
Abstract
Background
Proteolytic degradation of amyloid β (Aβ) peptides has been intensely studied due to the central role of Aβ in Alzheimer’s disease (AD) pathogenesis. While several enzymes have been shown to degrade Aβ peptides, the main pathway of Aβ degradation in vivo is unknown. Cerebrospinal fluid (CSF) Aβ42 is reduced in AD, reflecting aggregation and deposition in the brain, but low CSF Aβ42 is, for unknown reasons, also found in some inflammatory brain disorders such as bacterial meningitis.
Method
Using 18O-labeling mass spectrometry and immune-affinity purification, we examined endogenous proteolytic processing of Aβ in human CSF.
Results
The Aβ peptide profile was stable in CSF samples from healthy controls but in CSF samples from patients with bacterial meningitis, showing increased leukocyte cell count, 18O-labeling mass spectrometry identified proteolytic activities degrading Aβ into several short fragments, including abundant Aβ1–19 and 1–20. After antibiotic treatment, no degradation of Aβ was detected. In vitro experiments located the source of the proteolytic activity to blood components, including leukocytes and erythrocytes, with insulin-degrading enzyme as the likely protease. A recombinant version of the mid-domain anti-Aβ antibody solanezumab was found to inhibit insulin-degrading enzyme-mediated Aβ degradation.
Conclusion
18O labeling-mass spectrometry can be used to detect endogenous proteolytic activity in human CSF. Using this technique, we found an enzymatic activity that was identified as insulin-degrading enzyme that cleaves Aβ in the mid-domain of the peptide, and could be inhibited by a recombinant version of the mid-domain anti-Aβ antibody solanezumab.
Similar content being viewed by others

Background
The significance of proteolytic degradation products of proteins and peptides in the central nervous system (CNS) relate not only to their potential use as biomarkers of disease activity, but also to the understanding of disease mechanisms. Amyloid β (Aβ) peptides are intensely studied due to their central role in Alzheimer’s disease (AD) [18, 19, 26]. Aβ is produced from neurons by sequential cleavage of amyloid precursor protein (APP) by the β- and γ-secretases [5], and released into the interstitial fluid of the brain, which communicates freely with the cerebrospinal fluid (CSF). Aβ is cleared from the brain by several pathways including both extra- and intra- cellular proteolytic degradation, resulting in the production of a large number of different peptides found in brain tissue and CSF [37]. Aβ is also cleared across the blood-brain barrier into the blood, and by CSF absorption into the circulatory or lymphatic system (for review see [49]). While Aβ is effectively degraded in the blood, it is not clear to which extent it is degraded in the interstitial fluid or CSF [24].
AD patients have reduced CSF levels of the 42 amino acid long peptide Aβ42 [6, 36], but low CSF Aβ concentration has also been found in infectious or inflammatory CNS disorders, for example in patients suffering from bacterial meningitis, neuroborreliosis, multiple sclerosis and HIV with CNS engagement [17, 27, 28, 33, 46, 50]. In bacterial meningitis, low CSF Aβ42 normalizes after proper antibiotic treatment, and it has been hypothesized that the decrease in CSF Aβ42 during the acute infection may be caused by impaired clearance of Aβ from the brain, and not related to the inflammation in itself [46]. Another explanation may be increased activities of APP and Aβ metabolizing enzymes with inflammation.
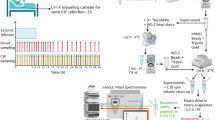
In the current work, we explore for the first time the use of proteolytic 18O labeling and mass spectrometry (MS) to detect endogenous proteolytic activity in human CSF. CSF was collected from patients with bacterial meningitis before and after successful treatment with antibiotics. The samples were incubated with 18O-enriched water and subjected to immunoaffinity purification of Aβ followed by MS to measure the degree of incorporation of 18O into proteolytically produced Aβ peptides, enabling determination of the relative amount of peptides formed by endogenous proteolytic activity in the CSF. Using this approach, we identified products of a specific Aβ-degrading activity in CSF from meningitis patients in the acute phase. This activity, likely derived from blood cell components in the CSF, was identified to arise from insulin-degrading enzyme (IDE), possibly representing a major degradation pathway of Aβ. We also show that this proteolytic activity can be blocked by a recombinant version of the mid-domain anti-Aβ antibody solanezumab that has an epitope covering the discovered cleavage site on Aβ.
Methods
Study subjects
CSF was provided from the Clinical Neurochemistry Laboratory, Mölndal, Sweden. The samples were surplus from the clinical routine, used after de-identification. The samples were taken from patients with neurochemical signs of meningitis including increased CSF cell counts and CSF/serum albumin ratio (see Table 1 for demographics). Blood samples were obtained from healthy volunteers. Erythrocyte concentrate (225–340 ml prepared from 400 to 450 ml whole blood) was provided by the Blood Center at the Sahlgrenska University Hospital. The study was approved by the local Institutional Review Boards and was performed in accordance with the ethical standards laid down in the 1964 Declaration of Helsinki.
Proteolytic 18O-labeling in CSF
The CSF samples (1 ml) were pH-stabilized by addition of 1 M HEPES buffer, pH 7.4 to a final concentration of 100 mM. H2 18O (Sigma) were added to the samples at a 1:1 ratio (v/v), and the samples were incubated for 24 h at room temperature. The samples were stored at −80 °C pending analysis.
Aβ immunoaffinity purification
Aβ peptides were immunoprecipitated using Aβ-specific antibodies coupled to magnetic beads as described previously [38]. Briefly, 4 μg of the anti-Aβ antibodies 6E10 and 4G8 (Signet Laboratories, Dedham, MA, USA) were separately added to 50 μL each of magnetic Dynabeads M-280 Sheep Anti-Mouse IgG (Invitrogen, Carlsbad, CA, USA). The 6E10 and 4G8 antibody-coated beads were mixed and added to the CSF samples to which 0.025% Tween20 in phosphate-buffered saline (pH 7.4) had been added. After washing, the Aβ peptides were eluted using 100 μL 0.5% formic acid.
Mass spectrometry
Mass spectrometry was performed using a matrix-assisted-laser-desorption/ionizationtime-of-flight/time-of-flight (MALDI TOF/TOF) instrument (UltraFleXtreme, Bruker Daltonics, Bremen, Germany). Samples were prepared as described previously [38].
Test of Aβ degrading activity in CSF containing trace amounts of blood
The isotope labeled Aβ peptide Aβ1-40 Arg13C15N (Anaspec, Inc., San Jose, USA) was dissolved in dimethylsulfoxide (DMSO, Aldrich) to a concentration of 1 mg/mL. The stock solution was aliquoted and immediately stored at −20 °C. Before use the labeled peptide was diluted and mixed to a final concentration of 0.8 μM. A 10-μL aliquot of the peptide solution was added to 930 μL CSF to which blood was added at different final concentrations (0, 0.05, 0.5, 5% (v/v)) followed by incubation in room temperature overnight at room temperature. IP-MS was conducted as described above.
In vitro test of Aβ1-40 degradation by IDE
An aliquot (10 μL) of a 0.8 μM solution of the isotopic labeled Aβ1-40 (Arg13C15N) was added to 980 μL CSF and incubated 5 min in room temperature. Recombinant human IDE (R&D systems) was diluted in 50 mM ammonium bicabonate to 23.6 ng/mL and 5.9 ng/mL. 10 μL of each solution was added to two separate pre-incubated CSF samples and left over night in room temperature. Immunoprecipitation and mass spectrometry were conducted as described above.
Inhibition of IDE activity by insulin
The inhibition of IDE activity was addressed by adding isotopic labeled Aβ1-40 (Arg13C15N, end concentration 10 ng/mL) to CSF followed by addition of recombinant human IDE (0.1, 1 or 10 μM, Sigma) and 0.5% human blood. The samples were incubated overnight at room temperature. Immunoprecipitation and mass spectrometry were conducted as described above.
Identification of Aβ degrading activity in leukocytes, thrombocytes, and erythrocytes
Leukocytes and thrombocytes were isolated from 10 mL blood as described previously [13, 35]. The cells were resuspended in 500 μL ultra-pure water, lysed by four freeze/thaw cycles and finally centrifuged 10 min (+4 °C, 31,000 x g). Erythrocytes were prepared using a Reveos automated blood component extractor.
To 100 μL ammonium bicarbonate, spiked with 5 pmol/μL isotopic labeled Aβ1-40, 5 μL leukocytes, thrombocytes, or erythrocytes were added, followed by incubation overnight at room temperature. Immunoprecipitation and mass spectrometry were conducted as described above.
The effect of antibodies on Aβ degradation
To 1 ml PBS containing isotopic labeled Aβ1-40 (0.016 μM) 4 μg of each of the recombinant antibodies solanezumab, bapineuzumab and crenezumab were added [8, 53], followed by incubation for 1 h at room temperature. The antibodies were expressed and purified as previously described [53]. Each sample was spiked with serum (5%, end concentration) followed by incubation overnight at room temperature. The samples were placed in a 100 °C heating block for five minutes to disrupt potential antibody/Aβ complexes, and subsequently centrifuged for 10 min at 4 °C (31.000 x g). The supernatants were transferred to new tubes and Aβ peptides were immunoprecipitated and analyzed by mass spectrometry as described above.
Isotope distribution calculations
Theoretical peptide isotope distributions with and without incorporation of one 18O atom at 50% abundance were calculated using the software Isotope Distribution Calculator v 0.3 [21]. The fractional abundance of Aβ peptides, produced by proteolysis during the incubation period, was calculated by linear regression analysis according to Mirgorodskaya et al. [31], based on the observed isotope distributions of the detected peptides.
Results
Proteolytic 18O labeling detects aberrant Aβ processing in the CSF during acute bacterial meningitis
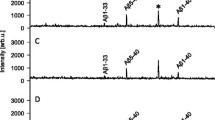
Initial experiments had shown that a patient in the acute phase of bacterial meningitis displayed an abnormal Aβ peptide pattern (Additional file 1: Figure S1). To test for Aβ/APP proteolytic processing in the CSF, H2 18O was added to CSF samples at a volume ratio of 1:1, followed by incubation at room temperature for 24 h. The samples were then subjected to Aβ immunoprecipitation, and analyzed by MALDI-TOF MS [39]. In control samples (Fig. 1a), a normal Aβ peptide pattern was observed, as previously reported [39], and the isotope distributions of the Aβ peptide signals did not differ from samples not incubated with H2 18O (data not shown), indicating that there is no proteolytic activity affecting the Aβ peptides in normal CSF. In contrast, in CSF samples from patients (Table 1) in the acute phase of bacterial meningitis (Fig. 1b), a strong signal at m/z 2461.17 (monoisotopic mass) was present, corresponding to Aβ1-20 as verified by MS/MS (data not shown). In three of the patients the appearance of this signal was accompanied by an apparent decrease in Aβ1-40 (m/z 4328.16). In CSF samples from bacterial meningitis patients taken after antibiotic treatment (Fig. 1c), the normal Aβ peptide pattern was restored. In addition, two unidentified compounds at m/z 3369.45 and 3440.50 were detected in three of the patients in the acute phase. Peaks at these m/z values were also detected in blood-containing samples (see below), indicating blood as their likely source of origin.

MALDI-TOF MS CSF Aβ peptide patterns of a a non-symptomatic control, b Patient 1 in the acute phase of BM and c after antibiotic treatment. The recorded isotope patterns of d Aβ 1–20 and e Aβ 1–40 in acute BM are shown in blue bars, and the expected isotope patterns of the peptides without 18O incorporation and with one oxygen atom exchanged for 18O at 50% abundance is indicated by red and green bars, respectively
Incubation of the samples in H2 18O results in the incorporation of 18O at the C-terminal carboxy group of peptides formed by proteolysis, resulting in a molecular mass shift of +2 or + 4 Da. The mix of labeled/unlabeled peptides resulting from partial labeling and unlabeled peptide formed prior to addition of H2 18O manifests in the mass spectra as a broadening of the natural isotopic peak distribution. In Fig. 1d and e, the red bars represent the theoretical isotope distributions of Aβ1-20 and Aβ1-40 without incorporation of 18O. The green bars indicate the theoretical distributions expected from incorporation of one 18O at 50% abundance, representing full incorporation under the give experimental conditions. By comparing the theoretical isotope distributions with the recorded isotopic peak distributions, the fractional abundance of proteolytically produced Aβ1-20 and 1-40 was calculated by linear regression analysis [31] for all patients before and after treatment (Table 2). In five out of six patients the fractional abundance was close to one, i.e., the recorded isotope distribution of Aβ1-20 corresponded to the calculated isotope distribution of the peptide with exchange of one 16O oxygen atom for 18O at 50% abundance (Fig. 1d), implying that this peptide was mostly formed by proteolysis in CSF during the experiment. In contrast, the fractional abundance of 18O-Aβ1-40 was close to zero, i.e., its isotope distribution indicates no incorporation of 18O (Fig. 1e). Thus the Aβ1-40 peptide was not formed by proteolytic cleavage in the CSF.
Blood contamination in the CSF produces acute bacterial meningitis -like Aβ degradation
Considering the possibility that the proteoloytic activity found in bacterial meningitis CSF represented an enzymatic activity derived from blood we next incubated CSF from a healthy individual, spiked with stable isotope labeled Aβ1-40 Arg13C15N, with different percentages of fresh blood (v/v). The addition of blood produced an Aβ peptide pattern similar to that of patients with acute phase bacterial meningitis, with increased levels of Aβ1-19 and Aβ1-20 (Fig. 2). Degradation of Aβ1-40 was verified by the detection of Aβ1-19 Arg13C15N and Aβ1-20 Arg13C15N from the added stable isotope labeled Aβ1-40 Arg13C15N (Fig. 2c-d). Other proteolytic activity degraded endogenous Aβ1-16 further, as is evident by the decreasing abundance of the peptide signal (Fig. 2, inset). In the sample incubated with 0.5% blood in the CSF (Fig. 2d), endogenous Aβ1-16 was undetectable.

MALDI-TOF mass spectra of the CSF Aβ pattern after incubation with a 0%, b 0.05%, c 0.5% and d 5% human blood. The inset shows the degradation of Aβ1-16, and the vertical dashed line indicates its monoisotopic m/z. * represents an unidentified peak, of three Da higher mass than the monoisotopic peak of Aβ1-16. The Arg13C15N will appear as a mass shift of 10 Da relative the endogenous peptide as seen for Aβ1-19 and Aβ1-20
These proteolytic activities were not dependent on CSF-specific factors, since a similar degradation pattern was observed when replacing CSF with PBS buffer in the experiment above, resulting in proteolysis of the stable isotope labeled Aβ1-40 into similar shorter peptides as observed in CSF (Fig. 3).

Mass spectra of PBS spiked with Aβ1-40 Arg13C15N, incubated overnight in the absence (a) and presence (b) of 0.5% blood. In b, Several Aβ degradation products are detected as well as a cluster of signature peaks from blood in the mass range 3300 – 3600 Da, the identities of which are unknown
The Aβ degrading activity is present in blood components
To pinpoint the origin of the observed proteolytic activity, blood was fractionated into leukocytes, thrombocytes, thrombocyte-free plasma, and erythrocytes. The fractions were spiked with Aβ1-40, incubated as described above, and analyzed by IP-MALDI MS (Fig. 4). While minor degradation was observed in the thrombocyte fraction, the major effect was observed in the leukocyte and erythrocyte fractions where Aβ1-40 was clearly degraded into shorter Aβ peptides, preferentially Aβ1-20. Further experiments showed some Aβ1-40 degradation after incubation with 5% serum but almost no degradation in the presence of plasma (Additional file 2 Figure S2).

Mass spectra of Aβ1-40 proteolytic products after incubation with a leukocytes, b thrombocytes, c plasma without thrombocytes, and d erythrocytes
IDE is a candidate protease for CSF Aβ degradation in acute bacterial meningitis patients
It is known that the protease insulin-degrading enzyme (IDE) cleaves Aβ between residues 19–20 and 20–21 [34]. To test if the observed Aβ degradation was caused by IDE, stable isotope labeled Aβ1-40 Arg13C15N and IDE were added to CSF from healthy individuals, followed by incubation and subsequent IP-MALDI MS analysis. Massive degradation of Aβ was observed, the most prominent cleavage products being Aβ1-19, Aβ1-20 and Aβ1-28 (Fig. 5a, b).

Mass spectra of CSF spiked with Aβ1-40 and its proteolytic products, after incubation with a IDE, and b IDE and insulin. c Scatter plot of Aβ1-20 and Aβ1-40 versus log insulin conc. in PBS spiked with Aβ1-40 and IDE after incubation with insulin at different concentrations
To investigate if the IDE-mediated proteolytic activity could be inhibited, we added increasing amounts of insulin to aliquots of PBS spiked with Aβ1-40 Arg13C15N. As hypothesized, the abundance of Aβ1-20 decreased with increasing insulin concentration, while Aβ1-40 increased (Fig. 5c).
An anti-Aβ antibody blocks degradation of Aβ in serum
It has previously been shown that anti-amyloid treatment using some Aβ-specific monoclonal antibodies, such as the mid-domain anti-Aβ antibody solanezumab [14, 45] and the C-terminal (Aβ X-40) antibody ponezumab [9, 22] increases plasma Aβ concentration up to thousand-fold compared to physiological levels (For review, see [7]). It has been suggested that this increase may be caused by some antibodies blocking the cleavage sites of proteases that normally degrade Aβ [7]. To explore this hypothesis, we tested the effect of recombinant versions of the antibodies solanezumab [14], bapineuzumab [47], and crenezumab [53]. The anti-Aβ antibodies were incubated together with Aβ1-40 in the presence of 5% serum (Fig. 6). While incubation in 5% serum in the absence of antibody gave rise to multiple shorter Aβ peptides including Aβ1-19 and Aβ1-20 (Fig. 6b), the proteolytic degradation was effectively abrogated when the sample was pre-treated with recombinant solanezumab antibody (Fig. 6c). In contrast, incubation in the presence of recombinant crenezumab (Fig. 6d) and bapineuzumab (Fig. 6e) antibodies did not block the degradation of Aβ.

Mass spectra of Aβ1-40 and its proteolytic products in PBS (a), after incubation with 5% serum with b no antibody, c in the presence of Solanezumab, d Crenezumab, and e Bapineuzumab
Discussion
We report on an enzymatic 18O labeling-mass spectrometry assay for determination of the relative amount of the peptides formed by endogenous proteolytic activity. 18O-labeling has previously been used for determination of proteins’ C-termini [41], for the identification of cross-linked peptides in proteolytic peptide mixtures [1], to assist the interpretation of fragment ion mass spectra [43], for generating calibrators for mass spectrometric protein quantification [31], and for differential analysis of protein mixtures [2, 30, 52, 55]. In a recent paper, 18O-labeling was used to monitor proteolytic degradation during sample preparation of mouse brain [48]. Here we show for the first time that 18O-labeling can also be used to detect endogenous proteolytic activity associated with disease. While the current study focuses on the processing of Aβ, the method may be applied to detect proteolytic processing events leading to the formation of any peptide and could thus be a universal tool to assess the integrity of CSF peptides over time; an important aspect in the development of biomarkers.
In bacterial meningitis, activated neutrophils are activated and released. Using 18O-labeling, we found that proteolytic activity in the CSF from patients with bacterial meningitis in the acute phase degrades full-length Aβ peptides into shorter forms, with Aβ1-19, Aβ1-20 and Aβ1-24 being prominent cleavage products. In CSF samples from bacterial meningitis patients taken after antibiotic treatment, the activity was abrogated, also concurring previous data showing that Aβ1-42 in CSF is normally stable over time, even at room temperature [4].
Spiking CSF and solutions containing Aβ with blood produced the similar Aβ fragments as in CSF from patients with bacterial meningitis, suggesting a blood component as the likely source of the observed activity. Similar activity was observed when Aβ1-40 was incubated with serum. The lesser activity observed in plasma likely reflects the presence of EDTA in the collection tubes, which previously has been shown to inhibit IDE proteolytic activity [25]. Further spiking experiments showed that the proteolytic activity resided in leukocytes as well as erythrocytes. IDE activity has previously been associated with leukocytes [42] and erythrocytes [44]. In-vitro experiments have shown that IDE can cleave Aβ between position 19 and 20, and 20 and 21 in the amino acid sequence [29], suggesting it as a possible mediator of the observed activity. We also detected prominent Aβ1-19 and Aβ1-20 signals in CSF samples spiked with IDE (Fig. 5). There are some differences between the results obtained with PBS spiked with blood. For example, a prominent Aβ1-28 signal is present in CSF incubated with IDE but absent in Aβ1-40 incubated with blood. This may be attributed to other proteolytic activities in blood that further degrade Aβ1-28 to shorter Aβ species. Similarly, Aβ1-24 in an Aβ1-40 sample spiked with blood may be the product of other blood-derived proteases.
IDE is a zinc-metallopeptidase which has been implicated in several prevalent diseases including Type 2 diabetes mellitus and AD [12]. IDE has been found to degrade and influence brain Aβ in experimental animal models [3, 23, 32, 54] and it has also been shown to be present in serum [25] and CSF [40]. In the present experiments, no formation of Aβ1-19 or Aβ1-20 was observed in CSF samples from patients taken after antibiotics treatment with normalized leukocyte counts. Furthermore, it has previously been shown that Aβ1-42 in CSF is normally stable over time, even at room temperature [4].
Our results indicate that cleavages at positions Aβ19 and Aβ20 represents a significant pathway for physiological Aβ degradation in blood, and that these cleavages may be mediated by IDE. Since IDE-deficient mice show accumulation of Aβ protein [15], hyperinsulinemia and glucose intolerance enhancing or emulating the activity of IDE may lower the Aβ burden and may be of interest to pursue as an AD-treatment.
The dramatic increase observed in plasma Aβ, in response to treatment with Aβ-specific monoclonal antibodies [14, 45] has been attributed to increased clearance of soluble Aβ from the brain into the periphery. According to the peripheral sink hypothesis, the therapeutic antibody changes the equilibrium for Aβ between the brain and plasma, with increased clearance from the brain and a resulting increase in plasma Aβ levels [11]. However, studies in mouse models, in which the peripheral level of Aβ was decreased by either by reducing production or increasing degradation of Aβ in the periphery, failed to show any effect on Aβ levels in the brain [16, 20, 51]. An alternative hypothesis is that some anti-Aβ antibodies cause a prolonged half-life of Aβ peptides in the bloodstream by blocking protease cleavage sites [7].
Here we show that treatment with a recombinant version of solanezumab protects Aβ from degradation in serum. In contrast, bapineuzumab or crenezumab did not affect the Aβ peptide profiles, with the exception that Aβ1-13 seems to increase. This difference may be due to the much lower affinity of crenezumab [10] and bapineuzumab than solanezumab for Aβ. The pathophysiological relevance of the observations needs to be determined in future studies.
Conclusions
In the present study, we show that a technique based on enzymatic 18O labeling-mass spectrometry is useful for identifying and determining the relative amount of peptides formed by endogenous proteolytic activity in human CSF. Using this technique we found an enzymatic activity in blood leukocytes and erythrocytes that was identified as IDE that cleaves Aβ in the mid-domain of the peptide, and could be inhibited by a recombinant version of the mid-domain anti-Aβ antibody solanezumab. If, as these results suggest, the increase in plasma Aβ upon treatment with some Aβ-specific antibodies is caused by blocking the protease cleavage site, that is that the higher affinity antibody (solanezumab) reduces IDE processing of the peptide, it should be considered that therapeutic antibodies may in fact interfere with Aβ clearance by stabilizing Aβ peptides.
References
Back JW, Notenboom V, de Koning LJ, Muijsers AO, Sixma TK, de Koster CG, et al. Identification of cross-linked peptides for protein interaction studies using mass spectrometry and 18O labeling. Anal Chem. 2002;74(17):4417–22.
Bantscheff M, Dumpelfeld B, Kuster B. Femtomol sensitivity post-digest (18)O labeling for relative quantification of differential protein complex composition. Rapid Commun Mass Spectrom. 2004;18(8):869–76.
Baranello RJ, Bharani KL, Padmaraju V, Chopra N, Lahiri DK, Greig NH, et al. Amyloid-beta protein clearance and degradation (ABCD) pathways and their role in Alzheimer’s disease. Curr Alzheimer Res. 2015;12(1):32–46.
Bjerke M, Portelius E, Minthon L, Wallin A, Anckarsater H, Anckarsater R, et al. “Confounding factors influencing amyloid Beta concentration in cerebrospinal fluid.” Int J Alzheimers Dis. 2010;2010:1–11.
Blennow K, de Leon MJ, Zetterberg H. Alzheimer’s disease. Lancet. 2006;368(9533):387–403.
Blennow K, Hampel H, Weiner M, Zetterberg H. Cerebrospinal fluid and plasma biomarkers in Alzheimer disease. Nat Rev Neurol. 2010;6(3):131–44.
Blennow K, Hampel H, Zetterberg H. Biomarkers in amyloid-beta immunotherapy trials in Alzheimer’s disease. Neuropsychopharmacology. 2014;39(1):189–201.
Bouter Y, Lopez Noguerola JS, Tucholla P, Crespi GA, Parker MW, Wiltfang J, et al. Abeta targets of the biosimilar antibodies of Bapineuzumab, Crenezumab, Solanezumab in comparison to an antibody against Ntruncated Abeta in sporadic Alzheimer disease cases and mouse models. Acta Neuropathol. 2015;130(5):713–29.
Burstein AH, Zhao Q, Ross J, Styren S, Landen JW, Ma WW, et al. Safety and pharmacology of ponezumab (PF-04360365) after a single 10-minute intravenous infusion in subjects with mild to moderate Alzheimer disease. Clin Neuropharmacol. 2013;36(1):8–13.
Crespi GA, Hermans SJ, Parker MW, Miles LA. Molecular basis for mid-region amyloid-beta capture by leading Alzheimer’s disease immunotherapies. Sci Rep. 2015;5:9649.
DeMattos RB, Bales KR, Cummins DJ, Dodart JC, Paul SM, Holtzman DM. Peripheral anti-A beta antibody alters CNS and plasma A beta clearance and decreases brain A beta burden in a mouse model of Alzheimer’s disease. Proc Natl Acad Sci U S A. 2001;98(15):8850–5.
Duckworth WC, Bennett RG, Hamel FG. Insulin degradation: progress and potential. Endocr Rev. 1998;19(5):608–24.
Ekman R, Gobom J, Persson R, Mecocci P, Nilsson CL. Arginine vasopressin in the cytoplasm and nuclear fraction of lymphocytes from healthy donors and patients with depression or schizophrenia. Peptides. 2001;22(1):67–72.
Farlow M, Arnold SE, van Dyck CH, Aisen PS, Snider BJ, Porsteinsson AP, et al. Safety and biomarker effects of solanezumab in patients with Alzheimer’s disease. Alzheimers Dement. 2012;8(4):261–71.
Farris W, Mansourian S, Chang Y, Lindsley L, Eckman EA, Frosch MP, et al. Insulin-degrading enzyme regulates the levels of insulin, amyloid beta-protein, and the beta-amyloid precursor protein intracellular domain in vivo. Proc Natl Acad Sci U S A. 2003;100(7):4162–7.
Georgievska B, Gustavsson S, Lundkvist J, Neelissen J, Eketjall S, Ramberg V, et al. Revisiting the peripheral sink hypothesis: inhibiting BACE1 activity in the periphery does not alter beta-amyloid levels in the CNS. J Neurochem. 2015;132(4):477–86.
Gisslen M, Krut J, Andreasson U, Blennow K, Cinque P, Brew BJ, et al. Amyloid and tau cerebrospinal fluid biomarkers in HIV infection. BMC Neurol. 2009;9:63.
Glenner GG, Wong CW. Alzheimer’s disease: initial report of the purification and characterization of a novel cerebrovascular amyloid protein. Biochem Biophys Res Commun. 1984;120(3):885–90.
Hardy J, Selkoe DJ. The amyloid hypothesis of Alzheimer’s disease: progress and problems on the road to therapeutics. Science. 2002;297(5580):353–6.
Henderson SJ, Andersson C, Narwal R, Janson J, Goldschmidt TJ, Appelkvist P, et al. Sustained peripheral depletion of amyloid-beta with a novel form of neprilysin does not affect central levels of amyloid-beta. Brain. 2014;137(Pt 2):553–64.
Kubinyi H. Calculation of isotope distributions in mass-spectrometry - a trivial solution for a nontrivial problem. Anal Chim Acta. 1991;247(1):107–19.
Landen JW, Zhao Q, Cohen S, Borrie M, Woodward M, Billing Jr CB, et al. Safety and pharmacology of a single intravenous dose of ponezumab in subjects with mild-to-moderate Alzheimer disease: a phase I, randomized, placebo-controlled, double-blind, dose-escalation study. Clin Neuropharmacol. 2013;36(1):14–23.
Leissring MA, Farris W, Chang AY, Walsh DM, Wu X, Sun X, et al. Enhanced proteolysis of beta-amyloid in APP transgenic mice prevents plaque formation, secondary pathology, and premature death. Neuron. 2003;40(6):1087–93.
Leissring MA, Turner AJ. Regulation of distinct pools of amyloid beta-protein by multiple cellular proteases. Alzheimers Res Ther. 2013;5(4):37.
Liu Z, Zhu H, Fang GG, Walsh K, Mwamburi M, Wolozin B, et al. Characterization of insulin degrading enzyme and other amyloid-beta degrading proteases in human serum: a role in Alzheimer’s disease? J Alzheimers Dis. 2012;29(2):329–40.
Masters CL, Simms G, Weinman NA, Multhaup G, McDonald BL, Beyreuther K. Amyloid plaque core protein in Alzheimer disease and Down syndrome. Proc Natl Acad Sci U S A. 1985;82(12):4245–9.
Mattsson N, Axelsson M, Haghighi S, Malmestrom C, Wu G, Anckarsater R, et al. Reduced cerebrospinal fluid BACE1 activity in multiple sclerosis. Mult Scler. 2009;15(4):448–54.
Mattsson N, Bremell D, Anckarsater R, Blennow K, Anckarsater H, Zetterberg H, et al. Neuroinflammation in Lyme neuroborreliosis affects amyloid metabolism. BMC Neurol. 2010;10:51.
McDermott JR, Gibson AM. Degradation of Alzheimer’s beta-amyloid protein by human and rat brain peptidases: involvement of insulin-degrading enzyme. Neurochem Res. 1997;22(1):49–56.
Mirgorodskaya E, Wanker E, Otto A, Lehrach H, Gobom J. Method for qualitative comparisons of protein mixtures based on enzyme-catalyzed stable-isotope incorporation. J Proteome Res. 2005;4(6):2109–16.
Mirgorodskaya OA, Kozmin YP, Titov MI, Korner R, Sonksen CP, Roepstorff P. Quantitation of peptides and proteins by matrix-assisted laser desorption/ionization mass spectrometry using (18)O-labeled internal standards. Rapid Commun Mass Spectrom. 2000;14(14):1226–32.
Moore KM, Girens RE, Larson SK, Jones MR, Restivo JL, Holtzman DM, et al. A spectrum of exercise training reduces soluble Abeta in a dose-dependent manner in a mouse model of Alzheimer’s disease. Neurobiol Dis. 2016;85:218–24.
Mori F, Rossi S, Sancesario G, Codeca C, Mataluni G, Monteleone F, et al. Cognitive and cortical plasticity deficits correlate with altered amyloid-beta CSF levels in multiple sclerosis. Neuropsychopharmacology. 2011;36(3):559–68.
Nalivaeva NN, Beckett C, Belyaev ND, Turner AJ. Are amyloid-degrading enzymes viable therapeutic targets in Alzheimer’s disease? J Neurochem. 2012;120 Suppl 1:167–85.
Nilsson C, Karlsson G, Blennow K, Heilig M, Ekman R. Differences in the neuropeptide Y-like immunoreactivity of the plasma and platelets of human volunteers and depressed patients. Peptides. 1996;17(3):359–62.
Olsson B, Lautner R, Andreasson U, Ohrfelt A, Portelius E, Bjerke M, et al. “CSF and blood biomarkers for the diagnosis of Alzheimer’s disease: a systematic review and meta-analysis.” Lancet Neurol. 2016;15:673–684.
Portelius E, Mattsson N, Andreasson U, Blennow K, Zetterberg H. Novel abeta isoforms in Alzheimer’s disease - their role in diagnosis and treatment. Curr Pharm Des. 2011;17(25):2594–602.
Portelius E, Tran AJ, Andreasson U, Persson R, Brinkmalm G, Zetterberg H, et al. Characterization of amyloid beta peptides in cerebrospinal fluid by an automated immunoprecipitation procedure followed by mass spectrometry. J Proteome Res. 2007;6(11):4433–9.
Portelius E, Westman-Brinkmalm A, Zetterberg H, Blennow K. Determination of beta-amyloid peptide signatures in cerebrospinal fluid using immunoprecipitation-mass spectrometry. J Proteome Res. 2006;5(4):1010–6.
Qiu WQ, Walsh DM, Ye Z, Vekrellis K, Zhang J, Podlisny MB, et al. Insulin-degrading enzyme regulates extracellular levels of amyloid beta-protein by degradation. J Biol Chem. 1998;273(49):32730–8.
Rose K, Simona MG, Offord RE, Prior CP, Otto B, Thatcher DR. A new mass-spectrometric C-terminal sequencing technique finds a similarity between gamma-interferon and alpha 2-interferon and identifies a proteolytically clipped gamma-interferon that retains full antiviral activity. Biochem J. 1983;215(2):273–7.
Roth RA, Mesirow ML, Cassell DJ, Yokono K, Baba S. Characterization of an insulin degrading enzyme from cultured human lymphocytes. Diabetes Res Clin Pract. 1985;1(1):31–9.
Schnolzer M, Jedrzejewski P, Lehmann WD. Protease-catalyzed incorporation of 18O into peptide fragments and its application for protein sequencing by electrospray and matrix-assisted laser desorption/ionization mass spectrometry. Electrophoresis. 1996;17(5):945–53.
Shii K, Yokono K, Baba S, Roth RA. Purification and characterization of insulin-degrading enzyme from human erythrocytes. Diabetes. 1986;35(6):675–83.
Siemers ER, Friedrich S, Dean RA, Gonzales CR, Farlow MR, Paul SM, et al. Safety and changes in plasma and cerebrospinal fluid amyloid beta after a single administration of an amyloid beta monoclonal antibody in subjects with Alzheimer disease. Clin Neuropharmacol. 2010;33(2):67–73.
Sjogren M, Gisslen M, Vanmechelen E, Blennow K. Low cerebrospinal fluid beta-amyloid 42 in patients with acute bacterial meningitis and normalization after treatment. Neurosci Lett. 2001;314(1–2):33–6.
Sperling R, Salloway S, Brooks DJ, Tampieri D, Barakos J, Fox NC, et al. Amyloid-related imaging abnormalities in patients with Alzheimer’s disease treated with bapineuzumab: a retrospective analysis. Lancet Neurol. 2012;11(3):241–9.
Stingl C, Soderquist M, Karlsson O, Boren M, Luider TM. Uncovering effects of ex vivo protease activity during proteomics and peptidomics sample extraction in rat brain tissue by oxygen-18 labeling. J Proteome Res. 2014;13(6):2807–17.
Tarasoff-Conway JM, Carare RO, Osorio RS, Glodzik L, Butler T, Fieremans E, et al. Clearance systems in the brain-implications for Alzheimer disease. Nat Rev Neurol. 2015;11(8):457–70.
Trysberg E, Hoglund K, Svenungsson E, Blennow K, Tarkowski A. Decreased levels of soluble amyloid beta-protein precursor and beta-amyloid protein in cerebrospinal fluid of patients with systemic lupus erythematosus. Arthritis Res Ther. 2004;6(2):R129–136.
Walker JR, Pacoma R, Watson J, Ou W, Alves J, Mason DE, et al. Enhanced proteolytic clearance of plasma Abeta by peripherally administered neprilysin does not result in reduced levels of brain Abeta in mice. J Neurosci. 2013;33(6):2457–64.
Wang YK, Ma Z, Quinn DF, Fu EW. Inverse 18O labeling mass spectrometry for the rapid identification of marker/target proteins. Anal Chem. 2001;73(15):3742–50.
Watt AD, Crespi GA, Down RA, Ascher DB, Gunn A, Perez KA, et al. Do current therapeutic anti-Abeta antibodies for Alzheimer’s disease engage the target? Acta Neuropathol. 2014;127(6):803–10.
Vingtdeux V, Chandakkar P, Zhao H, Blanc L, Ruiz S, Marambaud P. CALHM1 ion channel elicits amyloid-beta clearance by insulin-degrading enzyme in cell lines and in vivo in the mouse brain. J Cell Sci. 2015;128(13):2330–8.
Yao X, Freas A, Ramirez J, Demirev PA, Fenselau C. Proteolytic 18O labeling for comparative proteomics: model studies with two serotypes of adenovirus. Anal Chem. 2001;73(13):2836–42.
Acknowledgements
The authors would like to thank Anni Westerlund and Kristin Augutis for technical assistance.
Funding
This work was supported by funding from The Swedish Research Council, Wallström’s and Sjöbloms Foundation, Gun and Bertil Stohne’s Foundation, Magnus Bergwall’s Foundation, Demensförbundet, Stiftelsen för Gamla tjänarinnor, a National Health and Medical Research Council of Australia (NHMRC) Project Grant (APP1021935) and grants from the JO & JR Wicking Trust, The Mason Foundation and The Bethlehem Griffith Research Foundation to. Infrastructure support to St. Vincent’s Institute from the NHMRC Independent Research Institutes Infrastructure Support Scheme and the Victorian State Government Operational Infrastructure Support Program are gratefully acknowledged. MWP is an NHMRC Research Fellow.
Availability of data and material
All data generated or analysed during this study are included in this published article and its supplementary information files.
Authors’ contributions
JG conceived of the isotopic labeling strategy, contributed to planning the study, performing the experiments and interpreting data, and writing the manuscript. EP contributed to planning the study, data interpretation and writing the manuscript. HZ contributed to data interpretation and writing the manuscript. KB contributed to planning the study, data interpretation and writing the manuscript. NM contributed to planning the study, data interpretation and writing the manuscript. MG selected samples for the study and contributed to writing the manuscript. GANC participated in preparing and sequencing the recombinant biosimilar monoclonal antibodies solanezumab, bapineuzumab and crenezumab, and contributed to writing the manuscript. MWP participated in preparing and sequencing the recombinant biosimilar monoclonal antibodies solanezumab, bapineuzumab and crenezumab, and contributed to writing the manuscript. LAM participated in preparing and sequencing the recombinant biosimilar monoclonal antibodies solanezumab, bapineuzumab and crenezumab, and contributed to writing the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
The manuscript does not contain any individual’s data. All patient samples were de-identified.
Ethics approval and consent to participate
Surplus CSF from patients undergoing clinical evaluation was used after de-identification, as approved by the regional ethics committee at the University of Gothenburg.
Author information
Authors and Affiliations
Corresponding author
Additional files
Additional file 1: Figure S1.
MALDI-TOF MS CSF Aβ peptide patterns of a patient (A) in the acute phase of BM and (B) after antibiotic treatment. (PPTX 74 kb)
Additional file 2: Figure S2.
Effect of serum and plasma on Aβ degradation. PBS spiked with (A) Aβ1-40 Arg13C15N and 5% serum and (B) Aβ1-40 Arg13C15N and 5% plasma. (PPTX 58 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Portelius, E., Mattsson, N., Pannee, J. et al. Ex vivo 18O-labeling mass spectrometry identifies a peripheral amyloid β clearance pathway. Mol Neurodegeneration 12, 18 (2017). https://doi.org/10.1186/s13024-017-0152-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13024-017-0152-5