
Abstract
Background
Liver, as a vital organ, is responsible for a wide range of biological functions to maintain homeostasis and any type of damages to hepatic tissue contributes to disease progression and death. Viral infection, trauma, carcinoma, alcohol misuse and inborn errors of metabolism are common causes of liver diseases are a severe known reason for leading to end-stage liver disease or liver failure. In either way, liver transplantation is the only treatment option which is, however, hampered by the increasing scarcity of organ donor. Over the past years, considerable efforts have been directed toward liver regeneration aiming at developing new approaches and methodologies to enhance the transplantation process. These approaches include producing decellularized scaffolds from the liver organ, 3D bio-printing system, and nano-based 3D scaffolds to simulate the native liver microenvironment. The application of small molecules and micro-RNAs and genetic manipulation in favor of hepatic differentiation of distinct stem cells could also be exploited. All of these strategies will help to facilitate the application of stem cells in human medicine. This article reviews the most recent strategies to generate a high amount of mature hepatocyte-like cells and updates current knowledge on liver regenerative medicine.
Similar content being viewed by others


Background
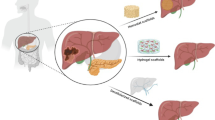
The liver is the largest gland of the body, which normally weighs about 1.5 kg in adults and divided into a large right lobe and a smaller left lobe [1]. Each lobe is further divided into lobules, which are the functioning units of the liver. The lobule is consisting of a hexagonal row of hepatocytes. Primary hepatocytes constitute 60–80% of the liver mass and play many important functions in our body. The main functions played by the liver include (a) bile production and secretion; (b) excretion of bilirubin, cholesterol, hormones, and drugs; (c) metabolism of fats, proteins, and carbohydrates; (d) enzyme activation; (e) storage of glycogen, vitamins, and minerals; (f) macromolecules and protein synthesis (i.e. Alb and bile acids); and (g) detoxification [2]. Detoxification is a critical liver-specific function [3]. Exogenous and endogenous substances are detoxified in the liver by two main mechanisms, phase I and phase II biotransformation [4, 5]. Hepatocyte-based hepatotoxicity testing is useful in the rapid screening of chemicals and in the mechanistic evaluation of toxicological phenomena. A large amount of natural and synthetic chemicals are hepatotoxins. In many cases, the hepatotoxicity is due to an impaired hepatocyte metabolism and conversion of inert and non-toxic compounds into highly reactive metabolites. The hepatocytes are usually the first cell types that are damaged upon hepatotoxic insult [6]. The loss of liver functions such as detoxification, metabolism, and regulation causes life-threatening complications, including kidney failure, encephalopathy, cerebral edema, severe hypotension and susceptibility to infections culminating in multiple organ failure. Hepatocytes-based screening can be used to characterize the metabolic fate of compounds and whether metabolism contributes to toxicity. Therefore, approaches to screen hepatocytes are of great biomedical importance. The bioartificial engineering liver constructs have been recently developed [7, 8]. The efforts in the modern liver tissue engineering field mainly include: (1) creating a whole, implantable, and functional tissue-engineered liver constructs; (2) establishing bioartificial liver systems to sustain liver patient’s lives before liver transplantation, establishing in vitro hepatocyte-based model; (3) establishing a culture model for drug metabolism/toxicity screening for drug discovery and (4) for basic researchers of liver regeneration, disease, pathophysiology and pharmacology (Figs. 1, 2).

Hepatic tissues from various sources such as human, porcine and rat underwent decellularization using detergents. Repopulation of the decellularized liver scaffold is performed through various routes. Decellularized liver scaffold is chopped to cubes or thin slices then recellularized by microinjection. Whole-organ reseeded by exerting negative pressure suction. Various cell types such as induced pluripotent stem cells (iPSCs), mesenchymal stem cells (MSCs) or liver progenitor cells (LPCs) are used for recellularization

The impact of various nanostructures on hepatic differentiation. Nanofibers (high surface area, high porosity), nanoparticles (efficiently growth factors delivery) and carbon nanotube (mechanical properties, easily functionalized, aligned as collagen
The liver is an important organ because of critical metabolic functions such as protein synthesis and xenobiotic biotransformation [9]. The liver possesses remarkable regenerative capacity but viral infection, toxic compounds, and cancer resulted in vast injury and liver dysfunction, if continued, leading to end-stage liver pathologies and hepatic failure which can threaten life. In the case of liver transplantation immunological incompatibility between the donor and recipient, however, limits the application of this procedure [10,11,12]. Transplantation at cell levels such as hepatocyte provides several benefits over liver transplantation, can be performed several times and offer a less invasive alternative to whole liver transplantation, but it demonstrates low levels of engraftments [13]. Therefore, researchers bring forward to test novel tissue engineering approaches aiming to fabricate 3D hepatic tissue in microscale size or whole bioengineered liver synthesis to restore damaged liver function even after massive injury and resolve the donor shortage problem. Over the past years, the liver bioengineering field has experienced significant progress in the area of cell engineering, biomaterials fabrication, and tissue architecture to recapitulate transplantable microscale liver tissue and whole organ bioengineering as well [14]. Genetic engineering strategies and hepatic differentiation of stem cells are also under tight investigations [15, 16]. A wide range of cells from hepatocyte primary cells to hepatocyte-like cells that generated from hiPSCs constitute the promising source of stem cells which can be expanded in a high quantity that has appropriate compatibility with the host immune system. Considerable efforts have been dedicated to evaluating decellularized liver scaffolds in the construction of natural 3D extracellular matrix to better integration of mechanical and chemical signals for stem cell differentiation and maturation (Table 1). In addition, as a new 3D culture system, the perfusion-based culture approaches have been emerged and showed better potential to model in vivo tissue microenvironment state. In spite of conventional 2D monolayer systems, the cells can pile on top of each other in perfusion based culture system which in turn leads to improve material exchange and cell to cell communication, especially in prolonged culture periods [17, 18]. Besides liver tissue engineering, various nanomaterials and nanoparticles are used in in vitro hepatic differentiation of stem cells. Nanomaterials can emulate native liver ECM and nanoparticles due to their biodegradability and good biocompatibility are used to direct delivery of hepatogenic small molecules, growth factors, cytokines and proteins to stem cells [19]. 3D bioprinting system, as a sophisticated engineering methodology, has been improved to solve the issues related to conventional 2D culture technique, by providing clues essential for the dynamic of distinct cell types in the context of in vivo milieu. Evidence point that cell performance is enhanced by promoting the juxtacrine cell-to-cell interaction in the massive scale and 3D printing system has the potential to fabricate the distinctive constructions applicable to target tissues and organs [20,21,22]. In the 3D printing system, a computer identifies a 3D spatial model of the target organ and cuts it into sequences of 2D slices, which is then assembled from bottom to the top. This system is able to print and copy any preferred shapes with the facility to direct cell distribution, scaffold pore size, interconnectivity, and geometry. It is noteworthy that if the operator error is reduced, it can contribute to the formation of optimal uniform shapes on a large scale structure [23,24,25]. Apart from the mentioned approaches, some valuable steps have been taken in the application of small molecules and micro-RNAs as well as genetic manipulation in hepatic differentiation of stem cells. Small molecules have been reported to play significant roles in developmental processes through modulation of some signaling pathways involved in cell fate determination such as Wnt, Notch, and FGF during cell differentiation [26]. The application of small molecules in in vitro hepatic induction protocols demonstrated promising results through the assessment of hepatic markers [16]. MicroRNAs are constitutively or transiently expressed during differentiation and involved in cell fate determination programs of stem cells and researchers recently investigated the impact of microRNAs alone or in combination with each other as well suppression of some of them on hepatic differentiation of stem cells [27, 28]. In parallel, it has been shown that transcription factors such as FGF, HGF, Wnt, BMP, RA, and TGFβ to correlate with effectors influencing liver development. Therefore, it has been expected that transferring and integration, overexpression or knockdown of one or more than one transcription factors in stem cells can be able to enhance expression hepatic commitment markers [29]. In this review, the authors attempt to discuss the most recently addressed approaches aiming to improve the hepatic differentiation rate by different tissue engineering modalities as a new avenue for transplantation after hepatic failure or as therapy for liver regeneration and replacement (Figs. 3, 4).

Schematic overviews of multi-material 3D bioprinting approach. Sequential 3D bioprinted hepatic lobule-like structures (a). Simultaneous deposited and dual fabricated 3D structures (b)

A summary of methods used to hepatic differentiation of stem cells using intracellular signaling pathway. DMSO dimethyl sulfoxide, DKK-1 Dikkopf-related protein-1, HNF-3β hepatocyte nuclear factor 3-β, PrP-1 poly-ADP-ribose polymerase-1
Cell sources for liver regeneration
To generate donor-free and expandable hepatocyte cells source, several types of cells are exploited in liver tissue engineering. Based on previous studies in this area, these cells include a primary culture of hepatocytes, ESCs, iPSCs, and MSCs. ESCs are originated from the inner cell mass of blastocysts. To obtain iPSCs, adult somatic cells are genetically manipulated and reprogramed. For this propose, expression of pluripotency factors such as Oct4, Sox2, c-Myc, and klf4 is stimulated in the target cells [30]. It should be noted that MSCs are commonly isolated from almost all connective tissues mainly bone marrow medullary niche and adipose tissue. Using primary cell culture strategy, expanded hepatocytes retain and preserve specific functions such as drug metabolism activity and etc. which are comparable to the in vivo condition; however, prolonged in vitro expansion may lead to cell survival decrease and cell-specific function removal. In addition, it should not be forgotten that In addition, these cells should be freshly prepared from the patients to prevent immune cell reactivity and transplant rejection. To circumvent these pitfalls, great efforts have been devoted to improving functional behavior in the primary culture of hepatocytes. For instance, the application of 2D, 3D culture models, and perfusion-based microfluidic systems are at the center of attention [31, 32].
Perfusion-based systems are able to simultaneously replace fresh medium with the exhausted medium and continuously eliminate metabolic byproducts from the culture condition. Several experiments have highlighted an enhanced of hepatic cells function expanded in 2D, 3D culture models and perfusion-based systems, indicated by the up-regulation of liver function factors. ESCs and iPSCs possess high self-renewal capability that facilitates trans-differentiation into multiple cell lineages under specific conditions. It has been shown that the presence of specific growth factors, cytokines, and small molecules could increase differentiation properties. For instance, in a recent study, it was shown that ESCs could differentiate into hepatocyte-like cells in a stepwise manner using small molecules LY294002, touted as definitive endoderm inducer, bromo-indirubin-3′-oxime, odium butyrate, dimethyl sulfoxide and growth factor activin A. Among these factors, bromo-indirubin-3′-oxime, odium butyrate could dictate cells to acquire hepatoblast-like phenotype while dimethyl sulfoxide could accelerate orientation of progenitor cells toward mature hepatocyte-like cells. Differentiated cells have the ability to express hepatic cells specific factors and products such as urea, Alb and cytochrome p450 enzymes. In addition drug detoxification activity was similar to the human primary hepatocytes [33]. Scientific reports have pointed that iPSCs have some advantages over the ESCs. The use of iPSCs does not provoke immune cell activity and there are ethical issues exist surrounding the transplantation of ESCs. Recently, Rashidi et al. differentiated human iPSCs cells, lines FSPS13B and P106, into definitive endoderm cells by using activin A and bFGF followed by cell maturation into hepatocytes in the presence of HGF and OSM in a spheroid culture system. These spheroids were functional for more than 1 year and showed hepatic cells function and expressed maturation markers. It was found that these spheroids can partially support liver function in hepatectomized animal model after subcutaneous or intraperitoneal transplantation [34]. Similar to iPSCs and ESCs, MSCs also have shown a high hepatic differentiation potential either in vivo or in vitro model [35]. Bone marrow-derived MSCs demonstrated an enhanced expression of hepatocyte-specific markers and exhibited hepatocellular function after introduction to the liver decellularized scaffold in the presence of EGF and HGF (Fig. 5).

Some of the chemical structures related to small molecules are used commonly for hepatic-like phenotype induction from progenitor cells
In another study, iPSCs-derived MSCs and iPSCs were successfully transplanted into a hepatectomized rat to reduce hepatic injury. The cells were successful tracked in the rat hepatic tissue even after 2 months. The reduction of systemic bilirubin and increase of liver-specific markers such as Alb, cytokeratin-18, and α-fetoprotein were reported [36]. All these findings highlight the potency of ESCs, iPSCs, and MSCs to differentiate into hepatocyte-like cells. It seems that an inherent capacity of stem cells to circumvent problems related to post-transplantation hosts makes these cells superior to hepatocyte primary cells in the regeneration of hepatic tissue. Nevertheless, we must not forget that stem cells are not a magic bullet for the regeneration of hepatic tissue. Establishing precise criteria and measurement of differentiation capacity in the hepatic tissue must be carefully monitored after stem cell introduction.
Multicellular interactions in the hepatic tissue
Although in vitro culture models demonstrate hepatic tissue function, but these models could not completely restore the vital organ function such as metabolism and synthesis. All these systems mainly concentrate on the hepatocyte cells expansion. To achieve fully functional liver tissue, we need all cell types to exist in the liver, including PCs and NPCs cells [37]. Liver tissue is composed of NPCs such as Kupffer cells, endothelial cells and hepatic stellate cells that perform liver functions in collaboration with hepatocytes. The space between the liver NPC and hepatocyte is filled with a protein-rich material so-called Space of Disse. This space plays an important role in liver function through the regulation of materials and nutrients exchanging between endothelial cells and hepatocytes [38]. The conventional culture models of hepatocytes expansion are unable to mimic normal hepatic tissue microarchitecture. The function of hepatocytes correlates with the promotion of hepatocyte–hepatocyte, hepatocyte–ECM and hepatocyte–NPCs interaction. The lack of reciprocal interactions, as seen in conventional culture systems, leads to hepatocyte rapidly lose hepatocyte-specific bioactivities [37]. Due to the important role of NPC in liver function, recently magnificent efforts are ongoing to develop culture systems that incorporate multiple cell types including PCs and NPCs in hepatocyte culture [39]. It has shown that the co-culture system consisted of hepatocytes and NPCs promoted hepatocytes function through the initiation of hepatocyte-different cell interaction both in juxtacrine and paracrine manners. Calling attention, each cell of liver tissue plays vital roles in maintaining liver function [40]. By using a culture system composed of three rat liver cells including hepatocytes, Kupffer and endothelial cells on a polymer surface, A Space of Disse-like structure was reconstructed. It was shown that all these cells proliferated and preserve their morphologies in a 3D system compared to the conventional 2D culture. The expression of Alb and Cytochrome 450 was higher when stiffness of the scaffold was similar to the normal liver [41]. In another study, the hepatic differentiation was shown in a co-culture system using human iPSCs and adipose microvascular endothelial cells at a ratio of 3:1. Endothelial cells were used ad supporting cells in hepatic differentiation. Differentiated cells in co-culture system exhibited higher hepatocyte-like function in a wide range compared to single human iPSCs cultured either in vitro or in vivo. Enzymes related to the detoxification, ammonia synthesis and the level of Alb and coagulation factors were higher using multicellular culture compared to the single-cell model [42]. All these findings point that the application of multicellular differentiation strategies could be a novel technique in the restoration of liver function.
Tissue engineering strategy
Diabetes, heart failure, and hepatic failure are diseases of an enormous burden to the world, and current therapies for these often-lethal diseases are clearly inadequate. Tissue engineering holds great promise for the treatment of these diseases [43]. Millions of people currently suffer from the effects of chronic disease. Due to the limited availability of donors, only a fraction of individuals who could benefit from organ transplantations actually receive them. One possible avenue for remedying this situation is to artificially engineer human tissues [44,45,46,47,48,49,50,51]. One of the central themes of tissue engineering is to reproduce the body’s architectural and geometric intricacies, including vital cell–cell interactions. Tissue engineering techniques have been successfully applied to engineer many types of tissues; however, many challenges regarding their development still remain [44]. Numerous strategies have been developed to engineer tissues, with the most commonly used technique exploiting transplanted biofactors, such as cells, genes or proteins, into a porous degradable material called a scaffold. These scaffolds serve as synthetic ECM that organize the embedded cells into a 3D architecture and present them with stimuli for their growth and maturation. The ideal scaffold (1) contains properties that facilitate cell attachment, (2) contains a nutrient-rich environment to maintain cell viability and (3) biologically degrades overtime at a rate similar to the rate at which cells deposit matrix and proliferate in their new environment. One of the most commonly used approaches to tissue engineering is to seed cells on biodegradable scaffolds. These scaffolds bring cells in close proximity to each other and enable the formation of cell–cell contacts that mimic cells in the body. Ideally, the scaffold degrades at a rate similar to the rate of cell growth and matrix deposition, eventually completely replacing the synthetic scaffold with natural cells and matrix. However, challenges remain with this approach. For example, lack of vascularity in most tissue engineering constructs results in cell necrosis and loss of function, thus limiting the maximum size of tissue engineering constructs. For more detailed information on bioengineered tissue vascularization, readers may refer to the review paper of Kottamasu et al. [52]. In addition, difficulties in uniformly seeding cells throughout the various scaffolds prevent high initial cell seeding densities and finally the inability to mimic the complex cell–microenvironmental interactions, such as 3D orientation and architecture of cells, as well as homotypic and heterotypic cell–cell contact with microscale resolution [53].
Three-dimensional tissue constructs
Over the years much has been attempted in generating hepatic tissue-engineered organs. One strategy for engineering 3D hepatic tissues is to cultivate cells within biodegradable scaffolds made from either natural [54, 55] or synthetic [56,57,58] materials. The scaffolds function as a 3D structure on which the hepatic tissue may be induced to grow. These scaffolds aim to mimic in vivo conditions and facilitate the delivery of nutrients, oxygen, and other factors [57,58,59,60]. The addition of specific microenvironmental factors can be used to enhance the function of these engineered organs. For example, by co-culturing non-parenchymal cells such as endothelial cells and fibroblasts, hepatocyte function can be prolonged [60]. A major challenge in liver tissue engineering is that the liver cells quickly lose their differentiated function. This is in contrast to the behaviors of hepatocytes in the body which have the capability to regenerate. Thus, it is desirable to formulate alternative approaches to more precisely control the organization of cells and vascularization of engineered hepatic tissues.
Traditional 3D scaffolding approaches are not suitable for generating such complex structures due to a lack of control of the tissue architecture and cell–cell interactions. In particular, hepatic cells in 2D culture as well as within traditional 3D scaffolds simply do not organize as they do in normal tissue; their metabolic properties are therefore unsuitable for liver tissue engineering applications. Hepatic cells or liver tissues are usually cultured in vitro in diagnostic applications or before implantation in therapeutic applications, where they interact directly within different natural or synthetic biomaterials or scaffolds for growth and functional liver maintenance. Biomaterials technology aims to make an advance in hepatic tissue engineering by developing the basis for fabricating tissues made from soft materials such as hydrogels with engineered microvasculature. Although engineering microscale features into tissue engineering scaffolds have been attempted before [53, 61,62,63,64,65], there are still several approaches that will eliminate the difficulties associated with other micro-fabricated tissue engineering scaffolds such as uniform cell-seeding.
Hydrogels
Hydrogels have attracted great interest as scaffolding materials for tissue engineering because of their high water content, biocompatibility, and mechanical properties, which resemble those of natural tissues [66, 67]. Hydrogels have been used for tissue engineering of bone [68,69,70], cartilage [71,72,73], vascular [74] and other tissues [75, 76]. By adding cells to a hydrogel precursor prior to the gelling process, cells can be distributed homogeneously throughout the gel. In addition, hydrogels can be used to deliver soluble or immobilized signaling molecules to cells, act as support structures for cell growth and function, and provide space-filling for future tissue ingrowth [66, 67, 77, 78]. In general, hydrogels from natural sources can be derived from polymers such as collagen, HA, fibrin, alginate, agarose or chitosan [66]. Depending on their origin and composition, various natural polymers have specific utilities and properties. Many natural polymers, such as collagen, HA and fibrin, are derived from various components of the mammalian ECM. The advantages of natural polymers include low toxicity and biocompatibility. Collagen and other protein-based polymers are effective matrices for cellular growth, as they contain many cell signaling domains present in the in vivo ECM. Collagen gels can be naturally created without chemical modifications. However, in many cases, these gels are mechanically weak. To synthesize gels with enhanced mechanical properties various methods have been developed such as chemical crosslinking [79, 80], crosslinking with UV or temperature [79, 81], or in mixture with other polymeric agents [79, 82]. Hydrogels can be used to deliver soluble or immobilized signaling molecules to cells, act as support structures for cell growth and function, and provide space-filling for future tissue ingrowth [66, 67, 77, 78]. For example, growth factors, such as TGF-β have been tethered to PEG hydrogels to regulate smooth muscle cell function [83] and BMP-2 has been covalently attached to alginate to regulate osteoblast migration and calcification into the gels [84]. Also, differentiated cardiac tissues have been engineered by casting neonatal rat cardiac myocytes into collagen gels and subsequently subjecting them to cyclic mechanical stretch [85]. Another type of hydrogels has been used extensively in tissue engineering approaches [47, 86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101]. The most abundant heteropolysaccharides in the body are the GAGs. They are long unbranched polysaccharides containing a repeating disaccharide unit that contains either of two modified sugars: N-acetylgalactosamine or N-acetylglucosamine and a uronic acid such as glucuronate or iduronate. GAGs are located primarily on the surface of cells or in the ECM. HA is a GAG that is particularly prevalent during wound healing and in joints. Covalently crosslinked HA hydrogels can be formed by multiple chemical modification means [102,103,104,105]. HA can be degraded by cells through the release of enzymes such as hyaluronidase. HA is particularly appealing for tissue engineering as it is naturally present in great abundance in a variety of tissues [106,107,108]. Previously, HA scaffolds have been used for tissue engineering of various tissues [55, 104, 109]. In addition, composite HA-PEG scaffolds have been used for tissue engineering [109,110,111]. Specifically designed amphiphilic peptides that contain a carbon alkyl tail and several other functional peptide regions have been synthesized and shown to form nanofibers through self-assembly by mixing cell suspensions in media with dilute aqueous solutions of the peptide amphiphile. Nanoscaled fibers produced by self-assembly of peptide amphiphile have great potential in tissue engineering [112, 113]. Peptide groups may be customized to direct cell behavior and polymerized directly into the hydrogel. For example, it was shown that directed differentiation of neural stem cells could be modulated using such a hydrogel functionalized with isoleucine–lysine–valine–alanine–valine, a laminin-derived sequence, without the use of additional biochemical factors [114]. Despite their excellent biological properties and the ease with which they can be modified for specific applications they are mechanically weak, which has limited their application in vivo.
Along with chemical properties, physical parameters of fabricated hydrogels such as stiffness, elastic modulus, degradation, and ligand density have a profound impact on stem cell fate decision and phenotype acquisition. Stiffness and matrix composition together have potential to regulate the hepatic differentiation of progenitor cells [115]. In a very recent experiment, cellular reaction of stem cells to matrix stiffness was evaluated using acrylamide and bisacrylamide in a variable stiffness. It was found that stem cells cultured on a soft substrate tended to significantly acquire hepatocyte-like cell phenotype on a soft matrix compared to cells of stiff matrix system. Additionally, soft matrix can maintain the function of stem cells longer than stiff matrix [116]. The soft matrix actively promoted cell migration and alignment. Young’s modulus value and integrin β1 content were decreased by increasing matrix stiffness [117]. These results give deep insights to cell biologists in designing improved matrices with abilities to enhance differentiation efficiency. To our knowledge, there are several mechanistic studies evaluating the effect of microenvironment biophysical properties on hepatic differentiation and functional behavior but more investigations are highly demanded to discover beyond molecular mechanisms.
Hepatic differentiation of various stem cells on decellularized scaffolds
Decellularized liver scaffolds are achieved by isolating of all cellular and immunogenic components of extracellular matrix proteins used commonly in studies related to liver regeneration. The ECM can be obtained through various methods including chemical, mechanical and enzymatic alone or in combination with each other aimed to preserve maximum ECM composition and natural architecture with minimum damage [118, 119]. Evidence points out that the preserved components of ECM differ in number and type depending on the method of decellularization. While in a method the greater amount of components is preserved, however, the substrates required to support hepatic cells or tissue carrying stem cells such as fibronectin, collagen and GAGs may be also excluded [120]. Experiments showed endogenous cytokines and growth factors attached to side chains of GAGS either can be conserved after decellularization [17]. In a very recent study, mass spectrometric analysis revealed that the amount of ECM proteins in a liver decellularized by deoxycholate were significantly higher than those decellularized by ammonium hydroxide, but the retained ECM proteins in the latter mainly were fibronectin, collagen (types 1 and 4) and proteoglycans, participating in cell attachment, survival, growth and hepatic differentiation [120]. Since the type and amount of the material content of scaffold affect the orientation of stem cells toward the hepatic lineage, quantitative and qualitative evaluation of decellularized scaffolds component appears to be essential prior to recellularization [120]. ECM scaffolds in the field of liver tissue regeneration are originated from various sources. For instance, the whole or part of the human liver organ which is not suitable for transplantation or the addition of synthetic or semisynthetic bio-molecules is at the center of attention [121].
Considering the role of common signals in hepatic differentiation across the various species, xenogenic animal livers including porcine, rat and mice also are used [122]. It seems that hepatic cells could per se trigger the stem cell-to-hepatocyte differentiation by providing essential ECM in in vitro condition. Liver-derived cell lines such as HepG2, Sk-Hep-1, liver primary cells, and progenitor cells were shown to release the array of ECM components into the culture media, promoting the hepatic commitment of various stem cells. Although decellularized whole liver maintains as much of native liver architecture, it requires challenging perfusion systems to fulfill decellularization which is not suitable to study a certain area of tissue [123]. Decellularized liver scaffolds not only markedly up-regulate the gene expression of integrin receptors, α1, α2 and notably α3β1 subunits, and related downstream modulators such as FAK and ILK but also yields the induction of HGF, bFGF, oncostatin-M and MAPK genes. Using scaffolds with higher porcine liver ECM content, hepatic differentiation of bone marrow-MSCs is not accelerated in optimum rate and expression of hepatic markers disappeared [124].
Though it is well-established that the ECM composition plays an important role in the determination of cell attachment, differentiation, and proliferation, however, a great body of studies has particular attention on the induction and/or inhibition of signaling pathways by growth factor treatment [125, 126]. Notably, there is little knowledge regarding the role of ECM type, protein content, and protein types of the scaffold in hepatic phenotype acquisition. As a matter of fact, novel strategic approaches investigating these features seem to be highly recommended. The use of plates coated with Matrigel, laminin and different types of collagen with a limit number of adhesion molecules was done previously. It has been shown that these matrices were unable to support the attachment of hiPSC-derived definitive endoderm cells [121]. In contrast, DE cells not only attached more efficiently to plates containing HepaRG acellular matrix and tightly adhered to each other but also committed more efficiently toward hepatic lineage [123, 127]. The effect of matrixes was examined on later stages of hepatic differentiation and shown that matrices mostly induce mature hepatic markers expression when used at maturation step [14]. On the other hand, the use of the human decellularized liver matrix in the early stage of differentiation can further stimulate earlier high expression of stage-specific markers in hepatic differentiation of hiPSCs compared to cells grown on Matrigel and collagen substrates [121]. With the progress of hepatic differentiation, the cells lose their sensitivity to the ECM composition. Therefore, ECM composition has the least influence on hepatic differentiation at the late stages. If the entire process of differentiation takes place on liver decellularized matrix, the large number of cells further display hepatic lineage features and the pattern of markers expression is found in embryonic development of liver and a low expression of HLF and HHEX, as mature liver markers, indicates these cells do not enter to a complete maturation procedure [121]. However, evaluation of ECM effect on the maturation of hepatocytes-derived from iPSCs led to achieve cells closer to intact mature liver cells evidenced by expression of lower levels of fetal liver markers, AFP and CYP3A7, and greater extent expression of Cyp proteins as well as better metabolic activity as compared with the same cells cultured on synthetic 3D scaffolds [128].
Some alternatives have been represented to partially overcome immaturity including pretreatment of cells with divalent cations such as Mn2+, as an integrin activator, remove of HGF, and the addition of AMP and Matrigel in the maturation step of hepatic differentiation [123, 124]. One recent study has shown that liver progenitors seeded onto ECM discs developed from ferret rat decellularized liver can self-assemble into 3D spheroids with a greater capacity for differentiation into hepatocyte and simultaneous generation of bile canaliculi-like structures [122]. The cultured cells also showed the first step of xenobiotic catabolic activity and expression of adult livers markers (HNF4a, CYP3A4, and CK19) [122].
In another study, the hepatic induction of porcine-iPSCs in a mixture of porcine liver ECM solution and hepatocyte differentiation media (10% of ECM in a total volume of media produced mature and functional cells compared to 2D cultures [14]. Following the transplantation of these cells to decellularized rat liver, the cells establish a cell-to-cell connection and efficiently attach to the matrix and exhibited enhanced expression of AFP and Alb, indicating hepatic phenotype maintaining [14]. The differentiated cells showed morphology similar to hepatocytes, abundant cytoplasm and large microvilli with a high ratio of cytoplasm to the nucleus. The transplantation of hepatocyte-like cells restored the liver function and reduced the CCL4-induced liver fibrosis in mice when compared to non-ECM cultures [14]. Despite considerable advances in liver tissue engineering, the use of liver decellularized scaffolds encounters some obstacles [129]. The problematic issues in the liver are that the outcome of decellularization show donor to donor variation [129]. Indeed, using the same chemical decellularization protocol on five livers obtained from patients undergoing resection and three healthy porcine liver yielded five different outcomes for human livers but reproducible for porcine livers [129]. In addition, substantial leakage of cells from the vascular network and parenchymal space takes place to the outside of the scaffold in perfusion-based protocols [130]. Based on the size of scaffolds, appropriate methodologies must use for decellularization. For example, the perfusion of small-size livers is laborious and many tissue masses are required to yield tissue powder. However, a low number of large livers are needed for the preparation of tissue powder while comparable to the human liver [14, 131]. In addition, the vacant spaces of tissue undergo collapse after removal of cells, which in turn makes the tissue condensed and stiff. Obtained stiffness is similar to the consistency of fibrotic tissues such as liver parenchyma. It has been reported that tissue stiffness could be altered from 4.5 kPa in normal tissue to more than 19 kPa in diseased liver tissue. Stem cells differentiated toward hepatic lineage on soft matrices (stiffness = 0.4 kPa) demonstrated a high degree of hepatocyte characteristic within a few hours, whereas stiffer matrices (stiffness > 80 kPa) was unable to support differentiation. Integrins have been suggested play an important role in hepatic induction of stem cells and the matrices with moderate stiffness enhance cell–cell (catenin-based) and cell–matrix (integrin-based) adhesion through maintaining balanced regulation of integrin-β1 and β-catenin expression and exhibit high value of Young’s modulus [117].
The ultimate goal of liver decellularization is to subject bioscaffolds for recellularization which are more applicable to hepatic microstructure. Liver decellularization and recellularization reported by Uygun et al. was the first study in this area. They transplanted acellular scaffolds to hepatic tissue coincided with the administration of 5 × 106 rat primary cells through the portal vein. Evaluations confirmed the successful engraftment exceeding 90% percent. During the 1st days after recellularization, transplanted cells surrounded the large veins and distributed across the whole scaffold. Based on the released data, approximately 20% of transplanted cells exhibited apoptotic changes. Furthermore, the cells continuously expressed the UDP-glucurotransferase 1, polypeptide A1, Alb, and urea. Interestingly, the expression of cytochrome 450 isoenzymes was similar to the pattern observed in normal liver. Also, Robertson and colleagues examined hepatic recellularization by using adult rat and human hepatocytes in a bioreactor culture model. After perfusion of rat liver with rat or human hepatocytes through the portal vein, cellular constructs were developed after 28 days and reticular networks consisted of type III collagen III formation were observed. In addition, these cells were functional and actively showed Alb and urea synthesis for up to 28 days of study [132]. In another study, decellularized mouse liver was recellularized with 5 × 105 iPSCs-derived hepatocytes and transplanted into the mice by a four steps infusion protocol through the portal vein. The transplanted cells successfully engrafted around the portal vein and produced Alb and α-fetoprotein [14]. It was also shown that decellularized hepatic scaffolds were appropriately recellularized in rat partial hepatectomy model [133]. Although results from recellularization protocols seem promising many efforts are essential to achieve functional liver tissue. Calling attention, the various routes of recellularization demonstrated different performance and regenerative outcomes. In a comparative experiment, the efficiency of cell administration was assessed via portal vein and bile duct routes in murine recellularized liver tissue. It was shown that administrated cells developed aggregates that block the microvasculature system and led to uneven distribution of cells throughout the parenchyma when cells were introduced through the portal vein. In contrast, by cell transplantation through bile duct, these cells were uniformly distributed throughout decellularized hepatic parenchyma [134]. Some authorities declared the potency of decellularized non-hepatic bioscaffolds such as placental tissue for the restoration of hepatic function [135]. Despite these technical barriers, considerable efforts are ongoing to generate liver decellularized scaffolds closely simulate in vivo microarchitecture of the liver.
Hepatic differentiation via the combination of cell source and nanotechnology-based approaches
The sophisticated nanofiber designed is touted as alternative methodologies for improving the efficiency of fabricated biomaterials in the favor of target tissue regeneration. The electrospun nanofibers with high surface area and porosity have been extensively used to act as backbone ECM, improving the growth and differentiation of cells. For example, the 3D liver model developed by electrospun chitosan nanofibers and co-culture system was shown to support hepatocytes and fibroblasts for long-term liver functions. The use of different ECM substrates in synthesized nanofibers such as fibronectin enhanced cellular adhesion and polarization [136]. Chitosan is a suitable polymer for hepatocyte culture, expansion and maintenance, because of structural similarity to glycosaminoglycans; the components seen commonly in hepatic ECM [137]. Based on the great body of documents, hepatocytes can maintain their morphologies and functions for long periods of time in the co-cultures 3-D system [136]. By applying various strategies, it is mighty to preserve the functional behavior of the target cells in a controlled manner. For instance, encapsulation of hepatocytes in 3D structures provides an appropriate niche for optimal function. In one study, polyelectrolyte complex hydrogel fibers were used as self-assembling 3D structures to encapsulate hepatocytes and endothelial cells differentiated from human iPSCs. The co-culture of cells in multi-interfacial polyelectrolyte complex fibers with chitin and sodium alginate origin showed that the existence of endothelial cells in the scaffold significantly improved hepatocyte function and in vivo studies revealed the superior effect of endothelial cells in vascularization of the transplanted scaffolds [138].
Natural ECM has a nanoscale structure that is common to many tissues’ basement membranes. To mimic liver ECM, various nanomaterial-based scaffolds have been studied until yet. CNTs showing controlled nanoscale topography that can be employed as liver ECM [139]. In a study, PA was used as a base matrix to resemble the human liver due to its Young’s modulus [140]. In addition to the existence of anchor sites stimulating cell attachment, scaffold consistency and pattern could dictate specific phenotype for target cells. It has been shown that the scaffold stiffness can influence the in vitro behaviors of hepatocytes [141]. Also, PEG-CNTs were coated on the PA to make CNT-PA. The influence of CNT-PA on the differentiation of hAECs to functional HLCs was investigated. CNT-PA had the potency to induce the up-regulation of hepatic markers at transcription and protein levels. Moreover, this milieu was able to yield higher uptake of indocyanine green, Alb secretion and comparable CYP3A4 enzymatic function [19].
Authorities demonstrated broad advantages for tissue engineering governed by CNTs. For example, they are biocompatible materials with great potential as cell-supporting substrates, provide strong mechanical properties, can be easily functionalized and are aligned as the collagen fibers [142]. Calling attention, MWNTs used as a scaffold to investigate the effect of it on primary liver cell culture showed suitable adhesion, proliferation rate with an enhanced function. This microstructure possesses the capacity to force hepatocyte to enhance Alb production coincided with the activity of CYP1A2 enzyme [139].
Hepatocyte growth factor has been found to play an important role in morphogenesis, and liver regeneration. Because of its short half-life in circulation, specific carriers have been studied for improving the release profile [143]. The polymeric delivery systems based on PLA, PLGA, and polycaprolactone and inorganic nanomaterials such as silica nanoparticles have been investigated. Due to specific properties such as biocompatibility, easily functionalized surfaces and tunable pore volumes silica nanoparticles are considered in the field of tissue engineering [144]. The modification of MSNs supplemented with PEI and loaded with Activin A, aFGF and HGF were used for hepatocyte differentiation of mouse embryonic stem cells. Functionalized MSNs were found to deliver growth factors in a sustained manner. Compared to the control non-treated cells, the gene expression of hepatocyte markers, such as ALB and alpha-fetoprotein was upregulated in the GF–PEI–MSN complexes. Also, the expression of hepatic functional markers such as α1-antitrypsin, cytochrome P450 subunit CYP7A1, and glucose-6-phosphatase were higher in GF–PEI–MSN complexes. Monitoring the synthesis of glycosaminoglycans by periodic acid-Schiff showed that the increasing number of positive cells in the GF–PEI–MSN complex group. In the presence of this nanoparticle, mouse embryonic stem cells acquire a potency to give rise to endodermal lineage and hepatocyte-like phenotype after transplantation into the injured liver in the mouse model [145]. In conclusion, nanomaterials possess advantages for tissue regeneration. For example, the nanofibers act as ECM and provide high surface area and porosity that are suitable for cell proliferation and differentiation. CNTs simulate collagen fibers and due to their high mechanical properties provide good stiffness as a scaffold. Also, nanomaterials can be used as carriers for improving release completeness of growth factors that have a role in the differentiation of cells. It suggests that the combination of these nanomaterials can be appropriate for hepatic differentiation.
3D printing role on cell fate toward a hepatic-like phenotype
3D printing has been touted as a novel method to dictate specific cell alignment on distinct substrates. During the last decades, many attempts have been collected to commercialize 3D-printing scaffolds. In this regard, Organovo™, one of the pioneer bioprinting corporations, has effectively attained 3D-vascularized liver constructs with high cell survival rate and applicable microstructures coincided with the formation of hepatic lobules after the use of mixed cell populations notably hepatocytes, endothelial, and hepatic progenitor cells [146, 147]. As a matter of fact, 3D tissue-engineering technology possesses the capacity to effectively simulate complex organ formation in a 3D pattern and circumvent plenty of obstacles reported for the 2D culture system [23]. The main obstacle is a confined hepatocyte proliferation and expansion rate in the 2D system. For instance, the hepatocyte proliferation rate is commonly decreased 3 to 5 days post-isolation and these cells are prone to de-differentiate and acquire fibroblast-like phenotype by modulation of hepatic-associated genes, leading to cell functionality removal and limitation of activity [148, 149]. It has shown that the secretion of Alb, synthesis of urea, and expression of gene CYP3A4 are decreased in freshly isolated hepatocytes [150, 151]. In support of the above-mentioned statement, the fabrication of scaffolds with natural substrates enabling a long-term culture of hepatocytes is the most attractive issues remain to be resolved. Investigations of primary hepatocytes in the 3D niche have been briefly progressed to elucidate hepatic metabolism and uptake [150, 152]. Calling attention, primary hepatocytes have the potential to form individual micro-aggregates in organ culture systems [23]. Culturing in 3D alginate scaffolds was found to promote hepatic cells aggregation 7 days post cultivation. The cells acquired prerequisite function and typical hepatocyte-like morphology to synthesize Alb [24, 151]. Considering the importance of cell aggregation in the function of hepatocytes, this leads to the expansion of a diversity of techniques to encourage and dictate the formation and maintenance of aggregates and micro-spheroids within an engineering scaffold prior to implantation to the target sites in vivo [153, 154]. In line with this claim, encapsulation or loading single-cell suspensions onto a porous scaffold are famous techniques to induce cell aggregation [155, 156]. However, some considerations in scaffold design are highly recommended for appropriate results. For example, the control of aggregate size to avoid generation of necrotic cells located at the core or applying some approaches to induce reciprocal interconnectivity between cells from neighboring aggregates while facilitating nutrient diffusion and neo-vascularization following implantation [157]. Notably, some inherent modulations could not be omitted during the fabrication of synthetic scaffolds. In scaffold production, restriction and bottlenecks such as freeze casting, electrospinning, salt leaching, or gas foaming could change the size and cell microspheres integrity within 3D backbones [158]. In these circumstances, only a limited degree of control is possible over scaffold geometry, pore size, and interconnectivity.
Hexagonal lobule-like geometries have been displayed to have advantageous effects on cultured hepatocytes [118, 159]. Use of an engineered technique to re-form detailed lobule structure must be considered as lobule organization in vivo is recognized to have a biological effect on blood and bile flow as well as hepatocyte phenotype and zonality [160]. The influence of 3D-printed scaffold pore geometry has therefore not been thoroughly examined for applications in soft tissue engineering. This may partially be due to the complications in 3D printing of soft resources, mainly hydrogels, making laborious attempts. Recently Sainz and co-workers studied the effect of gelatin-based scaffold pore geometry on non-adherent cells while keeping pore size constant and compared the result with a control 2D surface. They revealed that 3D manipulation of quite simple material, such as gelatin, can have major influences on the application of engineered tissues [161]. Compared to the 2D system, an appropriate phenotype acquisition and functional improvements are seen in 3D printing systems. In 3D hydrogel-based culture model, the co-culture of hiPSCs and hematopoietic progenitor cells with endothelial lineage and adipose-derived stem cells contributed to microscale size hexagonal construction while the biochemical activity of hepatocyte was initiated by advanced liver-specific gene expression levels, increased cytochrome P450 stimulation and activated secretome [118].
Bioinks
Hydrogel bioinks used for cell-based systems presented biochemical and biophysical motivations to mimic native ECM in in vivo microenvironment. Bioinks with the ability to allow appropriate cellular activities have also been improved for 3D bioprinting. In this technique, interfacing between the substrates and fabrication hardware, physicochemical features of biomaterials and bioink cocktails are one of the most obstacles attendant with 3D bio-fabrication methods [162]. Different studies have used a diversity of hydrogels such as HA, MeHA, PEG, GelMA, NovoGel® 2.0 (a non-adherent and thermo-responsive hydrogel), collagen, fibrin, fibrinogen, PCL, gelatin, and alginate in different combinations, for 3D bioprinting of hepatocytes and liver associated cell lines [118, 162,163,164,165]. Based on previously published experiments, different bioink formulations were used for the rapid production of hepatic tissue. Two forms of cell-loaded scaffolds including hepatic cells, gelatin and/or fibrinogen were effectively synthesized by programmed rapid prototyping method and became stable with thrombin. During the procedure, no obvious cell damage was observed. It was reported that an equal gelatin/fibrin combination provided the highest mechanical properties. Hepatic cells were demonstrated to efficiently proliferate in the matrix of gelatin/fibrinogen. The Alb biosynthesis was found to improve in embedded hepatocytes. It seems that fibrin performs as a satisfactory material for a gelatin-based cell assembly matrix with easy manipulation, bio-resorbable, supporting in vitro cell functions [166]. Another group combined ADSCs within a gelatin/alginate/fibrinogen hydrogel to construct a vascular-like network, using a digital pattern. They also located a cocktail of hepatocytes with gelatin/alginate/chitosan around it to simulate a natural liver tissue. In these systems, endothelial growth factors were used to induce the ADSCs to trans-differentiate into endothelial-like cells. In these bulk constructions, a 3D syringe-based bioprinting technique is demanded to provide bulk interchange channels using to preserve the integrity of the constructed liver structure. This double-nozzle assembling techniques have the potential to be a powerful apparatus for producing multipart constructs with distinctive intrinsic/extrinsic properties [167].
3D fabrication procedure
Recently, several 3D bioprinting technologies have been exploited with effective advances in shaping biomaterials at the macroscale size to design biomaterials with high complexity. These technologies can be divided into different categories that include stereolithography, laser-assisted forward transfer, nozzle-based bioprinting techniques, shear-thinning extrusion bioprinting, sacrificial bioprinting, microfluidic bioprinting, and multi-material bioprinting [168, 169]. Depending on the type of bioinks, some of these methods are applicable for liver tissue engineering.
Shear-thinning extrusion bioprinting
Extrusion bioprinting is a promising method to produce organized tissue constructs that supply cells and matrix materials concurrently to restore or substitute damaged or diseased tissues and organs. This procedure has an interesting vision to fabricate constructs with 3-D distinctions of cells through numerous axes with great architectural complexity. Extrusion bioprinting naturally needs the distinct bioink formula to be self-supportive and preserve the architectural integrity of the scaffold upon extrusion [170]. Lee et al. produced a GelMA-based shear-thinning bioink that allowed direct extrusion, by rapidly cooling down GelMA solutions to beneath their gelation points in the printing nozzle [170]. UV-initiated crosslinking could then be used to subsequent solidification [170, 171]. The bio-printed products could maintain the viability of encapsulated hepatocellular carcinoma cells; HepG2 and mesenchymal stem cells (MSCs) within the GelMA microfibers over 14 days [170].
Multi-material bioprinting
Printing a single bioink has been the base of most researches in 3D-bioprinting customarily. Nevertheless, because of the complexity of organs and tissues, combining various construction biomaterials and numerous cell types is essential in a single bioprinting procedure. Several studies have been done to explore the intricate associations between the printing factors and the resultant resolution. Tissue constructs made of numerous materials in a well-ordered fashion could be gained by serially altering the input bioinks for the digital micro-mirror device bioprinter [118, 172]. For instance, to simulate the liver microstructure, Ma et al. used the sequential printing of human hiPSC-derived hepatic cells in a hexagonal lobule structure with supporting cells from the endothelial and mesenchymal original, filling the lining of the lobules [118]. The 3D printed liver construct proved structural similarity with it’s in vivo counterpart and showed an appropriate functionality evident by prolonged cell viability, cytochromes activity, and stable secretion of liver biomarkers.
DCW system
DCW is used to fabricate simple linear or complex conformal constructions on a substrate by material deposition. Digital writing or digital printing is a group of malleable multifaceted procedures in which different methods and devices such as inkjet, laser, mechanical pressure, and tips are used to create structures in the range of nm to the mm. Also, an extremely wide range of materials from ceramics, metals, polymers, and dielectrics to bio-materials is used in these techniques [173]. In new technical systems, solid freeform fabrication, cell-based micro-chip systems, sensors, and microfluidic devices, cells are combining as part of the construction blocks for several tissue engineering procedures. Multilayer 3D printing techniques combined with perfusion culture systems are capable of producing in vivo liver tissue. To produce a biomimetic liver micro-organ as a drug screening approach, a DCW method in combination with perfusion culture has been established [174]. To align alginate hydrogels encapsulating HepG2 cells, the DCW system with four nozzles capable of operating in extrusion or droplet model was utilized. Three-layer tissue-engineered scaffolds were created by printing these alginate-encapsulated cells into liver sinusoidal shape structure that combined into a microchip device, allowing for medium exchange. Outcomes presented that over 80% of HepG2 cells in the cross-linked, printed products stayed alive after 3 days. Additionally, viability was preserved after 24-h perfusion circulation, indicating that the perfusion method did not per se influence cell viability and could be utilized to perfuse a targeted drug through fabricates and measure pharmacokinetic performance. Another investigation successfully utilized the DCW process and incorporated both hepatic and epithelial cells in the Matrigel substrate to more precisely stimulated hepatic sinusoid hierarchical architecture. A microfluidic environment was designed by these cells to make enhanced higher reliability in the favor of micro-liver tissue stimulation. This application is useful to serve as a portable ground model for the study of drug conversion and radiation protection of living liver tissue analogs. It seems that understanding the underlying mechanisms of the multi-cellular biological system responses for a prolonged period, various disease models and biosensors need to be elucidated [175].
Stereolithography
Stereolithography is one of the first 3D printing methods in which a forerunner mixture of a UV-curable material is filled in a tank, which is then cross-linked in an aligned manner. After the depositing of each distinct layer, a mechanized stage moves the deposited construction from the light source in the z-direction so there will be space for the subsequent print layer [169]. Scaffolds that more closely simulate liver construction and encourage improved hepatocyte culture have also been considered yet. PEG-based, photo-polymerizable polyethylene glycol-based hydrogels have been produced using stereolithography to develop hepatocyte cell seeding, proliferation, and duration of perfusion cultures in liver designs. Hydrogel scaffolds with open channels were fabricated for post-seeding and perfused culture of primary hepatocytes that form 3D structures in a bioreactor. Cell seeding densities, flow rates photo-initiator concentrations, stereolithography energy dose and pretreatment conditions were determined to be important factors to control and maximize cell viability. The perfused culture of primary hepatocytes in hydrogel constructs in the existence of soluble epidermal growth factor increased the preservation of Alb creation throughout the 7-day culture relative to 2D controls. This method can be hired to construct soft scaffolds for a number of bioreactor configurations [176].
Other physical factors important in the efficiency of bioink
Different studies have shown that cell viability is affected by several factors including the material flow rate, material concentration, dispensing pressure, and nozzle geometry. These results can attend as an instruction for future explorations and optimization of the 3D system. Sun and his co-workers made a multi-nozzle bio-printing system to concurrently deposit cells and multiple biomaterials. Their rheology study and cell viability assay were executed to examine mechanical-stress-induced cell damage during the printing procedure [177, 178]. Both Burdick and Wells groups examined how the differentiation of HSCs was affected by matrix stiffness. Tissue fibrosis that might be in part attributed to ECM stiffening during disease improvement has been the cause of differentiation of HSCs into myofibroblasts. Their outcomes established that HSCs cultured on stiffer hydrogels (24 kPa in elasticity) as compared to HSCs on softer hydrogels (2 kPa), expressed advanced levels of alpha-smooth muscle actin and type I collagen [179,180,181]. Shear stress applied during the printing procedure is another factor that can affect cell viability. Using liver spheroids can protect the cells from the adverse influences applied by the shear stress during the printing process and recapitulate the cell-to-cell interactions. By their stable secretion of hepatic biomarkers including Alb, ceruloplasmin, alpha-1 antitrypsin, and transferrin, the bio-printed liver spheroids surrounded by GelMA hydrogel showed long-term functionality for up to 30 days [163]. In another experiment, it was revealed that UV exposure can affect cell viability in UV-initiated crosslinking systems which are used to subsequent solidification of self-standing multilayered constructions and thereby advanced UV exposure could improve the crosslink density and thus causes in decreased cell viability [170].
Hepatic phenotype acquisition via an intracellular signaling pathway
Small molecule
To induce hepatocyte-differentiation of stem cells, different methodologies have been developed yet. In most circumstances, exploiting step-wise differentiation approaches along with recombinant growth factors or small-molecule analogs could be promising [182]. Early protocols for the generation of the specific cell type from stem cells was applicable by using embryoid body formation [183]. The basis of this protocol was the spontaneous formation of cell aggregates (embryoid bodies) from pluripotent cells which has the capacity to give rise to all three germ layers [16]. To expand the hepatocytes, the most well-organized method is a monolayer culture of distinct cells in a step-wise manner exposure to specific cocktails of recombinant growth factors and cytokines, including Activin A, Wnt3a, HGF, OSM, FGF4, VEGF, EGF and BMP4 [184,185,186,187,188,189,190,191]. These approaches were designed to simulate the embryonic stages of hepatocyte lineage. The protocol has, however, some deficiencies with low differentiation efficacy and heterogeneity in the nature of differentiating cells [182].
Small molecules can regulate specific target(s) in signaling pathways and epigenetic mechanisms have been recognized as valid chemical tools for mediating cell fate. The use of small-molecule compounds has the advantage of being safer than the use of cytokines, nucleic acids, or protein therapies [192]. Other advantages of small-molecule-based hepatic differentiation protocol in comparison with other methods are simplicity, stability, reproducibility, and low cost [193]. Thus, the identification of small molecule compounds has enhanced the development of stem cells to liver regeneration. In the following, we will discuss the role of small molecules differentiation of stem cells toward liver cells in various studies. During hepatic differentiation of human MSCs, Wnt/b-catenin signaling is inhibited. Inhibition of Wnt/b-catenin signaling molecules or target genes induces the hepatic differentiation of human MSCs. Effective modulation of Wnt signaling and glycogen synthase kinase 3 inhibition using small molecules efficiently produced definitive endodermal lineage. Dickkopf Wnt signaling pathway inhibitor 1, an antagonistic inhibitor of the Wnt signaling pathway was shown to promote hepatic differentiation of bone marrow-derived MSCs [33, 194]. More recently, different research groups showed that glycogen synthase kinase 3 inhibition with CHIR99021 efficiently directs hPSCs toward the definitive endodermal lineage and primary hepatocytes, via a combination of small-molecule treatments with dimethyl sulfoxide. They stated that the small-molecule based approach is a simple, inexpensive and reproducible platform for the derivation of hepatocytes from hPSCs [182, 195]. Itaba et al. screened 23 newly synthesized derivatives of small molecule combinations produced from several known Wnt/β-catenin signal inhibitors. In their study, HC-1, IC-2, and PN-3-13 similarly suppressed Wnt/b-catenin signaling, however, IC-2 was the most effective inducer of hepatic differentiation. Interacted molecules with IC-2 were not determined in their studies. IC-2 is a derivative of ICG-001. It has been made clear that ICG-001 suppresses Wnt/b-catenin signals by binding CREB binding protein, and because IC-2 is derived from the ICG-001, it may act with the same mechanism as the ICG-001 [196].
Stauprimide a small molecule described by Shoutian Zhu et al. that can significantly increase the efficiency of directed ESC differentiation in conjunction with lineage specification cues. Using an affinity-based approach, NME2 was reported as the biological objective of stauprimide [197]. Inhibition of NME2 occurs when stauprimide binding to NME2, NME2 nuclear localization, which, in turn, suppresses c-Myc expression. Downregulation of c-Myc expression, a key factor in the preservation of ESC self-renewal and its inhibition facilitates the process of differentiation [198].
Tasnim and co-workers stated a protocol for efficient differentiation of hESCs into hepatocyte-like lineage using a mainly small molecule-based approach. These three stepwise differentiation strategies contain the use of optimized concentrations of LY294002 and bromo-indirubin-3′-oxime) for the generation of definitive endoderm; sodium butyrate and dimethyl sulfoxide for the generation of hepatoblasts and SB431542 for differentiation into hepatocyte-like cells. They also showed that SM-Hep were morphologically and functionally similar or applicable to the hepatocytes generated by growth factor cocktails (GF-Hep) in terms of expression of hepatic markers, urea and Alb production and cytochrome P450 (CYP1A2 and CYP3A4) activities [33]. Ouyang et al. introduced 2-(4-bromophenyl)-N-(4-fluorophenyl)-3-propyl-3H-imidazo [4,5-b] pyridin-5-amine (SJA710-6), as a small molecule able to selectively promote MSCs differentiation toward hepatocyte-like cells after monitoring a library of 2500 small molecules [199]. Also, small molecules of RNA called small hairpin (shRNA/Hairpin Vector) can be used as another small molecule to increase the regenerative capacity of hepatocytes. By small hairpin RNAs (shRNAs) directly in the animal model, Wuestefeld et al. recognized the MKK4 which is dual-specific kinase as a key mediator of liver regeneration. MKK4 inhibition expressively amplified the regenerative ability of hepatocytes in animal models of liver regeneration. Notably, MKK4 inhibition in hepatocytes decreased fibrosis after chronic liver injury. Hepatocytes with stable RNA interference (RNAi)—mediated MKK4 silencing indicated quicker cell-cycle entrance and accelerated liver regeneration [200].
In summary, using small molecules instead of growth factors would provide a striking alternative source since small molecules are cell-permeable and non-expensive compared to growth factors [201]. Due to several advantages including the ability for sequential, tunable and sectional control of specific protein function, elimination of the risks and drawbacks associated with genetic manipulation, the low efficiency and slow kinetics of distinct phenotype induction, the application of small molecules in the biomedical field could simplify therapeutic benefits related to stem cells [201, 202].
miRNAs
As above-mentioned many studies have tried to discover cheap and accessible techniques for producing definite liver cells or hepatocyte-like cells from different stem cell types. In order to achieve this end, several hepatic differentiation media with different growth factors and cytokines have been introduced to hinder hADSCs-differentiation into hepatocytes. In a recent experiment, the significance of specific miRNAs has been shown on the differentiation and growth of certain cells. microRNAs are noncoding RNAs that control gene activities at the post-transcriptional stage, regulating stem cell biological features in plants and animals [203]. Further, 2500 unique mature human miRNAs have been recognized until now and it is probable that more than one-third of human protein-coding genes are exposed to regulation by miRNAs during the development of hepatic tissue. For example, the essential role of miR-30a has been recognized for bile duct development in the model of zebrafish [204]. The most abundant miRNA in the liver, miR-122, accounts for approximately 70% of total miRNAs [205], which is essential for the appropriate progression of hepatocyte differentiation [205, 206]. In recent years, evidence showed miRNAs performance in stem cell maintenance, differentiation, and organ development [206]. Data suggest miRNAs can be important regulators of hepatocytic differentiation. Additionally, studies showed that miR-194 convinces intestinal epithelial cell differentiation as well [207] is a key moderator of chondrogenic differentiation of ADMSCs [208]. More recently, Jung et al. [27] showed that the amplification of miR-194 progressively increased in HepaRG liver progenitor cells and hESCs differentiation into hepatocytes. In a better word, miR-194 overexpression results in down-regulation of 16 genes in both HepaRG cells and hESCs by modulation of IGF1R and YAP1, a downstream mediator of Hippo signaling that plays a key role in cell fate determination [209]. It was elucidated that Let-7f miRNA acts as a negative regulator for hepatic differentiation in hADSCs through the suppression of HNF4a and silencing of Let-7f, promoting hepatic differentiation [210]. It seems that simultaneous modulation of different miRNAs, miR-122, miR-148a, miR-424, miR-542-5p and miR-1246, could stimulate hMSCs-HLCs transition. The HLCs originated from five miRNAs expressed hepatocyte-specific genes HNF4A, AFP, ALB, TF, CYP3A4, and G6P [211]. Möbus et al. found that miR-199a-5p repression increased hepatic differentiation led to the generation of hepatic cells. Further, they showed that these hepatic cells have the ability of engrafting and regeneration the mouse liver [28]. They recognized the ATPase subunit of the mammalian SWI/SNF complex, SMARCA4, as a target of miR-199a-5p. This complex consists of 15 subunits and participates in is chromatin-remodeling, used in iPS reprogramming and chromosomal stability [212]. These studies show that one or several definite microRNAs can be used to transform stem-cells derived from sources into hepatocytes to proficiently obtain hepatocytes in vitro.
Genetic manipulation in dictating hepatic-like features
The population of stem cells in the human body possesses a highly different developmental capacity. The process from pluripotent to multipotent and developmentally statuses is attended by total alterations in gene expression. Genes active in previous progenitors are progressively silenced at developmentally later stages, and subsets of cell-type-specific genes are turned off. This development is the result of the careful selective expression of transcription factors together. Hepatocyte differentiation is recognized to be coordinately controlled by the act of multiple hepatocyte nuclear transcription factors, for example, hepatocyte nuclear factor (HNF)-1α and -1β; HNF-3α, -β, and -γ; HNF-4α, HNF-6; and CCAAT/enhancer-binding protein α and β [213]. A liver-enriched transcription factor, HNF-3β, prompts the primary development of the endoderm, the predecessor to produce the liver. HNF-3β in the developing endoderm is essential for the expression of HNF-4α, which shows a potential action in hepatocyte differentiation [213, 214]. Recently, HNF-3β Overexpression has been presented to prompt endoderm differentiation of ES cells. While HNF-3β-transfected ES cells were cultured in hepatocyte-culture medium with FGF-2 in three-dimensional culture, ES cells differentiated into HTC cells with liver-specific metabolic roles [214]. Parp1, (Poly (ADP-ribose polymerase 1)), is an enzyme that catalyzes PARylation, a crucial effector intricate in DNA repair, replication, transcription, and genomic methylation. DNA damage and decreased proliferative responses to mitogens have been detected in hepatocytes from Parp1-deficient mice. Breakdown of Parp1 signaling can exacerbate diet-induced obesity and insulin insensitivity. These findings suggested that Parp1 is an essential element in hepatic protection [215, 216]. Huang et al. demonstrated that Parp1 can improve iPSC generation from somatic cells in the existence of three other reprogramming factors Oct4/Sox2/Klf4 and after stimulation of hepatic differentiation into hepatocyte-like cells. Oct4/Sox2/Klf4-iPSC-Heps expressed liver-specific markers and characteristics, exhibited mature hepatocyte roles [213].
Conclusion
Alcohol abuse and infection with different viruses, the occurrence of metabolic disorders, carcinoma and injuries are common causes of liver failure. The only curative treatment suggested for liver disease is transplantation that remains expressively restricted by a severe deficiency of organ donors, forcing researchers to use the regenerative medicine modalities to address this issue. Calling attention, producing decellularized scaffolds from the liver organ, 3D bio-printing system and Nano-based 3D scaffolds that simulate the native liver microenvironment seems logical for liver regeneration and to obtain functional hepatic microtissues/organoids. Applying some strategies such as genetic modulation that target small molecules and micro-RNAs could yield an efficient hepatic differentiation rate. Commensurate with these comments, several crucial challenges need to be fully addressed prior to application to the clinical field. For instance, reproducibility in the fabrication of native liver tissue microstructures over microfabrication techniques or self-organization of hepatic organoids remains challenging. Given the complications of cell variations inside the hepatic tissue, appropriate cell density and organization can barely be applied to in vivo milieu. Both juxtacrine and paracrine interactions of non-hepatic cells with hepatocyte regardless 3D bioprinting and decellularized liver scaffolds allow for the reconstruction of ECM at a higher degree. Notably, microtissues/organoids engraftment or whole bioengineered liver transplantation with continuous functionality post-transplantation is challenging. More studies should be made to achieve efficient methods for clinical uses of hepatocytes-derived from stem-cells.
Availability of data and materials
Data will be available on request.
Abbreviations
- ADSCs:
-
adipose-derived stromal cells
- Alb:
-
albumin
- BMP:
-
bone morphogenetic protein
- BMP4:
-
bone morphogenic protein 4
- CNTs:
-
carbon nanotubes
- DE:
-
definitive endoderm
- DW:
-
direct cell writing
- ESCs:
-
embryonic stem cells
- EGF:
-
epidermal growth factor
- ECM:
-
extracellular matrix
- FGF:
-
fibroblast growth factor
- FAK:
-
focal adhesion kinase
- GelMA:
-
gelatin methacrylate
- GAGs:
-
glycosaminoglycans
- HSCs:
-
hepatic stellate cells
- HGF:
-
hepatocyte growth factor
- HNF4α:
-
hepatocyte nuclear factor 4 alpha
- HLCs:
-
hepatocyte-like cells
- hAECs:
-
human amniotic epithelial cells
- hiPSCs:
-
human induced pluripotent cells
- hPSCs:
-
human pluripotent stem cells
- HA:
-
hyaluronic acid
- PEI:
-
hyperbranched polyethyleneimine
- iPSCs:
-
induced pluripotent stem cells
- ILK:
-
integrin-linked kinase
- MSCs:
-
mesenchymal stem cells
- MSNs:
-
mesoporous silica nanoparticle
- MeHA:
-
methacrylated hyaluronic acid
- MAPK:
-
mitogen-activated protein kinase
- MKK4:
-
mitogen-activated protein kinase kinase 4
- MWNTs:
-
multi-walled-carbon nanotubes
- NPCs:
-
non-parenchymal cells
- NME2:
-
nucleoside diphosphate kinase B
- OSM:
-
oncostatin-M
- PCs:
-
parenchymal cells
- PLGA:
-
poly lactide-co-glycolide
- PA:
-
polyacrylamide hydrogel
- PEG:
-
polyethylene glycol
- PEG-CNTs:
-
polyethylene glycol-linked multi-walled carbon nanotubes
- PLA:
-
polylactic acid
- RA:
-
retinoic acid
- 3D:
-
three-dimensional
- TGFβ:
-
transforming growth factor β
- 2D:
-
two-dimensional
- VEGF:
-
vascular endothelial growth factor
- Wnt:
-
wingless-int
References
Nichols W, et al. Hepatotoxicity observed in clinical trials of aplaviroc (APL, 873140). Special Oral Communication. 2005.
Lal AA, Murthy PB, Pillai KS. Screening of hepatoprotective effect of a herbal mixture against CCl4 induced hepatotoxicity in Swiss albino mice. J Environ Biol. 2007;28(2):201–7.
Kobori L, et al. Drug-induced liver graft toxicity caused by cytochrome P450 poor metabolism. Br J Clin Pharmacol. 2007;65(3):428–36.
Yanagitani N, et al. Phase I study of biweekly paclitaxel and carboplatin for frail patients with advanced non-small cell lung cancer. Am J Clin Oncol. 2007;30(5):487–91.
Tomizawa Y, et al. A phase I dose escalation study of biweekly gemcitabine and carboplatin in completely resected stage IB-IIIA nonsmall cell lung cancer. Am J Clin Oncol. 2007;30(5):498–502.
DeLeve LD. Hepatic microvasculature in liver injury. Semin Liver Dis. 2007;27(4):390–400.
Khetani SR, Bhatia SN. Microscale culture of human liver cells for drug development. Nat Biotechnol. 2007;26(1):120–6.
Underhill GH, Bhatia SN. High-throughput analysis of signals regulating stem cell fate and function. Curr Opin Chem Biol. 2007;11(4):357–66.
Nakamura K, et al. Large-scale multiplex absolute protein quantification of drug-metabolizing enzymes and transporters in human intestine, liver, and kidney microsomes by SWATH-MS: comparison with MRM/SRM and HR-MRM/PRM. Proteomics. 2016;16(15–16):2106–17.
Goldberg D, et al. Changes in the prevalence of hepatitis C virus infection, nonalcoholic steatohepatitis, and alcoholic liver disease among patients with cirrhosis or liver failure on the waitlist for liver transplantation. Gastroenterology. 2017;152(5):1090–1099.e1.
Bangaru S, et al. Survey of liver transplantation practices for severe acute alcoholic hepatitis. Liver Transplant. 2018;24(10):1357–62.
Goradel NH, Darabi M, Shamsasenjan K, Ejtehadifar M, Zahedi S (2015) Methods of liver stem cell therapy in rodents as models of human liver regeneration in hepatic failure. Adv Pharm Bull 5(3):293–298
Matsui A, et al. Prolonged engraftment of transplanted hepatocytes in the liver by transient pro-survival factor supplementation using ex vivo mRNA transfection. J Control Release. 2018;285:1–11.
Park K-M, et al. Decellularized liver extracellular matrix as promising tools for transplantable bioengineered liver promotes hepatic lineage commitments of induced pluripotent stem cells. Tissue Eng Part A. 2016;22(5–6):449–60.
Guye P, et al. Genetically engineering self-organization of human pluripotent stem cells into a liver bud-like tissue using Gata6. Nat Commun. 2016;7:10243.
Siller R, et al. Small-molecule-driven hepatocyte differentiation of human pluripotent stem cells. Stem Cell Rep. 2015;4(5):939–52.
Verstegen MM, et al. Decellularization of whole human liver grafts using controlled perfusion for transplantable organ bioscaffolds. Stem Cells Dev. 2017;26(18):1304–15.
Prodanov L, et al. Long-term maintenance of a microfluidic 3D human liver sinusoid. Biotechnol Bioeng. 2016;113(1):241–6.
Zhao C, et al. Enhanced hepatic differentiation of human amniotic epithelial cells on polyethylene glycol-linked multiwalled carbon nanotube-coated hydrogels. J Tissue Eng Regen Med. 2018;12(7):1556–66.
Jeon H, et al. Generation of multilayered 3D structures of HepG2 cells using a bio-printing technique. Gut Liver. 2017;11(1):121.
Shimizu TS, et al. Molecular model of a lattice of signalling proteins involved in bacterial chemotaxis. Nat Cell Biol. 2000;2(11):792.
Symes MD, et al. Integrated 3D-printed reactionware for chemical synthesis and analysis. Nat Chem. 2012;4(5):349.
Kim Y, et al. Three-dimensional (3D) printing of mouse primary hepatocytes to generate 3D hepatic structure. Ann Surg Treat Res. 2017;92(2):67–72.
Lewis PL, Green RM, Shah RNJA. 3D-printed gelatin scaffolds of differing pore geometry modulate hepatocyte function and gene expression. Acta Biomater. 2018;69:63–70.
Gariboldi MI, Best SM. Effect of ceramic scaffold architectural parameters on biological response. Front Bioeng Biotechnol. 2015;3:151.
Li W, et al. Chemical approaches to stem cell biology and therapeutics. Cell Stem Cell. 2013;13(3):270–83.
Jung KH, et al. MicroRNA regulates hepatocytic differentiation of progenitor cells by targeting YAP1. Stem Cells. 2016;34(5):1284–96.
Möbus S, et al. MicroRNA-199a-5p inhibition enhances the liver repopulation ability of human embryonic stem cell-derived hepatic cells. J Hepatol. 2015;62(1):101–10.
Yahoo N, et al. Forced expression of Hnf1b/Foxa3 promotes hepatic fate of embryonic stem cells. Biochem Biophys Res Commun. 2016;474(1):199–205.
Klein D. iPSCs-based generation of vascular cells: reprogramming approaches and applications. Cell Mol Life Sci. 2018;75(8):1411–33.
Godoy P, et al. Recent advances in 2D and 3D in vitro systems using primary hepatocytes, alternative hepatocyte sources and non-parenchymal liver cells and their use in investigating mechanisms of hepatotoxicity, cell signaling and ADME. Arch Toxicol. 2013;87(8):1315–530.
Yu F, et al. A perfusion incubator liver chip for 3D cell culture with application on chronic hepatotoxicity testing. Sci Rep. 2017;7(1):14528.
Tasnim F, et al. Cost-effective differentiation of hepatocyte-like cells from human pluripotent stem cells using small molecules. Biomaterials. 2015;70:115–25.
Rashidi H, et al. 3D human liver tissue from pluripotent stem cells displays stable phenotype in vitro and supports compromised liver function in vivo. Arch Toxicol. 2018;92(10):3117–29.
Azandeh S, Gharravi AM, Orazizadeh M, Khodadi A, Tabar MH (2016) Improvement of mesenchymal stem cell differentiation into the endoderm lineage by four step sequential method in biocompatible biomaterial. BioImpacts 6(1):9–13
Spitzhorn L-S, et al. Transplanted human pluripotent stem cell-derived mesenchymal stem cells support liver regeneration in Gunn rats. Stem Cells Dev. 2018;27(24):1702–14.
Tegge AN, et al. Transcriptomic analysis of hepatic cells in multicellular organotypic liver models. Sci Rep. 2018;8(1):11306.
Bale SS, et al. Isolation and co-culture of rat parenchymal and non-parenchymal liver cells to evaluate cellular interactions and response. Sci Rep. 2016;6:25329.
Dash A, et al. Liver tissue engineering in the evaluation of drug safety. Expert Opin Drug Metab Toxicol. 2009;5(10):1159–74.
Cohen M, Levy G, Nahmias Y. Coculture and long-term maintenance of hepatocytes. In: Protocols in in vitro hepatocyte research. Springer; 2015. p. 161–73.
Larkin AL, et al. Designing a multicellular organotypic 3D liver model with a detachable, nanoscale polymeric Space of Disse. Tissue Eng Part C Methods. 2013;19(11):875–84.
Pettinato G, et al. Generation of fully functional hepatocyte-like organoids from human induced pluripotent stem cells mixed with endothelial cells. Sci Rep. 2019;9(1):8920.
Sharifzadeh G, Hosseinkhani H. Biomolecule-responsive hydrogels in medicine. Adv Healthc Mater. 2017;6(24):1700801.
Langer R, Vacanti JP. Tissue engineering. Science. 1993;260(5110):920–6.
Saberianpour S, et al. Tissue engineering strategies for the induction of angiogenesis using biomaterials. J Biol Eng. 2018;12(1):36.
Toosi S, et al. Long bone mesenchymal stem cells (Lb-MSCs): clinically reliable cells for osteo-diseases. Cell Tissue Banking. 2017;18(4):489–500.
Farokhi M, et al. Importance of dual delivery systems for bone tissue engineering. J Control Release. 2016;225:152–69.
Toosi S, et al. PGA-incorporated collagen: toward a biodegradable composite scaffold for bone-tissue engineering. J Biomed Mater Res Part A. 2016;104(8):2020–8.
Mottaghitalab F, et al. Silk as a potential candidate for bone tissue engineering. J Control Release. 2015;215:112–28.
Jahani H, et al. Controlled surface morphology and hydrophilicity of polycaprolactone toward selective differentiation of mesenchymal stem cells to neural like cells. J Biomed Mater Res Part A. 2015;103(5):1875–81.
Ghodsizadeh A, et al. Galactosylated collagen matrix enhanced in vitro maturation of human embryonic stem cell-derived hepatocyte-like cells. Biotechnol Lett. 2014;36(5):1095–106.
Kottamasu P, Herman I. Engineering a microcirculation for perfusion control of ex vivo-assembled organ systems: challenges and opportunities. J Tissue Eng. 2018;9:2041731418772949.
Khademhosseini A, et al. Microscale technologies for tissue engineering and biology. Proc Natl Acad Sci USA. 2006;103(8):2480–7.
Li RK, et al. Construction of a bioengineered cardiac graft. J Thorac Cardiovasc Surg. 2000;119(2):368–75.
Masters KS, et al. Crosslinked hyaluronan scaffolds as a biologically active carrier for valvular interstitial cells. Biomaterials. 2005;26(15):2517–25.
Gerecht-Nir S, et al. Biophysical regulation during cardiac development and application to tissue engineering. Int J Dev Biol. 2006;50(2–3):233–43.
Radisic M, et al. Oxygen gradients correlate with cell density and cell viability in engineered cardiac tissue. Biotechnol Bioeng. 2006;93(2):332–43.
Radisic M, et al. High-density seeding of myocyte cells for cardiac tissue engineering. Biotechnol Bioeng. 2003;82(4):403–14.
Radisic M, et al. Mathematical model of oxygen distribution in engineered cardiac tissue with parallel channel array perfused with culture medium containing oxygen carriers. Am J Physiol Heart Circ Physiol. 2005;288(3):H1278–89.
Radisic M, et al. Functional assembly of engineered myocardium by electrical stimulation of cardiac myocytes cultured on scaffolds. Proc Natl Acad Sci USA. 2004;101(52):18129–34.
Borenstein JT, et al. Microfabrication technology for vascularized tissue engineering. Biomed Microdevice. 2002;4(3):167–75.
King K, et al. Biodegradable microfluidics. Adv Mater. 2004;16(22):2007.
Kaihara S, et al. Silicon micromachining to tissue engineer branched vascular channels for liver fabrication. Tissue Eng. 2000;6(2):105–17.
Fidkowski C, et al. Endothelialized microvasculature based on a biodegradable elastomer. Tissue Eng. 2005;11(1–2):302–9.
Vozzi G, et al. Fabrication of PLGA scaffolds using soft lithography and microsyringe deposition. Biomaterials. 2003;24(14):2533–40.
Lee KY, Mooney DJ. Hydrogels for tissue engineering. Chem Rev. 2001;101(7):1869–79.
Rowley JA, Madlambayan G, Mooney DJ. Alginate hydrogels as synthetic extracellular matrix materials. Biomaterials. 1999;20(1):45–53.
Burdick JA, et al. An initial investigation of photocurable three-dimensional lactic acid based scaffolds in a critical-sized cranial defect. Biomaterials. 2003;24(9):1613–20.
Burdick JA, et al. Delivery of osteoinductive growth factors from degradable PEG hydrogels influences osteoblast differentiation and mineralization. J Control Release. 2002;83(1):53–63.
Burdick JA, Anseth KS. Photoencapsulation of osteoblasts in injectable RGD-modified PEG hydrogels for bone tissue engineering. Biomaterials. 2002;23(22):4315–23.
Bryant SJ, et al. Encapsulating chondrocytes in degrading PEG hydrogels with high modulus: engineering gel structural changes to facilitate cartilaginous tissue production. Biotechnol Bioeng. 2004;86(7):747–55.
Bryant SJ, Anseth KS. Controlling the spatial distribution of ECM components in degradable PEG hydrogels for tissue engineering cartilage. J Biomed Mater Res A. 2003;64(1):70–9.
Elisseeff J, et al. Photoencapsulation of chondrocytes in poly(ethylene oxide)-based semi-interpenetrating networks. J Biomed Mater Res. 2000;51(2):164–71.
Mann B, et al. Smooth muscle cell growth in photopolymerized hydrogels with cell adhesive and proteolytically degradable domains: synthetic ECM analogs for tissue engineering. Biomaterials. 2001;22(22):3045–51.
Hern DL, Hubbell JA. Incorporation of adhesion peptides into nonadhesive hydrogels useful for tissue resurfacing. J Biomed Mater Res. 1998;39(2):266–76.
Sawhney AS, et al. Optimization of photopolymerized bioerodible hydrogel properties for adhesion prevention. J Biomed Mater Res. 1994;28(7):831–8.
Elisseeff J, et al. Controlled-release of IGF-I and TGF-beta1 in a photopolymerizing hydrogel for cartilage tissue engineering. J Orthop Res. 2001;19(6):1098–104.
Anseth KS, Burdick JA. New directions in photopolymerizable biomaterials. MRS Bull. 2002;27(2):130–6.
Lee C, Grodzinsky A, Spector M. The effects of cross-linking of collagen-glycosaminoglycan scaffolds on compressive stiffness, chondrocyte-mediated contraction, proliferation and biosynthesis. Biomaterials. 2001;22(23):3145–54.
Lee C, Grodzinsky A, Spector M. Modulation of the contractile and Biosynthetic activity of chondrocytes seeded in collagen-glycosaminoglycan matrices. Tissue Eng. 2003;9(1):27–36.
Schoof H, et al. Control of pore structure and size in freeze-dried collagen sponges. J Biomed Mater Res. 2001;58(4):352–7.
Chen G, Ushida T, Tateishi T. A hybrid sponge of poly(dl-lactic-co-glycolic acid), collagen and apatite. Bioceramics. 2000;192–1:753–6.
Mann B, Schmedlen R, West J. Tethered-TGF-beta increases extracellular matrix production of vascular smooth muscle cells. Biomaterials. 2001;22(5):439–44.
Suzuki Y, et al. Alginate hydrogel linked with synthetic oligopeptide derived from BMP-2 allows ectopic osteoinduction in vivo. J Biomed Mater Res. 2000;50(3):405–9.
Zimmermann WH, et al. Tissue engineering of a differentiated cardiac muscle construct. Circ Res. 2002;90(2):223–30.
Ou K-L, Hosseinkhani H. Development of 3D in vitro technology for medical applications. Int J Mol Sci. 2014;15(10):17938–62.
Hosseinkhani H, Hong P-D, Yu D-S. Self-assembled proteins and peptides for regenerative medicine. Chem Rev. 2013;113(7):4837–61.
Hosseinkhani H, et al. Engineering three-dimensional collagen-IKVAV matrix to mimic neural microenvironment. ACS Chem Neurosci. 2013;4(8):1229–35.
Hosseinkhani H, et al. Development of 3D in vitro platform technology to engineer mesenchymal stem cells. Int J Nanomed. 2012;7:3035.
Mahmoudi M, et al. Magnetic resonance imaging tracking of stem cells in vivo using iron oxide nanoparticles as a tool for the advancement of clinical regenerative medicine. Chem Rev. 2010;111(2):253–80.
Subramani K, et al. Bone regeneration around dental implants as a treatment for peri-implantitis: a review of the literature. J Biomimetics Biomater Tissue Eng. 2011;11:21–33.
Hosseinkhani H, et al. Micro and nano-scale in vitro 3D culture system for cardiac stem cells. J Biomed Mater Res Part A. 2010;94(1):1–8.
Mohajeri S, et al. Proliferation and differentiation of mesenchymal stem cell on collagen sponge reinforced with polypropylene/polyethylene terephthalate blend fibers. Tissue Eng Part A. 2010;16(12):3821–30.
Hosseinkhani H, et al. DNA nanoparticles encapsulated in 3D tissue-engineered scaffolds enhance osteogenic differentiation of mesenchymal stem cells. J Biomed Mater Res Part A. 2008;85(1):47–60.
Hosseinkhani H, et al. Combination of 3D tissue engineered scaffold and non-viral gene carrier enhance in vitro DNA expression of mesenchymal stem cells. Biomaterials. 2006;27(23):4269–78.
Hosseinkhani H, et al. Ectopic bone formation in collagen sponge self-assembled peptide–amphiphile nanofibers hybrid scaffold in a perfusion culture bioreactor. Biomaterials. 2006;27(29):5089–98.
Hosseinkhani H, et al. Enhanced angiogenesis through controlled release of basic fibroblast growth factor from peptide amphiphile for tissue regeneration. Biomaterials. 2006;27(34):5836–44.
Hosseinkhani H, Hosseinkhani M, Kobayashi H. Proliferation and differentiation of mesenchymal stem cells using self-assembled peptide amphiphile nanofibers. Biomed Mater. 2006;1(1):8.
Hosseinkhani H, Hosseinkhani M, Kobayashi H. Design of tissue-engineered nanoscaffold through self-assembly of peptide amphiphile. J Bioactive Compatible Polym. 2006;21(4):277–96.
Hosseinkhani H, et al. Impregnation of plasmid DNA into three-dimensional scaffolds and medium perfusion enhance in vitro DNA expression of mesenchymal stem cells. Tissue Eng. 2005;11(9–10):1459–75.
Hosseinkhani H, et al. Perfusion culture enhances osteogenic differentiation of rat mesenchymal stem cells in collagen sponge reinforced with poly (glycolic acid) fiber. Tissue Eng. 2005;11(9–10):1476–88.
Vercruysse KP, et al. Synthesis and in vitro degradation of new polyvalent hydrazide cross-linked hydrogels of hyaluronic acid. Bioconjug Chem. 1997;8(5):686–94.
Prestwich GD, et al. Controlled chemical modification of hyaluronic acid: synthesis, applications, and biodegradation of hydrazide derivatives. J Control Release. 1998;53(1–3):93–103.
Burdick JA, et al. Controlled degradation and mechanical behavior of photopolymerized hyaluronic acid networks. Biomacromolecules. 2005;6(1):386–91.
Gamini A, et al. Structural investigations of cross-linked hyaluronan. Biomaterials. 2002;23(4):1161–7.
Pei M, et al. Bioreactors mediate the effectiveness of tissue engineering scaffolds. Faseb J. 2002;16(12):1691–4.
Mullegger J, Lepperdinger G. Hyaluronan is an abundant constituent of the extracellular matrix of Xenopus embryos. Mol Reprod Dev. 2002;61(3):312–6.
Luo Y, Kirker KR, Prestwich GD. Cross-linked hyaluronic acid hydrogel films: new biomaterials for drug delivery. J Control Release. 2000;69(1):169–84.
Leach JB, Schmidt CE. Characterization of protein release from photocrosslinkable hyaluronic acid-polyethylene glycol hydrogel tissue engineering scaffolds. Biomaterials. 2005;26(2):125–35.
Leach JB, et al. Development of photocrosslinkable hyaluronic acid-polyethylene glycol-peptide composite hydrogels for soft tissue engineering. J Biomed Mater Res A. 2004;70(1):74–82.
Baier Leach J, et al. Photocrosslinked hyaluronic acid hydrogels: natural, biodegradable tissue engineering scaffolds. Biotechnol Bioeng. 2003;82(5):578–89.
Zhang S. Fabrication of novel biomaterials through molecular self-assembly. Nat Biotechnol. 2003;21(10):1171–8.
Hartgerink JD, Beniash E, Stupp SI. Self-assembly and mineralization of peptide-amphiphile nanofibers. Science. 2001;294(5547):1684–8.
Silva GA, et al. Selective differentiation of neural progenitor cells by high-epitope density nanofibers. Science. 2004;303(5662):1352–5.
Kourouklis AP, Kaylan KB, Underhill GH. Substrate stiffness and matrix composition coordinately control the differentiation of liver progenitor cells. Biomaterials. 2016;99:82–94.
Cozzolino AM, et al. Modulating the substrate stiffness to manipulate differentiation of resident liver stem cells and to improve the differentiation state of hepatocytes. Stem Cells Int. 2016;2016:5481493.
Xia T, et al. Effect of substrate stiffness on hepatocyte migration and cellular Young’s modulus. J Cell Physiol. 2018;233(9):6996–7006.
Maghsoudlou P, et al. Optimization of liver decellularization maintains extracellular matrix micro-architecture and composition predisposing to effective cell seeding. PLoS ONE. 2016;11(5):e0155324.
Struecker B, et al. Improved rat liver decellularization by arterial perfusion under oscillating pressure conditions. J Tissue Eng Regen Med. 2017;11(2):531–41.
Coronado RE, et al. Decellularization and solubilization of porcine liver for use as a substrate for porcine hepatocyte culture: method optimization and comparison. Cell Transplant. 2017;26(12):1840–54.
Jaramillo M, et al. Decellularized human liver extracellular matrix (hDLM) mediated hepatic differentiation of human induced pluripotent stem cells (hIPSc). J Tissue Eng Regen Med. 2017;12(4):e1962.
Vyas D, et al. Self-assembled liver organoids recapitulate hepatobiliary organogenesis in vitro. Hepatology. 2018;67(2):750–61.
Kanninen LK, et al. Hepatic differentiation of human pluripotent stem cells on human liver progenitor HepaRG-derived acellular matrix. Exp Cell Res. 2016;341(2):207–17.
Bi H, et al. Liver extracellular matrix promotes BM-MSCs hepatic differentiation and reversal of liver fibrosis through activation of integrin pathway. J Tissue Eng Regen Med. 2017;11(10):2685–98.
Fukuda T, et al. Isolation and expansion of human pluripotent stem cell-derived hepatic progenitor cells by growth factor defined serum-free culture conditions. Exp Cell Res. 2017;352(2):333–45.
Mitani S, et al. Human ESC/iPSC-derived hepatocyte-like cells achieve zone-specific hepatic properties by modulation of WNT signaling. Mol Ther. 2017;25(6):1420–33.
Toivonen S, et al. Regulation of human pluripotent stem cell-derived hepatic cell phenotype by three-dimensional hydrogel models. Tissue Eng Part A. 2016;22(13–14):971–84.
Wang B, et al. Functional maturation of induced pluripotent stem cell hepatocytes in extracellular matrix—a comparative analysis of bioartificial liver microenvironments. Stem Cells Transl Med. 2016;5(9):1257–67.
Mattei G, et al. Decellularized human liver is too heterogeneous for designing a generic extracellular matrix mimic hepatic scaffold. Artif Organs. 2017;41(12):E347–55.
Kojima H, et al. Establishment of practical recellularized liver graft for blood perfusion using primary rat hepatocytes and liver sinusoidal endothelial cells. Am J Transplant. 2018;18(6):1351–9.
Xu B, Håkansson J, Kuna VK, et al. Assessing rat liver-derived biomatrix for hepatic tissue engineering with human fetal liver stem cells. Adv Tissue Eng Regen Med. 2016;1(3):64–70.
Robertson MJ, et al. Recellularization of rat liver: an in vitro model for assessing human drug metabolism and liver biology. PLoS ONE. 2018;13(1):e0191892.
Naeem EM, et al. Decellularized liver transplant could be recellularized in rat partial hepatectomy model. J Biomed Mater Res Part A. 2019;107(11):2576–88.
Acun A, Oganesyan R, Uygun BE. Liver bioengineering: promise, pitfalls, and hurdles to overcome. Curr Transplant Rep. 2019;6(2):119–26.
Barreto RS, et al. Mouse placental scaffolds: a three-dimensional environment model for recellularization. J Tissue Eng. 2019;10:2041731419867962.
Rajendran D, et al. Long-term liver-specific functions of hepatocytes in electrospun chitosan nanofiber scaffolds coated with fibronectin. J Biomed Mater Res Part A. 2017;105(8):2119–28.
Arca HÇ, Senel S. Chitosan based systems for tissue engineering part II: soft tissues. FABAD J Pharm Sci. 2008;33(4):211.
Du C, et al. Induced pluripotent stem cell-derived hepatocytes and endothelial cells in multi-component hydrogel fibers for liver tissue engineering. Biomaterials. 2014;35(23):6006–14.
Che Abdullah CA, et al. Primary liver cells cultured on carbon nanotube substrates for liver tissue engineering and drug discovery applications. ACS Appl Mater Interfaces. 2014;6(13):10373–80.
Fung J, et al. Defining normal liver stiffness range in a normal healthy Chinese population without liver disease. PLoS ONE. 2013;8(12):e85067.
You J, et al. Characterizing the effects of heparin gel stiffness on function of primary hepatocytes. Tissue Eng Part A. 2013;19(23–24):2655–63.
Wang L, Carrier RL. Biomimetic topography: bioinspired cell culture substrates and scaffolds. In: Advances in biomimetics. InTech; 2011.
Vasanthan KS, et al. Development of porous hydrogel scaffolds with multiple cues for liver tissue engineering. Regen Eng Transl Med. 2017;3(3):176–91.
Mamaeva V, Sahlgren C, Lindén M. Mesoporous silica nanoparticles in medicine—recent advances. Adv Drug Deliv Rev. 2013;65(5):689–702.
Wang M, et al. Sustained delivery growth factors with polyethyleneimine-modified nanoparticles promote embryonic stem cells differentiation and liver regeneration. Adv Sci. 2016;3(8):1500393.
Nguyen D, et al. Functional characterization of three-dimensional (3d) human liver tissues generated by an automated bioprinting platform. FASEB J. 2015;29(1_supplement):LB424.
Robbins JB, et al. A novel in vitro three-dimensional bioprinted liver tissue system for drug development. FASEB J. 2013;27(872):812.
Vinken M, et al. Primary hepatocytes and their cultures in liver apoptosis research. Arch Toxicol. 2014;88(2):199–212.
Bell CC, et al. Characterization of primary human hepatocyte spheroids as a model system for drug-induced liver injury, liver function and disease. Sci Rep. 2016;6:25187.
Boess F, et al. Gene expression in two hepatic cell lines, cultured primary hepatocytes, and liver slices compared to the in vivo liver gene expression in rats: possible implications for toxicogenomics use of in vitro systems. Toxicol Sci. 2003;73(2):386–402.
Rodriguez-Antona C, et al. Cytochrome P450 expression in human hepatocytes and hepatoma cell lines: molecular mechanisms that determine lower expression in cultured cells. Xenobiotica. 2002;32(6):505–20.
Swift B, Brouwer KLJ. Influence of seeding density and extracellular matrix on bile acid transport and mrp4 expression in sandwich-cultured mouse hepatocytes. Mol Pharm. 2009;7(2):491–500.
Lin RZ, Chang HY. Recent advances in three-dimensional multicellular spheroid culture for biomedical research. Biotechnol J. 2008;3(9–10):1172–84.
Li CY, et al. Micropatterned cell–cell interactions enable functional encapsulation of primary hepatocytes in hydrogel microtissues. Tissue Eng Part A. 2014;20(15–16):2200–12.
Underhill GH, et al. Assessment of hepatocellular function within PEG hydrogels. Biomaterials. 2007;28(2):256–70.
Glicklis R, et al. Hepatocyte behavior within three-dimensional porous alginate scaffolds. Biotechnol Bioeng. 2000;67(3):344–53.
Baranski JD, et al. Geometric control of vascular networks to enhance engineered tissue integration and function. Proc Natl Acad Sci USA. 2013;110:7586–91.
Ranucci CS, et al. Control of hepatocyte function on collagen foams: sizing matrix pores toward selective induction of 2-D and 3-D cellular morphogenesis. Biomaterials. 2000;21(8):783–93.
Tsang VL, et al. Fabrication of 3D hepatic tissues by additive photopatterning of cellular hydrogels. FASEB J. 2007;21(3):790–801.
Hailfinger S, et al. Zonal gene expression in murine liver: lessons from tumors. Hepatology. 2006;43(3):407–14.
Sainz B, TenCate V, Uprichard SL. Three-dimensional Huh7 cell culture system for the study of Hepatitis C virus infection. Virol J. 2009;6(1):103.
Skardal A, et al. A hydrogel bioink toolkit for mimicking native tissue biochemical and mechanical properties in bioprinted tissue constructs. Acta Biomater. 2015;25:24–34.
Bhise NS, et al. A liver-on-a-chip platform with bioprinted hepatic spheroids. Biofabrication. 2016;8(1):014101.
Lee H, Cho D-W. One-step fabrication of an organ-on-a-chip with spatial heterogeneity using a 3D bioprinting technology. Lab Chip. 2016;16(14):2618–25.
Nguyen DG, et al. Bioprinted 3D primary liver tissues allow assessment of organ-level response to clinical drug induced toxicity in vitro. PLoS ONE. 2016;11(7):e0158674.
Xu W, et al. Rapid prototyping three-dimensional cell/gelatin/fibrinogen constructs for medical regeneration. J Bioactive Compatible Polym. 2007;22(4):363–77.
Li S, et al. Direct fabrication of a hybrid cell/hydrogel construct by a double-nozzle assembling technology. J Bioactive Compatible Polym. 2009;24(3):249–65.
Vanderburgh J, Sterling JA, Guelcher SA. 3D printing of tissue engineered constructs for in vitro modeling of disease progression and drug screening. Ann Biomed Eng. 2017;45(1):164–79.
Leijten J, et al. Spatially and temporally controlled hydrogels for tissue engineering. Mater Sci Eng R. 2017;119:1–35.
Billiet T, et al. The 3D printing of gelatin methacrylamide cell-laden tissue-engineered constructs with high cell viability. Biomaterials. 2014;35(1):49–62.
Du M, et al. 3D bioprinting of BMSC-laden methacrylamide gelatin scaffolds with CBD-BMP2-collagen microfibers. Biofabrication. 2015;7(4):044104.
Arcaute K, Mann B, Wicker RJ. Stereolithography of spatially controlled multi-material bioactive poly (ethylene glycol) scaffolds. Acta Biomater. 2010;6(3):1047–54.
Hon K, Li L, Hutchings IJ. Direct writing technology—advances and developments. CIRP Ann. 2008;57(2):601–20.
Chang R, et al. Biofabrication of a three-dimensional liver micro-organ as an in vitro drug metabolism model. Biofabrication. 2010;2(4):045004.
Snyder J, et al. Bioprinting cell-laden matrigel for radioprotection study of liver by pro-drug conversion in a dual-tissue microfluidic chip. Biofabrication. 2011;3(3):034112.
Neiman JAS, et al. Photopatterning of hydrogel scaffolds coupled to filter materials using stereolithography for perfused 3D culture of hepatocytes. Biotechnol Bioeng. 2015;112(4):777–87.
Chang R, Nam J, Sun W. Direct cell writing of 3D microorgan for in vitro pharmacokinetic model. Tissue Eng Part C Methods. 2008;14(2):157–66.
Nair K, et al. A multilevel numerical model quantifying cell deformation in encapsulated alginate structures. J Mech Mater Struct. 2007;2(6):1121–39.
Guvendiren M, et al. Hydrogels with differential and patterned mechanics to study stiffness-mediated myofibroblastic differentiation of hepatic stellate cells. J Mech Behav Biomed Mater. 2014;38:198–208.
Guvendiren M, Burdick JA. Stiffening hydrogels to probe short-and long-term cellular responses to dynamic mechanics. Nat Commun. 2012;3:792.
Caliari SR, et al. Stiffening hydrogels for investigating the dynamics of hepatic stellate cell mechanotransduction during myofibroblast activation. Sci Rep. 2016;6:21387.
Asumda FZ, et al. Differentiation of hepatocyte-like cells from human pluripotent stem cells using small molecules. Differentiation. 2018;101:16–24.
Basma H, et al. Differentiation and transplantation of human embryonic stem cell-derived hepatocytes. Gastroenterology. 2009;136(3):990–999.e4.
Takayama K, et al. Efficient generation of functional hepatocytes from human embryonic stem cells and induced pluripotent stem cells by HNF4α transduction. Mol Ther. 2012;20(1):127–37.
Sa-ngiamsuntorn K, et al. A robust model of natural hepatitis C infection using hepatocyte-like cells derived from human induced pluripotent stem cells as a long-term host. Virol J. 2016;13(1):59.
Hannoun Z, et al. The potential of induced pluripotent stem cell derived hepatocytes. J Hepatol. 2016;65(1):182–99.
Carpentier A, et al. Hepatic differentiation of human pluripotent stem cells in miniaturized format suitable for high-throughput screen. Stem Cell Res. 2016;16(3):640–50.
Lang J, et al. Modeling dengue virus-hepatic cell interactions using human pluripotent stem cell-derived hepatocyte-like cells. Stem Cell Rep. 2016;7(3):341–54.
Sun C, et al. Hepatic differentiation of rat induced pluripotent stem cells in vitro. World J Gastroenterol. 2015;21(39):11118.
Carpentier A, et al. Engrafted human stem cell–derived hepatocytes establish an infectious HCV murine model. J Clin Invest. 2014;124(11):4953–64.
Hannan NR, et al. Generation of multipotent foregut stem cells from human pluripotent stem cells. Stem Cell Rep. 2013;1(4):293–306.
Shan J, et al. Identification of small molecules for human hepatocyte expansion and iPS differentiation. Nat Chem Biol. 2013;9(8):514–20.
Liu J, et al. Direct differentiation of hepatic stem-like WB cells into insulin-producing cells using small molecules. Sci Rep. 2013;3:1185.
Mathapati S, et al. Small-molecule-directed hepatocyte-like cell differentiation of human pluripotent stem cells. Curr Protoc Stem Cell Biol. 2016;38(1):1G.6.1–6.18.
Du C, et al. Highly efficient and expedited hepatic differentiation from human pluripotent stem cells by pure small-molecule cocktails. Stem Cell Res Ther. 2018;9(1):58.
Itaba N, et al. Identification of the small molecule compound which induces hepatic differentiation of human mesenchymal stem cells. Regen Ther. 2015;2:32–41.
Zhu S, et al. A small molecule primes embryonic stem cells for differentiation. Cell Stem Cell. 2009;4(5):416–26.
Kim J, et al. An extended transcriptional network for pluripotency of embryonic stem cells. Cell. 2008;132(6):1049–61.
Ouyang J, et al. Hepatic differentiation of rat mesenchymal stem cells by a small molecule. ChemMedChem. 2012;7(8):1447–52.
Wuestefeld T, et al. A direct in vivo RNAi screen identifies MKK4 as a key regulator of liver regeneration. Cell. 2013;153(2):389–401.
Emre N, Coleman R, Ding S. A chemical approach to stem cell biology. Curr Opin Chem Biol. 2007;11(3):252–8.
Shi Y, et al. A combined chemical and genetic approach for the generation of induced pluripotent stem cells. Cell Stem Cell. 2008;2(6):525–8.
Soheilyfar S, et al. In vivo and in vitro impact of miR-31 and miR-143 on the suppression of metastasis and invasion in breast cancer. J Buon. 2018;23(5):1290–6.
Zeng Z-L, et al. MicroRNAs: important regulators of induced pluripotent stem cell generation and differentiation. Stem Cell Rev Rep. 2018;14(1):71–81.
Skrzypczyk A, et al. Noncoding RNA transcripts during differentiation of induced pluripotent stem cells into hepatocytes. Stem Cells Int. 2018;2018:5692840.
Deng XG, et al. Overexpression of miR-122 promotes the hepatic differentiation and maturation of mouse ESCs through a miR-122/FoxA1/HNF4a-positive feedback loop. Liver Int. 2014;34(2):281–95.
Hino K, et al. Inducible expression of microRNA-194 is regulated by HNF-1α during intestinal epithelial cell differentiation. RNA. 2008;14(7):1433–42.
Xu J, et al. MiR-194 regulates chondrogenic differentiation of human adipose-derived stem cells by targeting Sox5. PLoS ONE. 2012;7(3):e31861.
Lian I, et al. The role of YAP transcription coactivator in regulating stem cell self-renewal and differentiation. Genes Dev. 2010;24(11):1106–18.
Davoodian N, et al. Let-7f microRNA negatively regulates hepatic differentiation of human adipose tissue-derived stem cells. J Physiol Biochem. 2014;70(3):781–9.
Zhou X, et al. Induction of hepatocyte-like cells from human umbilical cord-derived mesenchymal stem cells by defined microRNAs. J Cell Mol Med. 2017;21(5):881–93.
Kleger A, et al. Increased reprogramming capacity of mouse liver progenitor cells, compared with differentiated liver cells, requires the BAF complex. Gastroenterology. 2012;142(4):907–17.
Huang C-S, et al. Generation of high quality of hepatocyte-like cells from induced pluripotent stem cells with Parp1 but lacking c-Myc. J Chin Med Assoc. 2018;81(10):871–7.
Ishizaka S, et al. Development of hepatocytes from ES cells after transfection with the HNF-3β gene. FASEB J. 2002;16(11):1444–6.
Devalaraja-Narashimha K, Padanilam BJ. PARP1 deficiency exacerbates diet-induced obesity in mice. J Endocrinol. 2010;205(3):243–52.
Kraus WL. Transcriptional control by PARP-1: chromatin modulation, enhancer-binding, coregulation, and insulation. Curr Opin Cell Biol. 2008;20(3):294–302.
Li Y, et al. Immunogenicity of hepatic differentiated human umbilical cord mesenchymal stem cells promoted by porcine decellularized liver scaffolds. Xenotransplantation. 2017. https://doi.org/10.1111/xen.12287.
Wang B, et al. Enhanced hepatogenic differentiation of bone marrow derived mesenchymal stem cells on liver ECM hydrogel. J Biomed Mater Res Part A. 2018;106(3):829–38.
Wu Q, et al. Hepatic differentiation of mouse bone marrow-derived mesenchymal stem cells using a novel 3D culture system. Mol Med Rep. 2017;16(6):9473–9.
Acknowledgements
Authors wish to thank the personnel of Stem Cell Research Center for kindest collaboration.
Funding
This work was supported by a Grant from the National Council for Development of Stem Cell Sciences and Technologies, Iran.
Author information
Authors and Affiliations
Contributions
VH, NFM, SS, NS and SNSA: data collection and manuscript writing and figure preparation; MD, HH and RR: equal conceptualization and manuscript edition. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Hosseini, V., Maroufi, N.F., Saghati, S. et al. Current progress in hepatic tissue regeneration by tissue engineering. J Transl Med 17, 383 (2019). https://doi.org/10.1186/s12967-019-02137-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12967-019-02137-6