
Abstract
Background
Myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) is a chronic debilitating disease with huge social-economic impact. It has been suggested that immune dysregulation, nitrooxidative stress, and metabolic impairment might contribute to disease pathogenesis. However, the etiology of ME/CFS remains largely unclear, and diagnostic/prognostic disease markers are lacking. Several long noncoding RNAs (lncRNA, > 200 bp) have been reported to play roles in immunological diseases or in stress responses.
Methods
In our study, we examined the expression signature of 10 very long lncRNAs (> 5 kb, CR933609, His-RNA, AK124742, GNAS1-AS, EmX2OS, MIAT, TUG1, NEAT1, MALAT1, NTT) in the peripheral blood mononuclear cells of 44 ME/CFS patients.
Results
LncRNAs NTT, MIAT and EmX2OS levels were found to be significantly elevated in ME/CFS patients as compared with healthy controls. Furthermore, NTT and EmX2OS levels increased with disease severity. Stimulation of human monocytic cell line THP-1 and glioma cell line KALS1 with H2O2 (oxidative stress) and poly (I:C) (double strand RNA, representing viral activation) increased the expression levels of NTT and MIAT.
Conclusions
Our study revealed a ME/CFS-associated very long lncRNA expression signature, which might reflect the regulatory response in ME/CFS patients to oxidative stress, chronic viral infection and hypoxemia. Further investigations need to be done to uncover the functions and potential diagnostic value of these lncRNAs in ME/CFS.
Similar content being viewed by others


Background
Myalgic encephalomyelitis/chronic fatigue syndrome (ME/CFS) is a chronic debilitating disease with huge social-economic burden. The prevalence rate is 1–3/1000 in Germany and in the US population, affecting more than 1 million Americans [1]. In Taiwan, the estimated overall incidence rate of ME/CFS is 0.87–1.37/1000 person-year [2]. According to the Canadian criteria for diagnosis, ME/CFS patients suffer from more than 6 months of significant fatigue and post-exertional malaise, which describes the aggravation of symptoms upon exertion associated with sleep disturbance, myalgia/arthralgia, cognitive impairment, and autonomic dysfunction [3]. The pathogenesis of ME/CFS is complex, and remains mostly elusive so far. There is evidence that in a subset of patients infections result in chronic immune activation and autoimmunity [4]. Autoantibodies to neurotransmitters have been found in a subset of ME/CFS patients [5]. In addition, elevated cytokines and oxidative/nitrosative stresses possibly facilitate blood brain barrier disruption, neuroinflammation, and glial activation and hypersensitivity, which further triggers dysregulation of neurotransmitters and amplification of inflammatory signals [4]. More recently, mitochondrial dysfunction and metabolic disturbances have also been reported to underlie the potential mechanisms of ME/CFS [6, 7]. Despite these evidences, diagnostic markers are still lacking, and the diagnosis is usually made by clinical criteria.
Long non-coding RNAs are defined as RNAs that are more than 200 nucleotides long, which do not encode proteins. Through RNA–DNA, RNA–RNA, or RNA–protein interactions, lncRNA can affect different stages of gene regulation [8]. LncRNAs are key regulators of chromatin state, which show great capacity to interact with more than one protein in different context, and fine-tune the cellular response [8]. It has been reported that lncRNAs play essential roles in complex diseases, such as cancer and autoimmune diseases [9,10,11]. Although more and more lncRNAs are being discovered, most of their functions and mechanisms of actions are still unknown, especially for the very large lncRNAs that have sizes of more than 5000 nucleotides. We had particular interests in ten very large lncRNAs (> 5 kb), which have been either reported to be involved in immune regulation, or are located close to genes regulating stress response, metabolic and neurologic processes, thus potentially playing a role in ME/CFS.
The ten lncRNAs are NTT (17 kb), NEAT1 (23 kb), MALAT1 (7.5 kb), TUG1 (7.1 kb), MIAT (9.9 kb), His-1 RNA (8.4 kb), GNAS1-AS (8.9 kb), EMX2OS (7.3 kb), CR933609 (8.8 kb) and AK124742 (6 kb). NTT was first described in activated T cells, while NEAT1 has been reported to be involved in human lupus and in immune response to viral infections [11,12,13]. MALAT1 has been found to regulate LPS-induced inflammatory response, and TUG1 is involved in the regulation of cold-induced oxidative stress and inflammation [14,15,16]. As for MIAT, it is known to play roles in a variety of disease processes, including myocardial infarction, microvascular dysfunction, schizophrenia, and neurogenic commitment [17, 18]. His-1 RNA has been implicated in leukemogenesis, and GNAS1-AS is an imprinted anti-sense transcript at the locus of GNAS1, encoding neuroendocrine secretory protein [19, 20]. According to the lncrnadb database, EMX2OS is an opposite strand transcript of EMX2 gene, and possibly regulates EMX2 [21]. Both EMX2OS and EMX2 RNAs have been detected in the central nervous system (CNS) tissues [22]. For CR933609, we have previously identified its role in protecting INO80D from downregulation by miRNA-5096 [23]. Since INO80D is a main component of the chromatin re-modeler INO80 complex which regulates cell glycolytic and respiratory capacities, CR933609 could be involved in maintaining metabolic stability [24]. Finally, AK124742 has been reported to be an antisense RNA to the gene PSMD6, which encodes components of proteasome, involving antigen presentation by MHC class I and DNA damage repair [25, 26].
In this study, we aimed to investigate the expression signatures and potential diagnostic values of the ten very large lncRNAs (> 5 kb) in ME/CFS patients. Furthermore, the effects of oxidative stress (H2O2) and Toll like receptor 3 (TLR3) ligand poly (I:C) (mimicking viral infection) on the levels of very large lncRNAs were also evaluated in human monocytic cell line THP-1 and glioma cell line KALS1.
Methods
Subjects
Forty-four ME/CFS patients diagnosed according to the Canadian criteria and 30 sex and age-matched healthy controls were recruited. Characteristics of the study population are shown in Table 1. The study was approved by the Ethics Committee of Charité Universitätsmedizin Berlin in accordance with the 1964 Declaration of Helsinki and its later amendments and patients gave written informed consent.
RNA extraction and RT-PCR for lncRNAs
RNAs were isolated from PBMCs using TRIzol Reagent (Thermo Fisher Scientific-Invitrogen, Waltham, Massachusetts, USA). High Capacity cDNA Reverse Transcriptase Kit (Thermo Fisher Scientific-Applied Biosystems, Waltham, Massachusetts, USA) was used to reverse-transcribe 2 μg RNA into cDNA using for real time PCR analyses. GAPDH expression was used as an endogenous control. All primers were designed and synthesized by Genomics BioSci & Tech, Taipei, Taiwan. The following primer sequences were used: NTT forward 5′-cttggcctaaaaggggatg-3′, reverse 5′-gcacctttggtctccttcac-3′; MALAT1 forward 5′-gacccttcacccctcacc-3′, reverse 5′-ttatggatcatgcccacaag-3′; TUG1 forward 5′-gtctccgatagtgcacacagc-3′, reverse 5′-gaccatctccttcaggacca-3′; NEAT1 forward 5′-ctctgacccgaagggtagg-3′, reverse 5′-ctggcagctttgctcctg-3′; MIAT forward 5′-ctggagagggaggcatctaa-3′, reverse 5′- aactcatccccacccacac-3′; His-1 RNA forward 5′-cagtcttctttgaactgctactcct-3′, reverse 5′-tttttgtaacgctctggtcaaa-3′; GNAS1-AS forward 5′-ctgctatctgcagaggggtct-3′, reverse 5′-tcctttagctgcatttgct-3′; EMX2OS forward 5′-gtgacttgcacaaggacacaa-3′, reverse 5′-cctgtvtggccattcctct-3′; CR933609 forward 5′-gacaaaacaaactagtgaagcacct-3′, reverse 5′-tatacaccttgacacggcaga-3′; AK124742 forward 5′- aagttgtaggcatccttctagcc-3′, reverse 5′- cacttttgttaagcacccaactt-3′; IFNGR1 forward 5′-catgcagggtgtgagcag-3′, reverse 5′-aacattagttggtgtaggcactga-3′; PBOV1 forward 5′-gaaaaagattctcatcactcaac-3′, reverse 5′-ggttctcaaacagccttcc-3′; GAPDH: forward 5′-agccacatcgctcagacac-3′, reverse 5′-gcccaatacgaccaaatcc-3′.
Cell line studies
Human monocytic cell line THP-1 and glioma cell line KALS1 were grown in LPS-free complete RPMI medium containing 10% fetal bovine serum, at 37 °C incubator with 5% CO2. Cells were treated with different concentrations of peroxide (H2O2) (Sigma Merk, Darmstadt, Germany) or poly (I:C) (InvivoGen, California, USA) for 6 h. RNAs were then extracted for further RT-PCR.
Bioinformatics and statistical analysis
Principle component analyses were performed using Stata 13 software. Mann–Whitney U tests were used via GraphPad Prism 5 to analyze the difference of lncRNA expression levels between ME/CFS and controls. Receiver operating characteristic (ROC) curve analysis was performed using MedCalc v.14.
Results
PBMC lncRNA profile in chronic fatigue patients and controls
Expression of the ten lncRNAs (NTT, NEAT1, MALAT1, TUG1, MIAT, His-1 RNA, GNAS1-AS, EMX2OS, CR933609 and AK124742) in peripheral blood mononuclear cells (PBMCs) of ME/CFS patients and healthy controls were evaluated by RT-PCR. The values of the ten ΔCT (CTlncRNA–CTGAPDH) of each individual subject were used to build a correlation matrix, and projected to principle component space by principle component analysis (PCA) (Fig. 1a). The PCA showed that the ME/CFS and the control lncRNA expression profile could be separated mainly by principle component 2 (dimension 2). Similarly, the eigenvalues showed that data variances is mostly explained by the first and the second components (Fig. 1b).

Principle component analysis (PCA) of ΔCT values of ten lncRNAs from PBMCs of ME/CFS patients and controls. a Variance between ME/CFS (red dots) and control (green dots) according to the ten lncRNA correlation matrix, displayed onto a two-dimension plot. b Eigenvalues of principle components
Association of lncRNA expression levels with ME/CFS and disease severity
In order to reduce the test numbers of lncRNAs needed for differentiating ME/CFS from healthy controls, and to evaluate the association of lncRNA expression level with ME/CFS disease severity, we analyzed the amount of each lncRNA in PBMCs of controls, mild-moderate ME/CFS (with Bell score ≥ 30), and severe ME/CFS (with Bell score < 30) (Fig. 2a–j). Interestingly, NTT, MIAT and EMX2OS levels were found to be significantly higher in ME/CFS PBMCs as compared with controls (ME/CFS median ΔCT vs. control median ΔCT: NTT 8.86 vs. 10.05, Mann–Whitney U test p < 0.0001; MIAT 6.22 vs. 6.89, Mann–Whitney U test p < 0.05; EMX2OS 20.69 vs. 18.59, Mann–Whitney U test p < 0.001). Furthermore, NTT and EMX2OS expression levels correlated with ME/CFS disease severity, with highest amount detected in ME/CFS patients with Bell score of 10 or 20 (< 30) (Fig. 2a, h).

Expression of the ten very long lncRNA in PBMCs of ME/CFS and controls (a–j). ME/CFS patients with Bell score < 30 belong to the severe group. *p < 0.05, **p < 0.01, ***p < 0.001 by Mann–Whitney U tests. Lines represent medians. ns not statistically significant
PBMC NTT, MIAT and EMX2OS expression signature as ME/CFS diagnostic markers
Since we have discovered that among the ten lncRNAs tested, only NTT, MIAT and EMX2OS levels were upregulated in ME/CFS, we plotted a new PCA plot using a correlation matrix of NTT, MIAT and EMX2OS ΔCT values (Fig. 3a). In Fig. 3a, ME/CFS patients could be separated from healthy controls by principle component 1 (dimension 1). The ME/CFS group could still be separated from the control group when the input lncRNA values were reduced to those of NTT and MIAT (Fig. 3b) or those of NTT and EMX2OS (Fig. 3c). However, the ME/CFS and the control group could not be well differentiated from each other when the PCA was performed on a correlation matrix of only MIAT and EMX2OS (Fig. 3d).

Diagnostic value of NTT, MIAT and EMX2OS expressions in ME/CFS. PCA showing the distribution of ME/CFS and controls on two-dimension plots differentiated by values derived from the expression levels of NTT, MIAT and EMX2OS (a) or from the expression levels of any two of the three lncRNAs (b–d). e Receiver-operating characteristic (ROC) curve analysis of the three-lncRNA signature to discriminate ME/CFS patients from healthy controls. AUC = 0.82 when the criteria is increased expression (ΔCT below the optimal cutoff) of any two of the three lncRNAs (NTT, MIAT and EMX2OS)
To evaluate the diagnostic value of the combination of the expression of NTT, MIAT and EMX2OS, we first obtained the optimal cutoff ΔCT value of each of the three lncRNAs by using receiver operating characteristic (ROC) curves. The optimal cutoff ΔCT value for NTT, MIAT and EMX2OS were 9.49 (area under the curve (AUC) = 0.82, 95% CI 0.72–0.90); 6.82 (AUC = 0.65, 95% CI 0.53–0.75); 19.06 (AUC = 0.77, 95% CI 0.64–0.87), respectively. Further ROC analysis was then performed and revealed that the criteria of increased expression (ΔCT below the optimal cutoff) of any two of these three lncRNAs in diagnosing ME/CFS had a sensitivity of 67.4, and specificity of 86.7, AUC = 0.82, with 95% CI 0.71–0.90 (Fig. 3e).
Expression levels of genes potentially regulated by the lncRNAs in ME/CFS
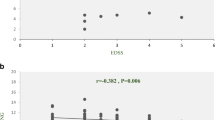
In order to elucidate potential mechanisms of actions of lncRNAs NTT, MIAT and EMX2OS in ME/CFS, we analyzed the expression profile of their potential downstream genes in patients and controls. NTT has been suggested to act on nearby genes, including IFNGR1 and PBOV1 [12]. ZEB1 has been reported to be downstream of MIAT; and EMX2 has been proposed to be regulated by EMX2OS [21, 27]. While EMX2 could not be detected in PBMCs (data not shown), ZEB1 level was found to have a mild but significant elevation in ME/CFS PBMCs as compared with controls (ME/CFS median ΔCT vs. control median ΔCT:7.60 vs. 7.82, p < 0.05, Fig. 4a). The expression levels of IFNGR1 and PBOV1 did not show significant differences between ME/CFS and controls (Fig. 4b, c). As for the correlation between the expressions of lncRNAs and downstream genes, a positive relationship between ZEB1 and MIAT level was detected using linear regression analysis in ME/CFS (r2= 0.47, p < 0.0001), but not in controls (Fig. 5a, b). Similarly, IFNGR1 level was found to be positively correlated with NTT level in ME/CFS (r2= 0.46, p < 0.0001), but not in controls (Fig. 5c, d). The linear regression analysis between NTT and PBOV1 showed no statistical significance both in ME/CFS and controls (Fig. 5e, f).

Expression level of genes potentially regulated by MIAT (a) or NTT (b, c) in PBMCs of ME/CFS patients and controls. *p < 0.05 by Mann–Whitney U test, lines represent medians

Correlation between lncRNA and downstream gene expressions. Correlation between MIAT and ZEB1 levels in ME/CFS (a) and in controls (b). Correlation between NTT and IFNGR1 in ME/CFS (c) and in controls (d). Correlation between NTT and PBOV1 levels in ME/CFS (e) and in controls (f)
Upregulation of NTT and MIAT by H2O2 and poly (I:C) in human monocytic and glioma cell lines
Oxidative stress and recurrent herpes viral infection are known to contribute to ME/CFS pathology [28]. To investigate the potential effect of these stressors on the expression of NTT, MIAT and EMX2OS, we used human monocytic cell line THP-1 and glioma cell line KALS1 as models, and treated them with titrations of H2O2 (oxidative stress) or poly (I:C) (TLR3 agonist, mimicking herpes viral infection) for 6 h. While NTT and MIAT were upregulated after stimulations, EMX2OS could not be detected in both cell lines. In THP-1 cells, NTT expression level increased to a mean of 1.47- and 3.06-fold after stimulation with 10 nM H2O2 and 100 μM poly (I:C), respectively (Fig. 6a). Furthermore, MIAT level increased to a mean of 1.26- and 3.38-fold after THP-1 stimulation with 10 nM H2O2 and 100 μM poly (I:C), respectively (Fig. 6b). Higher expression levels of the potential NTT downstream gene PBOV1 and the potential MIAT downstream gene ZEB1 were also detected in THP-1 after stimulations with 10 nM H2O2 and 100 μM poly (I:C), respectively; however, the level of IFNGR1 (another potential NTT downstream gene) did not show an obvious change after both stimulations (Fig. 6c–e). In KALS1 cells, NTT could be upregulated to a mean of 1.49-fold when H2O2 was titrated to 100 μM, and NTT level also increased to a mean of 1.65-fold after 50 μM poly (I:C) treatment (Fig. 6f). As for MIAT expression in KALS1, a 1.26-fold and a 1.31-fold increase were observed after 6 h 10 nM H2O2 and 100 μM poly (I:C) stimulations, respectively (Fig. 6g). The expression levels of potential NTT and MIAT downstream genes after H2O2 and poly (I:C) stimulations in KALS1 showed a similar pattern as those observed in THP-1 (Fig. 6h–j).

Upregulation of NTT and MIAT and potential downstream genes after peroxide or poly (I:C) stimulation in THP-1 and KALS1 cells compared to unstimulated cells. The expression levels of NTT (a) and MIAT (b) in THP-1 after 6 h stimulations. N = 6, bars represent mean ± SEM. The expression levels of IFNGR1 (c) (N = 6), PBOV1 (d) (N = 3) and ZEB1 (e) (N = 6), bars represent mean ± SEM. The expression levels of NTT (f), MIAT (g), IFNGR1 (h), PBOV1 (i) and ZEB1 (j) in KALS1 after 6 h stimulations. N = 3, bars represent mean ± SEM
Discussion
Emerging roles of very large (> 5 kb) lncRNAs in immune regulation and disease processes are being discovered [11]. Here we report the expression signature of ten lncRNAs NTT, NEAT1, MALAT1, TUG1, MIAT, His-1 RNA, GNAS1-AS, EMX2OS, CR933609 and AK124742 in PBMCs derived from ME/CFS patients. We have selected this screening panel according to their potential regulation of immune, stress, metabolic and neurologic responses, which have been hypothesized to be involved in ME/CFS pathogenesis. Nevertheless it is remarkable to find out on PCA plot that ME/CFS could be differentiated from healthy controls via this lncRNA profile. Among the ten lncRNAs, NTT, MIAT and EMX2OS expression levels explained the most variance between ME/CFS and controls. Further supporting their potential role in disease pathomechanism, higher NTT and EMX2OS levels were associated with more severe ME/CFS (Bell score < 30). Using expression of any two of these three lncRNAs (NTT, MIAT and EMX2OS) in discriminating ME/CFS from healthy had an AUC of 0.82 on ROC curve, suggesting a potential diagnostic value of these lncRNAs for ME/CFS.
Consistent with the hypothesis that disease pathology in ME/CFS could be driven by oxidative stress and viral infections, we found that NTT and MIAT levels in THP-1 and KALS1 cell lines could be increased after H2O2 or poly (I:C) stimulations, an expression pattern similar to those found in ME/CFS patients’ PBMCs. However, we do not know yet whether this lncRNA profile found in our study is specific for ME/CFS or can be found in other diseases involving immune dysregulation or oxidative stress, such as autoimmune diseases and cancer. It has been reported that MIAT levels could be upregulated in high glucose conditions and in lung cancer, and NTT expressions might be found in processes involving T cell activation [12, 17, 29]. Further comparing the PBMC expression signature of NTT, MIAT, and EMX2OS in ME/CFS with patients suffering from chronic fatigue due to autoimmune diseases or cancer is important to assess the lncRNA test specificity in diagnosing ME/CFS.
The mechanisms of NTT, MIAT and EMX2OS in ME/CFS pathogenesis require further investigations. In our study, we detected an association of higher level of ZEB1, a MIAT-regulated gene, with ME/CFS. Consistently, both THP-1 and KALS1 cell lines showed higher expression levels of MIAT and ZEB1 after stimulation of poly (I:C), a synthetic analog of double strand RNA representing active viral infection, a potential trigger of ME/CFS. Zinc finger E-box-binding protein (ZEB) 1 has been reported to be a transcription factor recruiting repressor complex to suppress IL-2 activation in T cells [30]. Upregulation of ZEB1 might be associated with response to chronic inflammation in ME/CFS. In non-small-cell lung cancer cell line, knockdown of MIAT resulted in decreased ZEB1 expression, indicating cis-action of MIAT on regulating ZEB1 [27]. Interestingly MIAT is involved in endothelial dysfunction as well, which is frequently observed in ME/CFS [17, 31]. For NTT, it has been proposed to exert its function on nearby genes due to its large size (17 kb) [12]. Several genes involving cell proliferation, apoptosis or inflammation, including IFNGR1, PBOV1, TNFAIP3, HIVEP2, BCLAF1 and MYB are located close to the chromosome position of NTT [12]. We found no significant difference in IFNGR1 and PBOV1 expressions between ME/CFS and controls. However, a marked positive correlation between NTT and IFNGR1 levels were observed in ME/CFS, not in controls. This observation suggests that the NTT/IFNGR1 axis might play a subtle role in ME/CFS pathogenesis. Whether other downstream genes are affected by the upregulated NTT in ME/CFS and possibly play more important roles in disease pathobiology needs more experiments. Finally, according to lncrnadb, the lncRNA database, the expression level of EMX2OS under normal physiological condition is higher in brain, moderate in lymph nodes, and very low in leukocytes. Consistent with this, we could not detect EMX2OS in PBMCs of several individuals. However, most ME/CFS patients were found to have elevated EMX2OS in their PBMCs. The role of EMX2OS in PBMC is currently unclear, and the potential downstream gene EMX2 usually expressed in CNS could not be detected in all study subjects [22]. Further EMX2OS overexpression experiment in THP-1 is underway in our lab to answer this question. Interestingly, EMX2 upregulation was found in brain hypoxemia [32]. ME/CFS patients have broad decreases in cerebral blood flow, which may result in hypoxemia [33].
In addition to our study on using lncRNA signature as diagnostic marker for ME/CFS, profiles of blood mRNA expression and plasma metabolites have been suggested to show diagnostic values for ME/CFS [6, 7, 34]. As described by Kerr et al., there were 88 differentially expressed genes in ME/CFS required for diagnostic and prognostic grouping [34]. Furthermore, Naviaux et al. proposed an eight-metabolite set for diagnosing male ME/CFS, and a thirteen-metabolite set for diagnosing female ME/CFS [6]. Our results showed that a lncRNA expression panel composed of as few as three very large lncRNAs (NTT, MIAT, EMX2OS) may achieve a good diagnostic value and provide information about ME/CFS disease severity (NTT, MIAT).
Conclusion
In conclusion, although the pathogenic mechanisms of very large lncRNAs in ME/CFS remains to be elucidated, we have first evidence that a lncRNA expression signature could be of diagnostic value in ME/CFS.
References
Wadman M. For chronic fatigue syndrome, a ‘shifting tide’ at NIH. Science. 2016;354(6313):691–2.
Yang TY, Kuo HT, Chen HJ, Chen CS, Lin WM, Tsai SY, Kuo CN, Kao CH. Increased risk of chronic fatigue syndrome following atopy: a population-based study. Medicine (Baltimore). 2015;94(29):e1211.
Carruthers BMJA, De Meirleir KL, Peterson DL, Klimas NG, Lemer AM, Bested AC, Flor-Henry P, Joshi P, Powles ACP, Sherkey JA, van de Sande MI. Myalgic encephalomyelitis/chronic fatigue syndrome: clinical working case definition, diagnostic and treatment protocols (Canadian case definition). J Chron Fatigue Synd. 2003;11(1):7–115.
Morris G, Maes M. Oxidative and nitrosative stress and immune-inflammatory pathways in patients with myalgic encephalomyelitis (ME)/chronic fatigue syndrome (CFS). Curr Neuropharmacol. 2014;12(2):168–85.
Loebel M, Grabowski P, Heidecke H, Bauer S, Hanitsch LG, Wittke K, Meisel C, Reinke P, Volk HD, Fluge O, Mella O, Scheibenbogen C. Antibodies to beta adrenergic and muscarinic cholinergic receptors in patients with chronic fatigue syndrome. Brain Behav Immun. 2016;52:32–9.
Naviaux RK, Naviaux JC, Li K, Bright AT, Alaynick WA, Wang L, Baxter A, Nathan N, Anderson W, Gordon E. Metabolic features of chronic fatigue syndrome. Proc Natl Acad Sci USA. 2016;113(37):E5472–80.
Yamano E, Sugimoto M, Hirayama A, Kume S, Yamato M, Jin G, Tajima S, Goda N, Iwai K, Fukuda S, Yamaguti K, Kuratsune H, Soga T, Watanabe Y, Kataoka Y. Index markers of chronic fatigue syndrome with dysfunction of TCA and urea cycles. Sci Rep. 2016;6:34990.
Geisler S, Coller J. RNA in unexpected places: long non-coding RNA functions in diverse cellular contexts. Nat Rev Mol Cell Biol. 2013;14(11):699–712.
Jiang C, Li X, Zhao H, Liu H. Long non-coding RNAs: potential new biomarkers for predicting tumor invasion and metastasis. Mol Cancer. 2016;15(1):62.
Wu GC, Pan HF, Leng RX, Wang DG, Li XP, Li XM, Ye DQ. Emerging role of long noncoding RNAs in autoimmune diseases. Autoimmun Rev. 2015;14(9):798–805.
Geng H, Tan XD. Functional diversity of long non-coding RNAs in immune regulation. Genes Dis. 2016;3(1):72–81.
Liu AY, Torchia BS, Migeon BR, Siliciano RF. The human NTT gene: identification of a novel 17-kb noncoding nuclear RNA expressed in activated CD4+ T cells. Genomics. 1997;39(2):171–84.
Zhang F, Wu L, Qian J, Qu B, Xia S, La T, Wu Y, Ma J, Zeng J, Guo Q, Cui Y, Yang W, Huang J, Zhu W, Yao Y, Shen N, Tang Y. Identification of the long noncoding RNA NEAT1 as a novel inflammatory regulator acting through MAPK pathway in human lupus. J Autoimmun. 2016;75:96–104.
Zhao G, Su Z, Song D, Mao Y, Mao X. The long noncoding RNA MALAT1 regulates the lipopolysaccharide-induced inflammatory response through its interaction with NF-kappaB. FEBS Lett. 2016;590(17):2884–95.
Su S, Liu J, He K, Zhang M, Feng C, Peng F, Li B, Xia X. Overexpression of the long noncoding RNA TUG1 protects against cold-induced injury of mouse livers by inhibiting apoptosis and inflammation. FEBS J. 2016;283(7):1261–74.
Tang W, Shen Z, Guo J, Sun S. Screening of long non-coding RNA and TUG1 inhibits proliferation with TGF-beta induction in patients with COPD. Int J Chron Obstruct Pulmon Dis. 2016;11:2951–64.
Yan B, Yao J, Liu JY, Li XM, Wang XQ, Li YJ, Tao ZF, Song YC, Chen Q, Jiang Q. lncRNA-MIAT regulates microvascular dysfunction by functioning as a competing endogenous RNA. Circ Res. 2015;116(7):1143–56.
Liao J, He Q, Li M, Chen Y, Liu Y, Wang J. LncRNA MIAT: myocardial infarction associated and more. Gene. 2016;578(2):158–61.
Li J, Witte DP, Van Dyke T, Askew DS. Expression of the putative proto-oncogene His-1 in normal and neoplastic tissues. Am J Pathol. 1997;150(4):1297–305.
Hayward BE, Bonthron DT. An imprinted antisense transcript at the human GNAS1 locus. Hum Mol Genet. 2000;9(5):835–41.
Noonan FC, Goodfellow PJ, Staloch LJ, Mutch DG, Simon TC. Antisense transcripts at the EMX2 locus in human and mouse. Genomics. 2003;81(1):58–66.
Spigoni G, Gedressi C, Mallamaci A. Regulation of Emx2 expression by antisense transcripts in murine cortico-cerebral precursors. PLoS ONE. 2010;5(1):e8658.
Chang CC, Liu TY, Lee YT, Chen YC, Yeh KT, Lee CC, Chen YL, Lin PC, Chang YS, Chan WL, Liu TC, Chang JG. Genome-wide analysis of lncRNAs in 3′-untranslated regions: CR933609 acts as a decoy to protect the INO80D gene. Int J Oncol. 2018;53:417–33.
Yao W, King DA, Beckwith SL, Gowans GJ, Yen K, Zhou C, Morrison AJ. The INO80 complex requires the Arp5-Ies6 subcomplex for chromatin remodeling and metabolic regulation. Mol Cell Biol. 2016;36(6):979–91.
Li J, Cao Y, Xu X, Xiang H, Zhang Z, Chen B, Hao Y, Wei Z, Zhou P, Chen D. Increased new lncRNA–mRNA gene pair levels in human cumulus cells correlate with oocyte maturation and embryo development. Reprod Sci. 2015;22(8):1008–14.
Narayanaswamy PB, Hodjat M, Haller H, Dumler I, Kiyan Y. Loss of urokinase receptor sensitizes cells to DNA damage and delays DNA repair. PLoS ONE. 2014;9(7):e101529.
Zhang HY, Zheng FS, Yang W, Lu JB. The long non-coding RNA MIAT regulates zinc finger E-box binding homeobox 1 expression by sponging miR-150 and promoteing cell invasion in non-small-cell lung cancer. Gene. 2017;633:61–5.
Sotzny F, Blanco J, Capelli E, Castro-Marrero J, Steiner S, Murovska M, Scheibenbogen C, European Network on MC. Myalgic encephalomyelitis/chronic fatigue syndrome—evidence for an autoimmune disease. Autoimmun Rev. 2018;17(6):601–9.
Lai IL, Yang CA, Lin PC, Chan WL, Lee YT, Yen JC, Chang YS, Chang JG. Long noncoding RNA MIAT promotes non-small cell lung cancer proliferation and metastasis through MMP9 activation. Oncotarget. 2017;8(58):98148–62.
Wang J, Lee S, Teh CE, Bunting K, Ma L, Shannon MF. The transcription repressor, ZEB1, cooperates with CtBP2 and HDAC1 to suppress IL-2 gene activation in T cells. Int Immunol. 2009;21(3):227–35.
Newton DJ, Kennedy G, Chan KK, Lang CC, Belch JJ, Khan F. Large and small artery endothelial dysfunction in chronic fatigue syndrome. Int J Cardiol. 2012;154(3):335–6.
Tonchev AB, Yamashima T. Differential neurogenic potential of progenitor cells in dentate gyrus and CA1 sector of the postischemic adult monkey hippocampus. Exp Neurol. 2006;198(1):101–13.
Biswal B, Kunwar P, Natelson BH. Cerebral blood flow is reduced in chronic fatigue syndrome as assessed by arterial spin labeling. J Neurol Sci. 2011;301(1–2):9–11.
Kerr JR, Petty R, Burke B, Gough J, Fear D, Sinclair LI, Mattey DL, Richards SC, Montgomery J, Baldwin DA, Kellam P, Harrison TJ, Griffin GE, Main J, Enlander D, Nutt DJ, Holgate ST. Gene expression subtypes in patients with chronic fatigue syndrome/myalgic encephalomyelitis. J Infect Dis. 2008;197(8):1171–84.
Authors’ contributions
CAY and CS conceived the study and organized the project in collaboration with JGC. CS was responsible for the diagnosis and patient recruitment. CAY and CS analyzed data and wrote the manuscript. CAY, SB, YCH and FS performed the experiments. All authors read and approved the final manuscript.
Acknowledgements
We thank Dr. Woei-Fuh Wang for statistical consulting.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Consent for publication
Our article does not contain individual personal data.
Ethics approval and consent to participate
This study was approved by the Ethics Committee of Charité Universitätsmedizin Berlin in accordance with the 1964 Declaration of Helsinki and its later amendments and all patients gave written informed consent.
Funding
This work was supported by the following grants. CAY received a Grant from the Ministry of Science and Technology (MOST), Taiwan, Grant 106-2314-B-039-047-MY3; CS received a Grant from the Lost Voices Foundation e.V., Germany, to support this work.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Yang, CA., Bauer, S., Ho, YC. et al. The expression signature of very long non-coding RNA in myalgic encephalomyelitis/chronic fatigue syndrome. J Transl Med 16, 231 (2018). https://doi.org/10.1186/s12967-018-1600-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12967-018-1600-x