinium CytoSNP-850 K microarray (Illumina, San Diego, CA, USA) with an average resolution of 18Kb and a practical resolution in genes CAMTA1 and NFIA of 1 Kb according to the manufacturer’s protocol. The data analysis was done using BlueFuse V4.2 software. The gene alignment was done using the University of California Santa Cruz (UCSC) genomic bioinformatics browser. The two deleted regions were mapped using the Genome Research Consortium Build 37 human/human genome 19 (GRCh37/hg19). The GenBank, Ensembl and OMIM browser accession numbers for CAMTA1 are NM_015215, ENST00000303635, ENSG00000171735, MIM 611501 and for NFIA are NM_001134673, ENST00000403491, ENSG 00000162599, MIM 600727.
Cerebral MRI was performed on clinical 1.5 T MRI systems (Magnetom Avanto and Aera, Siemens Medical, Germany) using standardized MRI protocols including multiplanar T1 and T2-weighted MR-sequences.
Patient consent
The authors obtained the patient consent for the publication of the data.
Change history
09 March 2021
A Correction to this paper has been published: https://doi.org/10.1186/s13039-021-00534-5
Abbreviations
- CANPMR:
-
cerebellar ataxia non progressive with mental retardation
- SARA:
-
scale for the assessment and rating of ataxia
- SON-R:
-
Snijders-Oomen non-verbal test
References
Steinlin M, Zangger B, Boltshauser E. Non-progressive congenital ataxia with or without cerebellar hypoplasia: a review of 34 subjects. Dev Med Child Neurol. 1998;40:148–54.
Yapici Z, Eraksoy M. Non-progressive congenital ataxia with cerebellar hypoplasia in three families. Acta Paediatr. 2005;94:248–53.
Guzzetta F, Mercuri E, Bonanno S, Longo M, Spano M. Autosomal recessive congenital cerebellar atrophy. A clinical and neuropsychological study. Brain Dev. 1993;15:439–45.
Imamura S, Tachi N, Oya K. Dominantly inherited early-onset non-progressive cerebellar ataxia syndrome. Brain Dev. 1993;15:372–6.
Bomar JM, Benke PJ, Slattery EL, Puttagunta R, Taylor LP, Seong E, et al. Mutations in a novel gene encoding a CRAL-TRIO domain cause human Cayman ataxia and ataxia/dystonia in the jittery mouse. Nat Genet. 2003;35:264–9.
Tranebjaerg L, Teslovich TM, Jones M, Barmada MM, Fagerheim T, Dahl A, et al. Genome-wide homozygosity mapping localizes a gene for autosomal recessive non-progressive infantile ataxia to 20q11-q13. Hum Genet. 2003;113:293–5.
Boycott KM, Flavelle S, Bureau A, Glass HC, Fujiwara TM, Wirrell E, et al. Homozygous deletion of the very low density lipoprotein receptor gene causes autosomal recessive cerebellar hypoplasia with cerebral gyral simplification. Am J Hum Genet. 2005;77:477–83.
Nicolas E, Poitelon Y, Chouery E, Salem N, Levy N, Mégarbané A, et al. CAMOS, a nonprogressive, autosomal recessive, congenital cerebellar ataxia, is caused by a mutant zinc-finger protein, ZNF592. Eur J Hum Genet. 2010;18:1107–13.
Lise S, Clarkson Y, Perkins E, Kwasniewska A, Sadighi Akha E, Schnekenberg RP, et al. Recessive mutations in SPTBN2 implicate β-III spectrin in both cognitive and motor development. PLoS Genet. 2012;8(12):e1003074. doi:10.1371/journal.pgen.1003074.
Burns R, Majczenko K, Xu J, Peng W, Yapici Z, Dowling JJ, et al. Homozygous splice mutation in CWF19L1 in a Turkish family with recessive ataxia syndrome. Neurology. 2014;83:2175–82.
Jobling RK, Assoum M, Gakh O, Blaser S, JRaiman JA, Mignot C, et al. PMPCA mutations cause abnormal mitochondrial protein processing in patients with non-progressive cerebellar ataxia. Brain. 2015;138:1505–17.
Finkler A, Ashery-Padan R, Fromm H. CAMTA1s: calmodulin-binding transcription activators from plants to human. FEBS Lett. 2007;581:3893–8.
Han J, Gong P, Redding K, Mitra M, Guo P, Li HS. The fly CAMTA transcription factor potentiates deactivation of rhodopsin, a G protein-couples light receptor. Cell. 2006;127:847–58.
Gong P, Han L, Redding K, Li HS. A potential dimerization region of dCAMTA is critical termination of fly visual response. J Biol Chem. 2007;282:21253–8.
Thevenon J, Lopez E, Keren B, Heron D, Mignot C, Altuzarra C, et al. Intragenic CAMTA1 rearrangements cause non-progressive congenital ataxia with or without intellectual disability. J Med Genet. 2012;49:400–8.
Campbell CG, Wang H, Hunter GW. Interstitial microdeletion of chromosome 1p in two siblings. Am J Med Genet. 2002;111:289–94.
Lu W, Quintero-Rivera F, Fan Y, Alkuraya FS, Donovan DJ, Xi Q, et al. NFIA haploinsufficiency is associated with a CNS malformation syndrome and urinary tract defects. PLoS Genet. 2007;3:830–43.
Qian F, Kruse U, Lichter P, Sippel AE. Chromosomal localization of the four genes (NFIA, B, C, and X) for the human transcription factor nuclear factor I by FISH. Genomics. 1995;28:66–73.
Grunder A, Qian F, Ebel TT, Mincheva A, Lichter P, Kruse U, et al. Genomic organization, splice products and mouse chromosomal localization of genes for transcription factor Nuclear Factor One. Gene. 2003;304:171–81.
Gronostajski RM. Roles of the NFI/CTF gene family in transcription and development. Gene. 2000;249:31–45.
Das Neves L, Duchala CS, Godinho F, Haxhiu MA, Colmenares C, Macklin WB, et al. Disruption of the murine nuclear factor I-A gene (Nfia) results in perinatal lethality, hydrocephalus, and agenesis of the corpus callosum. PNAS. 1999;96:11946–51.
Wong YW, Schulze C, Streichert T, Gronostajski RM, Schachner M, Tilling T. Gene expression analysis of nuclear factor I-A deficient mice indicates delayed brain maturation. Genome Biol. 2007;8:R72.
Steele-Perkins G, Butz KG, Lyons GE, Zeichner-David M, Kim HJ, Cho MI, et al. Essential role of NFI-C/CTF transcription-replication factor in tooth root development. Mol Cell Biol. 2003;23:1075–84.
Steele-Perkins G, Plachez C, Butz KG, Yang G, Bachurski CJ, Kinsman SL, et al. The transcription factor gene Nfib is essential for both lung maturation and brain development. Mol Cell Biol. 2005;25:685–98.
Koehler U, Holinski-Feder E, Ertl-Wagner B, Kunz J, von Moers A, von Voss H, et al. A novel 1p31.3p32.2 deletion involving the NFIA gene detected by array CGH in a patient with macrocephaly and hypoplasia of the corpus callosum. Eur J Pediatr. 2010;169:463–8.
Nyboe D, Kreiborg S, Kirchhoff M, Hove HB. Familial craniosynostosis associated with a microdeletion involving the NFIA gene. Clin Dysmorphol. 2015;24:109–12.
Paul LK, Brown WS, Adolphs R, Tyszka JM, Richards LJ, Mukherjee P, et al. Agenesis of the corpus callosum: genetics, development and functional aspects of the connectivity. Nat Rev Neurosci. 2007;8:287–99.
Acknowledgements
The authors express our deep gratitude to the family for the full collaboration throughout the project.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
EC, TL, AS, HL, JR participated in the design of the study and analyzed the patients’ phenotype. EC and UK perfomed the genetic studies. CF performed the neuro-radiological investigations. All authors drafted and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Coci, E.G., Koehler, U., Liehr, T. et al. CANPMR syndrome and chromosome 1p32-p31 deletion syndrome coexist in two related individuals affected by simultaneous haplo-insufficiency of CAMTA1 and NFIA genes. Mol Cytogenet 9, 10 (2016). https://doi.org/10.1186/s13039-016-0219-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13039-016-0219-y