
Abstract
Background
The role of long non-coding RNAs has been extensively appreciated in the contexts of cancer. Interferon γ-antisense RNA1 (IFNG-AS1) is an lncRNA located near to IFN-γ-encoding (IFNG) gene and regulates expression of IFNG in Th1 cells.
Methods
In the present study, we evaluated expression of IFNG and IFNG-AS1 in 108 breast samples including tumoral tissues and their adjacent non-cancerous tissues (ANCTs) using real-time PCR. IFNG-AS1 was significantly upregulated in tumoral tissues compared with ANCTs (expression ratio = 2.23, P = 0.03).
Results
Although the expression of IFNG was higher in tumoral tissues compared with ANCTs (relative expression = 1.89), it did not reach the level of significance (P = 0.07). IFNG expression was significantly higher in HER2-negative tumoral tissues compared with HER2-positive ones (P = 0.01) and in grade 1 samples compared with grade 2 ones (P = 0.03). No other significant difference was found in expressions of genes between other groups.
Conclusion
Significant strong correlations were detected between expression of IFNG and IFNG-AS1 in both tumoral tissues and ANCTs. The present study provides evidences for participation of IFNG and IFNG-AS1 in the pathogenesis of breast cancer and warrants future studies to elaborate the underlying mechanism.
Similar content being viewed by others


Background
Long non-coding RNAs (lncRNAs) are increasingly acknowledged as principal regulators of gene expression in the contexts of both cancer [1] and immunological disorders [2]. Considering the prominent role of immune system in control of carcinogenesis process, lncRNAs with regulatory roles on both immune cells and cancer cells are of particular value as tumor biomarkers or therapeutic targets. Interferon γ-antisense RNA1 (IFNG-AS1) is located near to IFN-γ-encoding (IFNG) gene. This lncRNA is regarded as a fundamental checkpoint that participates in IFNG expression in Th1 cells [3]. Targeting immune checkpoint molecules has been suggested as a new approach in cancer treatment. The specific pattern of expression of non-coding RNAs in tumoral tissues and their participation in initial phases of modulation of immune responses have potentiate them as novel candidates for changing the tumor microenvironment [4]. As IFN-based strategies along with immune checkpoint inhibitors are putative therapeutic options for malignancies [4], therapeutic modulation of IFNG-AS1 expression would exert beneficial effects in cancer patients from diverse aspects. Elevated expression of IFNG-AS1 lncRNA has been reported in Hashimoto’s thyroiditis (HT) patients in correlation with the proportion of circulating Th1 cells and IFNG gene expression [5]. The role of IFN-γ has been documented in both breast cancer pathogenesis and patients’ response to treatments. IFN-γ has been initially recognized for its role in antitumor host immunity which is exerted through induction of Th1 polarization and activation of both cytotoxic T cells (CTLs) and dendritic cells. Nevertheless, in certain conditions, IFN-γ function is in favor of tumor progression which has been documented by the observed negative effect of IFN-γ treatment on patient survival in some clinical trials. The underlying mechanism for such negative effect might be irresponsiveness to IFN-γ, downregulation of the MHC complex, or overexpression of other genes such as programmed cell death 1 ligand 1 (PD-L1) [6]. On the other hand, overexpression of IFN/STAT1-related genes has been suggested as prognostic markers of response to chemotherapy in estrogen receptor (ER) negative breast cancers [7]. More importantly, IFN-γ treatment in conjunction with anti-erbB2/neu mAb has significantly suppressed tumor growth in animal models [8]. In spite of several efforts to evaluate the efficiency of IFN-γ treatment in breast cancer, data regarding expression of IFNG gene in breast cancer tissues is scarce. In the present study, we assessed expression of IFNG gene and its natural occurring antisense RNA in 108 breast samples including tumoral tissues and their adjacent non-cancerous tissues (ANCTs) using real-time PCR in association with patients clinicopathological characteristics.
Methods
Patients
The current study enrolled 54 breast cancer patients. All patients had invasive ductal carcinoma of breast based on the histological examination. All of them have been recently diagnosed as having breast cancer and had no previous chemo/radiotherapy. The patients were admitted to Sina and Farmanieh hospitals over the years 2016–2017. The research protocol was approved by the ethical committee of Shahid Beheshti University of Medical Sciences (IR.SBMU.RETECH.REC.1397.403). All methods were performed in accordance with the relevant guidelines and regulations. Informed written consent was obtained from all patients. Tumoral tissues and ANCTs (0.5 cm × 0.5 cm) were excised from all patients during surgery, transferred in liquid nitrogen to the genetic laboratory of Shahid Beheshti University of Medical Sciences. Tissue samples were assessed by pathologists to endorse the diagnosis.
Expression analysis
Relative expressions of IFNG and IFNG-AS1 genes were assessed in tumoral tissues and paired ANCTs in the rotor gene 6000 Corbett Real-Time PCR System. Total RNA was extracted from tissue samples using TRIzol™ Reagent (Invitrogen, Carlsbad, CA, USA), and cDNA was synthesized by using RevertAid First Strand cDNA Synthesis Kit (TaKaRa, Japan). Al samples were treated with DNAse I to remove DNA contamination. SYBR Green RT-PCR Master Mix (TaKaRa, Japan) was used for expression analysis of genes. Expressions of genes were normalized to expression of Beta 2 microglobulin (B2M). The nucleotide sequences of primers are shown in Table 1. All experiments were performed in duplicate.
Statistical analysis
Student’s paired t test was used for analysis of differences in gene expression between paired samples. The association between clinicopathological data and transcript levels of each gene was assessed using chi-square test. Tukey’s honest significance test was used to assess the difference between mean values of transcript levels between different groups. The efficiency-corrected calculation model was used for assessment of fold changes of expression levels in tumoral tissues vs. ANCTs. The pairwise correlation between relative transcripts levels of IFNG and IFNG-AS1 genes was calculated using the regression model. For all statistical tests, the level of significance was set at P < 0.05. The receiver operating characteristic (ROC) curve was designed to assess the properness of gene expression levels for differentiating tumoral vs. ANCTs. The Youden index (j) was used to escalate the difference between sensitivity (true-positive rate) and 1 – specificity (false-positive rate).
Results
Elevated levels of IFNG and IFNG-AS1 can be used to identify breast cancer
IFNG-AS1 was significantly upregulated in tumoral tissues compared with ANCTs (expression ratio = 2.23, P = 0.03). Although the expression of IFNG was higher in tumoral tissues compared with ANCTs (relative expression = 1.89), it did not reach the level of significance (P = 0.07).
Figure 1 shows the –delta CT values (CT housekeeping - CT target gene) in tumoral tissues and ANCTs.

The relative expression of IFNG and IFNG-AS1 in tumoral tissues (n = 54) and ANCTs (n = 54) as presented by –delta CT values (CT housekeeping - CT target gene) in each set of samples
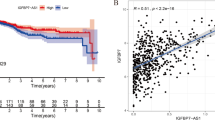
Assessment of correlation between expressions of IFNG and IFNG-AS1 genes revealed strong correlations between their expressions in both tumoral tissues and ANCTs (Fig. 2a, b).

Correlation between relative expressions of IFNG and IFNG-AS1 in tumoral (a) and ANCTs (b)
IFNG expression status is associated with clinical features of breast cancer
Table 2 shows the summary of demographic and clinicopathological data of study participants which have been gathered from questionnaires and patients’ medical records.
We compared expression level of IFNG and IFNG-AS1 in each tumoral tissue vs. its paired ANCT and classified patients based on these values to upregulation and downregulation groups. INFG and IFNG-AS1 were upregulated in tumoral tissues obtained from 35/54 (64%) and 37/54 (68%) of patients, respectively. Subsequently, we evaluated associations between clinicopathological data and relative expressions of IFNG and IFNG-AS1 genes. No significant associations were found between patients’ clinicopathological data and fold changes of expression of these genes in tumoral tissues compared with ANCTs. Table 3 shows the results of association analysis between relative expressions of genes in tumoral tissues compared with ANCTs and patients’ clinicopathological data.
Next, we compared relative expression of each gene in tumoral samples between clinicopathological-based groups (Table 4). IFNG expression was significantly higher in HER2-negative tumoral tissues compared with HER2-positive ones (P = 0.01) and in grade 1 samples compared with grade 2 ones (P = 0.03). No other significant difference was found in expressions of genes between other groups.
Assessment of the diagnostic value of IFNG and IFNG-AS1 in breast cancer
Based on the results of ROC curve analysis IFNG and IFNG-AS1 expressions had 83.3% specificity and 85.2% sensitivity for identification of disease status, respectively. The results of ROC curve analysis are shown in Fig. 3 and Table 5.

ROC curve for prediction of disease status based on the expression levels of IFNG and IFNG-AS1
Discussion
In the present study, we evaluated transcript levels of IFNG and IFNG-AS1 in breast cancer tissues and their paired ANCTs and found significant upregulation of IFNG-AS1 in tumoral tissues. IFNG-AS1 has been previously shown to regulate the expression of IFNG at both transcriptional and translational level in human CD4+ T cells [5]. Besides, strong positive correlations have been detected between the transcript levels of these two genes in thyroid tissues from HT patients [5]. Our data revealed the similar pattern of correlation between transcript levels of these genes in both tumoral tissues and ANCTs.
We could not find significant difference in expression of IFNG between tumoral tissues and ANCTs. García-Tuñón et al. have previously evaluated expression of IFNG in fibrocystic lesions, in situ tumors, and infiltrating tumors of breast and found higher expression of IFNG in in situ carcinoma than in benign and infiltrating tumors. They proposed IFNG as a prospective therapeutic modality in breast cancer [9]. In line with their observation, we found higher levels of IFNG in grade 1 samples compared with grade 2 ones. It is possible that tumor cells downregulate expression of IFNG as a mechanism for escaping from immune surveillance. We also detected higher IFNG expression in HER2-negative tumoral tissues compared with HER2-positive ones. IFN-γ has been previously shown to downregulate expression of HER2 in prostate cancer cells [10]. The existence of similar mechanism in breast cancer cells needs to be assessed. However, the direct effect of IFN-γ on HER2-positive breast cancer cells as reported by Nagai et al. [11] supports a similar function in the context of breast cancer.
Consistent with García-Tuñón et al. [9], we did not find any association between IFNG expression and ER/PR status. Mostafa et al. have shown an ERα inhibitory effect on IFN-γ signaling which results in immune escape in ERα-positive breast cancer cells [12]. However, such inhibitory effects are not necessarily exerted on expression of IFNG itself. Future studies are needed to assess the effect of estradiol or its receptor on IFNG expression.
We observed higher levels of IFNG-AS1 in breast cancer tissues compared with ANCTs. This finding might be either the cause or the consequence of the tumorigenesis process. Future functional studies are needed to elaborate the consequence of its overexpression in breast tissues. As previous studies have linked its overexpression with autoimmune conditions, it is possible that such overexpression is a compensatory mechanism to conquer immune evasion in tumor microenvironment. Critchley-Thorne et al. have evaluated the effectiveness of IFNG in peripheral blood lymphocytes from breast cancer patients and detected diminished IFN-γ-induced signaling in B cells of these patients in spite of normal signaling in T cells or natural killer cells [13]. Noticeably, no difference has been found within stages II, III, and IV breast cancer patients in this regard [13] which is in accordance with our finding regarding similar expression of the IFNG gene in histopathological-based groups. Consequently, there is a cell type-dependent regulatory mechanism for IFN function. So, future studies are needed to elaborate such mechanism in the epithelial tissues obtained from breast tumors to find whether these functional responses are impaired in the cancer tissue. Moreover, the significance of local expression of IFNG and IFNG-AS1 in response to systemic IFN-γ therapy of breast cancer patients must be investigated in imminent researches.
Conclusion
Apart from functional consequences of dysregulation of IFNG-AS1 in breast tumor tissues, transcript levels of this gene might be used for diagnosis purposes in the panels of putative biomarkers comprising both coding and non-coding genes. However, based on the results of ROC curve analysis, none of the assessed genes in the present study fulfill the requirements as an individual biomarker as the AUC values of both genes and their combinations were between 0.6 and 0.7 which means the poor accuracy of a diagnostic test.
References
Nikpayam E, Tasharrofi B, Sarrafzadeh S, Ghafouri-Fard S. The role of long non-coding RNAs in ovarian cancer. Iran Biomed J. 2017;21(1):3–15.
Eftekharian MM, Ghafouri-Fard S, Soudyab M, et al. Expression analysis of long non-coding RNAs in the blood of multiple sclerosis patients. J Mol Neurosci. 2017;63(3–4):333–41.
Collier SP, Collins PL, Williams CL, Boothby MR, Aune TM. Cutting edge: influence of Tmevpg1, a long intergenic noncoding RNA, on the expression of Ifng by Th1 cells. J Immunol. 2012;189(5):2084–8.
Smolle MA, Calin HN, Pichler M, Calin GA. Noncoding RNAs and immune checkpoints-clinical implications as cancer therapeutics. FEBS J. 2017;284(13):1952–66.
Peng HY, Liu YZ, Tian J, et al. The long noncoding RNA IFNG-AS1 promotes T helper type 1 cells response in patients with Hashimoto’s thyroiditis. Sci Rep. 2015;5:17702.
Mandai M, Hamanishi J, Abiko K, Matsumura N, Baba T, Konishi I. Dual faces of IFNgamma in cancer progression: a role of PD-L1 induction in the determination of pro- and antitumor immunity. Clin Cancer Res. 2016;22(10):2329–34.
Legrier ME, Bieche I, Gaston J, et al. Activation of IFN/STAT1 signalling predicts response to chemotherapy in oestrogen receptor-negative breast cancer. Br J Cancer. 2016;114(2):177–87.
Nagai Y, Tsuchiya H, Ji MQ, Zhang H, Greene MI. Synergistic effect of IFN-γ on breast cancer targeted therapy. (2017).
García-Tuñón I, Ricote M, Ruiz A, Fraile B, Paniagua R, Royuela M. Influence of IFN-gamma and its receptors in human breast cancer. BMC Cancer. 2007;7(1):158.
Kominsky SL, Hobeika AC, Lake FA, Torres BA, Johnson HM. Down-regulation of neu/HER-2 by interferon-gamma in prostate cancer cells. Cancer Res. 2000;60(14):3904–8.
Nagai Y, Tsuchiya H, Runkle EA, et al. Disabling of the erbB pathway followed by IFN-γ modifies phenotype and enhances genotoxic eradication of breast tumors. Cell Rep. 2015;12(12):2049–59.
Mostafa AA, Codner D, Hirasawa K, et al. Activation of ERα signaling differentially modulates IFN-γ induced HLA-class II expression in breast cancer cells. PLoS One. 2014;9(1):e87377.
Critchley-Thorne RJ, Simons DL, Yan N, et al. Impaired interferon signaling is a common immune defect in human cancer. Proc Natl Acad Sci. 2009;106(22):9010–5.
Acknowledgements
The current study was supported by a grant from Shahrekord University of Medical Sciences.
Funding
Not applicable
Availability of data and materials
The analyzed data sets generated during the study are available from the corresponding author on reasonable request.
Author information
Authors and Affiliations
Contributions
MT and SG-F supervised the study. HY and HA did the laboratory assessment. VKO analyzed the data. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All procedures performed in studies involving human participants were in accordance with the ethical standards of the institutional and/or national research committee and with the 1964 Helsinki declaration and its later amendments or comparable ethical standards.
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Yaghoobi, H., Azizi, H., Oskooei, V.K. et al. Assessment of expression of interferon γ (IFN-G) gene and its antisense (IFNG-AS1) in breast cancer. World J Surg Onc 16, 211 (2018). https://doi.org/10.1186/s12957-018-1508-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12957-018-1508-1