
Abstract
The human microbiome harbors a diverse array of microbes which establishes a mutually beneficial relation with the host in healthy conditions, however, the dynamic homeostasis is influenced by both host and environmental factors. Smoking contributes to modifications of the oral, lung and gut microbiome, leading to various diseases, such as periodontitis, asthma, chronic obstructive pulmonary disease, Crohn’s disease, ulcerative colitis and cancers. However, the exact causal relationship between smoking and microbiome alteration remains to be further explored.
Similar content being viewed by others


Background
Approximately 2 billion people worldwide use tobacco products, mostly in the form of cigarettes, with tobacco smoking-related diseases resulting in at least 4 million global deaths per year [1]. Dramatic rise of diseases associated with cigarette smoke or tobacco use, including cardiovascular disease, chronic obstructive pulmonary diseases (COPD), Crohn’s disease, and various forms of cancer [2], implying the potential detrimental role of smoking in occurrence of human diseases. Emerging evidence suggests that environmental factors play an influential role in shaping human-associated microbial communities and immune responses. Either active smoking or exposure to secondhand smoke is associated with colonization by potentially pathogenic bacteria [3,4,5]. Yet, in an era where microbes not only cause acute infectious illnesses but also are increasingly being recognized as etiologic agents or risk factors for chronic diseases including cancers [6,7,8] and neurologic disorders [9, 10], it is important to have a profound understanding of the effect of smoking on microbiome in diseases.
The microbiome refers to a community of microbes residing in a defined environment, comprising of bacteria, viruses, fungi, and protozoa, together with their genes and genomes in a given locus. The gastrointestinal microbiome is the most complex echo-system of 10–100 trillion microorganisms, in which the amount of bacteria was the most, in the next place was that of fungi and virus [11]. The oral communities come as the second in the human body [12]. With the initiation of the Human Microbiome Project in 2007, the use of culture-independent methods allied with next generation DNA sequencing methods to identify the composition of the human microbiome, is providing a far deeper analysis than hitherto possible [13], including 16S ribosomalRNA (rRNA) sequencing, metagenomic sequencing, and microbial metatranscriptomics [14]. The once called sterile lung based on conventional culture methods was unraveled to contain variable microbiomes depending on health and specific disease states [15, 16]. The human microbiome has the stability and resilience to restore themselves after perturbation maintains homeostasis in health, but its composition is susceptible to many factors such as antibiotics, diet, alcohol, and smoking [17] (Table 1). It has become clear that the microbiome is not a passive victim in many pathological processes, but its modification often play a contributive or causative role in pathophysiological processes [18]. Thus far, most studies have described the microbial composition of healthy or diseased organs, and smoking associated alterations of microbiome in different sites were demonstrated in a variety of diseases (Table 2). In this review, we will summarize the current understanding of the impact of smoking on microbiome and its involvement in various diseases, and thereby highlighting important research questions that require further investigation.
Possible mechanism of the impact of smoking on microbiome
Massive studies demonstrated the adverse health impacts of tobacco on systemic pathophysiologic changes that can lead to disease, were associated with the chemicals, heavy metals, particulate matter and other constituents in tobacco [19,20,21,22,23,24,25,26,27]. However, a paucity of studies investigated the microbes in tobacco recent years, and this may be incriminated as causative factors in smoking-associated diseases. Before advances in DNA sequencing technology, the golden standard of identification of microbes-culture method, was used to identify the Pantoea agglomerans, Acinetobacter calcoaceticus, and specific Pseudomonadaceae species such as P. fluorescens and Stenotrophomonas maltophilia in fresh tobacco leaves or other species in single tobacco flakes or fine tobacco particles [24, 26]. With the advent of high-throughput sequencing technology, a 16S rRNA-based taxonomic microarray and cloning and sequencing were utilized to identify a variety of uncultured species. Cigarettes made in the European Union contained 15 different classes of bacteria. Sapkota et al. revealed extensive bacterial diversity in cigarettes, ranging from soil microorganisms and commensals to potential human pathogens, including Acinetobacter, Bacillus, Burkholderia, Clostridium, Klebsiella, and Pseudomonas aeruginosa. Many of the detected organisms are capable of causing pneumonia, bacteremias, foodborne illnesses, meningitis, endocarditis, and urinary tract infections [28]. Therefore, mechanism that may lead to different bacteria profiles among smokers may be due to exposure to bacteria in cigarettes, leading to bacterial acquisition and colonization.
Another possibility for the mechanism through which current smokers may have different bacteria community may be related to impaired antimicrobial defenses due to the immunosuppressive nature of tobacco. Tobacco smoking has been observed to affect the peripheral immune system on several levels, including a decrease in the activity of natural killer cells, increase in white blood cell counts, and a higher susceptibility to infection [29]. Smoking increases the number of macrophages, neutrophils, eosinophils, and mast cells, decreases the number of airway dendritic cells, and alters macrophage and neutrophil function [30, 31]. Expanding macrophages and neutrophils demonstrated impaired phagocytic functions to the efficient clearance of bacteria or pathogen, as evidenced by reduced bacterial-stimulated production of superoxide and surface receptor expression, (e.g. TLR2) which is important for the recognition and response to gram-positive bacteria [32, 33]. Therefore, smoking related immunosuppression could permit novel bacteria colonization.
It is also possible that metabolic advantages of biofilm formation and increased adherence to the epithelium are conferred to certain taxa expansion in a smoky environment. Exposure to cigarette smoke could increase biofilm formation by specific bacteria [34, 35]. Biofilm is a self-generated polymer matrix that insulates the pneumococcus and other microbial pathogens from host defense and antibiotics, promoting bacterial persistence [36]. Mutepe et al. found that increased biofilm formation of Streptococcus pneumoniae and inactivation of pneumolysin induced by exposure to cigarette smoke condensate are likely to favor microbial colonization and persistence, both being essential precursors of pneumococcal disease [35]. Similarly, in another study, observations of increased biofilm formation of Staphylococcus aureus and human cell adherence in the presence of cigarette smoke (CS) indicate the role of bioactive effects of CS on resident microbiota in the pathogenesis of respiratory infection in CS-exposed humans [34]. These findings suggest that cigarette smoke may promote colonization and persistence of specific bacterial taxa in the human body through the biofilm formation, contributing to infections in different parts of the body.
“Microenvironment” may also be relevant regarding the influence of smoking on particular members of the microbiota, such as oxygen, pH, and acid production. Oxygen tension is an important promoter of the changes in bacterial community, with microaerophilic and anaerobic bacteria able to predominate due to lower oxygenation [37, 38]. Shanahan et al. demonstrated a reduction of the relative abundance Prevotella and Neisseria spp. and an increased relative abundance of Firmicutes, principally Streptococcus spp., and Veillonella spp., along with the genus Rothia (Actinobacteria) in the upper GI tract from current smokers, compared with that from persons who have never smoked [39]. The differences observed in Neisseria, Streptococcus, and Rothia spp. in current smokers indicated the implication of changes in oxygen tension. In that study, alterations in duodenal bicarbonate secretion [40] and lower pH [41] in smokers may also provide selective pressure on the growth of Neisseria, which is one of the capnophiles and sensitive to acid conditions [42], whereas Streptococcus and Rothia spp. are acidogenic and acid tolerant.
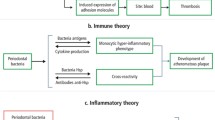
According to above discussion, we could reach the conclusion of mechanisms of cigarette smoking to influence the microbiome via changes to immune homeostasis, biofilm formation, oxygen tension, or through direct contact with microbes it contained, and these mechanisms may be involved in the occurence of various diseases (Fig. 1).

Schematic summary illustrating the possible mechanisms of smoking-induced dysbiosis of microbiome and its possible role in different diseases
Smoking and oral microbiome in diseases
The oral microbiome, comprising more than 2000 bacterial species [43], plays an important role in the maintenance of oral health [44]. Dysbiosis of oral microbiota has been associated locally with periodontal, respiratory, cardiovascular and systemic cancers, including head and neck cancer [45], pancreatic cancer [46], and esophageal cancer [47], yet regarding factors that influence the oral microbiome composition are poorly understood. Smoking is a major environmental factor that influences orodental pathophysiology [48]. Toxic components and bacteria in cigarette impact oral bacteria directly or indirectly through immunosuppression, oxygen deprivation, biofilm formation, or other potential mechanisms [49], leading to loss of beneficial oral species and pathogen colonization, ultimately to disease [50]. Despite of different sampling sites or laboratory methodologies, numerous studies have shown predominant or inhibited genera in oral from smokers compared with non-smokers. Culture results of smokers showed less numerous Neisseria species or Branhamella [51, 52]. Due to limitations on bacterial profiling of traditional method, recently, sequence analysis of bacterial 16S rRNA-encoding genes was performed to identify the different mouth communities between nonsmokers and smokers in species such as Porphyromonas, Neisseria, and Gemella [53]. Mason et al. revealed the microbial profiles of subgingival plaque samples from 200 systemically and periodontally healthy smokers and never-smokers were different at all taxonomic levels, and principal coordinate analysis revealed distinct clustering of the microbial communities based on smoking status. Smokers demonstrated a highly diverse, pathogen-rich, commensal-poor, anaerobic microbiome that is more closely aligned with a disease-associated community in clinically healthy individuals, suggesting that it creates an at-risk-for-harm environment that is primed for a future ecological catastrophe [37].
Periodontitis
Evidences indicate that periodontitis was associated with smoking and complex microbial communities in the subgingival sulcus [54,55,56], and cigarette smokers were found to have a statistically significant higher risk of severe periodontitis than non-tobacco users [57]. More and more researchers focused on the associations between smoking and sub-gingival bacterial species in the pathogenesis of periodontitis. Smoking-associated periodontitis is less diverse and distinct from that of non-smokers. Shchipkova et al. explored that the microbial profile of smokers with moderate to severe chronic periodontitis and demonstrated significant differences in the prevalence and abundance of disease-associated and health-compatible organisms, with greater abundance of Parvimonas, Fusobacterium, Campylobacter, Bacteroides, and Treponema and lower levels of Veillonella, Neisseria, and Streptococcus [58]. The differences existed in the composition of the subgingival microbiome between smoker and non-smoker patients with chronic moderate periodontitis were also elucidated in other studies [59,60,61]. In addition, smokers are at high risk for other oral diseases, such as peri-implant mucositis and peri-implantitis [62, 63]. Tsigarida et al. demonstrated smoking shapes the peri-implant microbiome of peri-implant biofilm samples from patients with peri-implant health, peri-implant mucositis, and peri-implantitis [63], paralleled with studies depicting that the underlying mechanism is through depleting commensals from this niche and favoring colonization of pathogens [63].
Infective endocarditis
In recent years, significant associations have been elucidated between periodontitis and other systemic diseases [64, 65], and the bacterial flora of the mouth entering the bloodstream may potentially be involved in the pathogenesis of invasive infections such as infective endocarditis, and the bacterial flora of the mouth entering the bloodstream may potentially be involved in the pathogenesis of invasive infections such as infective endocarditis [66]. Oral bacteria of endocarditis patients have been reported to be shown different community compositions from that of healthy individuals [67, 68]. Staphylococcus aureus, viri-dans Streptococci and Enterococcus spp. are the most common pathogens identified [69]. Gemella sanguine, Streptococcus tigurinus, L. goodfellowi were also found to be the cause of infective endocarditis [70,71,72]. However, there was barely study about the effect of smoking on infective endocarditis regarding oral microbiome. Biofilm formation, complex mechanisms with other bacteria might play a crucial role in the occurrence of invasive infections [71]. Indeed, endocarditis is also considered an example of a biofilm-mediated disease [73]. Given the importance of biofilm formation for adhesion in the oral cavity [70], and the fact that cigarette smoke increased biofilm formation by specific bacteria and promoted colonization [34], it would be reasonable to suspect that oral microbiome might be the missed connectivity between smoking and infective endocarditis. Further studies on whether smoking could increase the incidence of infectious endocarditis through altering oral micro-organisms remains to be elucidated.
Other
In addition to local health, oral microbiome also plays an important role in other systemic diseases, including HIV infection, gastrointestinal cancer, even immune diseases. HIV infection has been associated with dysbiosis of oral microbiome, with increased levels of pathogenic bacteria and fungi [74, 75], HIV-infected smokers showed rich abundance of specific bacterial taxa compared to infected non-smokers, including Granulicatella, Lactobacillus, Veillonella, Enhydrobacter, Streptococcaceae and Comamonadaceae, moreover, abundance of the fungal genus Candida was also increased in HIV-infected smokers [76]. Oral microbiome was also corroborated to be associated etiologically with gastrointestinal cancer in virtue of composition concordance among sites within the oral cavity and gut, and anatomical acquirement of gut microbiome from mouth. In colorectal cancer participants, current smoking was associated with a 33% decrease in relative counts of Betaproteobacteria (primarily Neisseria) and 23% increase in relative abundance of Veillonellaceae family [77]. These data indicated that community composition of oral microbiome may be associated with numerous diseases such as periodontitis, cancer, and diabetes, however, it remains to elucidate the causative relationship between a specific bacterium and the disease, and future work may also wish to consider this potential association.
Smoking and airway microbiome
Because of the presence of a sparse microbiome, especially in healthy conditions, traditional standard microbiological culture-based methods can hardly detect microbes in healthy individuals, so the lung has historically been presumed as sterile. Over the past several years, culture-independent molecular methods springing up, lung microbial communities in healthy individuals showed a phenotype predominant by Proteobacteria, Firmicutes and Bacteroidetes, as evidenced by bronchoalveolar lavage samples of healthy adults, bacterial communities vary with different airways in terms of different airway microarchitecture, and documented changes in the lung microbiome in several lung diseases have been uncovered [78, 79].
Smoking, cigarette smoke exposure, tobacco smoke or pollutants in the air contact directly with the airway, through the way to the lungs, causing a variety of airway diseases, such as COPD, asthma, cystic fibrosis and lung cancer. In recent years, the effect of smoking on microbiome of lower respiratory tract attracted increasing attention. Mammen and Aethi proposed a revised “Vicious Circle”, suggesting that insults such as tobacco smoke impairs innate immune defenses, causing variations in the abundance, taxonomic composition and phylogenetic diversity of the lung microbiome. This, in turn, leads to maladaptive inflammatory responses, further impairment of lung defenses and further dysbiosis of the lung microbiome, setting up the vicious circle with its attendant consequences [80].
COPD
COPD is a chronic airway inflammatory disease that can be prevented and treated lung disease, characterized by a largely irreversible chronic obstruction of airflow. The course of the disease is featured and frequently aggravated by intermittent exacerbations, acute changes in the airway microbiome, for example by introduction of a new strain of a respiratory pathogen, lead to larger inflammatory responses, which present clinically as exacerbations of COPD [80], so changes in the composition and activity of the microbiome may be implicated in their appearances. Pragman et al., once described the lung microbiome in moderate and severe COPD patients, with the former dominated by Actinobacteria and Proteobacteria, the later by Actinobacteria and Firmicutes, and they also found a trend without significant difference that severe subjects contained more Firmicutes and less Actinobacteria and Proteobacteria than the moderate subjects [81]. However, in another cohort study, in both stable and exacerbated samples, the most prevalent phyla were Proteobacteria, Firmicutes and Actinobacteria, the most prevalent genera were Streptococcus and Haemophilus, exceeding half of the abundance of present bacterial microbiome [82]. Although these researchers also found no significant differences in bronchial microbiome between stability and exacerbation using 16S rRNA sequencing and shotgun metagenomic sequencing, functional metabolic capabilities showed significant changes in several pathways, indicating specific changes in the lung microbiome in the progression of COPD [82].
Smoking is the principal cause or initiating factor responsible for the development of the disease in COPD patients. It alters host–microorganism interaction dynamics in the airways, contributing to COPD [83]. The influence of cigarette smoke on the microbiome and the role of the microbiome in COPD are relatively new field with limited data. Erb-Downward et al. showed the diversity of bacterial communities in bronchoalveolar lavage fluid (BALF) from healthy smokers was similar to that from healthy never-smokers, and COPD patients, however, further results from obtained lung tissues of COPD patients unraveled the heterogeneity and diversity in the bacterial microbiota across different regions of the abnormal lung [79], suggesting specific changes in lung microbiome resulted from smoking in COPD patients participate in the occurrence of COPD or exacerbation. Respiratory tract infections, either viral or bacterial, are major causes of acute exacerbation of COPD (AECOPD) [84]. Cigarette smoke exposure is a well-known risk factor for important bacterial and viral infections in the respiratory tract [85]. In the lung ecosystem, virus also plays a pivotal roles in lung diseases, for example, phages could lead to immune-mediated microbial competition [86], opportunistic infection [87], therefore, alterations of lung viral communities could change the bacteriome leading to dysbiosis and disease progression in individuals (e.g., COPD). Gregory et al. performed the first study of the effects of smoking on the lung DNA virome, lung viromes profiles were statistically indistinguishable across smokers and nonsmokers, and viral diversity was significantly lower in the lungs of healthy smokers [88]. Statistical analyses revealed that changes in viral communities correlate most with changes in levels of arachidonic acid and IL-8, both potentially relevant for COPD pathogenesis [88]. These data imply the potential role of changes in viral communities induced by smoking in the development of COPD. Although the role of smoking on microbiome in COPD needs to be further investigated, early studies have suggested an association between lung microbiota and the clinical outcomes of disease.
Asthma
Asthma is a chronic airway inflammatory disease, characterized by reversible airway obstruction, chronic airway inflammation, and airway hyper-responsiveness [89,90,91]. There are an emerging number of studies shown the correlations between airway microbiome and the incidence, severity or reactivity of corticosteroid medications of asthma [92,93,94,95]. Asthmatic patients harbored higher abundance of Proteobacteria and lower Bacteroidetes phylum compared to healthy control as evidenced from samples of bronchial epithelial brushings [96], and the sputum microbiota in severe asthma patients differs from healthy controls and non-severe asthmatics, with a significant correlation between Streptococcus spp. and eosinophilia in severe asthma patients [94]. Airway colonization with distinct specific bacteria were also associated with the severity of airways obstruction, neutrophilic airway inflammation [97] and corticosteroid resistance in asthma [98]. While the bacterial communities in airway inflammatory disease have been extensively studied, fungal microbiota is still poorly characterized. The analysis of induced sputum revealed that 90 fungal species were more abundant in asthmatics [99], of which members of genera Aspergillus and Penicillium were significantly associated with impaired post-bronchodilator expiratory volume in 1 s in asthmatics [100]. Severe asthmatics were characterized by enrichment of Aspergillus; the relative abundance of Aspergillus increased approximately 15-fold compared to mild asthmatics [101]. However, whether these organisms are cause or result of the pathophysiology or medications in asthma remains to be determined.
As the main source of indoor air pollution, tobacco smoking, mostly in the form of cigarette smoking, is an important environmental factor influencing the outcomes of asthma. Smoking (including active and passive smoking) can not only cause frequent attacks of asthma, lead to rapid decline of lung function in asthmatic patients, but also reduce the therapeutic effect of glucocorticoid in asthmatic patients, making the condition of asthmatic patients difficult to control [102]. Microbial colonization of the lower airway may be shaped by smoking in asthma, accordingly, some of the increased risk and severity of pulmonary disorders in tobacco smokers with asthma [103] could still be mediated through smoking-induced changes in the lung microbiome. Simpson et al. extracted DNA from induced sputum in asthma patients and profiled microbial communities using 16S rRNA pyrosequencing, the results showed ex-smokers have a higher prevalence of phylum Fusobacteria, the phyla Firmicutes and Bacteroidetes, and a lower abundance of bacteria from phylum Proteobacteria compared with never-smokers. They also revealed an association between smoking and increased diversity of bacteria [104]. However, another study failed to demonstrate the association, because of no difference in bacterial dominance from induced sputum in asthma patients between ex-smokers and non-smokers, this lack of difference may be explained by the small sample size and different samples in that study [97]. More studies examining effect of smoking on airway microbiome in individuals with asthma are warranted.
Cystic fibrosis
Cystic fibrosis (CF) is a multi-organ disease with variable clinical characteristics, with pulmonary manifestations (e.g. bronchiectasis and chronic infection) being the prominent feature [105, 106]. Progressive lung disease driven by microbial colonization and inflammation remains the leading cause of morbidity and mortality in CF patients [106]. The lung environment in cystic fibrosis patients, characterized by depletion of the airway surface liquid layer leading to impediment of mucociliary clearance, is ideal for microbial colonization [107]. Culture-independent molecular methods allowed emerging threads of the features of the respiratory microbiome in CF to be gradually discovered in recent years. The lung microbiota fluctuated in a chronological way and associated with clinical states. In CF patients aged < 2 years, nontraditional taxa (e.g. Streptococcus, Prevotella and Veillonella) predominated, these species shifted to traditional CF taxa (Pseudomonas, Staphylococcus, Haemophilus, Stenotrophomonas and Burkholderia) [108] and remained relatively stable in older children (> 6 years) and adults, especially in clinical stability [109,110,111]. With improvement of biological diagnosis, fungal and viral colonization in CF moved into the focus. Candida albicans and Aspergillus fumigatus are commonly detected in CF sputum cultures and have also been associated with acute pulmonary exacerbations [112], and the number of fungal species detected in sputum fluctuated over time [113]. As for respiratory “virome” in CF, precious few data reported distinct phage communities in CF compared with non-CF patients [114].
Mutations of cystic fibrosis transmembrane conductance regulator (CFTR), an epithelial anion channel, are predominant in the cause of CF [115]. Smoking is one of the most important adverse factors affecting respiratory health. Recent researches pay close attention to the issue related to the impact of smoking in the pathology of CF and clinical outcomes. Campbell et al. [116] found tobacco smoke in patients with CF exhibited poor clinical status, including reduced lung function, and a higher number of pulmonary-related hospitalizations. Some other studies focus on the variations of genes within the context of tobacco smoking and detected the associations between smoke exposure and CTFR dysfunction [117, 118], which could in turn lead to deleterious effects on airway surface liquid secretion, enhanced mucus expression, reduced mucociliary clearance, chronic bacterial infection, and excess inflammation [119], rendering the main morbidity and mortality in CF patients because of the fact that lung inflammation and chronic respiratory infections alone account for nearly 95% of the morbidity and mortality in patients with CF [120]. In addition, smoking is known to stimulate mucosal linings and increase sputum production in the respiratory tract, raising the possibility of bacterial infections. Yet, there is no evidence to investigate the microbiome (e.g. bacteria) in CF patients in the context of smoking. Both the effect of smoking on microbiome in CF patients and the possible causal relationship between smoking-induced variations of microbiome, possibly through CTFR dysfunction, and the clinical manifestations of CF, could not be proved. We may assume the possible mechanism of detrimental effect of smoking on CF may be because the alterations in microbiome due to airway surface liquid secretion, enhanced mucus expression, reduced mucociliary clearance induced by loss of CTFR functions.
Smoking and gut microbiome
The gastrointestinal microbiome is a complex echosystem of 10–100 trillion microorganisms composed of bacteria, virus and fungal species, that develops immediately after birth depending on multiple factors, and fluctuates or changes resulted from affection of a number of factors such as age, drugs (especially antibiotics), diet, alcohol, and smoking throughout the whole time growing up [11, 17]. In virtue of the most extensively focus on microbiota colonizing the intestinal tract, it has become clear that in healthy individuals, the microbiome is inclined to remain rather stable, with Bacteroides, Faecalibacterium, and Bifidobacterium being the most prevalent genera [121], and disturbance of the microbial equilibrium is associated with a variety of local and systemic diseases [17, 18].
Smoking prevalence is the leading cause responsible for developing Crohn’s disease (CD), colonic carcinoma, and systemic disease [122, 123]. The vulnerability of intestinal microbiome provokes interest in microbiome alterations in smoking environment. In animal models, cigarette smoke decreases organic acids levels and population of bifidobacterium in the caecum of rats [124]. Side-stream smoking increased the abundance of Clostridium and decreased the amount of Lactoccoci, Ruminococcus, Enterobacteriaceae and segmented filamentous bacteria (SFB) in the cecal microflora [125]. In human studies, smoking increased the probability of developing Clostridium difficile infection [126]. Current smokers displayed increased Bacteroidetes and decreased Firmicutes and Proteobacteria in gut microbiota composition community compared with never smokers [127]. Another study also revealed healthy smokers harbour higher Bacteroides–Prevotella (34.8%) than nonsmokers (24.1%) [128]. In addition, smoking status is also connected with variations in gut microbiome, as reported by Biedermann et al. that healthy individuals undergoing smoking cessation increased Firmicutes and Actinobacteria, while decreasing Bacteroidetes and Proteobacteria [129]. Many researchers investigated the role of smoking in gut microbiome in inflammatory bowel disease, we will discuss the effects of smoking on gut microbiome in several diseases.
Crohn’s disease
Crohn’s disease (CD) is a type of inflammatory bowel disease that mainly affect gastrointestinal tract, characterized by chronic inflammation due to defective mucosal barrier and greater intestinal permeability [130]. While the cause of CD is multifactorial, a combination of environmental, immune, and bacterial factors in genetically susceptible individuals [131,132,133]. Microbial dysbiosis is thought to be associated with either development or exacerbation of underlying Crohn’s disease [134]. CD patients have a relevant dysbiosis of the gastrointestinal microbiome, including reduction of normal commensal phyla (Bacteroidetes and Firmicutes), increase of pathogenic organisms (E. coli, Campylobacter species, and Mycobacterium species) [135], and a greater number of mucosal surface-associated bacteria with higher adherence and invasion compared with healthy control subjects [136]. Researchers also found CD patients possess a reduction in Roseburia spp., Clostridium and Bacteroides species, known to be producers of butyrate, which is fundamental to intestinal cell homeostasis and mucosal barrier integrity [137, 138]. These studies have shed light on the possible causative role of the dysbiosis of gut microbiota in CD.
Smoking is the best studied environmental risk factor for CD, exerting detrimental effects on mucosal barrier and greater intestinal permeability and disease susceptibility [139]. Accumulating data investigated the association between smoking and imbalance of intestinal microbiome [139]. Accordingly, several studies indicated that intestinal microbes could be an important link between smoking and CD [139]. Benjamin et al. found greater abundance of Bacteroides–Prevotella in smoking patients with CD compared with nonsmokers through fluorescent in situ hybridization using probes targeting 16S rRNA of bacteria [128]. In another study, Opstelten reported a reduced microbial gene richness and taxonomic diversity in smoking patients with CD, they further demonstrated a statistically significant reduction in specific genera Collinsella, Enterorhabdus, and Gordonibacter [140], which can produce urolithins with anti-inflammatory properties [141]. Faecalibacterium has immune-regulatory function to reduce IL-12 expression in peripheral blood mononuclear cell (PBMC) in vitro and increase IL-10 release, lower ileal mucosal Faecalibacterium prausnitzii is correlated with greater risk of recurrence following surgical resection in CD patients [142]. Murugananthan et al. revealed a reduction in the number of Faecalibacterium prausnitzii in inflamed mucosal tissue from smokers with active CD compared with non-smokers, the risk of post-operative CD recurrence may be predetermined at a pre-operative stage due to dysbiosis. These observed features of reduced gut microbiota may explain the persistent intestinal inflammation in CD patients in smoky environment, highlighting the possible role of microbes interacting with smoking and CD (or mediating the adverse effects of smoking in CD). However, the mechanisms through which smoking caused alterations in microbiota are unclear.
Ulcerative colitis
Ulcerative colitis (UC) is another type of chronic relapsing inflammatory disorder confined to the colorectal region and to the mucosal layer of the gastrointestinal tract. Previous studies have demonstrated aberrant microbiota deviations from gut homeostasis in UC patients, as evidenced by a low taxonomic diversity, decreases in Firmicutes and increases in Proteobacteria in UC gut microbiomes [143,144,145]. Proportions of Fusobacteriaceae family increased, Bifidobacteria and members of the Faecalibacterium taxon appeared to be compromised in gut microbiota of UC patients [146, 147], and further study suggested the reduced abundance of Bifidobacteria as a microbial biomarker to identify the intestinal dysbiosis triggering UC [147].
The understanding of pathogenesis and etiology of UC is still out of reach. It was presumed that genetically susceptible individuals of UC or patients with UC exhibited aberrant mucosal immune response against their gut microbiota [148, 149], which could result in productions of pro-inflammatory cytokines responsible for abnormal inflammation reaction in the digestive tract [150]. Environmental factors were deemed to trigger the onset and cause flares of inflammatory bowel disease. Smoking is among the most widely studied factors described in UC. The contradictory results of reverse associations between smoking and the natural history of UC has long been the topic of great interest. Some studies showed current smokers with UC are more likely to show milder disease, fewer hospitalizations, and reduced need for corticosteroid and immunosuppressant therapy compared with non-smokers [151]. Others did not find the protective role [152, 153]. Li et al. [154] suggested cigarette smoking could change the composition of intestinal microbiota, modulate mucus production and inhibit repairing of the gastrointestinal tract. Nevertheless, the main components of cigarette smoking has inflammatory-regulatory properties, for instance, Heme oxygenase-1 (HO-1) and carbon monoxide (CO) play a role in modulating cytokine expression and macrophage bactericidal activity, regulating intestinal homeostasis and mucosal immune responses to the enteric microbiota [155, 156], both of which were involved in the possible pathological process in UC. The exact mechanisms through which smoking is associated with alterations in microbiota are unclear. As yet, there is no data exploring the effect of smoking on gut microbiota in patients with UC. Smokers may have behavioral characteristics, such as diet, that predispose to a luminal and mucosal dysbiosis [157]. However, it is also possible that smoking has a direct influence on the microbiota, and this deserved to be mentioned in future research hotspots.
Colorectal cancer
Colorectal cancer (CRC) is considered a major public healthy issue, with approximately 700,000 deaths reported annually worldwide [158], ranking only second to lung cancer [159]. Chronic inflammation is a well-established factor associated with cancer onset, progression through mucosal disruption and the excess of reactive oxygen species (ROS) [160, 161]. At present, many scholars believe that the pathogenesis of CRC may be associated with the participation of intestinal microorganisms, which started the damage mechanisms of intestinal mucosal immune response, thus leading to immune response and inflammation [162]. The correlation between gut microbiome and the initiation of cancer can be dated from last century when people observed the potential etiology of bacteria Streptococcus bovis in the occurrence of CRC [163]. In recent years, the disturbance of gut microbiome was further pushed to a high new climax, as distinct gut microbiome composition detected in CRC patients.
The abundance of several microbes, such as Streptococcus gallolyticus [164], Fusobacterium, [165], B. fragilis [166], Escherichia–Shigella, Peptostreptococcus ten [167] were observed to be enriched in CRC patients versus control groups, while genera such as Bacteroides, Roseburia [164] and Pseudomonas [167] were significantly depleted in CRC patients. Moreover, the barely studied fields gut virome and mycobiome correlated with CRC were also reported when Nakatsu et al. identified a set of discriminatory virome signatures (e.g. Orthobunyavirus, Tunalikevirus, Phikzlikevirus, Betabaculovirus, Sp6likevirus, Sfi21dtunalikevirus, Punalikevirus, Lambdalikevirus, C2likevirus, and Mulikevirus) enriched in CRC subjects [168], and when Coker et al. reported higher Malasseziomycetes and depleted Saccharomycetes and Pneumocystidomycetes in CRC patients [169].
CRC is a complex disease susceptible to a variety of diet and lifestyle factors, especially smoking, a well-known factor involved in the initiation and increasing the risk of CRC with a prolonged latency period [170]. Although the mechanisms of smoking-induced susceptibility to higher risk of CRC remain to be elucidated, preliminary evidence suggests a collective role of host, microbial, and smoking, such as intestinal and immune disruption, impaired clearance of pathogens, changes in the virulence of bacteria and fungi, and ingestion of bacteria that are present in cigarettes [171]. The fact as mentioned above that cigarette smoke or side-stream smoking decreases the amount of Bifidobacterium [124], mainly butyrate-producing bacteria with anti-inflammatory and anti-tumor molecule role [172] was highly consistent with the results that butyrate-producing bacteria are depleted in cancer patients [170]. In addition, in vitro and in vivo studies found that cigarette smoke not only decrease the fecal abundance of Bifidobacterium but also reduce its production of short chain fatty acids (SCFAs) [124, 173], immune-regulatory molecules modulating immune and inflammatory response within many diseases, and reductions in the concentration of SCFAs especially butyrate in colorectal tissues were demonstrated to be associated with the possibility of early stage CRC development [174]. Moreover, the smoking-related microbial changes may lead to altered epithelial mucin composition of the mucus layer and increased inflammatory response [175], which play pivotal role in the onset of CRC.
Stated thus, these studies suggested that alterations of gut microbiome may be an essential contributing factor to the initiation and development of this cancer in the context of smoking.
Systemic diseases
In addition to inflammatory bowel disease, dysbiosis of the gut microbiome has been implicated in many autoimmune diseases, including multiple sclerosis (MS) [176], rheumatoid arthritis (RA) [177], ankylosing spondylitis [178], systemic lupus erythematous (SLE) [179], and [180]. Human studies and mouse models support the role of the gut microbiome in predisposition to RA, such as reduction in bacteria belonging to the family Bifidobacterium and Bacteroides [181], and higher prevalence Prevotella copri [182]. Ye et al. also revealed higher abundance of Bilophila spp., and several opportunistic pathogens (e.g., Parabacteroides spp. and Paraprevotella spp.), together with a reduction in butyrate-producing bacteria Clostridium spp., and two genera of methanogens (Methanoculleus spp., Methanomethylophilus spp.) in BD patients [183]. MS has also been recently associated with changes of the intestinal microbiota with the study reporting depletion in species belonging to Clostridium and Bacteroidetes in Japanese patients with MS [184].
Smoking affects both innate and adaptive immune systems and plays dual roles in regulating immunity by either exacerbation of pathogenic immune responses or weakening of defensive immunity [185]. Smoking is a well-established risk factor for developing RA, SLE and MS [186,187,188,189]. Epidemiologic data showed positive association between the incidence of MS and smoking, the risk of MS increased with the number of pack-years of smoking increasing [190], smoking renders individuals with HLA–DRB1 shared epitope (SE) alleles susceptible to RA, stronger association existed in individuals carrying double copies of the SE [186]. The gut microbiome is responsible for maintaining homeostasis and function of host immune system indicating the probable essential role in changing the immune response that leads to autoimmune diseases like RA. As mentioned earlier, cigarette smoking could change the composition of intestinal microbiota [154], and some of the components play an important role in modulating intestinal homeostasis and immune responses to the enteric microbiota [155, 156]. The role of smoking involved in these diseases was assessed in both animal models and clinical trials, but so far no exact underlying mechanisms were identified. The gut microbiome may provide the missing link to this puzzle and help solve the mystery of the influence of smoking in autoimmune diseases, and this deserve the future research to further the understanding of the role of microbiome in systemic diseases.
Conclusions
In conclusion, microbiome research has enormously developed in the last years tempting to move its steps to better characterize the human microbiome. Smoking is the risk factor of several diseases, it could impact human microbiome directly or indirectly through immunosuppression, oxygen deprivation, biofilm formation, or other potential mechanisms. None of the above mechanisms is well established, and adequate explanation of how smoking affects the microbiome is yet to be established. Microbiome has pivotal roles in the development of healthy immune responses, and oral, airway and gut microbial dysbiosis can contribute to local or systemic various diseases such as periodontitis, HIV infection, gastrointestinal cancer, asthma, COPD, CF, CD, UC, RA, MS. Evidence suggests the microbial dysbiosis in many diseases in smoky environment, but the causal relationship between microbiome alterations and disease progress remains enigmatic. More basic and clinical researches may help us gain more insight into the hugely complex net-work of smoking–microbiome–host interactions underlying the observed associations. Longitudinal studies integrating metagenomic, transcriptomic, metabolomic, methods with clinical results may help us ascertain the relationships between smoking, microbiome, and pathological mechanism in diseases.
Availability of data and materials
Not applicable.
Abbreviations
- COPD:
-
chronic obstructive pulmonary diseases
- CS:
-
cigarette smoke
- BALF:
-
bronchoalveolar lavage fluid
- CF:
-
cystic fibrosis
- CFTR:
-
cystic fibrosis transmembrane conductance regulator
- CD:
-
Crohn’s disease
- UC:
-
ulcerative colitis
- MS:
-
multiple sclerosis
- RA:
-
rheumatoid arthritis
- SLE:
-
systemic lupus erythematous
References
DeMarini DM. Genotoxicity of tobacco smoke and tobacco smoke condensate: a review. Mutat Res. 2004;567:447–74.
Durazzo TC, Mattsson N, Weiner MW. Alzheimer’s disease neuroimaging I: smoking and increased Alzheimer’s disease risk: a review of potential mechanisms. Alzheimers Dement. 2014;10:S122–45.
Brook I, Gober AE. Recovery of potential pathogens and interfering bacteria in the nasopharynx of smokers and nonsmokers. Chest. 2005;127:2072–5.
Brook I, Gober AE. Recovery of potential pathogens in the nasopharynx of healthy and otitis media-prone children and their smoking and nonsmoking parents. Ann Otol Rhinol Laryngol. 2008;117:727–30.
Shiloah J, Patters MR, Waring MB. The prevalence of pathogenic periodontal microflora in healthy young adult smokers. J Periodontol. 2000;71:562–7.
Correa P. Bacterial infections as a cause of cancer. J Natl Cancer Inst. 2003;95:E3.
Hohenberger P, Gretschel S. Gastric cancer. Lancet. 2003;362:305–15.
Parsonnet J. Bacterial infection as a cause of cancer. Environ Health Perspect. 1995;103(Suppl 8):263–8.
McKee DH, Sussman JD. Case report: severe acute Parkinsonism associated with streptococcal infection and antibasal ganglia antibodies. Mov Disord. 2005;20:1661–3.
Schulz JD, Hawkes EL, Shaw CA. Cycad toxins, Helicobacter pylori and parkinsonism: cholesterol glucosides as the common denomenator. Med Hypotheses. 2006;66:1222–6.
Gill SR, Pop M, Deboy RT, Eckburg PB, Turnbaugh PJ, Samuel BS, Gordon JI, Relman DA, Fraser-Liggett CM, Nelson KE. Metagenomic analysis of the human distal gut microbiome. Science. 2006;312:1355–9.
Human Microbiome Project C. Structure, function and diversity of the healthy human microbiome. Nature. 2012;486:207–14.
Proctor LM. The human microbiome project in 2011 and beyond. Cell Host Microbe. 2011;10:287–91.
Dy R, Sethi S. The lung microbiome and exacerbations of COPD. Curr Opin Pulm Med. 2016;22:196–202.
Huang YJ, Charlson ES, Collman RG, Colombini-Hatch S, Martinez FD, Senior RM. The role of the lung microbiome in health and disease. A National Heart, Lung, and Blood Institute workshop report. Am J Respir Crit Care Med. 2013;187:1382–7.
Kiley JP. Advancing respiratory research. Chest. 2011;140:497–501.
Faith JJ, Guruge JL, Charbonneau M, Subramanian S, Seedorf H, Goodman AL, Clemente JC, Knight R, Heath AC, Leibel RL, et al. The long-term stability of the human gut microbiota. Science. 2013;341:1237439.
Owyang C, Wu GD. The gut microbiome in health and disease. Gastroenterology. 2014;146:1433–6.
Humans IWGotEoCRt. Tobacco smoke and involuntary smoking. IARC Monogr Eval Carcinog Risks Hum. 2004;83:1–1438.
Bogden JD, Kemp FW, Buse M, Thind IS, Louria DB, Forgacs J, Llanos G, Moncoya Terrones I. Composition of tobaccos from countries with high and low incidences of lung cancer. I. Selenium, polonium-210, Alternaria, tar, and nicotine. J Natl Cancer Inst. 1981;66:27–31.
Eaton T, Falkinham JO 3rd, von Reyn CF. Recovery of Mycobacterium avium from cigarettes. J Clin Microbiol. 1995;33:2757–8.
Hasday JD, Bascom R, Costa JJ, Fitzgerald T, Dubin W. Bacterial endotoxin is an active component of cigarette smoke. Chest. 1999;115:829–35.
Kurup VP, Resnick A, Kagen SL, Cohen SH, Fink JN. Allergenic fungi and actinomycetes in smoking materials and their health implications. Mycopathologia. 1983;82:61–4.
Larsson L, Szponar B, Ridha B, Pehrson C, Dutkiewicz J, Krysinska-Traczyk E, Sitkowska J. Identification of bacterial and fungal components in tobacco and tobacco smoke. Tob Induc Dis. 2008;4:4.
Morishita Y. Mutagenicity of pyrolysates of salt-tolerant bacteria from food and cigarettes. Cancer Lett. 1983;18:229–34.
Pauly JL, Waight JD, Paszkiewicz GM. Tobacco flakes on cigarette filters grow bacteria: a potential health risk to the smoker? Tob Control. 2008;17(Suppl 1):i49–52.
Rooney AP, Swezey JL, Wicklow DT, McAtee MJ. Bacterial species diversity in cigarettes linked to an investigation of severe pneumonitis in U.S. Military personnel deployed in operation Iraqi freedom. Curr Microbiol. 2005;51:46–52.
Sapkota AR, Berger S, Vogel TM. Human pathogens abundant in the bacterial metagenome of cigarettes. Environ Health Perspect. 2010;118:351–6.
Jaspers I. Cigarette smoke effects on innate immune mechanisms in the nasal mucosa. Potential effects on the microbiome. Ann Am Thorac Soc. 2014;11(Suppl 1):S38–42. https://doi.org/10.1513/AnnalsATS.201306-154MG.
Mehta H, Nazzal K, Sadikot RT. Cigarette smoking and innate immunity. Inflamm Res. 2008;57:497–503.
Arnson Y, Shoenfeld Y, Amital H. Effects of tobacco smoke on immunity, inflammation and autoimmunity. J Autoimmun. 2010;34:J258–65.
Matthews JB, Chen FM, Milward MR, Ling MR, Chapple IL. Neutrophil superoxide production in the presence of cigarette smoke extract, nicotine and cotinine. J Clin Periodontol. 2012;39:626–34.
Droemann D, Goldmann T, Tiedje T, Zabel P, Dalhoff K, Schaaf B. Toll-like receptor 2 expression is decreased on alveolar macrophages in cigarette smokers and COPD patients. Respir Res. 2005;6:68.
Kulkarni R, Antala S, Wang A, Amaral FE, Rampersaud R, Larussa SJ, Planet PJ, Ratner AJ. Cigarette smoke increases Staphylococcus aureus biofilm formation via oxidative stress. Infect Immun. 2012;80:3804–11.
Mutepe ND, Cockeran R, Steel HC, Theron AJ, Mitchell TJ, Feldman C, Anderson R. Effects of cigarette smoke condensate on pneumococcal biofilm formation and pneumolysin. Eur Respir J. 2013;41:392–5.
Oggioni MR, Trappetti C, Kadioglu A, Cassone M, Iannelli F, Ricci S, Andrew PW, Pozzi G. Switch from planktonic to sessile life: a major event in pneumococcal pathogenesis. Mol Microbiol. 2006;61:1196–210.
Mason MR, Preshaw PM, Nagaraja HN, Dabdoub SM, Rahman A, Kumar PS. The subgingival microbiome of clinically healthy current and never smokers. ISME J. 2015;9:268–72.
Ganesan SM, Joshi V, Fellows M, Dabdoub SM, Nagaraja HN, O’Donnell B, Deshpande NR, Kumar PS. A tale of two risks: smoking, diabetes and the subgingival microbiome. ISME J. 2017;11:2075–89.
Shanahan ER, Shah A, Koloski N, Walker MM, Talley NJ, Morrison M, Holtmann GJ. Influence of cigarette smoking on the human duodenal mucosa-associated microbiota. Microbiome. 2018;6:150.
Boral MC. Studies on the erythropoietic effect of plasma from anemic toads both with and without testis. Endokrinologie. 1979;73:243–6.
Murthy SN, Dinoso VP Jr, Clearfield HR, Chey WY. Serial pH changes in the duodenal bulb during smoking. Gastroenterology. 1978;75:1–4.
Ainsworth MA, Hogan DL, Koss MA, Isenberg JI. Cigarette smoking inhibits acid-stimulated duodenal mucosal bicarbonate secretion. Ann Intern Med. 1993;119:882–6.
Warinner C, Rodrigues JF, Vyas R, Trachsel C, Shved N, Grossmann J, Radini A, Hancock Y, Tito RY, Fiddyment S, et al. Pathogens and host immunity in the ancient human oral cavity. Nat Genet. 2014;46:336–44.
Wade WG. The oral microbiome in health and disease. Pharmacol Res. 2013;69:137–43.
Hayes RB, Ahn J, Fan X, Peters BA, Ma Y, Yang L, Agalliu I, Burk RD, Ganly I, Purdue MP, et al. Association of oral microbiome with risk for incident head and neck squamous cell cancer. JAMA Oncol. 2018;4:358–65.
Fan X, Alekseyenko AV, Wu J, Peters BA, Jacobs EJ, Gapstur SM, Purdue MP, Abnet CC, Stolzenberg-Solomon R, Miller G, et al. Human oral microbiome and prospective risk for pancreatic cancer: a population-based nested case–control study. Gut. 2018;67:120–7.
Peters BA, Wu J, Pei Z, Yang L, Purdue MP, Freedman ND, Jacobs EJ, Gapstur SM, Hayes RB, Ahn J. Oral microbiome composition reflects prospective risk for esophageal cancers. Cancer Res. 2017;77:6777–87.
Lonnroth I, Falsen E, Westin J, Lindholm L. Two cases of “heavy chain disease”. Acta Pathol Microbiol Scand B Microbiol Immunol. 1971;79:443.
Macgregor ID. Effects of smoking on oral ecology. A review of the literature. Clin Prev Dent. 1989;11:3–7.
Nociti FH Jr, Casati MZ, Duarte PM. Current perspective of the impact of smoking on the progression and treatment of periodontitis. Periodontol. 2000;2015(67):187–210.
Colman G, Beighton D, Chalk AJ, Wake S. Cigarette smoking and the microbial flora of the mouth. Aust Dent J. 1976;21:111–8.
Ertel A, Eng R, Smith SM. The differential effect of cigarette smoke on the growth of bacteria found in humans. Chest. 1991;100:628–30.
Morris A, Beck JM, Schloss PD, Campbell TB, Crothers K, Curtis JL, Flores SC, Fontenot AP, Ghedin E, Huang L, et al. Comparison of the respiratory microbiome in healthy nonsmokers and smokers. Am J Respir Crit Care Med. 2013;187:1067–75.
Socransky SS, Haffajee AD. Periodontal microbial ecology. Periodontol. 2000;2005(38):135–87.
Tomar SL, Asma S. Smoking-attributable periodontitis in the United States: findings from NHANES III. National Health and Nutrition Examination Survey. J Periodontol. 2000;71:743–51.
Johnson GK, Hill M. Cigarette smoking and the periodontal patient. J Periodontol. 2004;75:196–209.
Hugoson A, Rolandsson M. Periodontal disease in relation to smoking and the use of Swedish snus: epidemiological studies covering 20 years (1983–2003). J Clin Periodontol. 2011;38:809–16.
Shchipkova AY, Nagaraja HN, Kumar PS. Subgingival microbial profiles of smokers with periodontitis. J Dent Res. 2010;89:1247–53.
Camelo-Castillo AJ, Mira A, Pico A, Nibali L, Henderson B, Donos N, Tomas I. Subgingival microbiota in health compared to periodontitis and the influence of smoking. Front Microbiol. 2015;6:119.
Moon JH, Lee JH, Lee JY. Subgingival microbiome in smokers and non-smokers in Korean chronic periodontitis patients. Mol Oral Microbiol. 2015;30:227–41.
Bizzarro S, Loos BG, Laine ML, Crielaard W, Zaura E. Subgingival microbiome in smokers and non-smokers in periodontitis: an exploratory study using traditional targeted techniques and a next-generation sequencing. J Clin Periodontol. 2013;40:483–92.
Duan X, Wu T, Xu X, Chen D, Mo A, Lei Y, Cheng L, Man Y, Zhou X, Wang Y, Yuan Q. Smoking may lead to marginal bone loss around non-submerged implants during bone healing by altering salivary microbiome: a prospective study. J Periodontol. 2017;88:1297–308.
Tsigarida AA, Dabdoub SM, Nagaraja HN, Kumar PS. The influence of smoking on the peri-implant microbiome. J Dent Res. 2015;94:1202–17.
Lalla E, Papapanou PN. Diabetes mellitus and periodontitis: a tale of two common interrelated diseases. Nat Rev Endocrinol. 2011;7:738–48.
Beck JD, Offenbacher S. Systemic effects of periodontitis: epidemiology of periodontal disease and cardiovascular disease. J Periodontol. 2005;76:2089–100.
Deshpande RG, Khan M, Genco CA. Invasion strategies of the oral pathogen porphyromonas gingivalis: implications for cardiovascular disease. Invasion Metastasis. 1998;18:57–69.
Parahitiyawa NB, Jin LJ, Leung WK, Yam WC, Samaranayake LP. Microbiology of odontogenic bacteremia: beyond endocarditis. Clin Microbiol Rev. 2009;22:46–64.
Verma D, Garg PK, Dubey AK. Insights into the human oral microbiome. Arch Microbiol. 2018;200:525–40.
Maraki S, Plevritaki A, Kofteridis D, Scoulica E, Eskitzis A, Gikas A, Panagiotakis SH. Bicuspid aortic valve endocarditis caused by Gemella sanguinis: case report and literature review. J Infect Public Health. 2019;12:304–8.
Zhu B, Macleod LC, Kitten T, Xu P. Streptococcus sanguinis biofilm formation & interaction with oral pathogens. Future Microbiol. 2018;13:915–32.
Zbinden A, Aras F, Zbinden R, Mouttet F, Schmidlin PR, Bloemberg GV, Bostanci N. Frequent detection of Streptococcus tigurinus in the human oral microbial flora by a specific 16S rRNA gene real-time TaqMan PCR. BMC Microbiol. 2014;14:231.
Matias WR, Bourque DL, Niwano T, Onderdonk AB, Katz JT. Subacute bacterial Endocarditis with Leptotrichia goodfellowii in a patient with a valvular allograft: a case report and review of the literature. Case Rep Infect Dis. 2016;2016:3051212.
Moser C, Pedersen HT, Lerche CJ, Kolpen M, Line L, Thomsen K, Hoiby N, Jensen PO. Biofilms and host response—helpful or harmful. APMIS. 2017;125:320–38.
Aas JA, Barbuto SM, Alpagot T, Olsen I, Dewhirst FE, Paster BJ. Subgingival plaque microbiota in HIV positive patients. J Clin Periodontol. 2007;34:189–95.
Mukherjee PK, Chandra J, Retuerto M, Sikaroodi M, Brown RE, Jurevic R, Salata RA, Lederman MM, Gillevet PM, Ghannoum MA. Oral mycobiome analysis of HIV-infected patients: identification of Pichia as an antagonist of opportunistic fungi. PLoS Pathog. 2014;10:e1003996.
Mukherjee PK, Chandra J, Retuerto M, Tatsuoka C, Ghannoum MA, McComsey GA. Dysbiosis in the oral bacterial and fungal microbiome of HIV-infected subjects is associated with clinical and immunologic variables of HIV infection. PLoS ONE. 2018;13:e0200285.
Kato I, Vasquez AA, Moyerbrailean G, Land S, Sun J, Lin HS, Ram JL. Oral microbiome and history of smoking and colorectal cancer. J Epidemiol Res. 2016;2:92–101.
Hilty M, Burke C, Pedro H, Cardenas P, Bush A, Bossley C, Davies J, Ervine A, Poulter L, Pachter L, et al. Disordered microbial communities in asthmatic airways. PLoS ONE. 2010;5:e8578.
Erb-Downward JR, Thompson DL, Han MK, Freeman CM, McCloskey L, Schmidt LA, Young VB, Toews GB, Curtis JL, Sundaram B, et al. Analysis of the lung microbiome in the “healthy” smoker and in COPD. PLoS ONE. 2011;6:e16384.
Mammen MJ, Sethi S. COPD and the microbiome. Respirology. 2016;21:590–9.
Pragman AA, Kim HB, Reilly CS, Wendt C, Isaacson RE. The lung microbiome in moderate and severe chronic obstructive pulmonary disease. PLoS ONE. 2012;7:e47305.
Millares L, Perez-Brocal V, Ferrari R, Gallego M, Pomares X, Garcia-Nunez M, Monton C, Capilla S, Monso E, Moya A. Functional metagenomics of the bronchial microbiome in COPD. PLoS ONE. 2015;10:e0144448.
Garmendia J, Morey P, Bengoechea JA. Impact of cigarette smoke exposure on host–bacterial pathogen interactions. Eur Respir J. 2012;39:467–77.
Ko FW, Chan KP, Hui DS, Goddard JR, Shaw JG, Reid DW, Yang IA. Acute exacerbation of COPD. Respirology. 2016;21:1152–65.
Arcavi L, Benowitz NL. Cigarette smoking and infection. Arch Intern Med. 2004;164:2206–16.
Read AF, Taylor LH. The ecology of genetically diverse infections. Science. 2001;292:1099–102.
Klainer AS, Beisel WR. Opportunistic infection: a review. Am J Med Sci. 1969;258:431–56.
Gregory AC, Sullivan MB, Segal LN, Keller BC. Smoking is associated with quantifiable differences in the human lung DNA virome and metabolome. Respir Res. 2018;19:174.
Thorburn AN, McKenzie CI, Shen S, Stanley D, Macia L, Mason LJ, Roberts LK, Wong CH, Shim R, Robert R, et al. Evidence that asthma is a developmental origin disease influenced by maternal diet and bacterial metabolites. Nat Commun. 2015;6:7320.
Fujimura KE, Sitarik AR, Havstad S, Lin DL, Levan S, Fadrosh D, Panzer AR, LaMere B, Rackaityte E, Lukacs NW, et al. Neonatal gut microbiota associates with childhood multisensitized atopy and T cell differentiation. Nat Med. 2016;22:1187–91.
Stokholm J, Blaser MJ, Thorsen J, Rasmussen MA, Waage J, Vinding RK, Schoos AM, Kunoe A, Fink NR, Chawes BL, et al. Maturation of the gut microbiome and risk of asthma in childhood. Nat Commun. 2018;9:141.
Gollwitzer ES, Saglani S, Trompette A, Yadava K, Sherburn R, McCoy KD, Nicod LP, Lloyd CM, Marsland BJ. Lung microbiota promotes tolerance to allergens in neonates via PD-L1. Nat Med. 2014;20:642–7.
Turturice BA, McGee HS, Oliver B, Baraket M, Nguyen BT, Ascoli C, Ranjan R, Rani A, Perkins DL, Finn PW. Atopic asthmatic immune phenotypes associated with airway microbiota and airway obstruction. PLoS ONE. 2017;12:e0184566.
Zhang Q, Cox M, Liang Z, Brinkmann F, Cardenas PA, Duff R, Bhavsar P, Cookson W, Moffatt M, Chung KF. Airway microbiota in severe asthma and relationship to asthma severity and phenotypes. PLoS ONE. 2016;11:e0152724.
Huang YJ, Nariya S, Harris JM, Lynch SV, Choy DF, Arron JR, Boushey H. The airway microbiome in patients with severe asthma: associations with disease features and severity. J Allergy Clin Immunol. 2015;136:874–84.
Huang YJ, Nelson CE, Brodie EL, Desantis TZ, Baek MS, Liu J, Woyke T, Allgaier M, Bristow J, Wiener-Kronish JP, et al. Airway microbiota and bronchial hyperresponsiveness in patients with suboptimally controlled asthma. J Allergy Clin Immunol. 2011;127(372–381):e371–3.
Green BJ, Wiriyachaiporn S, Grainge C, Rogers GB, Kehagia V, Lau L, Carroll MP, Bruce KD, Howarth PH. Potentially pathogenic airway bacteria and neutrophilic inflammation in treatment resistant severe asthma. PLoS ONE. 2014;9:e100645.
Goleva E, Jackson LP, Harris JK, Robertson CE, Sutherland ER, Hall CF, Good JT Jr, Gelfand EW, Martin RJ, Leung DY. The effects of airway microbiome on corticosteroid responsiveness in asthma. Am J Respir Crit Care Med. 2013;188:1193–201.
van Woerden HC, Gregory C, Brown R, Marchesi JR, Hoogendoorn B, Matthews IP. Differences in fungi present in induced sputum samples from asthma patients and non-atopic controls: a community based case control study. BMC Infect Dis. 2013;13:69.
Agbetile J, Fairs A, Desai D, Hargadon B, Bourne M, Mutalithas K, Edwards R, Morley JP, Monteiro WR, Kulkarni NS, et al. Isolation of filamentous fungi from sputum in asthma is associated with reduced post-bronchodilator FEV1. Clin Exp Allergy. 2012;42:782–91.
Chishimba L, Niven R, Fraczek M, Bowyer P, Smyth L, Simpson A, Denning D. Lung microbiome is associated with asthma severity in fungal associated asthma. Eur Respir J. 2015;46:OA1462.
Thomson NC, Chaudhuri R, Livingston E. Asthma and cigarette smoking. Eur Respir J. 2004;24:822–33.
Colak Y, Afzal S, Nordestgaard BG, Lange P. Characteristics and prognosis of never-smokers and smokers with asthma in the copenhagen general population study. A prospective cohort study. Am J Respir Crit Care Med. 2015;192:172–81.
Simpson JL, Daly J, Baines KJ, Yang IA, Upham JW, Reynolds PN, Hodge S, James AL, Hugenholtz P, Willner D, Gibson PG. Airway dysbiosis: Haemophilus influenzae and Tropheryma in poorly controlled asthma. Eur Respir J. 2016;47:792–800.
Lyczak JB, Cannon CL, Pier GB. Lung infections associated with cystic fibrosis. Clin Microbiol Rev. 2002;15:194–222.
Gibson RL, Burns JL, Ramsey BW. Pathophysiology and management of pulmonary infections in cystic fibrosis. Am J Respir Crit Care Med. 2003;168:918–51.
Lynch SV, Bruce KD. The cystic fibrosis airway microbiome. Cold Spring Harb Perspect Med. 2013;3:a009738.
Zemanick ET, Wagner BD, Robertson CE, Ahrens RC, Chmiel JF, Clancy JP, Gibson RL, Harris WT, Kurland G, Laguna TA, et al. Airway microbiota across age and disease spectrum in cystic fibrosis. Eur Respir J. 2017;50:1700832.
Mahenthiralingam E. Emerging cystic fibrosis pathogens and the microbiome. Paediatr Respir Rev. 2014;15(Suppl 1):13–5.
Fodor AA, Klem ER, Gilpin DF, Elborn JS, Boucher RC, Tunney MM, Wolfgang MC. The adult cystic fibrosis airway microbiota is stable over time and infection type, and highly resilient to antibiotic treatment of exacerbations. PLoS ONE. 2012;7:e45001.
Blainey PC, Milla CE, Cornfield DN, Quake SR. Quantitative analysis of the human airway microbial ecology reveals a pervasive signature for cystic fibrosis. Sci Transl Med. 2012;4:153ra130.
Willger SD, Grim SL, Dolben EL, Shipunova A, Hampton TH, Morrison HG, Filkins LM, O’Toole GA, Moulton LA, Ashare A, et al. Characterization and quantification of the fungal microbiome in serial samples from individuals with cystic fibrosis. Microbiome. 2014;2:40.
Kramer R, Sauer-Heilborn A, Welte T, Guzman CA, Abraham WR, Hofle MG. Cohort study of airway mycobiome in adult cystic fibrosis patients: differences in community structure between fungi and bacteria reveal predominance of transient fungal elements. J Clin Microbiol. 2015;53:2900–7.
Willner D, Furlan M, Haynes M, Schmieder R, Angly FE, Silva J, Tammadoni S, Nosrat B, Conrad D, Rohwer F. Metagenomic analysis of respiratory tract DNA viral communities in cystic fibrosis and non-cystic fibrosis individuals. PLoS ONE. 2009;4:e7370.
Rowe SM, Miller S, Sorscher EJ. Cystic fibrosis. N Engl J Med. 2005;352:1992–2001.
Campbell PW 3rd, Parker RA, Roberts BT, Krishnamani MR, Phillips JA 3rd. Association of poor clinical status and heavy exposure to tobacco smoke in patients with cystic fibrosis who are homozygous for the F508 deletion. J Pediatr. 1992;120:261–4.
Collaco JM, Vanscoy L, Bremer L, McDougal K, Blackman SM, Bowers A, Naughton K, Jennings J, Ellen J, Cutting GR. Interactions between secondhand smoke and genes that affect cystic fibrosis lung disease. JAMA. 2008;299:417–24.
Sloane PA, Shastry S, Wilhelm A, Courville C, Tang LP, Backer K, Levin E, Raju SV, Li Y, Mazur M, et al. A pharmacologic approach to acquired cystic fibrosis transmembrane conductance regulator dysfunction in smoking related lung disease. PLoS ONE. 2012;7:e39809.
Amaral MD, Kunzelmann K. Molecular targeting of CFTR as a therapeutic approach to cystic fibrosis. Trends Pharmacol Sci. 2007;28:334–41.
Tyc VL, Throckmorton-Belzer L. Smoking rates and the state of smoking interventions for children and adolescents with chronic illness. Pediatrics. 2006;118:e471–87.
Robles Alonso V, Guarner F. Intestinal microbiota composition in adults. World Rev Nutr Diet 2013;107:17–24.
Ananthakrishnan AN. Environmental risk factors for inflammatory bowel disease. Gastroenterol Hepatol. 2013;9:367–74.
Persson PG, Ahlbom A, Hellers G. Inflammatory bowel disease and tobacco smoke—a case–control study. Gut. 1990;31:1377–81.
Tomoda K, Kubo K, Asahara T, Andoh A, Nomoto K, Nishii Y, Yamamoto Y, Yoshikawa M, Kimura H. Cigarette smoke decreases organic acids levels and population of bifidobacterium in the caecum of rats. J Toxicol Sci. 2011;36:261–6.
Wang H, Zhao JX, Hu N, Ren J, Du M, Zhu MJ. Side-stream smoking reduces intestinal inflammation and increases expression of tight junction proteins. World J Gastroenterol. 2012;18:2180–7.
Rogers MA, Greene MT, Saint S, Chenoweth CE, Malani PN, Trivedi I, Aronoff DM. Higher rates of Clostridium difficile infection among smokers. PLoS ONE. 2012;7:e42091.
Lee SH, Yun Y, Kim SJ, Lee EJ, Chang Y, Ryu S, Shin H, Kim HL, Kim HN, Lee JH. Association between cigarette smoking status and composition of gut microbiota: population-based cross-sectional study. J Clin Med. 2018;7:282.
Benjamin JL, Hedin CR, Koutsoumpas A, Ng SC, McCarthy NE, Prescott NJ, Pessoa-Lopes P, Mathew CG, Sanderson J, Hart AL, et al. Smokers with active Crohn’s disease have a clinically relevant dysbiosis of the gastrointestinal microbiota. Inflamm Bowel Dis. 2012;18:1092–100.
Biedermann L, Zeitz J, Mwinyi J, Sutter-Minder E, Rehman A, Ott SJ, Steurer-Stey C, Frei A, Frei P, Scharl M, et al. Smoking cessation induces profound changes in the composition of the intestinal microbiota in humans. PLoS ONE. 2013;8:e59260.
Baumgart DC, Sandborn WJ. Crohn’s disease. Lancet. 2012;380:1590–605.
Stefanelli T, Malesci A, Repici A, Vetrano S, Danese S. New insights into inflammatory bowel disease pathophysiology: paving the way for novel therapeutic targets. Curr Drug Targets. 2008;9:413–8.
Cho JH, Brant SR. Recent insights into the genetics of inflammatory bowel disease. Gastroenterology. 2011;140:1704–12.
Dessein R, Chamaillard M, Danese S. Innate immunity in Crohn’s disease: the reverse side of the medal. J Clin Gastroenterol. 2008;42(Suppl 3 Pt 1):S144–7.
Colombel JF. Decade in review-IBD: IBD-genes, bacteria and new therapeutic strategies. Nat Rev Gastroenterol Hepatol. 2014;11:652–4.
Chassaing B, Darfeuille-Michaud A. The commensal microbiota and enteropathogens in the pathogenesis of inflammatory bowel diseases. Gastroenterology. 2011;140:1720–8.
Swidsinski A, Loening-Baucke V, Herber A. Mucosal flora in Crohn’s disease and ulcerative colitis—an overview. J Physiol Pharmacol. 2009;60(Suppl 6):61–71.
Imhann F, Vich Vila A, Bonder MJ, Fu J, Gevers D, Visschedijk MC, Spekhorst LM, Alberts R, Franke L, van Dullemen HM, et al. Interplay of host genetics and gut microbiota underlying the onset and clinical presentation of inflammatory bowel disease. Gut. 2018;67:108–19.
Fava F, Danese S. Intestinal microbiota in inflammatory bowel disease: friend of foe? World J Gastroenterol. 2011;17:557–66.
Parkes GC, Whelan K, Lindsay JO. Smoking in inflammatory bowel disease: impact on disease course and insights into the aetiology of its effect. J Crohns Colitis. 2014;8:717–25.
Opstelten JL, Plassais J, van Mil SW, Achouri E, Pichaud M, Siersema PD, Oldenburg B, Cervino AC. Gut microbial diversity is reduced in smokers with Crohn’s disease. Inflamm Bowel Dis. 2016;22:2070–7.
Gonzalez-Sarrias A, Larrosa M, Tomas-Barberan FA, Dolara P, Espin JC. NF-kappaB-dependent anti-inflammatory activity of urolithins, gut microbiota ellagic acid-derived metabolites, in human colonic fibroblasts. Br J Nutr. 2010;104:503–12.
Sokol H, Pigneur B, Watterlot L, Lakhdari O, Bermudez-Humaran LG, Gratadoux JJ, Blugeon S, Bridonneau C, Furet JP, Corthier G, et al. Faecalibacterium prausnitzii is an anti-inflammatory commensal bacterium identified by gut microbiota analysis of Crohn disease patients. Proc Natl Acad Sci USA. 2008;105:16731–6.
Huttenhower C, Kostic AD, Xavier RJ. Inflammatory bowel disease as a model for translating the microbiome. Immunity. 2014;40:843–54.
Jacobs JP, Goudarzi M, Singh N, Tong M, McHardy IH, Ruegger P, Asadourian M, Moon BH, Ayson A, Borneman J, et al. A disease-associated microbial and metabolomics state in relatives of pediatric inflammatory bowel disease patients. Cell Mol Gastroenterol Hepatol. 2016;2:750–66.
Sokol H, Seksik P. The intestinal microbiota in inflammatory bowel diseases: time to connect with the host. Curr Opin Gastroenterol. 2010;26:327–31.
Reshef L, Kovacs A, Ofer A, Yahav L, Maharshak N, Keren N, Konikoff FM, Tulchinsky H, Gophna U, Dotan I. Pouch inflammation is associated with a decrease in specific bacterial taxa. Gastroenterology. 2015;149:718–27.
Duranti S, Gaiani F, Mancabelli L, Milani C, Grandi A, Bolchi A, Santoni A, Lugli GA, Ferrario C, Mangifesta M, et al. Elucidating the gut microbiome of ulcerative colitis: bifidobacteria as novel microbial biomarkers. FEMS Microbiol Ecol. 2016;92:fiw191.
de Souza HS, Fiocchi C. Immunopathogenesis of IBD: current state of the art. Nat Rev Gastroenterol Hepatol. 2016;13:13–27.
Rogler G, Vavricka S. Exposome in IBD: recent insights in environmental factors that influence the onset and course of IBD. Inflamm Bowel Dis. 2015;21:400–8.
Singh UP, Singh NP, Murphy EA, Price RL, Fayad R, Nagarkatti M, Nagarkatti PS. Chemokine and cytokine levels in inflammatory bowel disease patients. Cytokine. 2016;77:44–9.
Zhai H, Huang W, Liu A, Li Q, Hao Q, Ma L, Yang F, Zhang S. Current smoking improves ulcerative colitis patients’ disease behaviour in the northwest of China. Prz Gastroenterol. 2017;12:286–90.
Wang YF, Ou-Yang Q, Xia B, Liu LN, Gu F, Zhou KF, Mei Q, Shi RH, Ran ZH, Wang XD, et al. Multicenter case–control study of the risk factors for ulcerative colitis in China. World J Gastroenterol. 2013;19:1827–33.
Ng SC, Tang W, Leong RW, Chen M, Ko Y, Studd C, Niewiadomski O, Bell S, Kamm MA, de Silva HJ, et al. Environmental risk factors in inflammatory bowel disease: a population-based case–control study in Asia-Pacific. Gut. 2015;64:1063–71.
Li LF, Chan RL, Lu L, Shen J, Zhang L, Wu WK, Wang L, Hu T, Li MX, Cho CH. Cigarette smoking and gastrointestinal diseases: the causal relationship and underlying molecular mechanisms (review). Int J Mol Med. 2014;34:372–80.
Altarescu G, Rachmilewitz D, Zevin S. Relationship between CYP2A6 genetic polymorphism, as a marker of nicotine metabolism, and ulcerative colitis. Isr Med Assoc J. 2011;13:87–90.
Onyiah JC, Sheikh SZ, Maharshak N, Otterbein LE, Plevy SE. Heme oxygenase-1 and carbon monoxide regulate intestinal homeostasis and mucosal immune responses to the enteric microbiota. Gut Microbes. 2014;5:220–4.
Dallongeville J, Marecaux N, Fruchart JC, Amouyel P. Cigarette smoking is associated with unhealthy patterns of nutrient intake: a meta-analysis. J Nutr. 1998;128:1450–7.
Torre LA, Bray F, Siegel RL, Ferlay J, Lortet-Tieulent J, Jemal A. Global cancer statistics, 2012. CA Cancer J Clin. 2015;65:87–108.
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68:394–424.
Rogler G. Chronic ulcerative colitis and colorectal cancer. Cancer Lett. 2014;345:235–41.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144:646–74.
Saus E, Iraola-Guzman S, Willis JR, Brunet-Vega A, Gabaldon T. Microbiome and colorectal cancer: Roles in carcinogenesis and clinical potential. Mol Aspects Med. 2019. https://doi.org/10.1016/j.mam.2019.05.001.
Hoppes WL, Lerner PI. Nonenterococcal group-D streptococcal endocarditis caused by Streptococcus bovis. Ann Intern Med. 1974;81:588–93.
Wang T, Cai G, Qiu Y, Fei N, Zhang M, Pang X, Jia W, Cai S, Zhao L. Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. ISME J. 2012;6:320–9.
Feng Q, Liang S, Jia H, Stadlmayr A, Tang L, Lan Z, Zhang D, Xia H, Xu X, Jie Z, et al. Gut microbiome development along the colorectal adenoma–carcinoma sequence. Nat Commun. 2015;6:6528.
Yu J, Feng Q, Wong SH, Zhang D, Liang QY, Qin Y, Tang L, Zhao H, Stenvang J, Li Y, et al. Metagenomic analysis of faecal microbiome as a tool towards targeted non-invasive biomarkers for colorectal cancer. Gut. 2017;66:70–8.
Gao Z, Guo B, Gao R, Zhu Q, Qin H. Microbiota disbiosis is associated with colorectal cancer. Front Microbiol. 2015;6:20.
Nakatsu G, Zhou H, Wu WKK, Wong SH, Coker OO, Dai Z, Li X, Szeto CH, Sugimura N, Lam TY, et al. Alterations in enteric virome are associated with colorectal cancer and survival outcomes. Gastroenterology. 2018;155(529–541):e525.
Coker OO, Nakatsu G, Dai RZ, Wu WKK, Wong SH, Ng SC, Chan FKL, Sung JJY, Yu J. Enteric fungal microbiota dysbiosis and ecological alterations in colorectal cancer. Gut. 2019;68:654–62.
Song M, Chan AT. Environmental factors, gut microbiota, and colorectal cancer prevention. Clin Gastroenterol Hepatol. 2019;17:275–89.
Budden KF, Gellatly SL, Wood DL, Cooper MA, Morrison M, Hugenholtz P, Hansbro PM. Emerging pathogenic links between microbiota and the gut–lung axis. Nat Rev Microbiol. 2017;15:55–63.
Bozkurt HS, Quigley EM, Kara B. Bifidobacterium animalis subspecies lactis engineered to produce mycosporin-like amino acids in colorectal cancer prevention. SAGE Open Med. 2019;7:2050312119825784.
Hu J, Wei T, Sun S, Zhao A, Xu C. Effects of cigarette smoke condensate on the production and characterization of exopolysaccharides by Bifidobacterium. An Acad Bras Cienc. 2015;87:997–1005.
Kurata N, Tokashiki N, Fukushima K, Misao T, Hasuoka N, Kitagawa K, Mashimo M, Regan JW, Murayama T, Fujino H. Short chain fatty acid butyrate uptake reduces expressions of prostanoid EP4 receptors and their mediation of cyclooxygenase-2 induction in HCA-7 human colon cancer cells. Eur J Pharmacol. 2019;853:308–15.
Allais L, Kerckhof FM, Verschuere S, Bracke KR, De Smet R, Laukens D, Van den Abbeele P, De Vos M, Boon N, Brusselle GG, et al. Chronic cigarette smoke exposure induces microbial and inflammatory shifts and mucin changes in the murine gut. Environ Microbiol. 2016;18:1352–63.
Berer K, Gerdes LA, Cekanaviciute E, Jia X, Xiao L, Xia Z, Liu C, Klotz L, Stauffer U, Baranzini SE, et al. Gut microbiota from multiple sclerosis patients enables spontaneous autoimmune encephalomyelitis in mice. Proc Natl Acad Sci USA. 2017;114:10719–24.
Zhang X, Zhang D, Jia H, Feng Q, Wang D, Liang D, Wu X, Li J, Tang L, Li Y, et al. The oral and gut microbiomes are perturbed in rheumatoid arthritis and partly normalized after treatment. Nat Med. 2015;21:895–905.
Wen C, Zheng Z, Shao T, Liu L, Xie Z, Le Chatelier E, He Z, Zhong W, Fan Y, Zhang L, et al. Quantitative metagenomics reveals unique gut microbiome biomarkers in ankylosing spondylitis. Genome Biol. 2017;18:142.
He Z, Shao T, Li H, Xie Z, Wen C. Alterations of the gut microbiome in Chinese patients with systemic lupus erythematosus. Gut Pathog. 2016;8:64.
Morgan XC, Tickle TL, Sokol H, Gevers D, Devaney KL, Ward DV, Reyes JA, Shah SA, LeLeiko N, Snapper SB, et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012;13:R79.
Vaahtovuo J, Munukka E, Korkeamaki M, Luukkainen R, Toivanen P. Fecal microbiota in early rheumatoid arthritis. J Rheumatol. 2008;35:1500–5.
Bernard NJ. Rheumatoid arthritis: Prevotella copri associated with new-onset untreated RA. Nat Rev Rheumatol. 2014;10:2.
Ye Z, Zhang N, Wu C, Zhang X, Wang Q, Huang X, Du L, Cao Q, Tang J, Zhou C, et al. A metagenomic study of the gut microbiome in Behcet’s disease. Microbiome. 2018;6:135.
Miyake S, Kim S, Suda W, Oshima K, Nakamura M, Matsuoka T, Chihara N, Tomita A, Sato W, Kim SW, et al. Dysbiosis in the gut microbiota of patients with multiple sclerosis, with a striking depletion of species belonging to Clostridia XIVa and IV clusters. PLoS ONE. 2015;10:e0137429.
Klareskog L, Padyukov L, Alfredsson L. Smoking as a trigger for inflammatory rheumatic diseases. Curr Opin Rheumatol. 2007;19:49–54.
Bang SY, Lee KH, Cho SK, Lee HS, Lee KW, Bae SC. Smoking increases rheumatoid arthritis susceptibility in individuals carrying the HLA-DRB1 shared epitope, regardless of rheumatoid factor or anti-cyclic citrullinated peptide antibody status. Arthritis Rheum. 2010;62:369–77.
Ludvigsson JF, Nordenvall C, Jarvholm B. Smoking, use of moist snuff and risk of celiac disease: a prospective study. BMC Gastroenterol. 2014;14:120.
Kaan U, Ferda O. Evaluation of clinical activity and functional impairment in smokers with ankylosing spondylitis. Rheumatol Int. 2005;25:357–60.
Simon KC, Munger KL, Ascherio A. XVI European Charcot Foundation lecture: nutrition and environment: can MS be prevented? J Neurol Sci. 2011;311:1–8.
Ascherio A, Munger KL. Environmental risk factors for multiple sclerosis. Part II: noninfectious factors. Ann Neurol. 2007;61:504–13.
Wu J, Peters BA, Dominianni C, Zhang Y, Pei Z, Yang L, Ma Y, Purdue MP, Jacobs EJ, Gapstur SM, et al. Cigarette smoking and the oral microbiome in a large study of American adults. ISME J. 2016;10:2435–46.
Valles Y, Inman CK, Peters BA, Ali R, Wareth LA, Abdulle A, Alsafar H, Anouti FA, Dhaheri AA, Galani D, et al. Types of tobacco consumption and the oral microbiome in the United Arab Emirates Healthy Future (UAEHFS) Pilot Study. Sci Rep. 2018;8:11327.
Charlson ES, Chen J, Custers-Allen R, Bittinger K, Li H, Sinha R, Hwang J, Bushman FD, Collman RG. Disordered microbial communities in the upper respiratory tract of cigarette smokers. PLoS ONE. 2010;5:e15216.
Zhang R, Chen L, Cao L, Li KJ, Huang Y, Luan XQ, Li G. Effects of smoking on the lower respiratory tract microbiome in mice. Respir Res. 2018;19:253.
Acknowledgements
None.
Funding
This study was supported by Grant 81770025 from National Natural Science Foundation of China, Grant ZH2018QNA48 from Cross research funds of Translational Medicine and Grant 2017ZZ02014 from Shanghai key discipline for respiratory diseases.
Author information
Authors and Affiliations
Contributions
CH wrote the review, GS revised the manuscript. Both authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Huang, C., Shi, G. Smoking and microbiome in oral, airway, gut and some systemic diseases. J Transl Med 17, 225 (2019). https://doi.org/10.1186/s12967-019-1971-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12967-019-1971-7