
Abstract
The phenomenon of phase separation is quite common in cells, and it is involved in multiple processes of life activities. However, the current research on the correlation between protein modifications and phase separation and the interference with the tendency of phase separation has some limitations. Here we focus on several post-translational modifications of proteins, including protein phosphorylation modification at multiple sites, methylation modification, acetylation modification, ubiquitination modification, SUMOylation modification, etc., which regulate the formation of phase separation and the stability of phase separation structure through multivalent interactions. This regulatory role is closely related to the development of neurodegenerative diseases, tumors, viral infections, and other diseases, and also plays essential functions in environmental stress, DNA damage repair, transcriptional regulation, signal transduction, and cell homeostasis of living organisms, which provides an idea to explore the interaction between novel protein post-translational modifications and phase separation.
Video Abstract
Similar content being viewed by others
Introduction
In recent years, the phase separation of biological macromolecules has been recognized as a prevalent biological phenomenon, which plays important functions in development, environmental stress, DNA damage repair, transcriptional regulation, signaling, and cellular homeostasis of living organisms through its physicochemical properties. As research progressed, it was discovered that protein post-translational modifications (PTM) play an important role in forming and regulating phase separation. Collecting, organizing, mining, and analyzing related data have become research hotspots in the physiological process and the occurrence and development of diseases. Here, we introduce the formation of phase separation, the factors affecting it and the regulation of phase separation mediated by PTMs of proteins, summarize the current research results on PTM of proteins related to eukaryotic phase separation, and look forward to the future development direction, and provide reference and clues for further experimental studies to explore the regulation of protein PTMs on phase separation.
The formation of phase separation and the factors affecting phase separation
Concept and formation of phase separation
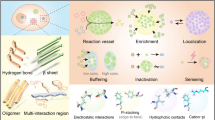
In some external conditions change, some of the molecules in the multi-component system will be enriched to a specific region of the system, the separation of several phases with different components and structures of the process is called phase separation [1]. In 2009, Brangwynne et al. in their study of the mechanisms of fertilized egg polarization and asymmetric division in Caenorhabditis elegans showed that P granules exhibit liquid-like phase change behavior, including fusion, dropping, and wetting [2]. This is the first application of phase separation theory to the field of biology, providing a new perspective for the in-depth study of the physicochemical mechanisms underlying the formation and function of membrane-free functional compartments in cells. Phase separation includes gas-liquid phase separation, [3] liquid-liquid phase separation(LLPS), solid-liquid phase separation, [4] and so on. In biology, LLPS is the most common. LLPS is the creation, under certain conditions, of two separate phases of mutually soluble components in a solution that are constantly exchanged at the phase boundary: a polymer-poor phase and a polymer-rich phase contained therein (Fig. 1) [5, 6]. The condensed phase formed by LLPS helps the complex biochemical reactions within the cell to be able to proceed in an orderly manner in different spaces [7]. The liquid phase state in which intracellular phase separation predominantly exists is usually dominated by surface tension, [8] which gives droplets a spherical shape, small droplets easily fuse to form larger droplets. In certain cases, phase separation within cells can lead to gel-like or even solid-like aggregates (Fig. 1) [9]. Not all intracellular proteins can undergo phase separation; the presence of intrinsically disordered regions (IDR), modular domains, or biomolecules capable of inter- or intra-molecular multivalent interactions with other biomolecules is necessary for phase separation to occur [10,11,12,13]. When the molecular solubility in the solution is below the threshold value for the occurrence of phase separation, the molecules can remain dispersed in the solution [8, 14]. When the molecular solubility in the solution exceeds the critical concentration for phase separation, a highly aggregated phase with distinct boundaries and a high degree of motility is precipitated from the solution (Fig. 1) [8, 14].

The formation process of LLPS and factors affecting LLPS
Factors affecting the occurrence of phase separation
The most direct factors affecting the occurrence of phase separation depend on the concentration of macromolecules and solutions and their physicochemical properties; [15] In addition, the formation and regulation of phase separation are very sensitive to changes in environmental states, including temperature, ionic strength, pH, as well as the passivator and crowding effects (Fig. 1) [5, 16,17,18,19]. For example, the phase separation of the nucleocapsid protein (N-protein) of coronaviruses is an environment-dependent process in which changes in protein concentration, salt ion, and pH concentration can affect the process by which phase separation occurs [20].
Direct adjustment of the critical concentration for phase separation or the concentration of biomolecules in the solution can directly affect the occurrence of phase separation [20]. For example, under low salt conditions in vitro, the short isoform of BRD4 protein (BRD4S) (1-719) formed LLPS droplets at 0.5 µM protein concentration. As the protein concentration of BRD4S(1-719) increased, the number of droplets formed for phase separation and the total area of the droplets increased accordingly, which indicates that the LLPS formed by BRD4S(1-719) under in vitro conditions is concentration dependent [21]. However, under physiological conditions, molecules are not isolated but exist in a crowded complex environment rich in multicomponent abundant proteins, so other competing proteins may interfere with IDR interactions [14]. The lower the critical concentration, the easier it is for the molecules to promote the formation of phase separation through interaction forces [19]. The effect of temperature on phase separation varies depending on the entropy effect system [17]. For phase separation systems with reduced entropy, increasing the temperature promotes phase separation depolymerization, [17, 22] examples include germ granule protein, bovine lens γB-crystallin, and so on [23, 24]. Conversely, for phase separation systems with increased entropy, elevated temperature promotes the formation of phase separation, [17] such as the N-protein of coronaviruses, hydrophobic elastin, and the Alzheimer’s-related protein Tau [20, 25, 26]. Salt ion concentration plays a key role in the formation and regulation of phase separation systems, and increasing ionic strength in phase separation systems where electrostatic interactions dominate weakens electrostatic interactions and is detrimental to the occurrence of phase separation [26]. For example, it has been shown that high salt inhibits the occurrence of tripartite motif-containing protein 21 (TRIM21)-driven LLPS phenomena in which electrostatic interaction forces are the primary mediators of this process [27]. For phase separation systems where hydrophobic interactions dominate increasing the ionic strength reduces the critical concentration at which phase separation of proteins occurs and facilitates the occurrence of phase separation [6, 10]. The effect of pH on phase separation is mainly reflected in the modulation of the occurrence of phase separation by changing the protonation state of amino acids, i.e., by changing the charged properties of amino acids and thus affecting the multivalent intermolecular interactions [25, 28]. In addition to this, the addition of bovine serum albumin (BSA) and protein crowding reagents such as polyethylene glycol (PEG), ficoll, and dextran can mimic the intracellular environment and facilitate the onset of phase separation [29,30,31]. For example, PEG8000 was added to purified fused in sarcoma (FUS) (1-180) as a macromolecular crowding agent to induce phase separation droplet formation [31].
PTMs of proteins
Overview of protein PTMs
Protein PTMs is a covalent modification that occurs in the backbone or side chain of a protein after translation and may alter protein physicochemical properties, spatial conformation, stability, cellular localization, and interactions with other proteins, greatly enriching the diversity of protein structures and functions [32,33,34]. There are more than 500 known PTMs of proteins, and the common ones include phosphorylation, acetylation, glycosylation, ubiquitination, SUMOylation, and methylation modifications [35]. Protein PTMs are involved in almost all normal life activity processes of cells, greatly enriching the complexity of the organism’s proteome and playing vital biological functions in physiological, pathological, or disease treatment processes. Intrinsically disordered proteins (IDPs) are highly exposed to PTMs as one of the drivers of phase separation [36, 37]. For example, tau is closely associated with a variety of PTMs, such as phosphorylation, ubiquitination, and acetylation. These PTMs enhance or attenuate the LLPS of tau [38,39,40]. Furthermore, previous studies on N-terminal DDX4 have shown that methylation reduces its LLPS [23].
Correlation between PTMs and phase separation
PTMs of proteins, including protein phosphorylation modifications, acetylation modifications, methylation modifications, ubiquitination modifications, and SUMO modifications at multiple sites, can affect the charge properties of the modified amino acid residues and charged sequences, change the size of biomolecules, generate new binding sites or change the local structure and conformation, and promote intermolecular multivalent interactions by mediating intermolecular charge-charge, π-π, and cation-π interactions [20, 35, 41, 42], which may regulate the formation of phase separation and the stability of phase separation structures [43].
In addition, phase-separated structures are also involved in regulating protein modifications, such as the cyclin T1 of the positive transcription elongation factor (P-TEFb), which targets RNA polymerase (Pol)II to the phase-separated droplets, and thus contributes to the hyperphosphorylation of the C-terminal domain (CTD) of the RPB1 subunit of human PolII [44]. In addition, it was demonstrated that Arabidopsis blue light receptor CRY2 and transcription factor TCP22 can form a blue light-dependent phase separation, which in turn recruits proteins such as photo regulatory protein kinases (PPKs) into the phase and regulates CCA1 gene expression through various biochemical processes such as protein phosphorylation modification [45]. In 2020, it was demonstrated that the conserved yeast E3 ubiquitin ligase Bre1 binds to the scaffold protein Lge1 and undergoes phase separation through multivalent interactions and that this layered fluid recruits Rad6 and nucleosome substrates to form spatially organized “reaction chambers” that accelerate the ubiquitination of H2B [46]. In 2021, a study found that paraspeckle component 1(PSPC1) can promote the phosphorylation of cyclin-dependent kinase 1 (Cdk1) (Ser-345) by driving the onset of phase separation to recruit the phosphatase serine/threonine-protein phosphatase 5 (PPP5C) in mouse oocyte maturation [47]. In 2022, Dajun Sang et al. demonstrated that MAPK3 and FUS3, as well as Cdk1, are recruited by phase separation into multiple types of synthetic condensates formed by multivalent “scaffolds” in which their phosphorylation is increased [48].
PTMs of proteins related to eukaryotic phase separation
Based on the excavation of phase separation protein structures and an increasing number of protein modification sites, the linkage between phase separation and protein modification processes has been successively discovered and caused a research boom in the field in the past decade (Table 1). In light of the results of the previous review by Owen I et al., [49] the following section will further review the latest insights on protein PTMs associated with phase separation to help us systematically understand the current common linkage between phase separation and protein PTMs.
Phosphorylation modifications
Phosphorylation, one of the most common PTMs of proteins, is an esterification reaction that attaches phosphoryl groups to the hydroxyl groups of the Ser, Thr, and Tyr side chains in proteins and can regulate protein interactions [64, 100]. The effect of phosphorylation on the phase separation of biomolecules is twofold; the addition of phosphate groups can positively or negatively control the formation of phase separation by changing the protein charge [53, 100, 101]. This regulatory role is closely linked to the development of neurodegenerative diseases, [31] tumors, [21] viral infections, [59] and other diseases caused by abnormal phase segregation [73]. In addition to this, it also plays essential functions in environmental stress, [71] DNA damage repair, [50,51,52] transcriptional regulation, [63] signal transduction, [62] and cell homeostasis of living organisms (Fig. 2) [102].

Crosstalk between PTMs and LLPS in the physiopathological process. The large brown semi-circular arrow denotes the illness circle’s ability to rotate, much like a turntable, which we call the “turntable model” here. Crosstalk between PTMs and LLPS has been implicated in numerous areas of illness and biological function, i.e., neurodegenerative diseases, tumors, viral infections, environmental stress, DNA damage repair, transcriptional regulation, signal transduction, and cell homeostasis of living organisms. Furthermore, each PTM is linked to a different disease or biological function, and vice versa
Negative modulation of phase separation by phosphorylation modifications
LLPS has a key role in the development of related neurodegenerative diseases. Given that previous studies noted that phosphorylation of the low-complexity structural domain (LC/LCD) of FUS prevents hydrogel retention [103], Murray DT et al. similarly concluded that phosphorylation of core Ser and Thr residues by DNA-dependent protein kinases prevents binding of FUS-LC hydrogels, leading to phase-separated liquid-like FUS- cleavage of LC droplets [50]. In addition, phosphorylation modifications and phosphomimetic mutations in the FUS low-complexity structural domains were able to reduce the aggregation or liquid-solid transition that occurs in LLPS [101]. However, the phosphomimetic substitution inhibited transient intramolecular and intermolecular LC contacts, disrupting transient intramolecular collapse and intermolecular interactions, providing valuable information for understanding how phosphorylation inhibits LLPS [101]. Ding XF et al. demonstrated that Ser61 site-specific phosphorylation modifications disrupt intramolecular and intermolecular interactions, providing important theoretical support for a role in blocking R2-induced conversion of FUS aggregates to pathological aggregates [31]. Furthermore, phosphorylation of FUS LCD during DNA damage may antagonize FUS phase separation [50,51,52]. The inhibitory effect of phosphorylation in FUS-LC on FUS LLPS and aggregation is now well known. Notably, the mechanism by which phosphorylation at the atomic level disrupts FUS LLPS and aggregation was first revealed by Lao ZH et al. in 2022 [104].
Some neurodegenerative diseases are associated with abnormal aggregation of TAR DNA binding protein-43 (TDP-43). Phosphomimetic substitutions at Ser48 were found to disrupt TDP-43 polymer assembly [53], and casein kinase 1δ mediated TDP-43 hyperphosphorylation or C-terminal phosphomimetic mutations reduced TDP-43 phase separation and aggregation [54]. Interestingly, Tariq A et al. found that phosphomimetic mutations at the Thr499 and Ser535 positions in the Hsp104 middle domain (MD) enabled Hsp104 to rescue the phase-separation-related aggregation and toxicity of TDP-43, FUS, and α-synuclein in yeast [55]. This regulation could provide a strategy for the treatment of destructive neurodegenerative diseases by designing enhanced Hsp104 disaggregase activity.
Recently phosphorylation has been gradually tapped into regulating the cytoskeleton assembly associated with phase separation. Phosphorylation at Ser262 of the K18 protein in the microtubule-binding domain (MTBD) of tau disrupts binding to microtubulin as assessed by microtubule protein aggregation assay [56]. Furthermore, Savastano A et al. found that LLPS of tau associated with Alzheimer’s disease recruited microtubulin to droplets and promoted microtubule assembly, while phosphorylation at Thr231 disrupted the binding of the proline-rich region P2 of tau to microtubulin and blocked the assembly process [39].
The Ryan VH research team found that tyrosine phosphorylation of the neuropathy-associated heterogeneous nuclear ribonucleoprotein (hnRNPA2) reduced hnRNPA2 phase separation and reduced aggregation of hnRNPA2 disease variants [57].
In recent years, in addition to the numerous studies and research in neurodegenerative diseases, the negative regulatory effects of phosphorylation modifications on phase separation have been gradually uncovered in the following areas. BRD4 protein is closely related to the development of various tumors, and a study in 2020 showed that phosphorylation of p-BRD4S led to a change in the distribution of charge on the surface of protein molecules, which reorganized the structure of protein molecules, thereby inhibiting the formation of LLPS under in vitro conditions and prompting the inactivation of gene transcription, [21] providing new ideas for the development of targeted anticancer drugs. In the context of viral infection, Carlson CR et al. found that unmodified SARS-CoV-2 N-proteins formed ordered gel-like condensates through multivalent RNA-protein and protein-protein interactions, whereas N-protein phosphorylation reduced the interactions and consequently transformed them into more liquid-like droplets, [59] suggesting the effect of phosphorylation on the dual functions of N-proteins in different oligomeric states. Phase segregation of barley yellow striate mosaic virus (BYSMV) phosphoprotein (P) in plants could be inhibited by host CK1-dependent phosphorylation of the intrinsically disordered P region [58]. In terms of signaling, TAZ is an important effector downstream of the Hippo signaling pathway, and its phase separation phenomenon is directly influenced by phosphorylation. When phosphorylated by the Hippo signaling pathway kinase LATS2, the TAZ phase separation phenomenon is inhibited, which prevents the recruitment of transcriptional regulators into the phase change compartment and affects downstream target gene expression. This is the first molecule to elucidate that signaling pathway-specific molecules can signal by way of LLPS, revealing a novel mechanism for phase separation to promote transcription and providing a new direction for the development of disease-therapeutically relevant interventions [62]. In terms of transcriptional regulation, human p53 is a transcription factor that regulates the transcription of multiple target genes. p53 phosphorylation was found by Dai ZJ et al. to promote p53 incorporation into transcriptional complex droplets and inhibit p53-mediated transcriptional droplet aggregation, revealing that p53 phosphorylation alters its LLPS behavior [63]. Lenard AJ et al. found that serine-arginine protein kinase 1 (SRPK1)-mediated phosphorylation of the RG/RGG region of cold-inducible RNA binding protein (CIRBP) impairs LLPS in vitro, which may affect RNA stability and regulate gene expression and cellular function [65]. In the functional homeostasis of cells, the generation of membraneless organelles by LLPS is widely recognized as the mechanism by which many disordered biomolecules within cells can order their physicochemical reactions, which is important for organizing cellular activities. It has been shown that changes in the phosphorylation state of peptides reversibly regulate the liquid organelle model of biomolecules, supporting the key role of phosphorylation modifications in dynamically regulating the liquid intracellular compartments generated by phase separation [102]. For example, LLPS of NBDY microproteins is thought to potentially regulate the formation of membrane-free organelles, the phosphorylation of NBDY drives the dissociation of these droplets [64]. In addition, the negative regulatory effect of phosphorylation on phase separation was confirmed in studies of the Intrinsically disordered plant protein PARCL and the vertebrate CPEB protein [67, 68].
Positive regulation of phase separation by phosphorylation modifications
Multiple disease processes were able to discover that phosphorylation modifications can also promote phase separation through intermolecular interactions. In Alzheimer’s studies, it was determined that MARK2 phosphorylation promotes LLPS of the Tau repeat structural domain [25]. Subsequently, by using high molecular weight hyperphosphorylated tau isolated from human Alzheimer’s brain, post-translationally modified recombinant tau was observed to form tau droplets in neurons in vitro, [105] providing strong evidence that phosphorylation promotes phase separation of full-length tau. In a study of the neuromuscular disease spinal muscular atrophy (SMA), Schilling M et al. found that the Ser49 and Ser63 phosphorylation control complexes of the human survival motor neuron protein (SMN) coalesce in Cajal [69]. In a study of fragile X syndrome, TsangB et al. experimentally demonstrated that phosphorylation increased the local negative charge density of fragile X mental retardation protein (FMRP), suggesting that phosphorylation of the low-complexity disordered region at the C-terminus of FMRP may increase its phase separation by increasing the propensity for multivalent electrostatic interactions [73]. The human selective autophagy receptor protein p62 was found to be enriched in cellular inclusion bodies of various human diseases, and phosphorylation of its Ser403 site promotes the binding of p62 protein to ubiquitin chains, thereby facilitating the phase separation of p62 protein to form p62 bodies, [75] providing a theoretical basis for studying the formation of other membrane-free structures in the field of autophagy and the mechanisms of related diseases.
In terms of gene regulation, phosphorylation of the N-terminal extension of human heterochromatin protein 1 (HP1α) associated with heterochromatin gene silencing promotes phase separation formation [70]. Site-specific protein modifications regulate the onset of phase separation, and notably other point mutations do not affect droplet formation, a process confirmed in a study in which Ser760 of Liprin-α3 eliminated Liprin-α3 phosphorylation by a single point mutation [72]. As scientists continue to develop and advance in the field of phase separation mechanisms, in 2022 HerC et al. further demonstrated the importance of electrostatic interactions due to HP1α phosphorylation in LLPS, reporting that phosphorylation redistributes the charge in the protein sequence and enhances the interaction between the NTE and hinge regions. At the same time, it provides an additional negative charge, introduces repulsive forces between other regions of the protein, and leads to protein conformational expansion at high concentrations, among other important mechanisms [42].
Environmental stress-induced changes in phosphorylation also affect the state of phase separation. Under heat stress conditions, mTORC1-mediated phosphorylation of PGL-1/-3 is elevated in P-particle components, and PGL-1/-3 undergoes accelerated phase separation to form PGL particles resistant to autophagic degradation, [71] which can maintain embryonic viability during heat stress, suggesting a protective role of phosphorylation modifications regulating phase separation in the organism under stress conditions. In 2020, Liu ZY’s team demonstrated that stress-induced phosphorylation of Hsp27 impairs the inhibitory activity of Hsp27 in LLPS of FUS-LC to preserve the liquid phase to prevent amyloid fibril formation, [106] providing a deeper understanding and insight into the molecular mechanisms and important roles of membraneless organelles to maintain the liquid or gel phase. In some neuronal injury responses, TIAR-2 can form liquid-like TIAR-2 granules by LLPS and inhibit axonal regeneration. However, the non-phosphorylatable TIAR-2 variant does not form granules and is unable to inhibit axonal regeneration. The critical role of phosphorylation-mediated phase separation in axon regeneration can be seen [76].
In the field of viral infection, measles virus inclusion bodies are formed by a LLPS process and phosphorylation of phosphoproteins Ser86 and Ser151 can improve the efficiency of the inclusion body assembly [61]. PTM is a key step affecting viral inclusion body assembly. In investigating the potential mechanisms of LLPS in virus-infected cells, phosphorylation of its nonstructural protein 2 (NS2) was found to induce and regulate phase separation of NS2 in bluetongue virus, [74] and Ser51 phosphorylation in the N-protein of globally transmitted SARS-CoV-2 was found to potentially inhibit the RNA-binding ability of the NTD structural domain, further promoting phase separation of the N-protein [60].
The effect of phosphorylation at different sites of the same protein on phase separation may be different. For example, phosphorylation of Ser78 of postsynaptic density protein 95 (PSD-95) inhibits phase separation of GluN2B and the auxiliary protein stargazin, whereas phosphorylation of Ser116 induces phase separation of stargazin [66]. Phase separation has been reported to play an important role in the phosphorylation modification of proteins involved in β-catenin, Pol II, PPP5C CHK1, CTD, MAPK3, and Fus3, providing data for insight into the prevalence and biological functions of phase separation [44, 45, 47, 48, 107].
Methylation modifications
Methylation modification transfers activated methyl groups provided by S-adenosylmethionine (SAM) to specific protein residues by methyltransferases (Fig. 2) [108]. In recent years, a large amount of experimental data has shown that methylation negatively modulates the tendency of phase separation. Nott TJ et al. demonstrated that Ddx4, the major component of Nuage protein, forms structures in vitro that are indistinguishable from intracellular membraneless organelles and form droplets in living cells. However, methylation of arginine in Ddx4 significantly destabilizes droplets and reduces LLPS, [77] which provides ideas for the dynamic regulation of biota compartmentalization. In another related study of membraneless organelles, protein arginine methyltransferase PRMT1 knockdown impairs the localization of RAP55A in the P-body, suggesting that elimination of methylation may have a positive effect on phase separation [82]. In neurodegenerative diseases, FUS arginine methylation has been shown to tend to inhibit phase separation by modifying the interaction of the cation π with tyrosines in the N-terminal LCD, [78, 79] and asymmetric dimethylation in cells decreases nuclear aggregation of FUS [79]. In the same year, a study found that asymmetric dimethylation of hnRNPA2 LCD reduced co-phase separation and co-aggregation with the LCD at the C-terminus of TDP-43 by disrupting arginine-aromatic interactions [80]. In 2020, a study showed that dimethylarginine (DMA) of poly-GR, a dipeptide repeat protein (DPR) associated with TDP-43 inclusions, in the brains of C9orf72 FTD/ALS patients reduced the strength of interactions between poly-GR molecules and decreased the phase separation of poly-GR [109]. Furthermore, in another disease study, Tsang B et al. found that methylation of the low-complexity disordered region (LCR) at the C-terminus of FMRP reduced phase separation in vitro, [73] supporting the role of methylation on the propensity for phase separation. Interestingly, although the positive effect of methylation on phase separation is less reported, it is undeniable that methylation modifications can promote the formation of phase separation [81]. For example, Arribas-Layton M et al. found that symmetric arginine dimethylation of the RGG structural domain of Lsm4 stimulated the accumulation of processing bodies [81].
Acetylation modification
Acetylation modification is the process of transferring acetyl groups of acetyl coenzyme A to protein amino acid residues in the presence of enzymatic/non-enzymatic action (Fig. 2) [110, 111]. Acetylation modifications are divided into two main categories: those that occur at the lysine end of the protein, and those that occur at the N-terminal end of the protein [110].
A growing number of studies have shown that lysine acetylation modifications of proteins associated with neurodegenerative diseases can mediate the tendency of phase separation and influence the disease process. In 2018, data while studying the aggregation properties of tau showed that hyperacetylation of tau by p300 histone acetyltransferase (HAT) inhibited LLPS and heparin-induced aggregation of tau and prevented microtubule assembly that effectively affected disease progression [38]. Acetylation of residues K321 and K274 of 4R0N tau has been reported to inhibit tau aggregation. Interestingly, [83] acetylation at 4R2N tau residue K274 significantly reduced the critical concentration of tau for LLPS and induced conformational changes in tau, and suggested that acetylation of tau at residue K274 could increase tau aggregation [84]. This suggests that acetylation of the K274 residue produces different aggregation tendencies for different tau isomers. A large amount of TDP-43 can be acetylated, and in 2022 Garcia Morato J et al. found that acetylation of the K136 site of the RNA recognition domain of TDP-43 impaired its RNA binding and splicing ability. This failure of RNA interactions triggered TDP-43 phase separation [85]. Imbalance in the dynamic regulation of lysine acetylation and deacetylation of TDP-43 protein may be closely associated with diseases such as amyotrophic lateral sclerosis (ALS) and frontotemporal lobar degeneration (FTLD) [112]. Acetylation modifications also regulate the assembly of membraneless organelles. For example, Saito M et al. reported that the RNA decarboxylase DDX3X, an important component of stress granules (SGs), is a novel substrate for the deacetylase HDAC6. Acetylation on multiple lysine residues of the N-terminal IDR of DDX3X (IDR1) impairs droplet formation. And the enhancement of LLPS propensity by deacetylation of DDX3X-IDR1 by HDAC6 is necessary for SG maturation [86]. In terms of gene expression, p300 histone acetylation antagonizes chromatin phase separation and reduces the formation of droplets in the nucleus [87]. However, in the presence of polybrominated structural domain proteins, such as BRD4, highly acetylated chromatin forms new phase separation states, and these droplets have different physical properties, and their non-fusion with unmodified chromatin droplets have important implications for our understanding of the organization and regulation of eukaryotic genomes [87]. Subsequently, Han X et al. in their study of the role of BRD4 short isoforms in phase separation and active gene transcription found that BRD4S LLPS is formed through multivalent interactions of IDRs and regulates chromosomal DNA-histone interactions through histone lysine acetylation [21]. In 2022, Qin Z et al. demonstrated that the addition of the acetyl portion at specific lysine residues of the IRF3/IRF7 DNA-binding domain (DBD) eliminates IRF3/IRF7 LLPS, while SIRT1 recognizes acetylated IRF3/IRF7 and promotes LLPS through catalytic deacetylation reactions, elucidating a deacetylation-mediated IRF3/IRF7 LLPS by a novel mechanism of innate antiviral immune control [88]. Currently, there is some progress in protein N-terminal acetylation-mediated phase separation studies, and in 2021 Bock AS et al. observed a difference in the LLPS propensity of N-terminal acetylated FUS-LC compared to those without modification. Furthermore, the team evaluated the aggregation propensity of N-terminal acetylated FUS-LC and found that N-terminal acetylation can slow down aggregation under certain conditions [89].
Ubiquitination modifications
Ubiquitination is a common PTM in eukaryotic cells, and the covalent attachment of one or more ubiquitin molecules to multiple sites and proteins by the action of ubiquitin-activating enzyme 1 (E1), ubiquitin-conjugating enzyme 2 (E2) and ubiquitin ligase 3 (E3) (Fig. 2) [113]. Deubiquitinase (DUB) plays a role in deubiquitination [114]. In recent years, an increasing number of experimental data have shown crosstalk between PTM and phase separation of proteins.
In 2021 Höllmüller E et al. proposed that site-specific ubiquitination plays a general regulatory role for spliceosomal H1 and that K64 site-specific ubiquitination affects H1-dependent chromosome assembly and phase separation [90]. In 2022 several studies further confirmed that ubiquitination modifications regulate the formation and stabilization of phase separation. For example, Parolini F et al. found that enzymatic ubiquitination of the Alzheimer’s-related protein tau stabilized droplets and prevented aggregation-associated lysis [91]. Gao K et al. described that SPOP binding and induction of nondegradative ubiquitination modification of SQSTM1 at L420 can reduce SQSTM1 body formation, liquid phase condensation, dimerization, and ubiquitin-binding capacity [92]. Furthermore, Tozawa T et al. found that activation of the intracellular ubiquitin cascade reaction promoted the tripartite motif (TRIM) family of ubiquitin ligases molecules into cytoplasmic bodies via LLPS, separating potentially harmful excess TRIM molecules from the cytoplasmic environment [93]. Gao Y et al. found that endoplasmic reticulum-localized AGO proteins recruit LTn1 through lipid-mediated phase separation formation of AGO condensates to catalyze de novo peptide ubiquitination [115, 116].
Phase separation allows ubiquitination and subsequent proteasomal degradation by triggering ubiquitin ligase substrates that activate ubiquitin ligases, which may play a key role in regulating ubiquitin-dependent protein homeostasis [117,118,119]. In 2018 Bouchard JJ et al. found that substrates trigger phase separation of speckle-type POZ protein (SPOP) in vitro and co-localization in membraneless organelles in cells while cancer-associated mutations in SPOP disrupt mislocalization due to phase separation and thus reduce the level of ubiquitination in cells [117]. The direct role of LLPS in stimulating histone ubiquitination was further elucidated by Gallego LD et al. Lge1, together with Bre1-Rad6 and nucleosomes, forms a spatially organized “reaction chamber” that stimulates H2B ubiquitination [46].
PTMs of ubiquitin-like proteins
With the discovery of various ubiquitin-like proteins and ubiquitination modifications, the correlation between PTMs of ubiquitin-like proteins and phase separation has been gradually investigated, such as small ubiquitin-related modifier (SUMO), neural precursor cell-expressed developmentally downregulated 8(NEDD8), etc., has been gradually investigated.
SUMOylation
SUMO, short for Small Ubiquitin-like Modifier, belongs to the ubiquitin-like protein family. Similar to ubiquitin, SUMO can be covalently attached to lysine side chains in various target proteins. Notably, SUMOylation distinguishes itself from other PTMs by not being a chemical group added to amino acid residues, [89] but rather by forming an isopeptide bond with the target protein. This bond is created through the coordinated activity of specific enzymes, namely the E1 SUMO activation enzyme, E2 SUMO conjugation enzyme, and E3 SUMO ligase (Fig. 2) [113]. Additionally, members of the SENP family possess the ability to cleave the isopeptide bond between SUMO and the target protein [114, 120]. With the continuous research and expansion of the research, the involvement of SUMOylation in the phase separation process is partially understood. It was found that SOP-2, one of the components of Polycomb-related complex 1 (PRC 1), could phase separate and that SUMOylation could regulate this phase separation process, as evidenced by an increase in the diameter and mobility of microdroplets [94]. In acquired immunodeficiency syndrome (AIDS), the phase segregation property of CBX4 allows it to recruit the catalytic subunit EZH2 at the retroviral HIV-1 promoter, which plays a key role in EZH2 SUMOylation and HIV-1 latency [96]. In addition to this, SLX4 can form phase separated droplets within the cell and SUMO-SIM interactions promote the assembly of SLX4 condensates [95].
Neddylation
Compared to other ubiquitin-like proteins, the NEDD8 is a molecule with the greatest similarity to ubiquitin, which binds to substrate proteins in a modification process called Neddylation [121, 122]. COP9 signalosome (CSN) and NEDD8 protease 1 (NEDP1), which release substrates and NEDD8 (Fig. 2) [123, 124]. In 2022, A study demonstrated that PML/RARα fusion proteins that drive chromosomal translocation production in acute promyelocytic leukemia (APL) are assembled by LLPS into nucleosomal structures. Further investigation of the mechanism revealed the presence of novel PTMs of ubiquitin-like proteins, Neddylation modification, that disrupt the phase separation process of PML/RARα by facilitating its binding to DNA, ultimately interfering with the assembly of PML nucleosomes [97]. This is the first time that the ubiquitinylation-like modification Neddylation regulates the phase separation process, which further expands the scope of PTMs in the field of phase separation.
Glycosylation modifications
O-GlcNAcylation
GlcNAcylation is linked and involved in regulating the phase separation process (Fig. 2). In 2022, Lv P et al. demonstrated through extensive experimental data that T1306 O-GlcNAc of SynGAP prevents the formation of hydrogen bonds with PSD-95, disrupts protein interactions, and thus inhibits LLPS formation, and that O-GlcNAc-dependent LLPS is subject to O-GlcNAc transferase (OGT) and O-GlcNAcase (OGA) reversible regulation [98]. This finding further opens up a new perspective in the study of the correlation between PTMs and phase separation of proteins and pushes the research process in this field to a new level.
Poly ADP-ribosylation (PARylation)
PARylation is an important PTM that involves the addition of poly(ADP-ribose) (PAR) onto specific proteins [125]. This process is primarily catalyzed by a family of enzymes called Poly(ADP-ribose) polymerase (PARP), with PARP1 being the most well-known [125, 126]. PARP1 converts NAD + into ADP-ribose and then adds individual ribose units onto specific amino acid residues of target proteins, forming PAR chains [127]. Hydrolytic enzymes such as PAR glycohydrolase (PARG), ADP-ribosylhydrolase 3 (ARH3), TARG1, MACROD1, MACROD1, etc. can reverse this process (Fig. 2) [128, 129]. Under physiological conditions, PARylation regulates the phase-separation-mediated assembly and disassembly of stress granules containing various RNA-binding proteins (RBPs) [130]. In addition, PARylation modifications and phase separation play an important role in the maintenance of genomic transcriptional stability after DNA damage. In 2022 Fu H et al. revealed that poly(ADP-ribose) polymerase 1 (PARP1)-mediated CDK9-cyclin T1 (CycT1) PARylation through the PARylation-dependent suppression of CycT1 phase separation and hyperphosphorylation of Pol II CTD by P-TEFb inhibits Pol II elongation and induces global transcriptional shutdown in response to DNA damage [99]. Additionally, it was shown that PARylation levels are a major regulator of the assembly-disassembly kinetics of ribonucleoprotein (RNP) granules (e.g. hnRNP A1 and TDP-43) containing disease-associated RBPs [131].
Hydroxylation modifications
There is no systematic study and interpretation of LLPS related to hydroxylation modifications. However, the possibility that JMJD6 catalysis of lysine hydroxylation regulates higher-order protein associations through effects on LLPS and the actions of this process on membraneless organelles or cellular condensates was proposed in 2022 based on the results of the study [132]. This suggests a new way of thinking about the further development of our field in the future.
Other novel protein modifications
The research on novel protein modifications such as sulfation, acylation, glutamylation, palmitoylation, carbonylation, glutarylation, myristoylation, crotonylation, malonylation, succinylation, etc. is still in the initial stage and relatively few, the crosstalk of these novel protein modifications and phase separation need to be further explored.
Issues and prospects
This paper provides an overview of the interactions between PTMs of various proteins and the phenomenon of cytosolic phase separation. It reviews current research on the intricate links between PTMs and cytosolic phase separation. In addition, the paper provides the latest knowledge on intracellular activities and macromolecular assembly, while extensively discussing the topic of the formation of membrane-free compartments and how they depend on PTMs. The understanding of the spatiotemporal dynamics of physiological responses regulated by PTMs at the subcellular level is deepened.
The current understanding of the crosstalk between phase separation and PTMs is mostly confined to the description of the phenomenon, and there is still uncertainty about the causal relationship between them and the specific regulatory mechanisms [133, 134]. Therefore, digging deeper into the biomolecular motifs where phase separation occurs, exploring the interactions between multiple modification sites of phase separation-associated proteins and phase separation, and determining the regulatory relationships and biological functions are our current urgent problems [135]. Pathological aggregation of proteins is closely related to the occurrence of diseases, such as tau [136]. In the future biological field, there is a great deal of potential for exploring how to utilize PTMs on the regulatory mechanism of phase separation to understand the pathogenesis of diseases and to develop targeted drugs [135,136,137,138].
Availability of data and materials
Data sharing does not apply to this article as no datasets were generated or analyzed during the current study.
References
Kim CG, Hwang DE, Kumar R, Chung M, Eom YG, Kim H, Koo DH, Choi JM. Recent trends in studies of biomolecular phase separation. BMB Rep. 2022;55:363–9.
Brangwynne CP, Eckmann CR, Courson DS, Rybarska A, Hoege C, Gharakhani J, Julicher F, Hyman AA. Germline P granules are liquid droplets that localize by controlled dissolution/condensation. Science. 2009;324:1729–32.
Shimada M, Oyama N. Gas-liquid phase separation at zero temperature: mechanical interpretation and implications for gelation. Soft Matter. 2022;18:8406–17.
Salehi M, Farzamfar S, Bozorgzadeh S, Bastami F. Fabrication of poly(L-Lactic acid)/Chitosan scaffolds by Solid-Liquid Phase Separation Method for Nerve Tissue Engineering: an in Vitro Study on Human neuroblasts. J Craniofac Surg. 2019;30:784–9.
Brangwynne CP, Tompa P, Pappu RV. Polymer physics of intracellular phase transitions. Nat Phys. 2015;11:899–904.
Mitrea DM, Kriwacki RW. Phase separation in biology; functional organization of a higher order. Cell Commun Signal. 2016;14:1–1.
Nesterov SV, Ilyinsky NS, Uversky VN. Liquid-liquid phase separation as a common organizing principle of intracellular space and biomembranes providing dynamic adaptive responses. Biochim Biophys Acta Mol Cell Res. 2021;1868:119102.
Hyman AA, Weber CA, Jülicher F. Liquid-liquid phase separation in Biology. Annu Rev Cell Dev Biol. 2014;30:39–58.
Alberti S. The wisdom of crowds: regulating cell function through condensed states of living matter. J Cell Sci. 2017;130:2789–96.
Burke KA, Janke AM, Rhine CL, Fawzi NL. Residue-by-Residue View of in Vitro FUS granules that bind the C-Terminal domain of RNA polymerase II. Mol Cell. 2015;60:231–41.
Li PP, Banjade SS, Cheng HH, Kim SS, Chen BB, Guo LL, Llaguno MM, Hollingsworth JVJV, King DSDS, Banani SFSF, et al. Phase transitions in the Assembly of Multi-valent Signaling proteins. Nat (London). 2012;483:336–40.
Elbaum-Garfinkle S, Kim Y, Szczepaniak K, Chen CC, Eckmann CR, Myong S, Brangwynne CP. The disordered P granule protein LAF-1 drives phase separation into droplets with tunable viscosity and dynamics. Proc Natl Acad Sci U S A. 2015;112:7189–94.
Kato M, Han TW, Xie S, Shi K, Du X, Wu LC, Mirzaei H, Goldsmith EJ, Longgood J, Pei J, et al. Cell-free formation of RNA granules: low complexity sequence domains form dynamic fibers within hydrogels. Cell. 2012;149:753–67.
Feng Z, Jia B, Zhang M. Liquid-liquid phase separation in Biology: specific stoichiometric molecular interactions vs promiscuous interactions mediated by disordered sequences. Biochemistry. 2021;60:2397–406.
Banani SF, Lee HO, Hyman AA, Rosen MK. Biomolecular condensates: organizers of cellular biochemistry. Nat Rev Mol Cell Biol. 2017;18:285–98.
Alberti S, Hyman AA. Are aberrant phase transitions a driver of cellular aging? BioEssays. 2016;38:959–68.
Zhang C, Lai L. Physiochemical mechanisms of Biomolecular Liquid-Liquid phase separation. Wu Li Hua Xue Xue Bao. 2020;36:154–68.
Wu R, Li P. Liquid-liquid phase separation and biomolecular condensates (in Chinese). Chin Sci Bull. 2019;64:2285–91.
Stender E, Ray S, Norrild RK, Larsen JA, Petersen D, Farzadfard A, Galvagnion C, Jensen H, Buell AK. Capillary flow experiments for thermodynamic and kinetic characterization of protein liquid-liquid phase separation. Nat Commun. 2021;12:7289.
Zhao M, Yu Y, Sun LM, Xing JQ, Li T, Zhu Y, Wang M, Yu Y, Xue W, Xia T, et al. GCG inhibits SARS-CoV-2 replication by disrupting the liquid phase condensation of its nucleocapsid protein. Nat Commun. 2021;12:2114.
Han X, Yu D, Gu R, Jia Y, Wang Q, Jaganathan A, Yang X, Yu M, Babault N, Zhao C, et al. Roles of the BRD4 short isoform in phase separation and active gene transcription. Nat Struct Mol Biol. 2020;27:333–41.
Berry J, Brangwynne CP, Haataja M. Physical principles of intracellular organization via active and passive phase transitions. Rep Prog Phys. 2018;81:046601.
Brady JP, Farber PJ, Sekhar A, Lin YH, Huang R, Bah A, Nott TJ, Chan HS, Baldwin AJ, Forman-Kay JD, Kay LE. Structural and hydrodynamic properties of an intrinsically disordered region of a germ cell-specific protein on phase separation. Proc Natl Acad Sci U S A. 2017;114:E8194–203.
Lin N, Yao K, Chen X. Research Progress of Molecular Recognition and Interaction of Crystallins linking cataract. Chem J Chin Universities. 2021;42:3379–89.
Ambadipudi S, Biernat J, Riedel D, Mandelkow E, Zweckstetter M. Liquid-liquid phase separation of the microtubule-binding repeats of the Alzheimer-related protein tau. Nat Commun. 2017;8:275–13.
Muiznieks LD, Keeley FW. Proline periodicity modulates the self-assembly properties of elastin-like polypeptides. J Biol Chem. 2010;285:39779–89.
Liu W. TRIM21 drives phase-separation and regulates the development of Breast Cancer. Nanjing Normal University; 2021.
Jin X, Zhou M, Chen S, Li D, Cao X, Liu B. Effects of pH alterations on stress- and aging-induced protein phase separation. Cell Mol Life Sci. 2022;79:380.
Cho EJ, Kim JS. Crowding-Induced phase separation of Lennard-Jones particles: implications to Nuclear structures in a Biological cell. J Phys Chem. 2012;116:3874–9.
Vweza AO, Song CG, Chong KT. Liquid-liquid phase separation in the Presence of Macromolecular Crowding and State-dependent kinetics. Int J Mol Sci. 2021;22:6675.
Ding X, Sun F, Chen J, Chen L, Tobin-Miyaji Y, Xue S, Qiang W, Luo S. Amyloid-forming segment induces aggregation of FUS-LC domain from phase separation modulated by site-specific phosphorylation. J Mol Biol. 2020;432:467–83.
Xiong Q, Chen Z, Ge F. Proteomic analysis of post translational modifications in cyanobacteria. J Proteom. 2016;134:57–64.
Peng T, Das T, Ding K, Hang HC. Functional analysis of protein post-translational modifications using genetic codon expansion. Protein Sci. 2023;32:e4618.
Sonenberg N, Hinnebusch AG. Regulation of translation initiation in eukaryotes: mechanisms and biological targets. Cell. 2009;136:731–45.
Ruan B, Dai P, Wang W, Sun J, Zhang W, Yan Z, Yang J. Progress on post-translational Modififi cation of proteins. Chin J Cell Biology. 2014;36:1027–37.
Zhang M, Xue B, Li Q, Shi R, Cao Y, Wang W, Li J. Sequence tendency for the Interaction between Low-Complexity intrinsically disordered proteins. JACS Au. 2023;3:93–104.
Bressler SG, Mitrany A, Wenger A, Nathke I, Friedler A. The oligomerization domains of the APC protein mediate liquid-liquid phase separation that is phosphorylation controlled. Int J Mol Sci. 2023;24:6478.
Josephine CF, Antrix J, Kyoung-Jae C, Phoebe ST, Kevin RM, Sung YJ, Allan CF. Acetylation disfavors tau phase separation. Int J Mol Sci. 2018;19:1360.
Savastano A, Flores D, Kadavath H, Biernat J, Mandelkow E, Zweckstetter M. Disease-Associated Tau Phosphorylation hinders Tubulin Assembly within Tau Condensates. Angew Chem Int Ed Engl. 2021;60:726–30.
Trivellato D, Floriani F, Barracchia CG, Munari F, D’Onofrio M, Assfalg M. Site-directed double monoubiquitination of the repeat domain of the amyloid-forming protein tau impairs self-assembly and coacervation. Bioorg Chem. 2023;132:106347.
Wang J, Choi JM, Holehouse AS, Lee HO, Zhang X, Jahnel M, Maharana S, Lemaitre R, Pozniakovsky A, Drechsel D, et al. A molecular Grammar governing the driving forces for phase separation of prion-like RNA binding proteins. Cell. 2018;174:688–699e16.
Her C, Phan TM, Jovic N, Kapoor U, Ackermann BE, Rizuan A, Kim YC, Mittal J, Debelouchina GT. Molecular interactions underlying the phase separation of HP1α: role of phosphorylation, ligand and nucleic acid binding. Nucleic Acids Res. 2022;50:12702–22.
Chen Y, Yang J. Role of liquid-liquid phase separation in cell physiology and Diseases. Shijie Huaren Xiaohua Zazhi. 2020;28:884–90.
Lu H, Yu D, Hansen AS, Ganguly S, Liu R, Heckert A, Darzacq X, Zhou Q. Phase-separation mechanism for C-terminal hyperphosphorylation of RNA polymerase II. Nature. 2018;558:318–23.
Mo W. Study on the mechanism of Arabidopsis blue light receptor CRY2-mediated phase separation to regulate the circadian clock. Jilin University; 2022.
Gallego LD, Schneider M, Mittal C, Romanauska A, Gudino CRM, Schubert T, Pugh BF, Köhler A. Phase separation directs ubiquitination of gene-body nucleosomes. Nature. 2020;579:592–7.
Li J, Cui P, Sun Q, Du Z, Chen Z, Li Z, Liu C, Cao Y, Yang Z, Liu R, Luo M. PSPC1 regulates CHK1 phosphorylation through phase separation and participates in mouse oocyte maturation. Acta Biochim Biophys Sin (Shanghai). 2021;53:1527–37.
Sang D, Shu T, Pantoja CF, Ibanez DOA, Zweckstetter M, Holt LJ. Condensed-phase signaling can expand kinase specificity and respond to macromolecular crowding. Mol Cell. 2022;82:3693–3711e10.
Owen I, Shewmaker F. The role of post-translational modifications in the phase transitions of intrinsically disordered proteins. Int J Mol Sci. 2019;20:5501.
Murray DT, Kato M, Lin Y, Thurber KR, Hung I, Mcknight SL, Tycko R. Structure of FUS protein fibrils and its relevance to self-assembly and phase separation of low-complexity domains. Cell. 2017;171:615–627e16.
Deng Q, Holler CJ, Taylor G, Hudson KF, Watkins W, Gearing M, Ito D, Murray ME, Dickson DW, Seyfried NT, Kukar T. FUS is phosphorylated by DNA-PK and accumulates in the cytoplasm after DNA damage. J Neurosci. 2014;34:7802–13.
Rhoads SN, Monahan ZT, Yee DS, Leung AY, Newcombe CG, O Meally RN, Cole RN, Shewmaker FP. The prionlike domain of FUS is multiphosphorylated following DNA damage without altering nuclear localization. Mol Biol Cell. 2018;29:1786–97.
Wang A, Conicella AE, Schmidt HB, Martin EW, Rhoads SN, Reeb AN, Nourse A, Ramirez MD, Ryan VH, Rohatgi R, et al. A single N-terminal phosphomimic disrupts TDP-43 polymerization, phase separation, and RNA splicing. EMBO J. 2018;37:e97452.
Gruijs DSL, Simonetti F, Hutten S, Riemenschneider H, Sternburg EL, Pietrek LM, Gebel J, Dotsch V, Edbauer D, Hummer G, et al. Disease-linked TDP-43 hyperphosphorylation suppresses TDP-43 condensation and aggregation. EMBO J. 2022;41:e108443.
Tariq A, Lin J, Noll MM, Torrente MP, Mack KL, Murillo OH, Jackrel ME, Shorter J. Potentiating Hsp104 activity via phosphomimetic mutations in the middle domain. FEMS Yeast Res. 2018;18:foy042.
Haj-Yahya M, Gopinath P, Rajasekhar K, Mirbaha H, Diamond MI, Lashuel HA. Site-specific hyperphosphorylation inhibits, rather than promotes, tau fibrillization, seeding capacity, and its Microtubule binding. Angew Chem Int Ed Engl. 2020;59:4059–67.
Ryan VH, Perdikari TM, Naik MT, Saueressig CF, Lins J, Dignon GL, Mittal J, Hart AC, Fawzi NL. Tyrosine phosphorylation regulates hnRNPA2 granule protein partitioning and reduces neurodegeneration. EMBO J. 2021;40:e105001.
Fang XD, Gao Q, Zang Y, Qiao JH, Gao DM, Xu WY, Wang Y, Li D, Wang XB. Host casein kinase 1-mediated phosphorylation modulates phase separation of a rhabdovirus phosphoprotein and virus Infection. Elife. 2022;11:e74884.
Carlson CR, Asfaha JB, Ghent CM, Howard CJ, Hartooni N, Safari M, Frankel AD, Morgan DO. Phosphoregulation of phase separation by the SARS-CoV-2 N protein suggests a Biophysical basis for its dual functions. Mol Cell. 2020;80:1092–1103e4.
Wu J, Zhong Y, Liu X, Lu X, Zeng W, Wu C, Xing F, Cao L, Zheng F, Hou P, et al. A novel phosphorylation site in SARS-CoV-2 nucleocapsid regulates its RNA-binding capacity and phase separation in host cells. J Mol Cell Biol. 2022;14:mjac003.
Zhou Y, Su JM, Samuel CE, Ma D. Measles Virus forms inclusion bodies with properties of Liquid Organelles. J Virol. 2019;93:e00948–19.
Lu Y, Wu T, Gutman O, Lu H, Zhou Q, Henis YI, Luo K. Phase separation of TAZ compartmentalizes the transcription machinery to promote gene expression. Nat Cell Biol. 2020;22:453–64.
Dai Z, Li G, Chen Q, Yang X. Ser392 phosphorylation modulated a switch between p53 and transcriptional condensates. Biochimica et Biophysica Acta (BBA) - Gene Regulatory mechanisms. 2022; 1865:194827.
Na Z, Luo Y, Cui DS, Khitun A, Smelyansky S, Loria JP, Slavoff SA. Phosphorylation of a human microprotein promotes dissociation of Biomolecular condensates. J Am Chem Soc. 2021;143:12675–87.
Lenard AJ, Hutten S, Zhou Q, Usluer S, Zhang F, Bourgeois B, Dormann D, Madl T. Phosphorylation regulates CIRBP arginine methylation, Transportin-1 binding and liquid-liquid phase separation. Front Mol Biosci. 2021;8:689687.
Vistrup-Parry M, Chen X, Johansen TL, Bach S, Buch-Larsen SC, Bartling CRO, Ma C, Clemmensen LS, Nielsen ML, Zhang M, Strømgaard K. Site-specific phosphorylation of PSD-95 dynamically regulates the postsynaptic density as observed by phase separation. iScience. 2021;24:103268–8.
Ostendorp A, Ostendorp S, Zhou Y, Chaudron Z, Wolffram L, Rombi K, von Pein L, Falke S, Jeffries CM, Svergun DI, et al. Intrinsically disordered plant protein PARCL colocalizes with RNA in phase-separated condensates whose formation can be regulated by mutating the PLD. J Biol Chem. 2022;298:102631–1.
Duran-Arque B, Canete M, Castellazzi CL, Bartomeu A, Ferrer-Caelles A, Reina O, Caballe A, Gay M, Arauz-Garofalo G, Belloc E, Mendez R. Comparative analyses of vertebrate CPEB proteins define two subfamilies with coordinated yet distinct functions in post-transcriptional gene regulation. Genome Biol. 2022;23:192.
Schilling M, Prusty AB, Boysen B, Oppermann FS, Riedel YL, Husedzinovic A, Rasouli H, König A, Ramanathan P, Reymann J, et al. TOR signaling regulates liquid phase separation of the SMN complex governing snRNP biogenesis. Cell Rep. 2021;35:109277.
Larson AG, Elnatan D, Keenen MM, Trnka MJ, Johnston JB, Burlingame AL, Agard DA, Redding S, Narlikar GJ. Liquid droplet formation by HP1α suggests a role for phase separation in heterochromatin. Nat (London). 2017;547:236–40.
Zhang G, Wang Z, Du Z, Zhang H. mTOR regulates phase separation of PGL granules to modulate their autophagic degradation. Cell. 2018;174:1492–1506e22.
Emperador-Melero J, Wong MY, Wang S, de Nola G, Nyitrai H, Kirchhausen T, Kaeser PS. PKC-phosphorylation of Liprin-alpha3 triggers phase separation and controls presynaptic active zone structure. Nat Commun. 2021;12:3057.
Tsang B, Arsenault J, Vernon RM, Lin H, Sonenberg N, Wang LY, Bah A, Forman-Kay JD. Phosphoregulated FMRP phase separation models activity-dependent translation through bidirectional control of mRNA granule formation. Proc Natl Acad Sci U S A. 2019;116:4218–27.
Rahman SK, Ampah KK, Roy P. Role of NS2 specific RNA binding and phosphorylation in liquid-liquid phase separation and virus assembly. Nucleic Acids Res. 2022;50:11273–84.
Sun D, Wu R, Zheng J, Li P, Yu L. Polyubiquitin chain-induced p62 phase separation drives autophagic cargo segregation. Cell Res. 2018;28:405–15.
Andrusiak MG, Sharifnia P, Lyu X, Wang Z, Dickey AM, Wu Z, Chisholm AD, Jin Y. Inhibition of Axon Regeneration by Liquid-like TIAR-2 granules. Neuron. 2019;104:290–304e8.
Nott TJ, Petsalaki E, Farber P, Jervis D, Fussner E, Plochowietz A, Craggs TD, Bazett-Jones DP, Pawson T, Forman-Kay JD, Baldwin AJ. Phase transition of a disordered nuage protein generates environmentally responsive membraneless organelles. Mol Cell. 2015;57:936–47.
Hofweber M, Hutten S, Bourgeois B, Spreitzer E, Niedner-Boblenz A, Schifferer M, Ruepp MD, Simons M, Niessing D, Madl T, Dormann D. Phase separation of FUS is suppressed by its nuclear import receptor and arginine methylation. Cell. 2018;173:706–719e13.
Qamar S, Wang G, Randle SJ, Ruggeri FS, Varela JA, Lin JQ, Phillips EC, Miyashita A, Williams D, Strohl F, et al. FUS phase separation is modulated by a molecular chaperone and methylation of Arginine Cation-Pi interactions. Cell. 2018;173:720–734e15.
Ryan VH, Dignon GL, Zerze GH, Chabata CV, Silva R, Conicella AE, Amaya J, Burke KA, Mittal J, Fawzi NL. Mechanistic view of hnRNPA2 low-complexity domain structure, interactions, and phase separation altered by mutation and arginine methylation. Mol Cell. 2018;69:465–479e7.
Arribas-Layton M, Dennis J, Bennett EJ, Damgaard CK, Lykke-Andersen J. The C-Terminal RGG domain of human Lsm4 promotes Processing body formation stimulated by Arginine Dimethylation. Mol Cell Biol. 2016;36:2226–35.
Matsumoto K, Nakayama H, Yoshimura M, Masuda A, Dohmae N, Matsumoto S, Tsujimoto M. PRMT1 is required for RAP55 to localize to processing bodies. RNA Biol. 2012;9:610–23.
Carlomagno Y, Chung DC, Yue M, Castanedes-Casey M, Madden BJ, Dunmore J, Tong J, Deture M, Dickson DW, Petrucelli L, Cook C. An acetylation-phosphorylation switch that regulates tau aggregation propensity and function. J Biol Chem. 2017;292:15277–86.
Rane JS, Kumari A, Panda D. An acetylation mimicking mutation, K274Q, in tau imparts neurotoxicity by enhancing tau aggregation and inhibiting tubulin polymerization. Biochem J. 2019;476:1401–17.
Garcia MJ, Hans F, von Zweydorf F, Feederle R, Elsasser SJ, Skodras AA, Gloeckner CJ, Buratti E, Neumann M, Kahle PJ. Sirtuin-1 sensitive lysine-136 acetylation drives phase separation and pathological aggregation of TDP-43. Nat Commun. 2022;13:1223.
Saito M, Hess D, Eglinger J, Fritsch AW, Kreysing M, Weinert BT, Choudhary C, Matthias P. Acetylation of intrinsically disordered regions regulates phase separation. Nat Chem Biol. 2019;15:51–61.
Gibson BA, Doolittle LK, Schneider MWG, Jensen LE, Gamarra N, Henry L, Gerlich DW, Redding S, Rosen MK. Organization of chromatin by intrinsic and regulated phase separation. Cell. 2019;179:470–484e21.
Qin Z, Fang X, Sun W, Ma Z, Dai T, Wang S, Zong Z, Huang H, Ru H, Lu H, et al. Deactylation by SIRT1 enables liquid-liquid phase separation of IRF3/IRF7 in innate antiviral immunity. Nat Immunol. 2022;23:1193–207.
Bock AS, Murthy AC, Tang WS, Jovic N, Shewmaker F, Mittal J, Fawzi NL. N-terminal acetylation modestly enhances phase separation and reduces aggregation of the low-complexity domain of RNA-binding protein fused in sarcoma. Protein Sci. 2021;30:1337–49.
Hollmuller E, Geigges S, Niedermeier ML, Kammer KM, Kienle SM, Rosner D, Scheffner M, Marx A, Stengel F. Site-specific ubiquitylation acts as a regulator of linker histone H1. Nat Commun. 2021;12:3497.
Parolini F, Tira R, Barracchia CG, Munari F, Capaldi S, D’Onofrio M, Assfalg M. Ubiquitination of Alzheimer’s-related tau protein affects liquid-liquid phase separation in a site- and cofactor-dependent manner. Int J Biol Macromol. 2022;201:173–81.
Gao K, Shi Q, Liu Y, Wang C. Enhanced autophagy and NFE2L2/NRF2 pathway activation in SPOP mutation-driven Prostate cancer. Autophagy. 2022;18:2013–5.
Tozawa T, Matsunaga K, Izumi T, Shigehisa N, Uekita T, Taoka M, Ichimura T. Ubiquitination-coupled liquid phase separation regulates the accumulation of the TRIM family of ubiquitin ligases into cytoplasmic bodies. PLoS ONE. 2022;17:e0272700.
Qu W, Wang Z, Zhang H. Phase separation of the C. Elegans polycomb protein SOP-2 is modulated by RNA and sumoylation. Protein Cell. 2020;11:202–7.
Alghoul E, Paloni M, Takedachi A, Urbach S, Barducci A, Gaillard PH, Basbous J, Constantinou A. Compartmentalization of the SUMO/RNF4 pathway by SLX4 drives DNA repair. Mol Cell. 2023;83:1640–58.
Wu L, Pan T, Zhou M, Chen T, Wu S, Lv X, Liu J, Yu F, Guan Y, Liu B, et al. CBX4 contributes to HIV-1 latency by forming phase-separated nuclear bodies and SUMOylating EZH2. EMBO Rep. 2022;23:e53855.
Chen Y. NEDDylation of PML/RAR regulates the assembly of PML nuclear bodies and its function in acute promyelocytic Leukemia. Zhejiang University; 2022.
Lv P, Du Y, He C, Peng L, Zhou X, Wan Y, Zeng M, Zhou W, Zou P, Li C, et al. O-GlcNAcylation modulates liquid-liquid phase separation of SynGAP/PSD-95. Nat Chem. 2022;14:831–40.
Fu H, Liu R, Jia Z, Li R, Zhu F, Zhu W, Shao Y, Jin Y, Xue Y, Huang J, et al. Poly(ADP-ribosylation) of P-TEFb by PARP1 disrupts phase separation to inhibit global transcription after DNA damage. Nat Cell Biol. 2022;24:513–25.
Jin F, Grater F. How multisite phosphorylation impacts the conformations of intrinsically disordered proteins. PLoS Comput Biol. 2021;17:e1008939.
Monahan Z, Ryan VH, Janke AM, Burke KA, Rhoads SN, Zerze GH, O’Meally R, Dignon GL, Conicella AE, Zheng W, et al. Phosphorylation of the FUS low-complexity domain disrupts phase separation, aggregation, and toxicity. EMBO J. 2017;36:2951–67.
Aumiller JWM, Keating CD. Phosphorylation-mediated RNA/peptide complex coacervation as a model for intracellular liquid organelles. Nat Chem. 2016;8:129–37.
Han TW, Kato M, Xie S, Wu LC, Mirzaei H, Pei J, Chen M, Xie Y, Allen J, Xiao G, Mcknight SL. Cell-free formation of RNA granules: bound RNAs identify features and components of Cellular assemblies. Cell. 2012;149:768–79.
Lao Z, Dong X, Liu X, Li F, Chen Y, Tang Y, Wei G. Insights into the atomistic mechanisms of phosphorylation in disrupting liquid-liquid phase separation and aggregation of the FUS Low-Complexity Domain. J Chem Inf Model. 2022;62:3227–38.
Wegmann S, Eftekharzadeh B, Tepper K, Zoltowska KM, Bennett RE, Dujardin S, Laskowski PR, Mackenzie D, Kamath T, Commins C, et al. Tau protein liquid-liquid phase separation can initiate tau aggregation. EMBO J. 2018;37:e98049.
Liu Z, Zhang S, Gu J, Tong Y, Li Y, Gui X, Long H, Wang C, Zhao C, Lu J, et al. Hsp27 chaperones FUS phase separation under the modulation of stress-induced phosphorylation. Nat Struct Mol Biol. 2020;27:363–72.
Nong J, Kang K, Shi Q, Zhu X, Tao Q, Chen YG. Phase separation of Axin organizes the beta-catenin destruction complex. J Cell Biol. 2021;220:e202012112.
Paik WK, Kim S, Protein Methylation. Chemical, Enzymological, and Biological Significance.; 1975. pp. 227–286.
Gittings LM, Boeynaems S, Lightwood D, Clargo A, Topia S, Nakayama L, Troakes C, Mann DMA, Gitler AD, Lashley T, Isaacs AM. Symmetric dimethylation of poly-GR correlates with Disease duration in C9orf72 FTLD and ALS and reduces poly-GR phase separation and toxicity. Acta Neuropathol. 2020;139:407–10.
Polevoda B, Sherman F. The diversity of acetylated proteins. Genome Biol. 2002;3:reviews0006.
Weinert BT, Iesmantavicius V, Moustafa T, Schölz C, Wagner SA, Magnes C, Zechner R, Choudhary C. Acetylation dynamics and stoichiometry in Saccharomyces cerevisiae. Mol Syst Biol. 2014;10:716.
Cohen TJ, Hwang AW, Restrepo CR, Yuan CX, Trojanowski JQ, Lee VM. An acetylation switch controls TDP-43 function and aggregation propensity. Nat Commun. 2015;6:5845.
Lu L, Li D, He F. Bioinformatics advances in protein ubiquitination. Hereditas(Beijing). 2013;35:17–26.
Conole D, Cao F, Am EC, Xue L, Kantesaria S, Kang D, Jin J, Owen D, Lohr L, Schenone M et al. Discovery of a potent deubiquitinase (DUB) small molecule activity-based probe enables broad spectrum DUB activity profiling in living cells. Angew Chem Int Ed Engl. 2023;62:e202311190.
Gao Y, Zhu Y, Wang H, Cheng Y, Zhao D, Sun Q, Chen D. Lipid-mediated phase separation of AGO proteins on the ER controls nascent-peptide ubiquitination. Mol Cell. 2022;82:1313–28.
Bengtson MH, Joazeiro CAP. Role of a ribosome-associated E3 ubiquitin ligase in protein quality control. Nature. 2010;467:470–3.
Jill JB, Joel HO, Daniel CS, Elzbieta S, Erik WM, Nafiseh S, Daniele G, Melissa RM, Kresten L, Xavier S, et al. Cancer mutations of the Tumor suppressor SPOP disrupt the formation of active, phase-separated compartments. Mol Cell. 2018;72:19–36e8.
Kwon JE, La Muhnho, Oh KH, Oh YM, Kim GR, Seol JH, Baek SH, Chiba T, Tanaka K, Bang OS, et al. BTB domain-containing speckle-type POZ protein (SPOP) serves as an adaptor of Daxx for ubiquitination by Cul3-based ubiquitin ligase. J Biol Chem. 2006;281:12664–72.
Hernandez MI, Lund AH, van der Stoop P, Boutsma E, Muijrers I, Verhoeven E, Nusinow DA, Panning B, Marahrens Y, van Lohuizen M. From the cover: stable X chromosome inactivation involves the PRC1 polycomb complex and requires histone MACROH2A1 and the CULLIN3/SPOP ubiquitin E3 ligase. Proc Natl Acad Sci U S A. 2005;102:7635–40.
Joana MPD, Jill T, Ronald TH. Ubch9 conjugates SUMO but not ubiquitin. FEBS Lett. 1997;417:297–300.
He S, Zhang L, He F. Progress in function and regulation of protein neddylation. Progress in Biochemistry and Biophysics. Progress in Biochemistry and Biophysics. 2009;36:1089–94.
Enchev RI, Schulman BA, Peter M. Protein neddylation: beyond cullin-RING ligases. Nat Rev Mol Cell Biol. 2015;16:30–44.
Kim WD, Mathavarajah S, Huber RJ. The Cellular and Developmental Roles of Cullins, Neddylation, and the COP9 signalosome in Dictyostelium Discoideum. Front Physiol. 2022;13:827435.
Yan F, Guan J, Peng Y, Zheng X. MyD88 NEDDylation negatively regulates MyD88-dependent NF-kappaB signaling through antagonizing its ubiquitination. Biochem Biophys Res Commun. 2017;482:632–7.
Gibson BA, Kraus WL. New insights into the molecular and cellular functions of poly(ADP-ribose) and PARPs. Nat Rev Mol Cell Biol. 2012;13:411–24.
Hottiger MO, Hassa PO, Luscher B, Schuler H, Koch-Nolte F. Toward a unified nomenclature for mammalian ADP-ribosyltransferases. Trends Biochem Sci. 2010;35:208–19.
Vyas S, Chang P. New PARP targets for cancer therapy. Nat Rev Cancer. 2014;14:502–9.
Niere M, Mashimo M, Agledal L, Dolle C, Kasamatsu A, Kato J, Moss J, Ziegler M. ADP-ribosylhydrolase 3 (ARH3), not poly(ADP-ribose) glycohydrolase (PARG) isoforms, is responsible for degradation of mitochondrial matrix-associated poly(ADP-ribose). J Biol Chem. 2012;287:16088–102.
Zaja R, Aydin G, Lippok BE, Feederle R, Luscher B, Feijs K. Comparative analysis of MACROD1, MACROD2 and TARG1 expression, localisation and interactome. Sci Rep. 2020;10:8286.
Liu C, Fang Y. New insights of poly(ADP-ribosylation) in neurodegenerative Diseases: a focus on protein phase separation and pathologic aggregation. Biochem Pharmacol. 2019;167:58–63.
Duan Y, Du A, Gu J, Duan G, Wang C, Gui X, Ma Z, Qian B, Deng X, Zhang K, et al. PARylation regulates stress granule dynamics, phase separation, and neurotoxicity of disease-related RNA-binding proteins. Cell Res. 2019;29:233–47.
Cockman ME, Sugimoto Y, Pegg HB, Masson N, Salah E, Tumber A, Flynn HR, Kirkpatrick JM, Schofield CJ, Ratcliffe PJ. Widespread hydroxylation of unstructured lysine-rich protein domains by JMJD6. Proc Natl Acad Sci U S A. 2022;119:e2201483119.
Zhang S, Pei G, Li B, Li P, Lin Y. Abnormal phase separation of biomacromolecules in human Diseases. Acta Biochim Biophys Sin (Shanghai). 2023;55:1133–52.
Zhang H, Ji X, Li P, Liu C, Lou J, Wang Z, Wen W, Xiao Y, Zhang M, Zhu X. Liquid-liquid phase separation in biology: mechanisms, physiological functions and human Diseases. Sci China Life Sci. 2020;63:953–85.
Wheeler RJ. Therapeutics-how to treat phase separation-associated Diseases. Emerg Top Life Sci. 2020;4:307–18.
Wang Y, Yu C, Pei G, Jia W, Li T, Li P. Dissolution of oncofusion transcription factor condensates for cancer therapy. Nat Chem Biol. 2023;19:1223–34.
Shi Y, Liao Y, Liu Q, Ni Z, Zhang Z, Shi M, Li P, Li H, Rao Y. BRD4-targeting PROTAC as a unique tool to study biomolecular condensates. Cell Discov. 2023;9:47.
Pan Y, Lu J, Feng X, Lu S, Yang Y, Yang G, Tan S, Wang L, Li P, Luo S, Lu B. Gelation of cytoplasmic expanded CAG RNA repeats suppresses global protein synthesis. Nat Chem Biol. 2023;19:1372–83.
Acknowledgements
We apologize to all researchers whose relevant findings are not cited and discussed here due to space limitation. We thank lab members who have contributed papers to this review.
Funding
This research was funded by the National Natural Science Foundation of China (82271643), Natural Science Foundation of Shandong Province (ZR2020MH061), Jinan Science and Technology Development Plan (202134060, 202225032).
Author information
Authors and Affiliations
Contributions
Y.L. collected the data and wrote the manuscript; Y.L. and B.W. designed the graph; B.W. and W.F. designed, and supervised the study; Y.W. and B.W. reviewed and finalized the manuscript; All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liu, Y., Feng, W., Wang, Y. et al. Crosstalk between protein post-translational modifications and phase separation. Cell Commun Signal 22, 110 (2024). https://doi.org/10.1186/s12964-023-01380-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12964-023-01380-1