
Abstract
Nanocarriers as drug/biomolecule delivery systems have been significantly developed during recent decades. Given the stability, reasonable delivery efficiency, and safety of nanocarriers, there are several barriers in the fulfillment of successful clinical application of these delivery systems. These challenges encouraged drug delivery researchers to establish innovative nanocarriers with longer circulation time, high stability, and high compatibility. Exosomes are extracellular nanometer-sized vesicles released through various cells. These vesicles serve as nanocarriers, possessing great potential to overcome some obstacles encountered in gene and drug delivery due to their natural affinity to recipient cells and the inherent capability to shuttle the genes, lipids, proteins, and RNAs between cells. So far, there has been a lot of valuable research on drug delivery by exosomes, but research on RNA delivery, especially mRNA, is very limited. Since mRNA-based vaccines and therapies have recently gained particular prominence in various diseases, it is essential to find a suitable delivery system due to the large size and destructive nature of these nucleic acids. That's why we're going to take a look at the unique features of exosomes and their isolation and loading methods, to embrace this idea that exosome-mediated mRNA-based therapies would be introduced as a very efficient strategy in disease treatment within the near future.
Graphical Abstract

Similar content being viewed by others

Introduction
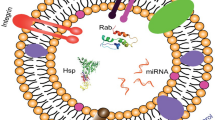
Exosomes are extracellular membranous vesicles (EMVs) possessing the special compositions of DNA, coding RNAs, non-coding RNAs, lipids, and proteins [1,2,3,4]. These endocytic membranes-derived vesicles can deliver various signals to target cells, thereby mediating new cell-to-cell communication mechanisms [3]. In -molecular-scale studies, exosomes possess phospholipases and lipid-related proteins, proteins participating in the biosynthesis of multi-vesicular bodies (MVBs) (TSG101, Alix), proteins involved in transport and fusion of membranes (flotillin, GTPases, annexins), heat shock proteins (i.e., Hsp90, Hsc70), and tetraspanins (CD82, CD81, CD63, CD9) [5]. Until now, a total of 4500 various proteins have been detected with exosomes, often through mass spectrometry (MS) [6], likely serving as cargos for intercellular communications. In addition to the above-mentioned proteins, the membranous vesicles are enriched in special raft-related lipids, like phosphoglycerides with saturated fatty-acyl and long chains, sphingolipids, ceramide, and cholesterol (mostly B lymphocytes-derived exosomes) [7, 8].
The previous reports have proven that the exosomes derived from various fluids in the body [9, 10] and various cell lines [11] possess RNA molecules, especially mRNAs and miRNAs, which can be translated into proteins or regulate protein expression in the recipient cells, respectively [12,13,14,15]. Current deep-sequencing studies have revealed that the membranous vesicles possess a variety of RNA cargoes comprising full-length RNAs with 25–250 nucleotides in lengths like tRNA and miRNA, as well as fragments of long RNA like rRNA, mRNA, even though mRNA molecules are presented in full-length types [16,17,18,19,20,21].
Exosomes play multi-pronged roles in the tissues or cells of origin to facilitate movement of pathogens such as viruses [22] and prions from one cell to another cell [23], inducing tumorigenesis [24], coagulation [25], inflammation [26], angiogenesis [4], programmed cell death [27], antigen presentation [3, 28], improvement of the immune responses, and deletion of debris molecules [29]. Interestingly, exosomes have been proved to enable signaling and cell-to-cell communications and deliver macromolecular messages such as proteins and RNAs [1]. Interest towards these membranous vesicles ranging from in vivo roles to further uses like their application in therapeutics, biomarker development, and diagnostics (according to the assay of their protein and RNA extent) has substantially increased in the past decades [22, 26, 29].
Owing to their potential application in translational research, these vesicles have fascinated a lot of fieldwork attention on their functions in various diseases and health [8, 26, 30,31,32]. These EMVs, especially mesenchymal stem cells (MSCs)-derived exosomes play an important role in kidney diseases, wound healing, liver diseases, neurodegenerative and autoimmune disorders, diabetes, spinal cord injury, and other diseases. Moreover, exosomes have attracted a great deal of attention as novel bio-carriers for gene and drug delivery because of the advantages such as less toxicity, the ability to cross biological barriers, and evade mononuclear phagocytic system (MPS) compared to synthetic nanoparticles [33]. Thus, highly efficient procedures for loading exosomes with drugs or biomolecules are prerequisites to achieving breakthroughs in the future [17, 22]. Although several studies have elucidated the detection, isolation, and characterization of exosomes along with their uses in drug delivery [1, 2, 4, 25, 26], the preparation and application of exosomes as the bio-inspired nanocarriers in RNA-based therapeutics have not been explored. In this study, the cutting-edge advances in the latest studies for the isolation and production of exosomes along with their clinical application in the delivery of RNA molecules, have been discussed. We revealed that exosomes with their unique properties and safety could open a new avenue for researchers in the development of efficient mRNA delivery system. Therefore, the establishment of large-scale RNA-loaded exosomes for clinical applications should be given further attention.
Biogenesis
Exosome biogenesis starts with the generation of early endosome through inward budding of the cellular membrane followed by second budding of the endosomal membrane. The second budding leads to the production of late endosome. These endosomes including intraluminal vesicles (ILV), are known as multivesicular bodies. These bodies either follow the endocytic pathway for the exosome generation or fuse with lysosome to degradation [25]. During inward budding of the endosomal membrane, proteins, miRNA, mRNA, and DNA fragments are incorporated into the forming vesicles by very specific protein complex. Finally, by fusion of the ILV with plasma membrane the exosomes are released into extracellular space [26].
Sources of exosomes
The term "exosome" was first presented in the 1980s for vesicles released by a cell line having ectoenzyme activities [34]. In fact, it refers to the vesicles secreted during the differentiation of reticulocytes [35], whereas, the EMVs are reported to be secreted through dendritic cells (DCs) and B lymphocytes via a similar path [24]. In the latter years, several cell kinds of nonhematopoietic/hematopoietic origin, like oligodendrocytes, intestinal epithelial cells, Schwann cells, neurons, platelets, cytotoxic T cells, and mast cells have been documented to release the EMVs [29, 36]. Moreover, the vesicles have been found in the various fluids of the body, such as bile [37], cerebrospinal fluid [28], amniotic fluid [38], ascites fluid [39], breast milk [40], urine [41], semen [42], saliva [43], and blood [44].
Stem cell-derived exosomes
Recently, efforts to repair human tissues by stem cells (SCs) have attracted considerable attention in regenerative medicine [23]. Many SCs like induced pluripotent SCs (iPSCs), embryonic SCs (ESCs), and MSCs with considerable potential in cell proliferation and differentiation have been used to repair human tissues [4, 13, 45]. SCs were found to discharge many products in a paracrine manner that results in their related influences. These products consist of extracellular vesicles, cytokines, and various growth factors [23]. EMVs released through SCs are commonly pointed out as exosomes, shedding vesicles, microparticles, cell-derived vesicles, and microvesicles [46]. The exosomes secreted from SCs have exhibited considerable potential in cell-free regenerative medicine. For example, Gibbings et al. [47] suggested that the exosomes derived from SCs can package particular miRNAs for regulating the various cell processes. This provides an opportunity for EMVs-based remedial approaches in musculoskeletal diseases because the miRNA molecules have essential functions in the prevention and progression of disorders.
Dietary sources-derived exosomes
EMVs and their cargos can be isolated from dietary resources, like milk. These vesicles and cellular glycoproteins are imperative for intestinal uptake. A considerable fraction of milk-derived vesicles accumulates in the brain tissue [47]. The part of milk-derived EMVs that gets away from absorption elicits alteration in microbial populations in the gastrointestinal tract [47]. Dietary discharge of EMVs and their cargos leads to a loss of circulating RNAs and induces some properties, like the changed immune responses, loss of fecundity, increased purine metabolites, and loss of cognitive performance [23]. Overall, the exosomes derived from milk meet the definition of bioactive ingredients in foods. In an applied study, Reif et al. [48] presented evidence that milk-derived exosomes possess a various biological efficacy on normal fetal colon epithelial cell in contrast to colon tumor cell in a miRNA-dependent manner. Thus, they suggested that the positive efficacy of exosome on normal cell without affecting tumor cell can present a promising aspect of its safety when regarding its application as a nutritional supplement to infant formula.
Plant cell-derived exosomes
Exosomes from non-animal organisms remain underestimated, and our understanding of them is still expansible, albeit animal-derived exosomes are moving increasingly into interest areas of research [23]. This is due to the lack of a practicable and easy visualization, purification, and isolation procedure. The exosomes from Viscum album L., Vinca minor L., and Nicotiana tabacum L., were isolated by differential centrifugation supplemented with agarose gel electrophoresis. It is claimed that the combination of electrophoresis with differential centrifugation can improve the isolation of exosomes [49]. Woith and Melzig purified exosomes from the dried material of plants, successfully. Further, exosomes can be separated from dye excess and small charged impurities because the contaminants can pass the gel matrix, whereas exosomes remain within the wells and do not enter the gel. [23, 49, 50].
Cancer cell-derived exosomes
According to the fact that cancerous cells release many more EVs in contrast to normal cells, it was found that (1) the active secretion of exosomes possesses functional senses, although it is unknown whether they are cancer-suppressing or promoting [51]; (2) the exosomes can be utilized as markers for cancer diagnosis [23]. Interestingly, it was detected that the interactions caused by the exchange of exosomes among tumor stroma and cancer cells can promote the transfer of oncomicroRNAs (e.g., miR375, miR16, miR15, miR1, and let7) as well as oncogenes (e.g., LMP-1, Melan-A/Mart-1, HER2, CEA, and β-catenin) from one cell to another, resulting in new programming in target cells [4]. The functional roles and molecular compositions of cancerous cell-derived EMVs in response to therapy, metastasis, and tumorigenesis are gradually deciphered [23]. The potential therapeutic approaches and latest achievements in the terms of cancer cell-derived exosomes were elucidated by Kalluri and LeBleu [4].
Isolation and purification methods for exosomes
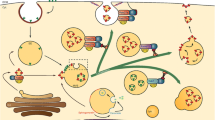
With the rapid progress in technology and science, many methods have been developed for exosome isolation in good purity and quantity. Each method exerts a specific characteristic of exosomes, like their surface proteins, size, shape, and density, to facilitate their isolation [22]. Some of the isolation procedures along with their mechanism of isolation and drawbacks and profits of the method, are summarized in Table 1 and Fig. 1. There are no efficient methods for exosome isolation, and the existing methods either encounter the problem of contamination with proteins and non-exosomal vesicles or are very expensive due to requiring special equipment [52]. The affinity-based methods are very pure; however, their high costs and low yields have prevented its extensive use in exosome purification. Researchers believe that a combination of two methods can boost the efficiency and purity of the isolated exosomes [53]. The field flow fractionation methods have received special attention due to their scalability and isolation of specific subpopulations of exosomes [54]. However, they require further improvement for use in clinical applications.

Schematic diagram of isolation methodology of exosomes. a Size exclusion chromatography which separates particles based on size, is one of the most common methods for obtaining a large volume of exosomes due to the lack of protein contamination and the ability to purify the exosome on a large scale. b Ultracentrifuge separation, despite being dependent on expensive equipment, has been widely used to isolate exosomes based on size and sedimentation properties or density in sucrose gradients. c Microfluidics-based methods rely on physical properties such as size and density, or chemical properties such as binding to exosome surface antigens. d In immunoaffinity methods, exosomes are captured based on their specific binding to antibodies or magnetic nanoparticles. As a result, the surface chemical properties are critical in these techniques. e In ultrafiltration, the particles are centrifuged through the filter and separated based on the pore size of the filter. f In the polymer co-percipitation method, based on steric exclusion, particles are gathered by PEG to form clumps that can be easily precipitated by low-speed centrifugation. g In field flow fractionation, particles accumulate at different position of the membrane depending on their size. Separation occurs when the diffusing and cross-flow forces are balanced
Procedures for the purification and preparation of extracellular vesicles in large amounts include exosome precipitation [55], ultrafiltration [56], sucrose density gradient ultracentrifugation, and differential ultracentrifugation [30]. In a study in which large-scale mRNA-loaded exosomes were prepared, two differential ultracentrifugation and density gradient ultracentrifugation methods were used to purify the exosomes. It was observed that the rate of mRNA recovery is similar in both procedures. At the same time, the chemicals used in the density gradient filtration method may not be removed entirely. Furthermore, more RNAs were concentrated in the exosome fractions when differential ultracentrifugation was used. Hence, differential ultracentrifugation was chosen as a preferable method to purify the exosomes [57].
Methods for loading RNA into exosomes
Exosomes are natural intercellular transporters of RNAs that are responsible for different roles in vivo. Because of their intrinsic features, these EMVs can present a more effective procedure for transfecting RNAs in gene therapy than classic delivery vehicles of nucleic acids [63]. Despite many benefits, the exosome-based RNA delivery is restricted since generating adequate amounts of RNA-loaded exosomes for in vivo application is technically challenging [45, 64]. First, only a few cell sources have been observed to release an adequate quantity of exosomes that are needed for clinical translation [27, 65, 66]. Second, to produce a clinical dose of the EMVs, a great deal of cell culture should be incubated for multiple days, followed by RNA isolating and loading before the ultimate gene-enriched vesicles could be achieved. Although post-insertion of siRNA and shRNA plasmid into exosomes through electroporation technique has shown more remedial efficacies than synthetic nanocarriers in repressing oncogenic targets in the pancreatic cancer preclinical models [66], loading of large RNAs like mRNAs into exosomes remains challenging technically [67]. Herein, our group discussed several approaches to incorporate RNA molecules into exosomes for therapeutic applications and transcriptional manipulations. These approaches include guidance of signature sequences, electroporation, transfecting donor cell, transfection with specific reagents, producing hybrid exosome-liposome and cellular nanoporation [4, 63, 68]. In some loading methods such as electroporation, liposome-exosome hybrids, and transfection with specific reagents, exosomes are first purified from the appropriate sources, and RNA molecules are then loaded into its structure. However, in other techniques such as the guidance of signature sequences and transfecting donor cells, the cells are employed to load nucleic acids by engineering the exosome-producing cells. Therefore, the exosomes secreted from these cells will contain the desired RNA molecules.
Guidance of signature sequences
The actively packaging of nucleic acids as one of the methods for loading nucleic acids into the EMVs has attracted the researcher's attention [63]. Active packaging by proteins binding to special RNAs was used to generate exosomes in engineered mammalian cells [22]. By transfecting and engineering exosome-producing cells, researchers have been able to use the cellular potential to load catalase mRNA into exosomes. According to their results, exosomes can deliver the mRNA cargo to brain cells and significantly attenuate the effects of neurotoxicity and neuro-inflammation in Parkinson’s disease [69]. The conserved sequence of exosome-enriched RNAs (eRNAs) was also presented as a procedure to perform active packaging. The authors proposed that exosome-enriched RNAs might possess a special common sequence that aims RNAs into exosomes as cis-elements. The results revealed that only three motifs are potential signature sequences. The findings help us understand the selective targeting of candidate mRNA to the exosome for future therapeutic research [63, 70].
Electroporation
For a long time, electroporation has been known as an efficient and fast procedure to load siRNAs and miRNAs into the extracellular vesicles. Jing-Hung Wang et al. demonstrated that the electroporation method is applicable for loading siRNA into exosomes. Simultaneously, it is not adequate for mRNA loading because less mRNA entered the electroporated exosomes compared to the non-electroporated ones, indicating that the mRNA remained not internalized. Therefore, they used a new method involving transfecting 293FT cells with a plasmid type to insert HChrR6 mRNA into exosomes [71]. The researchers fused the neuron-specific peptide RVG with exosome membrane protein Lamp2b [60] in another study. The siRNAs were loaded into purified exosomes to make these extracellular vesicles target neuron cells through the electroporation technique [22].
Moreover, Alvarez-Erviti et al. reported that exosomes loaded with BACE1-targeted siRNA prepared via electroporation method can be delivered to the patient's brain to treat Alzheimer's conditions [65]. Overall, exosome transfection with nucleic acids through electroporation is straightforward, but it is not high potent and efficient. This procedure needs purification and separation of extracellular vesicles before and after the transfection process. High-speed centrifugation might destroy exosomes and decrease the quality of samples. It was also found that repeated purification can lead to loss of exosomes [2, 63]. In fact, electroporation is limited to some RNAs, while miRNAs, shRNAs, and mRNAs cannot be loaded to exosomes with this method [72].
Transfecting donor cell
In this method, the parental cells are transfected with siRNAs and miRNAs, which subsequently release into the exosomes via the parental cells [63]. It was found that the secretion of exosomes from fat SCs overexpressing miR126 takes part in a remedial task in relieving myocardial ischemic acute injuries [73]. The exosomes released by HEK293T cells transfected with the miR-214 inhibitor showed the ability to reverse the cisplatin resistance of gastric cancers [71]. In another study, SGC-7901 and HEK293T cells transfected with siRNA and the released HGF siRNAs loaded exosomes were found to be able to suppress the migration and growth of cancer/vascular cells [62]. Moreover, Sun et al., transfected HEK293T cells with the IRESLuc reporter plasmid and demonstrated that about 50–70 mRNA copies loaded per 100 exosomes [74]. In Vakhshiteh's study, it was found that miR-34a-loaded exosomes after overexpression of mesenchymal stem cells were more efficient in inhibiting the proliferation of breast carcinoma cells than liposomes [75]. This method, however, is restricted by inadequate packaging, lacking specificity, and cytotoxicity [63].
Transfection with specific reagents
Transfection with specific reagents was proposed as an alternative approach for loading nucleic acids into exosomes. In an effort, exosomes were obtained from the HeLa cell line through the ultracentrifugation technique. The lipofectamine-combined siRNA AF488 was then incubated with the extracellular vesicles for 25 min. The mixture was washed and ultrafiltered after the transfection process. The siRNA-loaded extracellular vesicles were cultured together with the target cells for one day. Eventually, the authors suggested that the exogenous siRNA molecules can be introduced to the recipient cell by using transfection with specific reagents. The main disadvantage of this method is that the exosome cannot be separated from lipofectamine and it is not clear whether the transfection is the result of lipofectamine or the exosome [76, 77].
Recently, System Biosciences Co. developed a commercial Kit for exosome delivery, namely Exo-Fect™ Exosome Transfection Kit [75]. This company claimed that the researchers can turn exosomes into delivery vehicles with Exo-Fect, enabling the insertion of small molecules, DNAs (including plasmids), and RNAs into isolated exosomes. Their kit has several features, including (1) introduction of various biomolecules directly into isolated exosomes: metabolites and other small molecules; DNAs including plasmids; RNAs including mRNAs, miRNAs, and siRNAs; (2) easy-to-use with a straightforward and fast loading procedure [75].
Producing hybrid exosome with liposome
Directing the CRISPR/Cas to the host cells is vital for genome editing in vivo. The performance of packaging nucleic acids within exosomes, however, is low. Because liposomal particles are made artificially, and drugs are feasibly loaded into these structures, they are widely used in the pharmaceutical industry nowadays. Although the liposomal particles are applied as nano-carriers, the main challenge with these nanovesicles is their low efficiency in cell entry and drug delivery, which exosomes are experts to do. However, the amount of loaded drugs and nucleic acids is meager in the exosome. In this regard, the researchers tried to form a new structure, called the liposome-exosome hybrid, by simply incubating two nanoparticles and inducing membrane fusion to overcome the difficulty of using either of these nano-carriers alone [78,79,80]. Lin et al. developed hybrid exosomes-liposomes and they found that these mixed nanoparticles (NPs) can package the CRISPR-Cas9 expression vector as a large plasmid efficiently. The authors demonstrated that the MSCs can endocytose and express these hybrid NPs. The liposomes had no ability to transfect the SCs alone. Thus, the hybrid NPs of exosome-liposome were proved to deliver CRISPR-Cas9 systems in the SCs efficiently, proposing a potent application of this method in mRNA encapsulation in the future [81].
Cellular nanoporation: accelerating exosome release
The cellular-nanoporation procedure was firstly established for the large-scale production of exosomes containing therapeutic mRNA molecules [57]. Yang et al. transfected different cells with plasmids and provoked the cells with focal/transient electrical motivations, which induce the secretion of EMVs harboring transcribed mRNA. This procedure can generate up to 55-fold more vesicles and achieve up to 1000-fold increment in exosomal mRNA transcripts, even from cells with a low level of EMVs release and without genetic modification of the source cells [57]. Therefore, the cellular nanoporation approach enables the application of the vesicles as a potential mRNA carrier for uses that need transcriptional manipulations.
Therapeutic applications of exosomes
Exosomes with intrinsic therapeutic activity
The substantial remedial and biomedical potential of exosomes was not forecasted when they were at first detected in the 1980s [3, 22]. These membranous vesicles take part in intercellular communications through delivering their content, such as proteins, mRNAs, and miRNAs to target cells, with or without direct contact among cells. In addition, exosomes affect pathological and physiological processes. Owing to the important properties, exosomes can enhance motor and neural functions in the nervous system, allow multiple intravenous dosing without any side effects, cross the blood–brain barriers, and decrease inflammation. These membranous vesicles play a crucial role in diagnosis and prognosis of many pathological conditions like numerous cardiopulmonary disorders, kidney and liver disease, neurodegenerative disorders, and cancer [82].
Current researches have indicated that exosomes derived from different sources are a new remedial tool.
MSCs-derived exosomes
The MSCs-derived exosomes have been assayed in disease models like diabetes mellitus, renal disease, dermatological, gastrointestinal, hepatic, musculoskeletal, neurological, cardiovascular, and respiratory disorders [32]. These vesicles promoted tissue regeneration by improving extracellular matrix remodeling. They prevented the production of the pro-inflammatory cytokines resulting in anti-inflammatory effects [83]. MSCs-derived exosomes were found to have the cytoprotective and immunomodulatory activity of their parent cells [84]. It was shown that exosomes obtained from MSCs could enhance cognition by protecting oxidative damages in astrocytes and neurons in diabetic animals suffered from cognitively impaired conditions [85]. Administration of MSCs-derived exosomes can also decrease infarct sizes in myocardial reperfusion/ischemia models [86, 87]. MSCs-derived exosomes with GATA-4 overexpressing reduced cardiomyocyte apoptosis and increased cardiomyocyte survival [88]. Bone marrow MSCs-isolated vesicles were found to protect against different kinds of diseases like brain injury, hypoxia-derived pulmonary hypertension, and ischemia injury [31, 86, 89]. These MSCs-derived exosomes showed neurorestorative influences, such as an increment in myelin and axon density in rats with diabetes II and a decrement in blood–brain barrier haemorrhage and leakage [90].
Similar to MSCs-derived vesicles, exosomes secreted from cardiac progenitor cells, ESCs, and iPSCs possess therapeutic effects [91]. The intravenous injection of urinary SCs-isolated exosomes can decrease apoptosis and urinary albumin, and increase the glomerular endothelial cell growth in rats treated with streptozotocin, demonstrating exosomes can be a new procedure to treat diabetic nephropathy [92].
Milk-derived exosomes
Milk-derived exosomes have some favorable characteristics, including staying completely intact during gastric digestion, accessible entrance into the blood circulation through endocytosis in the gastrointestinal tract, transferring their protein and miRNA contents to the immune cells, and subsequently regulating immune response and growth [93]. Moreover, bovine milk-derived exosomes can attenuate arthritis [94]. Milk exosomes can also be employed as carriers for oral drug delivery. Agrawal et al. reported that oral administration of Paclitaxel-loaded exosomes showed less toxicity and side effects than intravenous administration [95].
DC-derived exosomes
Intradermal/subcutaneous injections of DC-derived exosomes resulted in stabilizing of objective tumor response or another disease [96]. DC-derived vesicles containing tumor antigens can induce cancer-specific T cell responses [97]. Exosomes derived from dendric cells (DCs) showed a more remarkable ability to produce anti-tumor antibodies than DC cells. Due to the presence of MHC class I / II in the membrane structure of DC-derived exosomes, researchers believe that these vesicles would be used as a very efficient platform in vaccination in the near future [98].
Tumor-derived exosomes
Tumor cell-derived exosomes were documented to induce immunosuppression by stimulating regulatory T cells and immunosuppressive myeloid-derived suppressor cells, suppressing DC differentiation and NK cell cytotoxicity, and inducing T cell apoptosis [99, 100]. While, exosomes that derived from tumor cells under stress condition could induce antitumor immune responses because they contain more HSPs (heat-shock proteins) in addition to tumor antigens. For instance, B16BL6 murine melanoma-originated exosomes incubated with melanoma antigens could induce particular T-cell responses and suppress cancer growth [101].
Exosomes as drug carriers
Exosomes have been potentially used as drug carriers in various diseases and cancers since their discovery due to their unique properties such as stability, tumor targeting, and biocompatibility [102]. Withaferin, doxorubicin, and paclitaxel-loaded exosomes released these agents slowly and suppressed the lung tumor growth in vivo and A549 lung tumor in vitro, effectively [103]. Furthermore, when these agents were loaded into exosomes, they indicated a lower IC50 value than when they were used as free drugs. EL-4 mouse lymphoma-isolated exosomes loaded with curcumin suppressed pro-inflammatory cytokines, like IL-6 and TNF-α [104]. Lower levels of cardiotoxicity along with the higher concentration of intracellular doxorubicin were obtained in doxorubicin-loaded exosomes in contrast to its systemic administration [105]. Similarly, cytotoxicity and high neoplastic tropism were recorded in MDR pulmonary metastases in pursuit of using exosomes encapsulated with Taxol [61].
Exosome-mediated RNA therapeutics
RNAs are large biomolecules and problematic to deliver in vivo. Some delivery systems like cationic polymer-based particles [106], dendrimers [107], and cationic liposomes [108, 109] have been proposed. These carriers, however, are not appropriate for application in clinical practices, because of off-target issues and inadequate stability and safety [110]. Exosomes as lipid-bilayer enclosed, nanometer-sized extracellular vesicles serve as potential nucleic acid carriers owing to their natural affinity to recipient cells and inherent capability of shuttling DNA and RNA between cells [4]. Until now, exosome-based nanocarriers delivering a variety of RNAs—including mRNA, siRNA, and miRNA—have been established to treat different diseases and various cancers [67] (Tables 2 and 3).
Exosomes as carriers for siRNA delivery
siRNA is a biological macromolecule with a polyanionic nature that leads to poor passive uptake. Furthermore, due to the possibility of degradation by nucleases, it is not possible to inject naked siRNA into the systemic circulation. Exosomes have been shown to be an ideal nanocarrier for siRNA encapsulation due to their structure consisting of a hollow, aqueous core surrounded by a phospholipid bilayer, as well as their stability in the blood and inherent targeting properties [111]. El-Andaloussi et al. suggested an approach to utilize exosomes to deliver siRNA molecules in vivo/in vitro to the mouse brain that resulted in a promising output [112]. Moreover, Alvarez-Erviti et al. utilized murine self-derived dendritic exosomes for delivering BACE1 and GAPDH siRNAs across the mouse blood–brain barrier. They observed that the BACE1 and GAPDH gene-loaded exosomes greatly suppressed the gene expression and β-amyloid production in the mouse brain following intravenous injection [65]. Wahlgren et al. observed that human plasma-isolated exosomes loaded with siRNA successfully targeted lymphocytes and monocytes and eventually silenced the MAPK1 gene. They also demonstrated that exosomes loaded with siRNA co-localized in the target cell cytoplasm. Further, RAS52 and RAD51 siRNAs-enriched exosomes induced the death of fibrosarcoma cells, showing that exosomes can be utilized as a vector in the gene therapy based on RNAi. It should be noted that, in the in vitro studies, the target cells take up the EMVs easier than in vivo [76, 113]. Recently researchers illustrated that silencing the CTGF gene via siRNA-loaded MSCs-derived exosomes effectively reduced the inflammation and neuronal apoptosis at the injured area of the spinal cord. Besides, it has been shown that loading miRNA into the exosome structure can easily induce some processes such as promoting cancer cell apoptosis, regulating lipid metabolism, and promoting angiogenesis [114].
Exosomes as carriers for miRNA delivery
miRNA is a small molecule with a low molecular weight that, in addition to being easily transfected into the exosome, remains stable inside it and is, therefore, able to travel long distances without being degraded by nucleases in the blood [115]. It has been shown that by loading miRNA into the exosome structure, processes such as promoting cancer cell apoptosis, regulating lipid metabolism and promoting angiogenesis can be easily induced [116]. Ohno et al. delivered the GE11-targeted exosomes containing miRNA let-7a to EGF-overexpressing breast tumors in mice. GE11-targeted exosomes showed higher tumor suppression than control. Furthermore, the donor cells-derived exosomes transfected with the miRNA suppressed cancer cells [72]. It was reported that miR-122 overexpression in exosomes obtained from adipose MSCs can increase chemotherapy sensitivity and inhibit carcinoma growth in the mouse model [117]. Similarly, miR146b-enriched exosome is capable of suppressing EGFR and prevents tumor growth in glioma rats [118]. Moreover, the miR-451/144-enriched cardiac progenitor cell-derived exosomes promoted cardioprotection through enhancing cardiomyocyte survival in H9c2 cells, in vitro, and in the myocardial reperfusion/ischemia model in vivo [119].
Moreover, other forms of RNAi, such as miRNA inhibitors and miRNA mimics can be loaded into the EMVs. Mahati et al. displayed that (ScFv)-modified exosomes derived from human cord blood MSCs can be used to deliver the anti-tumor effect miR-26a mimics into the GPC3-positive hepatocellular carcinoma cells with no side effects. [120]. Exosomal miRNA delivery would be especially useful when several processes are targeted, as in the case of cancers [121, 122] and Alzheimer’s disease (miR29) [123].
Exosomes as carriers for mRNA delivery
Exosome-mediated mRNA delivery for COVID-19 vaccination has been recently reported. Tsai et al. validated the application of exosome for delivering the intended mRNA into the host cell in vivo and in vitro, and further specifically, developed the LSNME-SW1 vaccine, which activated broad immunities to COVID-19 [124]. Lipid nanoparticles (LNP) are currently considered one of the safest and most efficient delivery systems for mRNA, and this lipid composition has been employed in Pfizer-BioNTech and Moderna mRNA vaccines. However, some recent studies have claimed that mRNA-loaded exosomes are much more efficient than mRNA-loaded LNPs in delivering mRNA to target cells in vitro [52, 124, 125]. Moreover, it was found that mRNA-harboring exosomes can increase survival, enhance tumor inhibition, and restore tumor suppressor function in PTEN-deficient glioma mice [68]. In another study, it was shown that exosomes loaded with engineered mRNA translationally activated by corresponding miRNAs in the target cells could increase efficacy while reducing off-target uptake [74]. The results mentioned above propose that exosome is an essential tool for gene delivery with an acceptable safety profile than those of polymer-based particles, cationic lipids, and viral vectors [126].
Challenges in RNA delivery by exosomes
The major challenges in RNA-based therapies are the instability of the RNA structure in blood circulation due to their rapid removal by nucleases and their high immunogenicity; [134], to overcome this problem, a wide variety of nanocarriers have been introduced [135]. Some delivery vehicles, such as polymers, could not be developed due to toxicity, [136], while others, such as liposomes, have not been very efficient in vivo due to poor loading efficiency and ability to penetrate the cell [137, 138]. Lipid nanoparticles (LNPs) with low toxicity and extremely high efficiency in delivering mRNA to the target cell have recently piqued the interest of all researchers [139, 140]. Exosomes, on the other hand, appear to be serious competitors for LNPs, according to some evidence [52, 125]. Because of their good biocompatibility, low immunogenicity, and ability to cross biological barriers such as the BBB, they offer many advantages as RNA delivery vehicles [141, 142]. Despite all of the benefits, the large-scale production and purification of exosomes, as well as the standard and efficient method for RNA loading, are extremely difficult and significant concerns.
Large scale exosome production
One of the main challenges in using exosomes in the clinic is the lack of access to an approved method for large-scale cell culture and production of exosomes with constant characteristics and properties. The presence of exosomes in the serums used in the cell culture media and the effect of its removal on the cell proliferation and consequently the property and biology of cell-derived exosomes has caused severe problems in the mass production of exosomes [143, 144].
Exosome purification
Almost all exosome purification methods commonly used today have either poor purity, such as methods based on volume-excluding polymers due to co-precipitation of other non-exosomal contaminants, or low yields and high costs, such as ultracentrifugation and immunoaffinity-based methods. There is no perfect purification technique today based on purity and scalability. Thus, it is another major problem in using exosomes as delivery systems [143]. Recently, there have been hopes for using the tangential flow filtration (TFF) method for reproducible large-scale exosome purification [145]. Therefore, an efficient method and good manufacturing practice (GMP) guidelines should be developed for exosomes purification [144].
Exosome analysis
Very precise and reliable techniques are essential for characterization and detection to assure the quality of the delivery system. The analysis and characterization process of exosomes face significant obstacles due to their small size (40 to 200 nm) and insufficient information about their structure and surface components [52, 146].
Long RNA loading
Exosomes are very successful in delivering miRNA and siRNA, unlike mRNA, which is very difficult to be loaded in the exosome due to their length and charge. Some mRNA loading methods were mentioned in the present study, but further studies should be performed to achieve more satisfactory and practical results in loading mRNA in exosomes [52, 147].
Conclusion
Exosomes, a subset of extracellular vesicles, offer several benefits as drugs/biomolecules carriers over other drug nanocarriers, including the capability of loading different cargos and crossing impermeable biological barriers, facilitation of the cellular internalization of the cargos via endocytosis or membrane fusion, long half-life and circulation time, low immunogenicity, and small size [4]. These bio-inspired nanocarriers can deliver many cargos like proteins, small molecules, and nucleic acids such as DNA, siRNA, miRNA, and mRNA [22]. mRNAs play an essential role in the disease treatment and vaccine production and exosomes, owing to unique properties, can open up a new way for researchers in the development of a safe and efficient mRNA delivery system [148]. Although abundant evidence shows that lipid nanoparticles (LNPs) are highly efficient for mRNA transfer, some studies show that exosomes are more stable and less immunogenic than LNPs [52]. Recent in vitro studies have shown that transferring the CRISPR/Cas9-based RNA reporter system by exosomes has been much more satisfactory than the LNP [125]. However, due to the various challenges of using exosomes in the clinic, this extremely stable and efficient nano-carrier has not been employed in treatments yet [144]. One of the major problems is the lack of a standard and reliable method for exosome purification [149]. Although several strategies have been performed to load short nucleic acids like siRNA, shRNA, and DNA into exosomes [66, 68, 71, 72, 74], the efficient encapsulation of large mRNAs into exosomes remains a challenge. Therefore, the establishment of large-scale RNA-loaded exosomes for clinical applications should be given further attention.
Availability of data and materials
Not applicable.
Abbreviations
- AAT:
-
Alpha-1-antitrypsin
- AATD:
-
Alpha-1 antitrypsin deficiency
- CD:
-
Cytosine deaminase
- Dox:
-
Doxorubicin
- EPO:
-
Encoding erythropoietin
- ExoPAC:
-
Exosomes for oral delivery of paclitaxel
- EVs:
-
Extracellular vesicles
- 5-FC:
-
5-Fluorocytosine
- 2FdU:
-
2ʹ Fluoro-deoxyuridine
- 5-FU:
-
5-Fluorouracil
- HepG2:
-
Human hepatic cell line
- 5hmC:
-
5-Hydroxymethylcytidine
- IVT:
-
In vitro-transcribed mRNA
- imDCs:
-
Immature dendritic cells
- LGP2:
-
Laboratory of genetics and physiology 2
- MVBs:
-
Multi-vesicular bodies
- MDA5:
-
Melanoma differentiation-associated protein 5
- 5moC:
-
5-Methoxycytidine
- MPV:
-
Mean platelet volume
- mRNA:
-
Messenger RNAs
- MSCs:
-
Mesenchymal stem cells
- MVs:
-
Microvesicles
- PLT:
-
Platelet counts
- PRRs:
-
Pattern recognition receptors
- RBCs:
-
Red blood cells
- RBCEVs:
-
RBC extracellular vesicles
- RIG-1:
-
Retinoic acid-inducible protein 1
- RLRs:
-
RIG-I-like receptors
- TFs:
-
Transcription factors
- TLR:
-
Toll-like receptor
- UPRT:
-
Uracil phosphoribosyltransferase
- UTR:
-
Untranslated region
- VEGF-A:
-
Vascular endothelial growth factor-A
References
Vlassov AV, Magdaleno S, Setterquist R, Conrad R. Exosomes: current knowledge of their composition, biological functions, and diagnostic and therapeutic potentials. Biochimica et Biophysica Acta BBA General Subjects. 2012;1820(7):940–8.
Schorey JS, Harding CV. Extracellular vesicles and infectious diseases: new complexity to an old story. J Clin Investig. 2016;126(4):1181–9.
Pegtel DM, Gould SJ. Exosomes. Annu Rev Biochem. 2019;88(1):487–514.
Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367:6478.
Subra C, Grand D, Laulagnier K, Stella A, Lambeau G, Paillasse M, et al. Exosomes account for vesicle-mediated transcellular transport of activatable phospholipases and prostaglandins. J Lipid Res. 2010;51(8):2105–20.
Mathivanan S, Simpson RJ. ExoCarta: A compendium of exosomal proteins and RNA. Proteomics. 2009;9(21):4997–5000.
Subra C, Laulagnier K, Perret B, Record M. Exosome lipidomics unravels lipid sorting at the level of multivesicular bodies. Biochimie. 2007;89(2):205–12.
Zhang Y, Liu Y, Liu H, Tang WH. Exosomes: biogenesis, biologic function and clinical potential. Cell Biosci. 2019;9(1):1–18.
Michael A, Bajracharya SD, Yuen PS, Zhou H, Star RA, Illei GG, et al. Exosomes from human saliva as a source of microRNA biomarkers. Oral Dis. 2010;16(1):34–8.
Rabinowits G, Gerçel-Taylor C, Day JM, Taylor DD, Kloecker GH. Exosomal microRNA: a diagnostic marker for lung cancer. Clin Lung Cancer. 2009;10(1):42–6.
Skog J, Würdinger T, Van Rijn S, Meijer DH, Gainche L, Curry WT, et al. Glioblastoma microvesicles transport RNA and proteins that promote tumour growth and provide diagnostic biomarkers. Nat Cell Biol. 2008;10(12):1470–6.
Ratajczak J, Wysoczynski M, Hayek F, Janowska-Wieczorek A, Ratajczak M. Membrane-derived microvesicles: important and underappreciated mediators of cell-to-cell communication. Leukemia. 2006;20(9):1487–95.
Valadi H, Ekström K, Bossios A, Sjöstrand M, Lee JJ, Lötvall JO. Exosome-mediated transfer of mRNAs and microRNAs is a novel mechanism of genetic exchange between cells. Nat Cell Biol. 2007;9(6):654–9.
NoltetHoen EN, Buermans HP, Waasdorp M, Stoorvogel W, Wauben MH, Hoen PA. Deep sequencing of RNA from immune cell-derived vesicles uncovers the selective incorporation of small non-coding RNA biotypes with potential regulatory functions. Nucleic Acids Res. 2012;40(18):9272–85.
Aslan C, Kiaie SH, Zolbanin NM, Lotfinejad P, Ramezani R, Kashanchi F, Jafari R. Exosomes for mRNA delivery: A novel biotherapeutic strategy with hurdles and hope. BMC Biotechnol. 2021;21(1):1–2.
Bellingham SA, Coleman BM, Hill AF. Small RNA deep sequencing reveals a distinct miRNA signature released in exosomes from prion-infected neuronal cells. Nucleic Acids Res. 2012;40(21):10937–49.
Zeringer E, Li M, Barta T, Schageman J, Pedersen KW, Neurauter A, et al. Methods for the extraction and RNA profiling of exosomes. World J Methodol. 2013;3(1):11.
Zhang M, Zang X, Wang M, Li Z, Qiao M, Hu H, et al. Exosome-based nanocarriers as bio-inspired and versatile vehicles for drug delivery: recent advances and challenges. J Mater Chem B. 2019;7(15):2421–33.
Schageman J, Zeringer E, Li M, Barta T, Lea K, Gu J, et al. The complete exosome workflow solution: from isolation to characterization of RNA cargo. BioMed Res Int. 2013;2013:89.
Sabzehzari M, Naghavi M. Phyto-miRNA: a molecule with beneficial abilities for plant biotechnology. Gene. 2019;683:28–34.
Sabzehzari M, Naghavi M. Phyto-miRNAs-based regulation of metabolites biosynthesis in medicinal plants. Gene. 2019;682:13–24.
Mashouri L, Yousefi H, Aref AR, MohammadAhadi A, Molaei F, Alahari SK. Exosomes: composition, biogenesis, and mechanisms in cancer metastasis and drug resistance. Mol Cancer. 2019;18(1):75.
Bunggulawa EJ, Wang W, Yin T, Wang N, Durkan C, Wang Y, et al. Recent advancements in the use of exosomes as drug delivery systems. J Nanobiotechnol. 2018;16(1):1–13.
Zitvogel L, Regnault A, Lozier A, Wolfers J, Flament C, Tenza D, et al. Eradication of established murine tumors using a novel cell-free vaccine: dendritic cell derived exosomes. Nat Med. 1998;4(5):594–600.
Szabo G. Momen-Heravi F. Extracellular Vesicles and Exosomes: Biology and Pathobiology. Liver; 2020. p. 1022–7.
Khatun Z, Bhat A, Sharma S, Sharma A. Elucidating diversity of exosomes: biophysical and molecular characterization methods. Nanomedicine. 2016;11(17):2359–77.
Andaloussi SE, Mäger I, Breakefield XO, Wood MJ. Extracellular vesicles: biology and emerging therapeutic opportunities. Nat Rev Drug Discovery. 2013;12(5):347–57.
Vella L, Sharples R, Lawson V, Masters C, Cappai R, Hill A. Packaging of prions into exosomes is associated with a novel pathway of PrP processing. J Pathol. 2007;211(5):582–90.
Théry C, Ostrowski M, Segura E. Membrane vesicles as conveyors of immune responses. Nat Rev Immunol. 2009;9(8):581–93.
Théry C, Amigorena S, Raposo G, Clayton A. Isolation and characterization of exosomes from cell culture supernatants and biological fluids. Curr Protocols Cell Biol. 2006;30(1):322.
Doeppner TR, Herz J, Görgens A, Schlechter J, Ludwig A-K, Radtke S, et al. Extracellular vesicles improve post-stroke neuroregeneration and prevent postischemic immunosuppression. Stem Cells Transl Med. 2015;4(10):1131–43.
Willis GR, Kourembanas S, Mitsialis SA. Toward exosome-based therapeutics: isolation, heterogeneity, and fit-for-purpose potency. Front Cardiovasc Med. 2017;4:63.
Andaloussi SE, Lakhal S, Mäger I, Wood MJ. Exosomes for targeted siRNA delivery across biological barriers. Adv Drug Deliv Rev. 2013;65(3):391–7.
Trams EG, Lauter CJ, Salem JN, Heine U. Exfoliation of membrane ecto-enzymes in the form of micro-vesicles. Biochimica et Biophysica Acta (BBA) Biomembranes. 1981;645(1):63–70.
Pan B-T, Teng K, Wu C, Adam M, Johnstone RM. Electron microscopic evidence for externalization of the transferrin receptor in vesicular form in sheep reticulocytes. J Cell Biol. 1985;101(3):942–8.
Simons M, Raposo G. Exosomes–vesicular carriers for intercellular communication. Curr Opin Cell Biol. 2009;21(4):575–81.
Masyuk AI, Huang BQ, Ward CJ, Gradilone SA, Banales JM, Masyuk TV, et al. Biliary exosomes influence cholangiocyte regulatory mechanisms and proliferation through interaction with primary cilia. Am J Physiol-Gastrointest Liver Physiol. 2010;299(4):G990–9.
Asea A, Jean-Pierre C, Kaur P, Rao P, Linhares IM, Skupski D, et al. Heat shock protein-containing exosomes in mid-trimester amniotic fluids. J Reprod Immunol. 2008;79(1):12–7.
Andre F, Schartz NE, Movassagh M, Flament C, Pautier P, Morice P, et al. Malignant effusions and immunogenic tumour-derived exosomes. Lancet. 2002;360(9329):295–305.
Admyre C, Johansson SM, Qazi KR, Filén J-J, Lahesmaa R, Norman M, et al. Exosomes with immune modulatory features are present in human breast milk. J Immunol. 2007;179(3):1969–78.
Pisitkun T, Shen R-F, Knepper MA. Identification and proteomic profiling of exosomes in human urine. Proc Natl Acad Sci. 2004;101(36):13368–73.
Aalberts M, van Dissel-Emiliani FM, van Adrichem NP, van Wijnen M, Wauben MH, Stout TA, et al. Identification of distinct populations of prostasomes that differentially express prostate stem cell antigen, annexin A1, and GLIPR2 in humans. Biol Reprod. 2012;86(3):82.
Ogawa Y, Miura Y, Harazono A, Kanai-Azuma M, Akimoto Y, Kawakami H, et al. Proteomic analysis of two types of exosomes in human whole saliva. Biol Pharm Bull. 2011;34(1):13–23.
Caby M-P, Lankar D, Vincendeau-Scherrer C, Raposo G, Bonnerot C. Exosomal-like vesicles are present in human blood plasma. Int Immunol. 2005;17(7):879–87.
Yeo RWY, Lai RC, Zhang B, Tan SS, Yin Y, Teh BJ, et al. Mesenchymal stem cell: an efficient mass producer of exosomes for drug delivery. Adv Drug Deliv Rev. 2013;65(3):336–41.
Burke J, Kolhe R, Hunter M, Isales C, Hamrick M, Fulzele S. Stem cell-derived exosomes: a potential alternative therapeutic agent in orthopaedics. Stem Cells Int. 2016;2016:89.
Zempleni J, Sukreet S, Zhou F, Wu D, Mutai E. Milk-derived exosomes and metabolic regulation. Ann Rev Anim Biosci. 2019;7:245–62.
Reif S, Shiff YE, Golan-Gerstl R. Milk-derived exosomes (MDEs) have a different biological effect on normal fetal colon epithelial cells compared to colon tumor cells in a miRNA-dependent manner. J Transl Med. 2019;17(1):1–10.
Woith E, Melzig MF. Extracellular vesicles from fresh and dried plants—simultaneous purification and visualization using gel electrophoresis. Int J Mol Sci. 2019;20(2):357.
Patel GK, Khan MA, Zubair H, Srivastava SK, Singh S, Singh AP. Comparative analysis of exosome isolation methods using culture supernatant for optimum yield, purity and downstream applications. Sci Rep. 2019;9(1):1–10.
Kharaziha P, Ceder S, Li Q, Panaretakis T. Tumor cell-derived exosomes: a message in a bottle. Biochem Biophys Acta. 2012;1826(1):103–11.
Lundberg S, Karlsson E, Dahlberg H, Glansk M, Larsson S, Larsson S, et al. Exosomes and lipid nanoparticles - the future of targeted drug delivery [Student thesis]; 2020.
Stam J, Bartel S, Bischoff R, Wolters JC. Isolation of extracellular vesicles with combined enrichment methods. J Chromatogr B. 2021;1169:122604.
Sitar S, Kejžar A, Pahovnik D, Kogej K, Tušek-Žnidarič M, Lenassi M, et al. Size characterization and quantification of exosomes by asymmetrical-flow field-flow fractionation. Anal Chem. 2015;87(18):9225–33.
Niu Z, Pang RT, Liu W, Li Q, Cheng R, Yeung WS. Polymer-based precipitation preserves biological activities of extracellular vesicles from an endometrial cell line. PLoS ONE. 2017;12(10):e0186534.
Heinemann ML, Vykoukal J. Sequential filtration: A gentle method for the isolation of functional extracellular vesicles. Extracellular Vesicles: Springer; 2017. p. 33–41.
Yang Z, Shi J, Xie J, Wang Y, Sun J, Liu T, et al. Large-scale generation of functional mRNA-encapsulating exosomes via cellular nanoporation. Nat Biomed Eng. 2020;4(1):69–83.
Lozano-Ramos I, Bancu I, Oliveira-Tercero A, Armengol MP, Menezes-Neto A, Portillo HAD, et al. Size-exclusion chromatography-based enrichment of extracellular vesicles from urine samples. J Extracell Vesicles. 2015;4(1):27369.
Liu C, Guo J, Tian F, Yang N, Yan F, Ding Y, et al. Field-free isolation of exosomes from extracellular vesicles by microfluidic viscoelastic flows. ACS Nano. 2017;11(7):6968–76.
Yoo CE, Kim G, Kim M, Park D, Kang HJ, Lee M, et al. A direct extraction method for microRNAs from exosomes captured by immunoaffinity beads. Anal Biochem. 2012;431(2):96–8.
Kim MS, Haney MJ, Zhao Y, Mahajan V, Deygen I, Klyachko NL, et al. Development of exosome-encapsulated paclitaxel to overcome MDR in cancer cells. Nanomedicine. 2016;12(3):655–64.
Zhang H, Wang Y, Bai M, Wang J, Zhu K, Liu R, et al. Exosomes serve as nanoparticles to suppress tumor growth and angiogenesis in gastric cancer by delivering hepatocyte growth factor si RNA. Cancer Sci. 2018;109(3):629–41.
Liu C, Su C. Design strategies and application progress of therapeutic exosomes. Theranostics. 2019;9(4):1015–28.
Stewart MP, Sharei A, Ding X, Sahay G, Langer R, Jensen KF. In vitro and ex vivo strategies for intracellular delivery. Nature. 2016;538(7624):183–92.
Alvarez-Erviti L, Seow Y, Yin H, Betts C, Lakhal S, Wood MJ. Delivery of siRNA to the mouse brain by systemic injection of targeted exosomes. Nat Biotechnol. 2011;29(4):341–5.
Kamerkar S, LeBleu VS, Sugimoto H, Yang S, Ruivo CF, Melo SA, et al. Exosomes facilitate therapeutic targeting of oncogenic KRAS in pancreatic cancer. Nature. 2017;546(7659):498–503.
Usman WM, Pham TC, Kwok YY, Vu LT, Ma V, Peng B, et al. Efficient RNA drug delivery using red blood cell extracellular vesicles. Nat Commun. 2018;9(1):1–15.
Yang T, Fogarty B, LaForge B, Aziz S, Pham T, Lai L, et al. Delivery of small interfering RNA to inhibit vascular endothelial growth factor in zebrafish using natural brain endothelia cell-secreted exosome nanovesicles for the treatment of brain cancer. AAPS J. 2017;19(2):475–86.
Kojima R, Bojar D, Rizzi G, Charpin-El Hamri G, El-Baba MD, Saxena P, et al. Designer exosomes produced by implanted cells intracerebrally deliver therapeutic cargo for Parkinson’s disease treatment. Nat Commun. 2018;9(1):1–10.
Batagov AO, Kuznetsov VA, Kurochkin IV. Identification of nucleotide patterns enriched in secreted RNAs as putative cis-acting elements targeting them to exosome nano-vesicles. New York: Springer; 2011.
Wang J-H, Forterre AV, Zhao J, Frimannsson DO, Delcayre A, Antes TJ, et al. Anti-HER2 scFv-directed extracellular vesicle-mediated mRNA-based gene delivery inhibits growth of HER2-positive human breast tumor xenografts by prodrug activation. Mol Cancer Ther. 2018;17(5):1133–42.
Ohno S-I, Takanashi M, Sudo K, Ueda S, Ishikawa A, Matsuyama N, et al. Systemically injected exosomes targeted to EGFR deliver antitumor microRNA to breast cancer cells. Mol Ther. 2013;21(1):185–91.
Luo Q, Guo D, Liu G, Chen G, Hang M, Jin M. Exosomes from MiR-126-overexpressing adscs are therapeutic in relieving acute myocardial ischaemic injury. Cell Physiol Biochem. 2017;44(6):2105–16.
Sun W, Xing C, Zhao L, Zhao P, Yang G, Yuan L. Ultrasound Assisted Exosomal Delivery of Tissue Responsive mRNA for enhanced efficacy and minimized off-target effects. Mol Ther Nucleic Acids. 2020;20:558–67.
Vakhshiteh F, Rahmani S, Ostad SN, Madjd Z, Dinarvand R, Atyabi F. Exosomes derived from miR-34a-overexpressing mesenchymal stem cells inhibit in vitro tumor growth: A new approach for drug delivery. Life Sci. 2021;266:118871.
Shtam TA, Kovalev RA, Varfolomeeva EY, Makarov EM, Kil YV, Filatov MV. Exosomes are natural carriers of exogenous siRNA to human cells in vitro. Cell Commun Signal. 2013;11(1):1–10.
Li S-p, Lin Z-x, Jiang X-y, Yu X-y. Exosomal cargo-loading and synthetic exosome-mimics as potential therapeutic tools. Acta Pharmacol Sin. 2018;39(4):542–51.
Sato YT, Umezaki K, Sawada S, Mukai S-A, Sasaki Y, Harada N, et al. Engineering hybrid exosomes by membrane fusion with liposomes. Sci Rep. 2016;6(1):21933.
Li Y-J, Wu J-Y, Liu J, Xu W, Qiu X, Huang S, et al. Artificial exosomes for translational nanomedicine. J Nanobiotechnol. 2021;19(1):242.
Evers MJW, van de Wakker SI, de Groot EM, de Jong OG, Gitz-François JJJ, Seinen CS, et al. Functional siRNA delivery by extracellular vesicle-liposome hybrid nanoparticles. Adv Healthcare Mater. 2021;11:2101202.
Lin Y, Wu J, Gu W, Huang Y, Tong Z, Huang L, et al. Exosome-Liposome Hybrid Nanoparticles Deliver CRISPR/Cas9 System in MSCs. Adv Sci. 2018;5(4):1700611.
Beuzelin D, Kaeffer B. Exosomes and miRNA-loaded biomimetic nanovehicles, a focus on their potentials preventing type-2 diabetes linked to metabolic syndrome. Front Immunol. 2018;9:2711.
Lou G, Chen Z, Zheng M, Liu Y. Mesenchymal stem cell-derived exosomes as a new therapeutic strategy for liver diseases. Exper Mol Med. 2017;49(6):e346.
Baglìo SR, Devescovi V, Granchi D, Baldini N. MicroRNA expression profiling of human bone marrow mesenchymal stem cells during osteogenic differentiation reveals Osterix regulation by miR-31. Gene. 2013;527(1):321–31.
Nakano M, Nagaishi K, Konari N, Saito Y, Chikenji T, Mizue Y, et al. Bone marrow-derived mesenchymal stem cells improve diabetes-induced cognitive impairment by exosome transfer into damaged neurons and astrocytes. Sci Rep. 2016;6(1):1–14.
Lai RC, Arslan F, Lee MM, Sze NSK, Choo A, Chen TS, et al. Exosome secreted by MSC reduces myocardial ischemia/reperfusion injury. Stem Cell Res. 2010;4(3):214–22.
Arslan F, Lai RC, Smeets MB, Akeroyd L, Choo A, Aguor EN, et al. Mesenchymal stem cell-derived exosomes increase ATP levels, decrease oxidative stress and activate PI3K/Akt pathway to enhance myocardial viability and prevent adverse remodeling after myocardial ischemia/reperfusion injury. Stem Cell Res. 2013;10(3):301–12.
Yu B, Kim HW, Gong M, Wang J, Millard RW, Wang Y, et al. Exosomes secreted from GATA-4 overexpressing mesenchymal stem cells serve as a reservoir of anti-apoptotic microRNAs for cardioprotection. Int J Cardiol. 2015;182:349–60.
Lee C, Mitsialis SA, Aslam M, Vitali SH, Vergadi E, Konstantinou G, et al. Exosomes mediate the cytoprotective action of mesenchymal stromal cells on hypoxia-induced pulmonary hypertension. Circulation. 2012;126(22):2601–11.
Venkat P, Chopp M, Zacharek A, Cui C, Zhang L, Li Q, et al. White matter damage and glymphatic dysfunction in a model of vascular dementia in rats with no prior vascular pathologies. Neurobiol Aging. 2017;50:96–106.
Vrijsen KR, Maring JA, Chamuleau SA, Verhage V, Mol EA, Deddens JC, et al. Exosomes from cardiomyocyte progenitor cells and mesenchymal stem cells stimulate angiogenesis via EMMPRIN. Adv Healthcare Mater. 2016;5(19):2555–65.
Jiang Z-Z, Liu Y-M, Niu X, Yin J-Y, Hu B, Guo S-C, et al. Exosomes secreted by human urine-derived stem cells could prevent kidney complications from type I diabetes in rats. Stem Cell Res Ther. 2016;7(1):1–13.
Samuel M, Chisanga D, Liem M, Keerthikumar S, Anand S, Ang C-S, et al. Bovine milk-derived exosomes from colostrum are enriched with proteins implicated in immune response and growth. Sci Rep. 2017;7(1):5933.
Arntz OJ, Pieters BC, Oliveira MC, Broeren MG, Bennink MB, de Vries M, et al. Oral administration of bovine milk derived extracellular vesicles attenuates arthritis in two mouse models. Mol Nutr Food Res. 2015;59(9):1701–12.
Agrawal AK, Aqil F, Jeyabalan J, Spencer WA, Beck J, Gachuki BW, et al. Milk-derived exosomes for oral delivery of paclitaxel. Nanomedicine. 2017;13(5):1627–36.
Lamparski HG, Metha-Damani A, Yao J-Y, Patel S, Hsu D-H, Ruegg C, et al. Production and characterization of clinical grade exosomes derived from dendritic cells. J Immunol Methods. 2002;270(2):211–26.
André F, Chaput N, Schartz NE, Flament C, Aubert N, Bernard J, et al. Exosomes as potent cell-free peptide-based vaccine. I. Dendritic cell-derived exosomes transfer functional MHC class I/peptide complexes to dendritic cells. J Immunol. 2004;172(4):2126–36.
Yao Y, Fu C, Zhou L, Mi Q-S, Jiang A. DC-Derived Exosomes for Cancer Immunotherapy Cancers. 2021;13(15):3667.
Gutiérrez-Vázquez C, Villarroya-Beltri C, Mittelbrunn M, Sánchez-Madrid F. Transfer of extracellular vesicles during immune cell-cell interactions. Immunol Rev. 2013;251(1):125–42.
Kiaie SH, Sanaei MJ, Heshmati M, Asadzadeh Z, Azimi I, Hadidi S, Jafari R, Baradaran B. Immune checkpoints in targeted-immunotherapy of pancreatic cancer: New hope for clinical development. Acta Pharm Sinica B. 2021;11(5):1083–97.
Morishita M, Takahashi Y, Matsumoto A, Nishikawa M, Takakura Y. Exosome-based tumor antigens–adjuvant co-delivery utilizing genetically engineered tumor cell-derived exosomes with immunostimulatory CpG DNA. Biomaterials. 2016;111:55–65.
Patil SM, Sawant SS, Kunda NK. Exosomes as drug delivery systems: a brief overview and progress update. Eur J Pharm Biopharm. 2020;154:259–69.
Munagala R, Aqil F, Jeyabalan J, Gupta RC. Bovine milk-derived exosomes for drug delivery. Cancer Lett. 2016;371(1):48–61.
Sun D, Zhuang X, Xiang X, Liu Y, Zhang S, Liu C, et al. A novel nanoparticle drug delivery system: the anti-inflammatory activity of curcumin is enhanced when encapsulated in exosomes. Mol Ther. 2010;18(9):1606–14.
Hadla M, Palazzolo S, Corona G, Caligiuri I, Canzonieri V, Toffoli G, et al. Exosomes increase the therapeutic index of doxorubicin in breast and ovarian cancer mouse models. Nanomedicine. 2016;11(18):2431–41.
Thomas M, Lu JJ, Ge Q, Zhang C, Chen J, Klibanov AM. Full deacylation of polyethylenimine dramatically boosts its gene delivery efficiency and specificity to mouse lung. Proc Natl Acad Sci. 2005;102(16):5679–84.
Patil ML, Zhang M, Taratula O, Garbuzenko OB, He H, Minko T. Internally cationic polyamidoamine PAMAM-OH dendrimers for siRNA delivery: effect of the degree of quaternization and cancer targeting. Biomacromol. 2009;10(2):258–66.
Yano J, Hirabayashi K, Nakagawa S-I, Yamaguchi T, Nogawa M, Kashimori I, et al. Antitumor activity of small interfering RNA/cationic liposome complex in mouse models of cancer. Clin Cancer Res. 2004;10(22):7721–6.
Sioud M, Sørensen DR. Cationic liposome-mediated delivery of siRNAs in adult mice. Biochem Biophys Res Commun. 2003;312(4):1220–5.
Kesharwani P, Gajbhiye V, Jain NK. A review of nanocarriers for the delivery of small interfering RNA. Biomaterials. 2012;33(29):7138–50.
Faruqu FN, Xu L, Al-Jamal KT. Preparation of exosomes for siRNA delivery to cancer cells. J Visual Exp. 2018;142:e58814.
El-Andaloussi S, Lee Y, Lakhal-Littleton S, Li J, Seow Y, Gardiner C, et al. Exosome-mediated delivery of siRNA in vitro and in vivo. Nat Protoc. 2012;7(12):2112.
Wahlgren J, Karlson TDL, Brisslert M, Vaziri Sani F, Telemo E, Sunnerhagen P, et al. Plasma exosomes can deliver exogenous short interfering RNA to monocytes and lymphocytes. Nucleic Acids Res. 2012;40(17):e130.
Huang W, Qu M, Li L, Liu T, Lin M, Yu X. SiRNA in MSC-derived exosomes silences CTGF gene for locomotor recovery in spinal cord injury rats. Stem Cell Res Ther. 2021;12(1):334.
Munir J, Yoon JK, Ryu S. Therapeutic miRNA-enriched extracellular vesicles: current approaches and future prospects. Cells. 2020;9(10):2271.
Fang Z, Zhang X, Huang H, Wu J. Exosome based miRNA delivery strategy for disease treatment. Chin Chem Lett. 2021. https://doi.org/10.1016/j.cclet.2021.11.050.
Lou G, Song X, Yang F, Wu S, Wang J, Chen Z, et al. Exosomes derived from miR-122-modified adipose tissue-derived MSCs increase chemosensitivity of hepatocellular carcinoma. J Hematol Oncol. 2015;8(1):1–11.
Katakowski M, Buller B, Zheng X, Lu Y, Rogers T, Osobamiro O, et al. Exosomes from marrow stromal cells expressing miR-146b inhibit glioma growth. Cancer Lett. 2013;335(1):201–4.
Chen L, Wang Y, Pan Y, Zhang L, Shen C, Qin G, et al. Cardiac progenitor-derived exosomes protect ischemic myocardium from acute ischemia/reperfusion injury. Biochem Biophys Res Commun. 2013;431(3):566–71.
Mahati S, Fu X, Ma X, Zhang H, Xiao L. Delivery of miR-26a using an exosomes-based nanosystem inhibited proliferation of hepatocellular carcinoma. Cancer. 2021;8:847.
Godlewski J, Nowicki MO, Bronisz A, Williams S, Otsuki A, Nuovo G, et al. Targeting of the Bmi-1 oncogene/stem cell renewal factor by microRNA-128 inhibits glioma proliferation and self-renewal. Can Res. 2008;68(22):9125–30.
Kefas B, Godlewski J, Comeau L, Li Y, Abounader R, Hawkinson M, et al. microRNA-7 inhibits the epidermal growth factor receptor and the Akt pathway and is down-regulated in glioblastoma. Can Res. 2008;68(10):3566–72.
Bettens K, Brouwers N, Engelborghs S, Van Miegroet H, De Deyn PP, Theuns J, et al. APP and BACE1 miRNA genetic variability has no major role in risk for Alzheimer disease. Hum Mutat. 2009;30(8):1207–13.
Tsai S-J, Guo C, Atai NA, Gould SJ. Exosome-Mediated mRNA Delivery For SARS-CoV-2 Vaccination. bioRxiv. 2020.
Murphy DE, de Jong OG, Evers MJW, Nurazizah M, Schiffelers RM, Vader P. Natural or Synthetic RNA delivery: a stoichiometric comparison of extracellular vesicles and synthetic nanoparticles. Nano Lett. 2021;21(4):1888–95.
Luan X, Sansanaphongpricha K, Myers I, Chen H, Yuan H, Sun D. Engineering exosomes as refined biological nanoplatforms for drug delivery. Acta Pharmacol Sin. 2017;38(6):754–63.
Liu Y, Li D, Liu Z, Zhou Y, Chu D, Li X, et al. Targeted exosome-mediated delivery of opioid receptor Mu siRNA for the treatment of morphine relapse. Sci Rep. 2015;5(1):1–10.
Munoz JL, Bliss SA, Greco SJ, Ramkissoon SH, Ligon KL, Rameshwar P. Delivery of functional anti-miR-9 by mesenchymal stem cell–derived exosomes to glioblastoma multiforme cells conferred chemosensitivity. Mol Ther-Nucleic Acids. 2013;2:e126.
Fonsato V, Collino F, Herrera MB, Cavallari C, Deregibus MC, Cisterna B, et al. Human liver stem cell-derived microvesicles inhibit hepatoma growth in SCID mice by delivering antitumor microRNAs. Stem Cells. 2012;30(9):1985–98.
Zhang Y, Li L, Yu J, Zhu D, Zhang Y, Li X, et al. Microvesicle-mediated delivery of transforming growth factor β1 siRNA for the suppression of tumor growth in mice. Biomaterials. 2014;35(14):4390–400.
Banizs AB, Huang T, Dryden K, Berr SS, Stone JR, Nakamoto RK, et al. In vitro evaluation of endothelial exosomes as carriers for small interfering ribonucleic acid delivery. Int J Nanomed. 2014;9:4223.
Zhou W, Zhou Y, Chen X, Ning T, Chen H, Guo Q, et al. Pancreatic cancer-targeting exosomes for enhancing immunotherapy and reprogramming tumor microenvironment. Biomaterials. 2021;268:120546.
Rezaei R, Baghaei K, Amani D, Piccin A, Hashemi SM, Asadzadeh Aghdaei H, et al. Exosome-mediated delivery of functionally active miRNA-375–3p mimic regulate epithelial mesenchymal transition (EMT) of colon cancer cells. Life Sci. 2021;269:119035.
Wadhwa A, Aljabbari A, Lokras A, Foged C, Thakur A. Opportunities and challenges in the delivery of mRNA-based vaccines. Pharmaceutics. 2020;12(2):102.
Wu Z, Li T. Nanoparticle-mediated cytoplasmic delivery of messenger RNA vaccines: challenges and future perspectives. Pharm Res. 2021;38(3):473–8.
Patel P, Vyas N, Raval M. Safety and toxicity issues of polymeric nanoparticles. Nanotechnol Med. 2021;9:156–73.
van der Koog L, Gandek TB, Nagelkerke A. Liposomes and extracellular vesicles as drug delivery systems: a comparison of composition, pharmacokinetics, and functionalization. Advanced Healthcare Mater. 2021;11:2100639.
Hatakeyama H, Akita H, Harashima H. The polyethyleneglycol dilemma: advantage and disadvantage of PEGylation of liposomes for systemic genes and nucleic acids delivery to tumors. Biol Pharm Bull. 2013;36(6):892–9.
Hou X, Zaks T, Langer R, Dong Y. Lipid nanoparticles for mRNA delivery. Nat Rev Mater. 2021;6(12):1078–94.
Hou X, Zaks T, Langer R, Dong Y. Lipid nanoparticles for mRNA delivery. Nat Rev Mater. 2021;89:1–17.
Palazzolo S, Bayda S, Hadla M, Caligiuri I, Corona G, Toffoli G, et al. The clinical translation of organic nanomaterials for cancer therapy: a focus on polymeric nanoparticles, micelles. Liposomes and Exosomes Curr Med Chem. 2018;25(34):4224–68.
Song H, Liu B, Dong B, Xu J, Zhou H, Na S, et al. Exosome-based delivery of natural products in cancer therapy. Front Cell Develop Biol. 2021;9:366.
Meng W, He C, Hao Y, Wang L, Li L, Zhu G. Prospects and challenges of extracellular vesicle-based drug delivery system: considering cell source. Drug Delivery. 2020;27(1):585–98.
Colao IL, Corteling R, Bracewell D, Wall I. Manufacturing exosomes: a promising therapeutic platform. Trends Mol Med. 2018;24(3):242–56.
Lee JH, Ha DH, Go H-K, Youn J, Kim H-K, Jin RC, et al. Reproducible large-scale isolation of exosomes from adipose tissue-derived mesenchymal stem/stromal cells and their application in acute kidney injury. Int J Mol Sci. 2020;21(13):4774.
Herrmann IK, Wood MJA, Fuhrmann G. Extracellular vesicles as a next-generation drug delivery platform. Nat Nanotechnol. 2021;16(7):748–59.
Kim J, Song Y, Park CH, Choi C. Platform technologies and human cell lines for the production of therapeutic exosomes. Extracellular Vesicles Circulating Nucleic Acids. 2021;2(1):3–17.
Pardi N, Hogan MJ, Porter FW, Weissman D. mRNA vaccines — a new era in vaccinology. Nat Rev Drug Discovery. 2018;17(4):261–79.
Ludwig N, Whiteside TL, Reichert TE. Challenges in exosome isolation and analysis in health and disease. Int J Mol Sci. 2019;20(19):4684.
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
Conception and manuscript design: RR, and SHK. Collection of data: AA, RB, EAD and SHK. Manuscript writing: AA, RB, and SHK. Figure design and regenerate: EAD and SHK. Made important revisions and confirmed final revision: RR, and RJ. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Amiri, A., Bagherifar, R., Ansari Dezfouli, E. et al. Exosomes as bio-inspired nanocarriers for RNA delivery: preparation and applications. J Transl Med 20, 125 (2022). https://doi.org/10.1186/s12967-022-03325-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12967-022-03325-7