
Abstract
Background
N6-methyladenosine (m6A) modification is the most prevalent internal modification of eukaryotic mRNA modulating gene expression. m6A modification is a dynamic reversible process regulated by three protein groups: methyltransferases (writers), demethylases (erasers), and m6A-binding proteins (readers). m6A modification is involved in all phases of RNA metabolism, including RNA folding, stability, splicing, nuclear exporting, translational modulation and degradation.
Main body
In recent years, numerous studies have reported that abnormal m6A modification causes aberrant expression of important viral genes. Herein, we review the role of m6A in viral lifecycle and its contribution to the pathogenesis of human diseases. Particularly, we focus on the viruses associated with human diseases such as HIV-1, IAV, HBV, HCV, EBV and many others.
Conclusions
A better understanding of m6A-virus relationship would provide new insights into the viral replication process and pathogenesis of human diseases caused by viruses. In addition, exploration of the role of m6A in disease-causing viruses will reveal novel approaches for the treatment of such diseases.
Similar content being viewed by others
Background
Like histone and DNA, RNA undergoes covalent modifications that fine-tunes its function [1]. Post-transcriptional modification of RNA is very common in eukaryotes, with more than one hundred types of RNA modification reported so far [2]. Most of these modifications occur on ribosomal RNA (rRNA) and transfer RNA (tRNA), which modulate RNA structures, functions, translation, as these RNAs are accessory molecules in eukaryotic processes [2, 3]. Messenger RNA (mRNA), a primary information-bearing molecule, is also post-transcriptionally modified [2]. As early as 1970s, researchers discovered that N6-methyladenosine (m6A) is present on cellular mRNA [4]. Since then, m6A modification has been identified in many mRNA, with 25% of all cellular mRNAs containing multiple m6A residues [5, 6]. Furthermore, it has been found that m6A modification mostly occur at RRACH motif (R = A or G, H = A, C, or U), and m6A sites are significantly clustered around transcription start sites, exonic regions flanking splicing sites, stop codons, 5’untranslated region (5’UTR) and 3’untranslated region (3’UTR) [7,8,9].
m6A modification is dynamically and reversibly regulated by methyltransferases (writers) [10, 11], and removed by demethylases (erasers) [12]; in addition, it functions by interacting with m6A binding proteins (readers), or by indirectly altering the structure of modified RNA to regulate RNA reader–protein interactions [6]. It has been demonstrated that m6A modification and its related enzymes play an important role in different phases of mRNA life [13].
Writers induce m6A modification, which is mediated by methyltransferase-like 3 (METTL3), methyltransferase-like 14 (METTL14), Wilms’ tumor 1-associating protein (WTAP), methyltransferase-like 16 (METTL16), RNA binding motif protein 15 (RBM15), and KIAA1429 [10, 14,15,16]. Bokar et al. discovered a ~ 200 kDa methyltransferase (MT) complex in nuclear extract lysate from HeLa cell which exhibited methyltransferase activity, from which a 70 kDa protein was identified, named MT-A70 or METTL3 [17]. Recently, Liu et al. found that METTL14, an RNA writer, forms a stable heterodimer core complex with METTL3 [10]. This complex component facilitates m6A deposition on nuclear RNA in mammalian cells. Furthermore, the combination of METTL3 and METTL14 dramatically enhanced methyltransferase activity compared to single protein. In addition, WTAP is a regulatory subunit in the m6A methyltransferase complex. Although it lacked methylation activity, it interacted with METTL3/14 complex to influence the deposition of m6A. Moreover, Ping and colleagues showed that WTAP might play a critical role in epitranscriptomic regulation of RNA metabolism [11]. Besides WTAP, RBM15 and KIAA1429, which are part of the complex, affected the activity of METTL3/14 [18]. Schwartz et al. showed that depletion of KIAA1429 decreased m6A level in vitro [19]. Moreover, knockdown of RBM15 caused a significant reduction of m6A deposition on mRNAs [15]. This means RBM15 and KIAA1429 might be involved in methylation process. A recent study [14] identified a new component of writers METTL16, which is independent from the METTL3/METTL14/WTAP complex, that caused m6A deposition on U6 snRNA [20] and U6-like hairpins of MAT2A mRNA [21] in a C-m6A-G context.
It has been demonstrated that m6A modification on RNA can be removed by at least two erasers, alkB homologue 5 (ALKBH5) or the fat mass and obesity-associated (FTO) proteins. Generally, ALKBH5 and FTO are mainly localized in the nuclear compartment [12, 16]. FTO is homologous to the DNA repair protein AlkB, and is involved in oxidative demethylation of 3-methylthymine in single-stranded DNA and in the 3-methyluracil activity in single-stranded RNA [22]. Several studies have reported that FTO is a potent regulator of nuclear mRNA processing events, participating in alternative splicing and processing of the 3′ end mRNA [23]. Additionally, as a demethylase, FTO manipulates the level of N6,2′-O-dimethyladenosine (m6Am); as depletion or overexpression of FTO selectively regulated the abundance of mRNAs containing m6Am in cells. Moreover, FTO reduced the stability of m6Am mRNAs, thus preferentially demethylated m6Am rather than m6A [24]. Zheng et al. demonstrated that ALKBH5 contributed to the removal of m6A modification from nuclear RNA (mostly mRNA) both in vitro and in vivo [16]. Furthermore, ALKBH5 significantly regulated the nuclear RNA export, metabolism and gene expression, indicating that reversible m6A modification on RNA has broad biological effects.
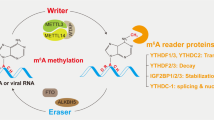
Proteins that selectively bind m6A sites are defined as m6A “readers”, which exert regulation by influencing the recognition of methylated RNA. It has been established that the YTH N6-methyladenosine RNA-binding protein family comprising YTHDF1, YTHDF2, YTHDF3, YTHDC1, and YTHDC2 is the major protein family among all “readers”. In general, YTHDF1 directly enhanced translation through binding with the m6A modification in 3’UTR region [25]; YTHDF2 recruited CCR4-NOT de-adenosine complexes to promote mRNA decay [26]; YTHDF3 acted as a helper of YTHDF1 and YTHDF2 [27, 28]. YTHDF1, 2, and 3 are dominantly located in the cytoplasm, while YTHDC1 is mainly localized in the nucleus where it regulates mRNA export from nucleus to cytoplasm and induce exon inclusion [29, 30]. YTHDC2 modulates the translation of RNA by interacting with the 40-80S subunit in the cytoplasm [31]. Recently, Huang et al. identified a new family- the insulin-like growth factor 2 mRNA-binding proteins (IGF2BPs, including IGF2BP1/2/3) [32]. In contrast to YTH domain family, IGF2BPs bind to m6A-modified mRNAs through GG(m6A)C, a typical m6A motif, to promote mRNA stability. Moreover, other proteins have been found to recognize m6A, such as eIF3 and HNRNP2AB1. eIF3, as part of the 43S pre-translational initiation complex, promotes protein translation by binding to the m6A site of mRNA 5’UTR region [24, 33, 34]. Therefore, m6A is dynamically regulated by writers, erasers, readers and other potential proteins associated with these regulators (Fig. 1).

A schematic model that illustrates the roles and regulation of m6A modification. m6A modification is regulated by “writers” “erasers” and “readers”. Writers and erasers are mainly located in the nucleus, where they modulate mRNAs splicing and nuclear export. YTHDC1 and HNRP2AB1 as nuclear readers also play a vital role in nuclear export. In the cytoplasm, YTHDF2 promotes degradation of mRNA; whereas YTHDF1/3, YTHDC2 and eIF3 participate in translation
Main text
Accumulating evidence shows that m6A modification not only exists in eukaryotic cells but also in viruses. Currently, it is known that deregulation of m6A modification is associated with diseases caused by pathogenic viruses. m6A has long been identified in RNA transcripts of viruses and it regulates nuclear replication in viruses such as influenza A virus, simian virus 40, Rous sarcoma virus, avian sarcoma virus, and adenovirus [35,36,37,38]. In recent years, numerous studies have revealed that m6A modification regulates viral life cycles and m6A modification in pathogenic viruses is increasingly being investigated. Other scholars have revealed that m6A modification influences the expression of key genes involved in the viral life. In addition, m6A modification plays a role in the inhibition or promotion of different types of pathogenic viruses (Table 1). Therefore, exploring the biological functions of m6A methylation in different viruses is of great significance to understanding the pathogenesis and innovative prevention of viral infection. Relationships between m6A and viruses, in relation to diseases caused by different types of viruses are reviewed in following sections.
Human immunodeficiency virus I related diseases
Human immunodeficiency virus I (HIV-1) is the prototype member of the retroviral lentiviruses family and is the etiologic agent of acquired immunodeficiency syndrome (AIDS). People infected with HIV-1 require long-term antiviral treatment, and interruption of the treatment triggers rapid rebound viremia [54]. Therapies for viral infections include vaccines and other drugs. So far, no completely effective treatments have not been developed. Further investigations are required to seek more effective and safe treatments for this disease.
Investigations into m6A modification on HIV have lagged behind other known viruses such as influenza virus, adenovirus, Rous sarcoma virus, and simian virus 40 for almost 40 years [35,36,37,38].Recently, Lichinchi et al. reported the presence of m6A modification on HIV-1 RNA, and characterized the molecular features, topology and function of the viral-host RNA m6A during CD4+T cell infection [39]. They used HIV infected CD4+T cells as experimental subjects to analyze their RNA by methylated RNA immunoprecipitation sequencing (MeRIP-seq). Data showed that HIV-1 infection increased m6A modification level both in host and viral mRNAs compared with the un-infected cells. Knockdown of METTL3/14 suppressed virus replication, while knocking down of ALKBH5 produced opposite results. This indicates that m6A promotes replication of HIV. Subsequently, they discovered that m6A enrichment region was mainly located at the 5’UTR and 3’UTR of HIV genomic RNA, this finding was consistent with that of Tirumuru et al (their RNA samples were isolated from T-cells or primary CD4+T-cells infected with replication-competent HIV-1NL4–3 and analyzed with high-throughput RNA sequencing (m6A-seq)) [40]. However, Kennedy et al. used photo-crosslinking-assisted m6A sequencing (PA-m6A-seq) technique to analyze the RNA samples isolated from infected human CD4+CEM-SS T-cells. They found the enrichment of m6A was merely located at the 3′UTR of HIV-genomic RNA [1]. The discrepancies between these studies might stem from the different mapping techniques used or cell types used for RNA isolation.
Besides writers and erasers, the studies mentioned above revealed that three YTHDF proteins could interact with HIV methylated RNA, but only two of these promoted HIV replication [1, 39]. Kennedy et al. confirmed that overexpression of YTHDF1/3 enhanced the expression of HIV-1 at mRNA and protein level in HIV infected HEK-293 T cells, whereas YTHDF2 did not influence viral replication and protein expression in CEM-SS T cells. In contrast, Tirumuru et al. found that YTHDFs inhabits HIV replication. The mechanisms for the differences among these studies warrants further research. Elsewhere, Lichinchi et al. confirmed that m6A modification existed in 56 host genes from infected T cells using MeRIP-seq. The 56 host genes were involved in viral replication, implying that m6A modification may enhance viral replication by regulating the expression of the 56 host genes.
Rev response element (RRE) is a well-characterized structural and functional RNA element within HIV-1 env gene. After translation, Rev. proteins are imported back into the nucleus where they assemble at the RRE to form an active nuclear export complex to facilitate the transit of viral transcripts into cytoplasm [55]. This process is an essential step for viral replication. Lichinchi et al. found that m6A modification on viral RNA positively affected the interaction between HIV Rev. protein and RRE RNA. This promoted the formation of Rev–RRE complexes and the nuclear export of viral RNA, and hence viral replication. Overall, the links between m6A-related proteins and HIV-1 need to be clarified to identify new mechanisms modulating HIV-1 replication and interaction with the host immune system.
Enterovirus 71 related diseases
Enterovirus 71 (EV71) is a single-stranded retrovirus with a genome of approximately 7.5 kb. EV71 belongs to the genus enterovirus in the Picornaviridae family, which contains three genotypes (A, B and C) and other sub-genotypes. EV71 infections are common in young children, and is one of the etiologic agents of hand, foot, and mouth disease (HFMD) characterized with fever, sores in mouth and a rash with blisters [56]. Since the discovery of EV71 in 1969, innumerable outbreaks and epidemics have been reported worldwide, especially in Asia and Pacific regions, such as China, Korea, Singapore, Japan and Vietnam [57]. Unfortunately, there are no effective antiviral drugs for diseases caused by EV71 infections.
With the rapid development of sequencing technology, several discoveries have been made. For instance, Hao et al. found that EV71 RNA also contains m6A modification in the coding region of VP, 3D and 2C. Specifically, they found that the expression of METTL3, METTL14, YTHDF1–3 and YTHDC1 was increased when the host was infected with EV71, whereas the expression of FTO was decreased [41]. In addition, the expression of m6A regulatory gene affected viral replication. Similarly, mutation in m6A modification sites were found to decrease the production of EV71 progeny virus and protein expression in the infectious clones. Moreover, METTL3 silencing decreased the virus titers and RNA copies, but knockdown of FTO produced opposite results. These results indicate that METTL3 and FTO affect the replication of EV71, suggesting the m6A modification may positively regulate EV71 lifecycle. Meanwhile, knockdown of YTHDF2 and YTHDF3 in Vero cells inhibited viral replication. A co-immunoprecipitation (co-IP) experiment revealed that METTL3 could interact with viral RNA-dependent RNA polymerase (RdRp) 3D, and overexpression of METTL3 increased this protein and contributed to sumoylation and ubiquitination of the 3D polymerase during viral replication. METTL3 functioned as a positive regulator of EV71 replication in the report by Hao and colleagues. Whether METTL3 should be used as a new therapeutic target remains to be elucidated.
Influenza a virus related diseases
Influenza A virus (IAV) is a major cause of upper and lower respiratory tract infections, posing a huge threat to human health. Currently, influenza prevention and treatment strategies include annual vaccination and antiviral drugs [58]. However, frequent antigens variation on viral surface was caused by antigenic shift and drift, allowing influenza viruses to escape antibody-mediated immunity following vaccination [59]. Therefore, it is important to explore new anti-viral drugs to replace the traditional methods for effective control of influenza virus diseases. Previously, it was demonstrated that the influenza virus mRNAs contain internal m6Amodification that is required for viral replication [38]. Courtney et al. reported that m6A modification affected the viral replication and gene expression of IAV [42]. In detail, they used CRISPR/Cas9 system or 3-deazaadenosine (3-DAA), an inhibitor of m6A modification which inhibits SAM, to suppress the expression of METTL3 in A549 cells. They found that the viral replication and mRNA level of IAV were significantly reduced. Conversely, the expression of YTHDF2 promoted IAV replication and increased the number of infectious particles in A549 cells, whereas YTHDF1 and YTHDF3 did not affect on IAV replication. These results indicate that m6A modification might play a critical role in promoting the replication of IAV. Later, it was found that m6A modification not only exists on the positive strands of IAV mRNA/cRNA and the negative strands of vRNA, but it is also present on the viral mRNAs of HA, NA, M1/M2 and NP as determined by PAR-CLIP and PA-m6A-seq sequencing. Finally, Courtney et al. silenced m6A sites in hemagglutinin segment RNA, and found that the expression of hemagglutinin and viral replication, as well as the pathogenicity of IAV were reduced in mice.
Collectively, these findings imply that m6A modification might play a critical role in viral replication, transcription, and production. Thus, these findings add to our understanding of m6A modification in IAV.
Kaposi’s sarcoma-associated herpesvirus related diseases
The Herpesviridae family is one of the most prevalent human pathogens, and 90% of adults are infected by at least one of the eight herpesvirus subtypes [60]. KSHV is the causative agent for multiple malignancies including Kaposi’s sarcoma (KS), primary effusion lymphoma (PEL) and multicentric Castleman’s disease (MCD) [61,62,63]. Latency is common among all herpes viruses. Recently, Ye et al. discovered that reactivation of KSHV would be stalled when the newly transcribed viral RNAs fail to undergo post-transcriptional modification with m6A [43]. Reactivation of the latent virus, resets the epigenetic processes, leading to the transactivation of viral genome [64]. Therefore, a better understanding of the relationship between KSHV and m6A is a prerequisite for developing prevention and treatment strategies for KSHV-induced diseases.
Ye et al. reported that the level of m6A-modified mRNA (m6A-mRNA) for a given viral transcript increased substantially when infected cells were stimulated to undergo lytic replication [43], and this was also observed by Hesser et al. in vitro [44]. KSHV replication transcription activator (RTA), an immediate early protein encoded by Open reading frame 50 (ORF50), played an essential role in KSHV lytic replication and inhibited virion production. Ye et al. [53]stimulated KSHV-infected cells to undergo lytic replication, and observed that the splicing of pre-mRNA was inhibited in the presence of 3-DAA. Moreover, they showed that knockdown of METTL3 or inhibition of methyltransferase complexes with 3-DAA suppressed viral lytic replication in BCBL1 cells. On the other hand, knockdown of FTO moderately increased viral lytic replication. These observations indicate that KSHV activates the expression of viral genes and the lytic cycle through manipulating the host m6A modification machinery. It has also been reported that YTHDC1 interacts with the splicing factors SRSF3 and SRSF10 to form a ternary complex, and can bind to the ORF50 pre-mRNA to promote its expression. Moreover, the expression of RTA increased the modification level of m6A, further inducing the splicing of pre-mRNA.
In KSHV iSLK.219 and iSLK.BAC16 reactivation models, Hesser et al. [44] discovered that YTHDF2 plays an important role in the replication of KSHV. They used si-RNA to knock down the expression of METTL3 and YTHDF2, and found that few virions were produced after inhibition of METTL3, while knockdown of YTHDF2 displayed almost no virions. Subsequent experiments from the same group showed that inhibition of YTHDF2 affected the expression of immediate early lytic gene and viral activation. ORF50 acted as a viral transcription trans-activator which played a critical role in the lytic induction of KSHV in response to all known stimuli [65]. They also revealed that m6A modification regulated the expression of ORF50 in iSLK.219 cells, and YTHDF2 deletion impaired the transcription of ORF50. This finding suggests that YTHDF2 have a pro-viral role in DNA and RNA viruses. Besides, Tan et al. [66] found that m6Am modifications occur in KSHV transcripts during latent and lytic replication. Moreover, YTHDF2 in m6Am negatively affected lytic replication by decaying transcripts of KSHV. These observations illustrate that YTHDC1 and YTHDF2 might be involved in KSHV lifecycle, suggesting that the regulation of ORF50 expression through m6A modification could be a new research direction into the mechanisms of KSHV infections.
Hepatitis virus related diseases
Hepatitis B virus (HBV) is a DNA virus belonging to the Hepadnaviridae family. HBV infection causes chronic hepatitis and increases the risk of developing cirrhosis and hepatocellular carcinoma [67]. Recently, researchers have demonstrated that the life cycle of HBV is influenced by m6A modification that can affect the expression of viral oncoprotein and the reverse transcription of pre-genomic RNA (pgRNA) [45]. Imam et al. found that m6A modification of HBV transcript negatively regulated the expression of HBV protein and the half-life of pgRNA. Knock down of METTL3 and METTL14 increased the expression of HBc, HBs and the half-life of pgRNA, similar to YTHDF2 and YTHDF3. In contrast, low expression of FTO and ALKBH5 produced opposite results.
Apart from HBV, Hepatitis C virus (HCV) RNA also harbors m6A modification [46]. Knockdown of METTL3 and METTL14 expression enhanced the titer of HCV, whereas FTO inhibition decreased the viral titer. They also found that YTHDF protein played a negative role in HCV replication. In addition, these proteins compete with the core protein of HCV by preferentially binding to the Env to inhibit the packaging of viral RNA.
Chronic infection with HBV and HCV is the most cause of hepatocellular carcinoma (HCC) [45, 68]. Accumulating evidence indicates that HBV and HCV together with HCC oncoproteins are regulated by m6A modification. Recently, two studies revealed that m6A modification is involved the progression of HCC via METTL3 and METTL14. Chen et al. discovered that METTL3 which is highly expressed in HCC promoted the cellular proliferation, migration and colony formation of HCC cells in vitro, and enhanced tumorigenicity in vivo [47]. Suppressors of cytokine signaling 2 (SOCS2), a target gene of METTL3-mediated m6A modification, inhibit various types of cancers. Chen et al. found that knockdown of METTL3 inhibited the transcription of SOCS2 mRNA. They also demonstrated that the effect of METTL3 on SOCS2 was mediated by m6A “reader” YTHDF2. Moreover, low expression of SOCS2 was significantly associated with poor overall survival and disease-free survival of HCC patients. Ma et al. reported that METTL14, which is significantly down-regulated in HCC, regulates HCC development [48]. Compared to the study by Chen [47], the levels expression of METTL3, Wilms tumor 1–associated protein, KIAA1429, and ALKBH5 in HCC were not significantly different [48].. METTL14 is associated with frequent recurrence and poor survival; and abnormal levels of METTL14 are related with tumor differentiation, tumor stage, tumor encapsulation, microsatellite and microvascular invasion. Ma et al. concluded that microRNA126 (miR126), a downstream target of METTL14, is regulated in an m6A-dependent manner. METTL14 regulates the maturation of pri-miR126 to miR126 by enhancing the recognition and binding of the microprocessor protein DGCR8 to pri-miRNA. Collectively, these studies reveal that m6A modification plays a pivotal role in HCC through different mechanisms mediated by various m6A related proteins.
Collectively, these results show that METTL3 and METTL14 modulate m6A modification during the development of HCC. Thus, SOCS2 and miR126 may be target genes for the treatment of HCC. Further studies are required to determine the implication of m6A modification in HBV, HCV and HCC.
Human papillomavirus related diseases
Human papillomavirus (HPV) is a small DNA virus with a genome of approximately 8 kb belonging to Papillomavirus genus of the Papillomavirus family. Cervical cancer (CC) is the fourth most prevalent cancer in women and is one of the most common gynecological malignancies worldwide [69, 70]. Although significant advances have been made in cancer detection and treatment during the past few decades, the 5-year survival rate remains low. Thus, it is urgent to determine molecular mechanisms underlying the development of CC, and to explore innovative therapeutic strategies for this disease.
A recent study revealed that m6A methylation participates in the oncogenesis of cervical cancer. The study found that m6A plays a negative role in cellular proliferation tumor formation of CC [49]. Specifically, they found that the m6A level was significantly lower in cervical cancer tissues compared with adjacent non-cancerous tissues. Moreover, for patients with cervical cancer, the disease-free survival (DFS) and overall survival (OS) were significantly higher in patients with high m6A level than those with low m6A level, indicating that the level of m6A methylation could be a prognostic marker of cervical cancer. Knockdown of erasers (FTO and ALKBH5) or overexpression of writers (METTL3 and METTL14) suppressed the cellular proliferation and tumor formation of cervical cancer cells both in vitro and in vivo. Interestingly, Zhou et al. reported that overexpression of FTO induced chemo-radiotherapy resistance in cervical squamous cell carcinoma (CSCC), the major type of cervical cancer [50]. Unfortunately, FTO-mediated upregulation of β-catenin via mRNA demethylation triggered the activation of excision repair cross-complementation group 1 (ERCC1), further contributing to this resistance. It was also recognized that high expression level of FTO was associated with poor prognosis. Understanding the role of m6A in cervical cancer is expected to provide clues for optimal treatment of CC. So far, no study has described the relationship between HPV and m6A methylation, and future studies should address this gap.
Zika virus related diseases
Zika virus (ZIKV), which causes Zika virus disease (ZVD)-a type of acute infectious disease, was firstly isolated from the serum of apyrexial rhesus monkey caged in the canopy of Zika Forest [71]. In 2007, a large outbreak of ZIKV occurred in Yap Island of the Western Pacific [72], and later on in French Polynesia in 2013 [73]. More recent outbreaks were reported in Brazil and South America which reached epidemic levels [74]. Currently, there are no specific antiviral drugs for ZIKV; therefore, it is imperative to explore innovative means to prevent ZIKA infections.
One study demonstrated the existence of m6A modification in ZIKV. Lichinchi et al. confirmed that ZIKV viral RNA was methylated and twelve discrete m6A peaks spanning the full length of ZIKV RNA were identified, most of which were present in the region encoding NS5 and the 3’UTR [51]. Like other virus, ZIKV viral-RNA is also modified at adenosines by writers and erasers; and perturbation of ZIKV m6A affects the replication efficiency and viral titer of ZIKV. Knockdown of writers (METTL3 or METTL14) significantly increased the viral-production, viral titer, ZIKV RNA levels in cell supernatants and the expression of ZIKV envelope protein, whereas silencing of erasers (ALKBH5 or FTO) produced opposite effect. It was also reported that readers (YTHDF proteins) that bind to ZIKV RNA may regulate the replication of ZIKV. Compared to YTHDF1 and YTHDF3, YTHDF2 has the greatest effect on ZIKV replication, RNA expression and stability of viral RNA. Generally, these results point to a new mechanism for the relationship between m6A associated enzymes and ZIKV. This knowledge is expected to generate ideas for the design of treatments for ZIKV-related diseases.
Simian virus 40 related diseases
Simian Virus 40 (SV40), a polyomavirus of the rhesus macaque, is a double-stranded DNA virus, and a potent DNA tumor virus reported to induce human primary brain tumors, malignant mesotheliomas, bone cancers, and non-Hodgkin’s lymphoma [75, 76]. In 1979, it was reported that late SV40 16S and 19S mRNAs contain numerous m6A modified residues [35]. However, the exact location of m6A modified residues is not known, and the functional significance of these residues is unclear. Recently, Tsai et al. identified and precisely mapped the m6A peaks on SV40, including eleven peaks in late transcripts, and two in the early region by photo-crosslinking-assisted m6A sequencing (PA-m6A-seq), photoactivatable ribonucleoside-enhanced crosslinking and immunoprecipitation (PAR-CLIP) [52]. In addition, they demonstrated that overexpression of YTHDF2 significantly enhanced the replication of SV40, and mutation of YTHDF2 inhibited the replication of SV40. Furthermore, knockdown of YTHDF2 or METTL3 by gene editing suppressed the expression of SV40 structural gene in the permissive cell line BSC40. Overall, this investigation revealed that m6A modification plays a positive role in the regulation of SV40 life cycle.
Epstein–Barr virus related diseases
Epstein–Barr virus (EBV), a herpesvirus, is an oncogenic virus first isolated and identified from a Burkitt’s lymphoma patient in 1964 [77]. As one of the most common human viruses, EBV causes infectious mononucleosis and is associated with specific forms of lymphoma. There are about 200,000 cases of cancer associated with this virus, with about 140,000 deaths reported annually [78]. Common antiviral therapies can suppress the replication of active-viruses. However, the existing vaccines or drugs are not effective in eradicating the latent infection of EBV.
To date, only one study has reported the association of m6A “writers” with EBV mainly through METTL14 [53]. METTL14 was increased in EBV latently infected cells and decreased during the lytic infection phase. It should be noted that METTL14 facilitates the cellular proliferation and colony formation of EBV transformed cells in vitro, and enhances the oncogenesis of EBV tumorigenicity in vivo. It was reported that METTL14 impairs the stability of the latent genes (such as EBNA1, EBNA3C, and LMP1) and lytic genes (such as BRLF1, gp350, and BMRF1). Elsewhere, EBNA3C was found to be a downstream target gene of METTL14, and EBV antigen up-regulated and stabilized METTL14. EBV latent antigens are the major contributors to EBV-associated malignancies, and low expression of EBV latent antigens attenuates the EBV-mediated tumorigenesis. Therefore, targeting METTL14 may be a key strategy for controlling EBV-associated cancer.
Conclusions
The viruses described in this review are harmful to human health. Some viruses RNA undergo m6A modification such as Rous sarcoma virus (RSV) [79], vesicular stomatitis virus (VSV) [80], and adenoviruses [37], and are less pathogenic in human. Several advances have been made in understanding human pathogenic viruses in recent years, however, only few drugs for these diseases have been put on the market in the past decades. For this reason, it is necessary to focus on the development of new treatments that may replace or improve the standard therapies.
The emergence of posttranscriptional modification of RNAs, especially the methylation of RNAs, has triggered several investigations into the role of such modifications on gene expression, cell behaviors, and physiological conditions in many species, including humans. Among more than 100 kinds of chemical modifications, m6A is the most common modification initially discovered in the poly(A) RNA fractions and has been predicted to be involved in mRNA processing [4]. m6A is a dynamic and reversible process initiated by “writers” and removed by “erasers” and binds to “readers”. Emerging evidence indicates that aberrant expression of proteins related to m6A modification is associated with the development of various cancers, such as acute myeloid leukemia [81, 82], lung cancer [83], and hepatocellular carcinoma [47, 48]. Particularly, m6A has been detected in viral mRNA and the RNA of retroviruses. It can influence the expression of viral genes and viral life cycle, providing a new method for the treatment of many malignant virus-related diseases.
The development of small molecule inhibitors targeted at writers, erasers and readers in m6A modification process is expected to improve antiviral therapy. Recent studies have shown that the use of writer inhibitors may affect viral replication and the development of tumors [1, 47, 84]. Thus, inhibition of m6A modification may be a potential therapy for virus induced diseases.
Numerous studies have shown that m6A modification plays a significant role in human pathogenic viruses, but there are some challenges to be addressed. First, the specific mechanism explaining the interaction of m6A modification proteins with viral RNA is largely unknown. Second, before the m6A modification-related regulatory genes and proteins can be used as prognostic and diagnostic markers for some viral diseases, studies are needed to explore the specificity and targeting precision of these molecules. Moreover, the effects of these biomolecules on normal cells remains to be clarified. Third, many researchers have reported that substances similar to inhibitors could be used to block m6A-modified abnormalities to control related diseases, but due to the lack of large clinical trials, the corresponding effects of these substances are not fully known. Thus, further studies are required to address these issues.
Availability of data and materials
Not applicable.
Abbreviations
- 3’UTR:
-
3’untranslated region
- 5’UTR:
-
5’untranslated region
- AIDS:
-
acquired immunodeficiency syndrome
- ALKBH5:
-
alkB homologue 5
- CC:
-
Cervical cancer
- co-IP:
-
co-immunoprecipitation
- CSCC:
-
cervical squamous cell carcinoma
- DAA:
-
3-deazaadenosine
- DFS:
-
disease-free survival
- EBV:
-
Epstein–Barr virus
- ERCC1:
-
excision repair cross-complementation group 1
- EV71:
-
Enterovirus 71
- FTO:
-
fat mass and obesity-associated proteins
- HBV:
-
Hepatitis B virus
- HCC:
-
Hepatocellular Carcinoma
- HCV:
-
Hepatitis C virus
- HIV-1:
-
Human immunodeficiency virus I
- HPV:
-
Human papillomavirus
- IAV:
-
Influenza A virus
- IGF2BPs:
-
the insulin-like growth factor 2 mRNA-binding proteins
- KS:
-
Kaposi’s sarcoma
- KSHV:
-
Kaposi’s sarcoma-associated herpesvirus
- m6A:
-
N6-methyladenosine
- m6Am:
-
N6,2′-O-dimethyladenosine
- m6A-seq:
-
high-throughput RNA sequencing
- MCD:
-
multicentric Castleman’s disease
- MeRIP-seq:
-
methylated RNA immunoprecipitation sequening
- METTL14:
-
methyltransferase-like 14
- METTL16:
-
methyltransferase-like 16
- METTL3:
-
methyltransferase-like 3
- miR126:
-
microRNA126
- mRNA:
-
messenger RNA
- MT:
-
methyltransferase
- ORF50:
-
Open reading frame 50
- OS:
-
overall survival
- PA-m6A-seq:
-
photo-crosslinking-assisted m6A sequencing
- PAR-CLIP:
-
photoactivatable ribonucleoside-enhanced crosslinking and immunoprecipitation
- PEL:
-
primary effusion lymphoma
- RBM15:
-
RNA binding motif protein 15
- RdRp:
-
RNA-dependent RNA polymerase
- RRE:
-
Rev response element
- rRNA:
-
ribosomal RNA
- RSV:
-
Rous sarcoma virus
- RTA:
-
replication transcription activator
- SOCS2:
-
suppressors of cytokine signaling 2
- SV40:
-
Simian Virus 40
- tRNA:
-
transfer RNA
- VSV:
-
vesicular stomatitis virus
- WTAP:
-
Wilms’ tumor 1-associating protein
- ZIKV:
-
Zika virus
- ZVD:
-
Zika virus disease
References
Kennedy EM, Bogerd HP, Kornepati AV, Kang D, Ghoshal D, Marshall JB, Poling BC, Tsai K, Gokhale NS, Horner SM, et al. Posttranscriptional m(6)a editing of HIV-1 mRNAs enhances viral gene expression. Cell Host Microbe. 2016;19:675–85.
Boccaletto P, Machnicka MA, Purta E, Piatkowski P, Baginski B, Wirecki TK, de Crecy-Lagard V, Ross R, Limbach PA, Kotter A, et al. MODOMICS: a database of RNA modification pathways. 2017 update. Nucleic Acids Res. 2018;46:D303–7.
Cantara WA, Crain PF, Rozenski J, McCloskey JA, Harris KA, Zhang X, Vendeix FA, Fabris D, Agris PF. The RNA Modification Database, RNAMDB: 2011 update. Nucleic Acids Res. 2011;39:D195–201.
Desrosiers R, Friderici K, Rottman F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc Natl Acad Sci U S A. 1974;71:3971–5.
Meyer KD, Jaffrey SR. The dynamic epitranscriptome: N6-methyladenosine and gene expression control. Nat Rev Mol Cell Biol. 2014;15:313–26.
Dominissini D, Moshitch-Moshkovitz S, Schwartz S, Salmon-Divon M, Ungar L, Osenberg S, Cesarkas K, Jacob-Hirsch J, Amariglio N, Kupiec M, et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature. 2012;485:201–6.
Linder B, Grozhik AV, Olarerin-George AO, Meydan C, Mason CE, Jaffrey SR. Single-nucleotide-resolution mapping of m6A and m6Am throughout the transcriptome. Nat Methods. 2015;12:767–72.
Meyer KD, Saletore Y, Zumbo P, Elemento O, Mason CE, Jaffrey SR. Comprehensive analysis of mRNA methylation reveals enrichment in 3′ UTRs and near stop codons. Cell. 2012;149:1635–46.
Yue Y, Liu J, He C. RNA N6-methyladenosine methylation in post-transcriptional gene expression regulation. Genes Dev. 2015;29:1343–55.
Liu J, Yue Y, Han D, Wang X, Fu Y, Zhang L, Jia G, Yu M, Lu Z, Deng X, et al. A METTL3–METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat Chem Biol. 2014;10:93–5.
Ping XL, Sun BF, Wang L, Xiao W, Yang X, Wang WJ, Adhikari S, Shi Y, Lv Y, Chen YS, et al. Mammalian WTAP is a regulatory subunit of the RNA N6-methyladenosine methyltransferase. Cell Res. 2014;24:177–89.
Jia G, Fu Y, Zhao X, Dai Q, Zheng G, Yang Y, Yi C, Lindahl T, Pan T, Yang YG, et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat Chem Biol. 2011;7:885–7.
Liu Z, Zhang J. Human C-to-U coding RNA editing is largely nonadaptive. Mol Biol Evol. 2018;35:963–9.
Warda AS, Kretschmer J, Hackert P, Lenz C, Urlaub H, Hobartner C, Sloan KE, Bohnsack MT. Human METTL16 is a N(6)-methyladenosine (m(6)a) methyltransferase that targets pre-mRNAs and various non-coding RNAs. EMBO Rep. 2017;18:2004–14.
Patil DP, Chen C, Pickering BF, Chow A, Jackson C, Guttman M, Jaffrey SR. m6A RNA methylation promotes XIST-mediated transcriptional repression. Nature. 2016;537:369–73.
Zheng G, Dahl JA, Niu Y, Fedorcsak P, Huang CM, Li CJ, Vagbo CB, Shi Y, Wang WL, Song SH, et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol Cell. 2013;49:18–29.
Bokar JA, Rath-Shambaugh ME, Ludwiczak R, Narayan P, Rottman F. Characterization and partial purification of mRNA N6-adenosine methyltransferase from HeLa cell nuclei. Internal mRNA methylation requires a multisubunit complex. J Biol Chem. 1994;269:17697–704.
Ianniello Z, Fatica A. N6-Methyladenosine role in acute myeloid Leukaemia. Int J Mol Sci. 2018;19:2345.
Schwartz S, Mumbach MR, Jovanovic M, Wang T, Maciag K, Bushkin GG, Mertins P, Ter-Ovanesyan D, Habib N, Cacchiarelli D, et al. Perturbation of m6A writers reveals two distinct classes of mRNA methylation at internal and 5′ sites. Cell Rep. 2014;8:284–96.
Pendleton KE, Chen B, Liu K, Hunter OV, Xie Y, Tu BP, Conrad NK. The U6 snRNA m(6)a methyltransferase METTL16 regulates SAM Synthetase intron retention. Cell. 2017;169:824–35.
Doxtader KA, Wang P, Scarborough AM, Seo D, Conrad NK, Nam Y. Structural basis for regulation of METTL16, an S-Adenosylmethionine homeostasis factor. Mol Cell. 2018;71:1001–11.
Jia G, Yang CG, Yang S, Jian X, Yi C, Zhou Z, He C. Oxidative demethylation of 3-methylthymine and 3-methyluracil in single-stranded DNA and RNA by mouse and human FTO. FEBS Lett. 2008;582:3313–9.
Bartosovic M, Molares HC, Gregorova P, Hrossova D, Kudla G, Vanacova S. N6-methyladenosine demethylase FTO targets pre-mRNAs and regulates alternative splicing and 3′-end processing. Nucleic Acids Res. 2017;45:11356–70.
Mauer J, Luo X, Blanjoie A, Jiao X, Grozhik AV, Patil DP, Linder B, Pickering BF, Vasseur JJ, Chen Q, et al. Reversible methylation of m(6)am in the 5′ cap controls mRNA stability. Nature. 2017;541:371–5.
Wang X, Zhao BS, Roundtree IA, Lu Z, Han D, Ma H, Weng X, Chen K, Shi H, He C. N(6)-methyladenosine modulates messenger RNA translation efficiency. Cell. 2015;161:1388–99.
Du H, Zhao Y, He J, Zhang Y, Xi H, Liu M, Ma J, Wu L. YTHDF2 destabilizes m(6)A-containing RNA through direct recruitment of the CCR4-NOT deadenylase complex. Nat Commun. 2016;7:12626.
Shi H, Wang X, Lu Z, Zhao BS, Ma H, Hsu PJ, Liu C, He C. YTHDF3 facilitates translation and decay of N(6)-methyladenosine-modified RNA. Cell Res. 2017;27:315–28.
Li A, Chen YS, Ping XL, Yang X, Xiao W, Yang Y, Sun HY, Zhu Q, Baidya P, Wang X, et al. Cytoplasmic m(6)a reader YTHDF3 promotes mRNA translation. Cell Res. 2017;27:444–7.
Roundtree IA, Luo GZ, Zhang Z, Wang X, Zhou T, Cui Y, Sha J, Huang X, Guerrero L, Xie P, et al. YTHDC1 mediates nuclear export of N(6)-methyladenosine methylated mRNAs. Elife. 2017;6.
Roundtree IA, He C. Nuclear m(6)a reader YTHDC1 regulates mRNA splicing. Trends Genet. 2016;32:320–1.
Hsu PJ, Zhu Y, Ma H, Guo Y, Shi X, Liu Y, Qi M, Lu Z, Shi H, Wang J, et al. Ythdc2 is an N(6)-methyladenosine binding protein that regulates mammalian spermatogenesis. Cell Res. 2017;27:1115–27.
Huang H, Weng H, Sun W, Qin X, Shi H, Wu H, Zhao BS, Mesquita A, Liu C, Yuan CL, et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat Cell Biol. 2018;20:285–95.
Li Y, Wang X, Li C, Hu S, Yu J, Song S. Transcriptome-wide N(6)-methyladenosine profiling of rice callus and leaf reveals the presence of tissue-specific competitors involved in selective mRNA modification. RNA Biol. 2014;11:1180–8.
Alarcon CR, Goodarzi H, Lee H, Liu X, Tavazoie S, Tavazoie SF. HNRNPA2B1 is a mediator of m(6)A-dependent nuclear RNA processing events. Cell. 2015;162:1299–308.
Canaani D, Kahana C, Lavi S, Groner Y. Identification and mapping of N6-methyladenosine containing sequences in simian virus 40 RNA. Nucleic Acids Res. 1979;6:2879–99.
Beemon K, Keith J. Localization of N6-methyladenosine in the Rous sarcoma virus genome. J Mol Biol. 1977;113:165–79.
Hashimoto SI, Green M. Multiple methylated cap sequences in adenovirus type 2 early mRNA. J Virol. 1976;20:425–35.
Krug RM, Morgan MA, Shatkin AJ. Influenza viral mRNA contains internal N6-methyladenosine and 5′-terminal 7-methylguanosine in cap structures. J Virol. 1976;20:45–53.
Lichinchi G, Gao S, Saletore Y, Gonzalez GM, Bansal V, Wang Y, Mason CE, Rana TM. Dynamics of the human and viral m(6)a RNA methylomes during HIV-1 infection of T cells. Nat Microbiol. 2016;1:16011.
Tirumuru N, Zhao BS, Lu W, Lu Z, He C, Wu L. N(6)-methyladenosine of HIV-1 RNA regulates viral infection and HIV-1 gag protein expression. Elife. 2016;5.
Hao H, Hao S, Chen H, Chen Z, Zhang Y, Wang J, Wang H, Zhang B, Qiu J, Deng F, et al. N6-methyladenosine modification and METTL3 modulate enterovirus 71 replication. Nucleic Acids Res. 2019;47:362–74.
Courtney DG, Kennedy EM, Dumm RE, Bogerd HP, Tsai K, Heaton NS, Cullen BR. Epitranscriptomic enhancement of influenza a virus gene expression and replication. Cell Host Microbe. 2017;22:377–86.
Ye F, Chen ER, Nilsen TW. Kaposi’s sarcoma-associated Herpesvirus utilizes and manipulates RNA N(6)-adenosine methylation to promote lytic replication. J Virol. 2017;91.
Hesser CR, Karijolich J, Dominissini D, He C, Glaunsinger BA. N6-methyladenosine modification and the YTHDF2 reader protein play cell type specific roles in lytic viral gene expression during Kaposi's sarcoma-associated herpesvirus infection. PLoS Pathog. 2018;14:e1006995.
Imam H, Khan M, Gokhale NS, McIntyre A, Kim GW, Jang JY, Kim SJ, Mason CE, Horner SM, Siddiqui A. N6-methyladenosine modification of hepatitis B virus RNA differentially regulates the viral life cycle. Proc Natl Acad Sci U S A. 2018;115:8829–34.
Gokhale NS, McIntyre A, McFadden MJ, Roder AE, Kennedy EM, Gandara JA, Hopcraft SE, Quicke KM, Vazquez C, Willer J, et al. N6-Methyladenosine in Flaviviridae viral RNA genomes regulates infection. Cell Host Microbe. 2016;20:654–65.
Chen M, Wei L, Law CT, Tsang FH, Shen J, Cheng CL, Tsang LH, Ho DW, Chiu DK, Lee JM, et al. RNA N6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2. Hepatology. 2018;67:2254–70.
Ma JZ, Yang F, Zhou CC, Liu F, Yuan JH, Wang F, Wang TT, Xu QG, Zhou WP, Sun SH. METTL14 suppresses the metastatic potential of hepatocellular carcinoma by modulating N6-methyladenosine-dependent primary MicroRNA processing. Hepatology. 2016;65:529–43.
Wang X, Li Z, Kong B, Song C, Cong J, Hou J, Wang S. Reduced m(6)a mRNA methylation is correlated with the progression of human cervical cancer. Oncotarget. 2017;8:98918–30.
Zhou S, Bai Z, Xia D, Zhao Z, Zhao R, Wang Y, Zhe H. FTO regulates the chemo-radiotherapy resistance of cervical squamous cell carcinoma (CSCC) by targeting β-catenin through mRNA demethylation. Mol Carcinog. 2018;57:590–7.
Lichinchi G, Zhao BS, Wu Y, Lu Z, Qin Y, He C, Rana TM. Dynamics of human and viral RNA methylation during Zika virus infection. Cell Host Microbe. 2016;20:666–73.
Tsai K, Courtney DG, Cullen BR. Addition of m6A to SV40 late mRNAs enhances viral structural gene expression and replication. PLoS Pathog. 2018;14:e1006919.
Lang F, Singh RK, Pei Y, Zhang S, Sun K, Robertson ES. EBV epitranscriptome reprogramming by METTL14 is critical for viral-associated tumorigenesis. PLoS Pathog. 2019;15:e1007796.
Mendoza P, Gruell H, Nogueira L, Pai JA, Butler AL, Millard K, Lehmann C, Suarez I, Oliveira TY, Lorenzi J, et al. Combination therapy with anti-HIV-1 antibodies maintains viral suppression. Nature. 2018;561:479–84.
Reddy TR, Xu W, Mau JK, Goodwin CD, Suhasini M, Tang H, Frimpong K, Rose DW, Wong-Staal F. Inhibition of HIV replication by dominant negative mutants of Sam68, a functional homolog of HIV-1 rev. Nat Med. 1999;5:635–42.
Wang H, Li K, Ma L, Wu S, Hu J, Yan H, Jiang J, Li Y. Berberine inhibits enterovirus 71 replication by downregulating the MEK/ERK signaling pathway and autophagy. Virol J. 2017;14:2.
Chang PC, Chen SC, Chen KT. The current status of the disease caused by Enterovirus 71 infections: epidemiology, pathogenesis, molecular epidemiology, and vaccine development. Int J Environ Res Public Health. 2016;13.
To EE, Luong R, Diao J, JJ OL, Brooks DA, Vlahos R, Selemidis S. Novel endosomal NOX2 oxidase inhibitor ameliorates pandemic influenza A virus-induced lung inflammation in mice. Respirology. 2019;24:1011–7.
Sun J, Braciale TJ. Role of T cell immunity in recovery from influenza virus infection. Curr Opin Virol. 2013;3:425–9.
Barry PA, William CW. Primate betaherpesviruses; 2007.
Chang Y, Cesarman E, Pessin MS, Lee F, Culpepper J, Knowles DM, Moore PS. Identification of herpesvirus-like DNA sequences in AIDS-associated Kaposi's sarcoma. Science. 1994;266:1865–9.
Cesarman E, Chang Y, Moore PS, Said JW, Knowles DM. Kaposi’s sarcoma-associated herpesvirus-like DNA sequences in AIDS-related body-cavity-based lymphomas. N Engl J Med. 1995;332:1186–91.
Soulier J, Grollet L, Oksenhendler E, Cacoub P, Cazals-Hatem D, Babinet P, D'Agay MF, Clauvel JP, Raphael M, Degos L, et al. Kaposi's sarcoma-associated herpesvirus-like DNA sequences in multicentric Castleman's disease. Blood. 1995;86:1276–80.
Ye F. RNA N(6)-adenosine methylation (m(6)a) steers epitranscriptomic control of herpesvirus replication. Inflamm Cell Signal. 2017;4.
Lukac DM, Kirshner JR, Ganem D. Transcriptional activation by the product of open reading frame 50 of Kaposi's sarcoma-associated herpesvirus is required for lytic viral reactivation in B cells. J Virol. 1999;73:9348–61.
Tan B, Liu H, Zhang S, Da SS, Zhang L, Meng J, Cui X, Yuan H, Sorel O, Zhang SW, et al. Viral and cellular N(6)-methyladenosine and N(6),2′-O-dimethyladenosine epitranscriptomes in the KSHV life cycle. Nat Microbiol. 2018;3:108–20.
Levrero M, Pollicino T, Petersen J, Belloni L, Raimondo G, Dandri M. Control of cccDNA function in hepatitis B virus infection. J Hepatol. 2009;51:581–92.
Raza SA, Clifford GM, Franceschi S. Worldwide variation in the relative importance of hepatitis B and hepatitis C viruses in hepatocellular carcinoma: a systematic review. Br J Cancer. 2007;96:1127–34.
Ferlay J, Soerjomataram I, Dikshit R, Eser S, Mathers C, Rebelo M, Parkin DM, Forman D, Bray F. Cancer incidence and mortality worldwide: sources, methods and major patterns in GLOBOCAN 2012. Int J Cancer. 2015;136:E359–86.
Jung KW, Won YJ, Kong HJ, Lee ES. Cancer statistics in Korea: incidence, mortality, survival, and prevalence in 2016. Cancer Res Treat. 2019;51:417–30.
DICK GW, KITCHEN SF, HADDOW AJ. Zika virus. I. Isolations and serological specificity. Trans R Soc Trop Med Hyg. 1952;46:509–20.
Duffy MR, Chen TH, Hancock WT, Powers AM, Kool JL, Lanciotti RS, Pretrick M, Marfel M, Holzbauer S, Dubray C, et al. Zika virus outbreak on Yap Island, Federated States of Micronesia. N Engl J Med. 2009;360:2536–43.
Oehler E, Watrin L, Larre P, Leparc-Goffart I, Lastere S, Valour F, Baudouin L, Mallet H, Musso D, Ghawche F. Zika virus infection complicated by Guillain-Barre syndrome--case report, French Polynesia, December 2013. Euro Surveill. 2014;19.
Schuler-Faccini L, Ribeiro EM, Feitosa IM, Horovitz DD, Cavalcanti DP, Pessoa A, Doriqui MJ, Neri JI, Neto JM, Wanderley HY, et al. Possible association between Zika virus infection and microcephaly - Brazil, 2015. MMWR Morb Mortal Wkly Rep. 2016;65:59–62.
Liddington RC, Yan Y, Moulai J, Sahli R, Benjamin TL, Harrison SC. Structure of simian virus 40 at 3.8-a resolution. Nature. 1991;354:278–84.
Vilchez RA, Kozinetz CA, Arrington AS, Madden CR, Butel JS. Simian virus 40 in human cancers. Am J Med. 2003;114:675–84.
EPSTEIN MA, ACHONG BG, BARR YM. VIRUS PARTICLES IN CULTURED LYMPHOBLASTS FROM BURKITT'S LYMPHOMA. Lancet. 1964;1:702–3.
Bu W, Joyce MG, Nguyen H, Banh DV, Aguilar F, Tariq Z, Yap ML, Tsujimura Y, Gillespie RA, Tsybovsky Y, et al. Immunization with components of the viral fusion apparatus elicits antibodies that neutralize Epstein-Barr virus in B cells and epithelial cells. Immunity. 2019;50:1305–16.
Kane SE, Beemon K. Precise localization of m6A in Rous sarcoma virus RNA reveals clustering of methylation sites: implications for RNA processing. Mol Cell Biol. 1985;5:2298–306.
Zheng Q, Hou J, Zhou Y, Li Z, Cao X. Corrigendum: the RNA helicase DDX46 inhibits innate immunity by entrapping m(6)A-demethylated antiviral transcripts in the nucleus. Nat Immunol. 2017;18:1361.
Li Z, Weng H, Su R, Weng X, Zuo Z, Li C, Huang H, Nachtergaele S, Dong L, Hu C, et al. FTO plays an oncogenic role in acute myeloid leukemia as a N(6)-Methyladenosine RNA Demethylase. Cancer Cell. 2017;31:127–41.
Kwok CT, Marshall AD, Rasko JE, Wong JJ. Genetic alterations of m(6)a regulators predict poorer survival in acute myeloid leukemia. J Hematol Oncol. 2017;10:39.
Liu J, Ren D, Du Z, Wang H, Zhang H, Jin Y. M(6)a demethylase FTO facilitates tumor progression in lung squamous cell carcinoma by regulating MZF1 expression. Biochem Biophys Res Commun. 2018;502:456–64.
Vu LP, Pickering BF, Cheng Y, Zaccara S, Nguyen D, Minuesa G, Chou T, Chow A, Saletore Y, MacKay M, et al. The N(6)-methyladenosine (m(6)a)-forming enzyme METTL3 controls myeloid differentiation of normal hematopoietic and leukemia cells. Nat Med. 2017;23:1369–76.
Acknowledgments
Not applicable.
Funding
This study was supported by the National Natural Science Foundation of China (No. 81271692), the Ocean Antithrombotic Fibrinolytic Enzyme Gene Bank of Taiwan Strait (No. 2014FJPT08), the Science and Technology Innovation Public Technology Service Platform of Function of Drugs and Food (No. 3502Z20141015), Education Department of Fujian Province (JA14020) and Subsidized Project for Postgraduates’ Innovative Fund in Scientific Research of Huaqiao University (No. 17013071028).
Author information
Authors and Affiliations
Contributions
FW, WC and FZ collected references and wrote the manuscript; WC, YD, MT and RX revised and completed the final manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Wu, F., Cheng, W., Zhao, F. et al. Association of N6-methyladenosine with viruses and related diseases. Virol J 16, 133 (2019). https://doi.org/10.1186/s12985-019-1236-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12985-019-1236-3