
Abstract
Food allergy appears to have its roots in an insufficient exposure to a diverse range of environmental microbiota during early life. Microbial exposure ensures the colonization of the gastrointestinal tract with commensal microbes, which is necessary for the induction of a balanced and tolerogenic immune function. High-throughput sequencing technology has facilitated in-depth studies of the gut microbiota as well as bacterial-derived metabolites. Although the role of the microbiota in allergies is now widely studied, its importance for food allergy was only recently noted. Studies in human cohorts have shown that there is an association of dysbiosis and pathogenesis of food allergy, while studies from animal models have demonstrated the capacity of specific species in the gut microbiota to alter immune response, which may lead to the desensitization of food allergy. This article reviews the role of the gut microbiota in food allergy, and discusses the influence of environmental factors as well as prevention and management strategies relating to such regulatory mechanism.
Similar content being viewed by others


Introduction
For decades, many cultivated microorganisms have been discovered by the microbiological culture technique, but these represent only a minority of the microbial species of the gut. The use of next generation high-throughput sequencing techniques has widened tremendously our knowledge of the human microbiota composition and its relationship to disease. The human body is estimated to consist of 10–100 trillion microbes, and more than 1000 bacterial species [1]. The highest number of microbes are found in the human gut [2]. The number of microbes found in the human gut are 10 times the number of cells making up the human body [3], although this ratio has been disputed [4]. Studies in the past few years have identified a critical role of the gut microbiota in shaping the immune system [5,6,7,8]. The gut microbiota is involved in the development of the organs of the immune system, and determines the tendencies of host immune responses. Research on the association between immune diseases and gut microbiota has reported that alterations in commensal bacteria can induce changes to the immune system [2, 9] which affect regulation of host metabolism, immune system maturation, and development of oral tolerance [10]. Thus, attempts to mitigate allergic diseases through adjusting the diversity and individuality of the gut microbiota have increased.
This review article outlines recent insights into the role of the gut microbiota in food allergy, the influence of environmental factors as well as prevention and management strategies involving the gut microbiota.
Dysbiosis and food allergy
Dysbiosis refers to a change in the microbiota composition and function such that it disrupts gut homeostasis and contributes to diseases [11]. There are increasing evidence from human studies suggesting that dysbiosis is associated with pathogenesis of food allergy (Table 1) [12,13,14,15,16,17,18,19,20,21,22,23]. Although these studies are not able to identify specific bacterial taxa that are consistently associated with food allergy due to heterogeneity in study design, such as different sampling time points, different techniques used to characterize the gut microbiota, and different allergic phenotypes, these studies show microbiota diversity and composition are significantly associated with the onset of food allergy.
Until now, it remains unclear how dysbiosis exactly affects the immune system in the development of food allergy but studies suggest that the gut microbiota influences the immune system by affecting host metabolism [2, 24,25,26] and the alteration of adaptive immunity [27, 28].
Metabolic and immune effects
Major attention has been directed to the possible role of short-chain fatty acids, such as butyrate, propionate, and acetate in affecting the immune system (Table 2) [29,30,31,32], since short-chain fatty acids are the main product of the digestive action of the gut microbiota [29, 30]. Production of short-chain fatty acids, particularly butyrate, is able to enhance the Vitamin A metabolism, in turns inducing the activity of ALDH in CD103+MLN DCs, and increasing the percentages of T regulatory (Tregs) cells and increasing IgA production [33]. Meanwhile, short-chain fatty acids are able to inhibit histone deacetylases activity, resulting in regulation of aldh1a1 expression, which contributes to immune tolerance. Other than this, short-chain fatty acids can bind metabolite-sensing G-protein coupled receptors, GPR43 or GPR109A [34], in turn promoting the tolerogenic CD103+ DC function and protecting against food allergies [35]. Moreover, short-chain fatty acids can reduce the production of pro-inflammatory cytokines including IL-1β, IL-6, IL-17 [36], and meanwhile increase the production of anti-inflammatory mediators including IL-10 [36, 37]. Thus, short-chain fatty acids are viewed as a key factor in promoting immunological tolerance towards harmless antigen and preventing inflammation.
Adaptive immunity
Adaptive immune responses are divided into two types: humoral immunity, regulated by B cells [38, 39], and cell-mediated immunity, regulated by T cells. The role of Tregs, subset of CD4+ T cells in oral tolerance development to food allergen, have been confirmed in animal models [40, 41] as well as human studies [42, 43] in which the induction of allergen-specific Treg cells is highly associated with a favourable allergy outcome. Microbiota, especially Clostridia species, in this case, were shown to be able to induce the production of Tregs [44, 45], which helps to inhibit allergic inflammation and promote oral tolerance [12, 46,47,48].
Specific Taxa related to food allergy across multiple studies
Bacteroidetes
An association between Bacteroidetes phyla in the gut and food allergy was found in a few epidemiologic studies [15, 18, 19]. Using gene sequencing methods, these studies identified that children with various types of food allergy had a lower relative abundance of Bacteroidetes compared with healthy children. The exact role of Bacteroidetes is unclear. However, a murine study indicated that Bacteroides fragilis, a species of Bacteroidetes, was able to produce polysaccharide A (PSA), which increased the suppressive capacity of the Treg cells and increased the production of IL-10 from Foxp3+ T cells [51]. The accumulated IL-10 mediates tolerance at mucosal surfaces and prevents intestinal inflammation. Further clinical studies in humans are needed to examine the regulatory role of Bacteroidetes in the development of food allergy.
Firmicutes
Firmicutes is the largest microbiota phylum containing more than 200 genera [52]. Most of the Firmicutes detected in the gut belong to the Clostridium clusters XIVa and IV. Both groups comprise members of the genera Clostridium, Eubacterium and Ruminococcus. Firmicutes are the most important butyrate producer in the gut and help to reduce inflammation through the inhibition of histone deacetylase [30]. A number of studies using 16S rRNA gene sequencing have identified a lower relative abundance of bacterial class Clostridia (bacterial phylum Firmicutes) in children with food allergy compared to healthy children [12, 13, 53, 54]. It is an accepted scientific knowledge that Clostridia promotes the accumulation of Treg cells [44, 45, 47, 55]. However, a study conducted by Stefka st al [55] demonstrated that Clostridia elevated IL-22 expression by RAR-related orphan receptor gamma (RORγt)+ ILCs and T cells, which in turn increasing an early, innate, barrier protective response. At the same time, Clostridia also elevated the production of IL-17, which in turn reducing intestinal epithelial permeability. Contradictory results also exist that patients with food allergy showed an increase in levels of Clostridia [18, 19, 56]. The genera of Clostridia that have been identified so far to have an association with food allergy are Clostridium [13, 18, 19, 53] and Ruminococcus [13, 15, 19, 54]. Diesner et al. reported that mice tolerant to food allergy have higher abundances of Ruminococcaceae [54] compared with intolerant counterparts, supporting the potential protective effects of Ruminococcus on food allergy. Nevertheless some discordant results the Clostridia class is that more clearly involved in mechanism of food tolerance induction and in food allergy protection.
Early-life colonization and environmental factors
Some studies argue that the gut microbiota is critical in early life for the development of the immune system as well as the development of allergic diseases [15, 18, 19, 53, 57]. The gut microbiota is unstable in the first 2–3 years of life during the initial colonization and development of the gut. Disruptions of microbial colonization during this period have been shown to increase the disease susceptibility during life. After the age of three the gut microbiota progresses towards an adult-like configuration and it essentially remains stable unless perturbed [1, 5, 53].
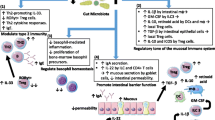
The dramatic rise of food allergy in modern society has led to the postulation of the hygiene hypothesis [58]. The hygiene hypothesis proposes that a lack of early childhood exposure to infectious agents suppresses the development of the immune system which leads to the rise of atopic diseases. Recent work has revisited the hygiene hypothesis model to include mode of delivery, antibiotic intake, diet [59] and synthetic chemicals as factors in altering gut microbiota (Fig. 1).

Early-life colonization and environmental factors. The gut microbiota is unstable in the first 2–3 years of life during the initial colonization and development of the gut. During this period, the gut microbiota is vulnerable to some disruptions such as mode of delivery, antibiotic intake, diet and synthetic chemicals, which may increase specific IgE, induce pro-inflammatory cytokines, reduce anti-inflammatory cytokines, inhibit the accumulation of Treg cells, and suppress the tolerogenic CD103+ DC (dendritic cell) functions, contributing to the pathogenesis of food allergy. After the age of 3 the gut microbiota progresses towards an adult-like configuration and this remains the same throughout the life
Mode of delivery
Perturbation of gut microbiota may influence susceptibility to food allergy [60]. It is evidenced that infants born vaginally are exposed to both maternal fecal and vaginal microbiota while infants born by caesarean section are exposed to maternal skin and environmental microbes only [50]. For instance, infants born by caesarean section are more likely to have lower abundance of Bacteroides and higher abundance of Firmicutes. Both Bacteroides and Firmicutes were revealed to associate with development of food allergy [50]. A population based birth cohort study revealed that children born by caesarean section had a threefold higher chance of developing food allergy [61]. However, the role of the delivery mode in influencing the food allergy outcome remains unclear as the delivery mode only exhibits an effect on the immune system for the first 90 days [62].
Antibiotic intake
Mothers undergoing a caesarean section are more likely to receive antibiotics [62], which can perturb the composition of the gut microbiota and modify the risk for allergic outcomes [63].
A number of studies have attempted to elucidate how antibiotics modify the composition of the gut microbiota and lead to the pathogenies of food allergy [64,65,66]. These studies revealed that antibiotic administration altered and reduced the microbiota diversity. A study suggested that antibiotic administration may cause reconstitution of the flora, thereby resulting in failure of the signal transmission via Toll-like receptor 4 (TLR4) [64]. The inability to signal via TLR4 resulted in markedly increased peanut-specific IgE and Th2 cytokine responses, contributing to allergic responses. Nevertheless, the association of antibiotic intake in early life and the development of food allergy is debated [67]. In a Chicago Family Cohort Food Allergy study, 1359 families were recruited and structure questionnaires were administered. This study did not find any association between antibiotic intake in the first year of life and the diagnosis of food allergy [67].
Diet
Diet plays an important role in the establishment of the gut microbiota and affects allergic symptoms [68, 69]. The dietary intake of infants starts with milk, either breast milk or formula [70]. Breast milk contains human milk oligosaccharides which stimulate the growth of Bifidobacterium bifidum [71] and genus Lactobacillus [72], which are the main probiotic organisms in the gut, forming an acidic environment with enriched short-chain fatty acids [73]. Consequently, breastfed infants are colonized with Bifidobacteria and Lactobacilli compared with formula fed infants [74]. Additionally, the breastfeeding duration also has an influence on the development of food allergy as infants with a very brief breastfeeding duration have a higher risk of developing cow’s milk allergy [75].
The milk intake is gradually replaced by a solid food diet that is similar to that of the adult and composed of a wide-variety of dietary macronutrients such as proteins, fats, carbohydrates and fibres [76]. These macronutrients are fundamental in determining microbiota composition and its effect on health outcomes.
After the introduction of complementary diet, the gut of infants exhibit more Lactobacillus, Ruminococcus, Bacteroides, Peptostreptococcus and Clostridium, which are tryptophan-catabolizing species [77]. Tryptophan is an amino acid commonly presented in high protein food, which is required for protein biosynthesis. Tryptophan metabolism plays a fundamental role in regulating the immune response as well as T cell proliferation. Other than this, tryptophan metabolism also induces IL-10 receptor-1 (IL-10R1) expression, which is essential in determining whether cells respond to IL-10, a potent anti-inflammatory cytokine that inhibits the release of pro-inflammatory cytokines [78].
Other than protein, fibre can also affect microbiota composition. The microbiota composition of the human gut associated with a high fibre diet is different from that seen with a high fat diet [79]. The gut microbiota with a high fat diet consisting of animal-based foods has higher abundance of Alistipes, Bilophila and Bacteroides and lower abundance of Roseburia, Eubacterium and Ruminococcus at genus level when compared to the gut microbiota with a high fibre diet [79]. The microbial conversions of dietary fibre to monosaccharides involve a number of metabolic pathways catalysed by the enzymatic activities of the gut microbiota and short-chain fatty acids are the major end-products of these microbial conversions [29, 30]. Mice studies were conducted in order to investigate the role of dietary fibre in suppressing immune response to food antigens [35, 80]. In these studies, mice were sensitized with food antigen and fed with either high fibre diet or no fibre diet. After few weeks of exclusion diet, mice receiving high fibre diet, showed significantly reduced symptoms of food allergy. Therefore, the “Western” diet with its high fat but low fibre content is proposed to be one of the reasons for the high prevalence of food allergy in Western countries [35, 81].
Synthetic chemicals
Synthetic chemicals play a role in perturbing the composition of gut microbiota as microbial metabolism of chemicals by gut microbiota may cause microbial dysbiosis [82], which in turn triggers allergy reaction [83, 84]. For example, food additives, one of the synthetic chemicals, are commonly found and used in our food as antioxidants, colourings, and flavourings, sweeteners and preservatives [83]. Mouse studies have indicated that such chemicals are associated with increased risks for food allergy [83, 84]. Food additives inhibited the accumulation of Treg cells, which is necessary for acquisition of oral tolerance [83]. The food additives also promoted the induction of allergic CD11b+ DCs, reduced the accumulation of tolerogenic CD103+ DCs as well as inhibited the induction of CD4+CD25hiT cells. Further to this, this study also suggested that intake of multiple food additives could increase the risk of developing food allergy.
Other synthetic chemicals, such as air pollutants, can be responsible for increasing the appearance of food allergy. Higher ambient levels of air pollutants, especially nitrogen dioxide have been consistently demonstrated to be associated with the increasing risk of allergies, including food allergies [85,86,87]. The mechanism of how air pollutant can cause food allergy remains unknown but studies suggested that inhaled air pollutants were able to directly or indirectly modify the composition of gut microbiota [88], which may cause an increase in the gut permeability and inflammation. Furthermore, inhaled air pollutants can lead to a substantial inflammatory response via reactive oxygen species (ROS) production and nuclear factor kappa B (NF-κB) activation in GI tract.
Therapeutic strategies
Human [12, 15, 18, 19, 53] and murine [54] models have demonstrated that study subjects with food allergy have distinct gut signatures and a different gut microbiota composition. Moreover, there is growing evidence that alteration of the gut microbiota may explain the development of food allergy [40, 41]. Microbial colonization has been shown to promote the induction of Treg cells, which is necessary to modulate the immune system and maintain tolerance to self-antigens [40, 41]. Microbiota may also influence the epigenetic modification of genes. It has been demonstrated that various forms of epigenetic changes, such as DNA methylation and histone modification, regulate the immune system. Also microbial derived metabolites, such as short-chain fatty acids, have been shown to reduce pro-inflammatory cytokines and induce anti-inflammatory mediators as well as inhibit histone deacetylases [36].
Research indicates that environmental factors affect the microbiota composition making it an ideal target of research to find new interventions to desensitize food allergy.
Probiotic supplementation
The use of probiotic supplementation seems an attractive option for the prevention and treatment of allergic diseases. Probiotics are defined as “live microorganisms which, when administered in adequate amounts as part of food, confer a health benefit on the host” [89, 90]. Probiotics can act as promoters of an adequate balance in the gut microbiota to prevent the development of allergies. The beneficial effects of probiotics involve restoring intestinal permeability to normal, improving the intestine’s immunological barrier function (both physical and mucous layer), promoting IgA production and inhibiting the release of proinflammatory cytokines through regulating gut microbiota composition [91, 92].
A well-characterised bacterial probiotic in desensitizing food allergy, especially cow’s milk allergy, is Lactobacillus rhamnosus GG [93]. Dietary intervention with Lactobacillus rhamnosus GG was found to reduce allergic responses towards cow’s milk in murine [94, 95] and human studies [93, 96, 97]. Lactobacillus rhamnosus GG was able to increase the production of different cytokines with proinflammatory (TNF-α and IL-6) or regulatory (IL-10) functions [98]. In addition, Lactobacillus rhamnosus GG also induced the accumulation of colonic Treg cells in the intestine [99]. In addition, Lactobacillus rhamnosus GG was able to increase FoxP3 demethylation rate, increase IL-4 and IL-5 DNA methylation rate, reduce IL-10 and IFN-γ DNA methylation rate, increase the expression of miR-155, -146a, -128 and -193a, to promote the acquisition of cow’s milk tolerance 12 months after treated with extensively hydrolyzed casein formula containing the probiotic Lactobacillus rhamnosus GG [97].
One study found that Lactobacillus rhamnosus GG showed a weak effect in desensitizing peanut allergy [100]. Most of the participants who received this probiotic and peanut oral immunotherapy passed the oral food challenge after receiving the treatment for 18 months and still passed the oral food challenge after 2 to 5 weeks after that. Moreover, participants who received probiotic and peanut oral immunotherapy showed a decrease in peanut sIgE levels and skin prick test wheal size, as well as an increase in peanut sIgG4 levels, at the end of treatment. Three months after treatment ended, these participants still had low sIgE levels and a small skin prick test wheal size. However, there was no convincing evidence that probiotic was effective in reducing allergic reactions to food as this study only compared between participants who received probiotic and peanut oral immunotherapy and participants who received placebo without including oral immunotherapy group only and probiotic group only. Also the prolonged tolerance towards peanut remains unclear as the participants in this study were not followed up beyond 3 months.
A mouse model of shellfish allergy demonstrated that oral administration of the probiotic strain Bifidobacterium longum reduced the specific IgE and stimulated dendritic cell maturation and CD103+ tolerogenic DCs accumulation in gut-associated lymphoid tissue. This, in turn, increased Tregs differentiation and suppressed Th2 responses [95, 101]. A murine study of cow’s milk allergy showed that oral administration of Bifidobacterium longum subsp. Infantis LA308 strain induced the expression of IL-10 and skewed the immune response to Th1. These results are encouraging to find a candidate probiotic strain to perform a clinical trial.
Two other murine studies have reported the capacity of Clostridium butyricum to reduce adverse reactions to egg and cow’s milk [46, 102]. The mechanism of how Clostridium butyricum is able to inhibit allergic inflammation remains unknown. Shi [102] suggested that the therapeutic effect of Clostridium butyricum may be generated by IL-10, as the IL-10-producing antigen specific Breg was found in mice with egg allergy which were administered with specific immunotherapy and Clostridium butyricum. On the other hand Zhang [46] suggested that Clostridium butyricum increased sIgA, CD4+CD25+Foxp3 Treg cell as well as reversed the imbalance of Th1/Th2 and Th17/Treg. These interesting results require further validation from human clinical trials.
There are three limitations on probiotic dietary intervention. First, the effect of probiotic is strain specific [103] and may vary depending on the individual’s lifestyle and baseline gut microbiota profile [104]. Secondly, abrupt termination of probiotic dietary intervention may further increase the gut dysbiosis [105]. Lastly, there are only a few studies about probiotic dietary intervention and further studies are needed to validate the findings in human system.
Prebiotic supplementation
Prebiotic is defined as “nonviable food component that confers a health benefit on the host through modulation of the gut microbiota” [106]. An animal study reported that cocoa, a source of antioxidant polyphenols, may be used to desensitize oral allergy [107]. In this study, rats administered with a 10% cocoa diet achieved oral tolerance and had a lower relative abundance of bacterial phylum Firmicutes and Proteobacteria and a higher relative abundance of Tenericutes and Cyanobacteria spp. compared to rats who received a standard diet, either orally sensitized or non-sensitized rats. It was suggested that the cocoa diet was able to increase the proportion of TCRγδ+ and CD103+CD8+ cells and decrease the proportion of CD62L+CD4+ and CD62L+CD8+ cells in mesenteric lymph nodes [108], regulate Treg cells function and reduce IgA production [107] through modulation of gut microbiota. This suggests that a cocoa diet has potential in regulating allergic immune responses in the human body and is of interest to investigate further.
Synbiotic supplementation
Synbiotics refers to a mixture of probiotics and prebiotics, designed to improve the survival of the beneficial microbiota as well as stimulate the growth of beneficial microbiota in the gastrointestinal tract [106].
A pioneer study tested a prebiotic blend of fructo-oligosaccharides with the probiotic strain Bifidobacterium breve M-16V. Infants with suspected non-IGE cow’s milk allergy were administered with a hypoallergenic, nutritionally complete amino-acid based formula either with or without the synbiotics [109]. The relative abundance of Bifidobacteria was increased in infants with cow’s milk allergy who were administered synbiotics. Moreover, the microbiota profile of these allergic infants became similar to that of healthy breastfed infants. More studies are required to confirm this therapeutic effect of synbiotics supplementation in desensitizing food allergy. It is a promising therapeutic strategy for improving the gut ecosystem and reducing food allergy responses.
Fecal microbiota transplantation
Fecal microbiota transplantation is a possible therapeutic for food allergy. Transplantation of fecal bacteria from a healthy donor to a disease recipient can re-establish gut microbiota diversity leading to the resolution of symptoms [110, 111]. As dysbiosis affects the development of food allergy [12], restoration of immune homeostasis and reconstruction of the impaired gut microbiota barrier by fecal microbiota transplantation may be able to promote the development of oral tolerance [110]. Recently, a human study has revealed that fecal microbiota transplantation is able to induce remission of infantile allergic colitis through restoration of gut microbiota diversity [112]. Although the available data in this field remain limited and the relevant scientific work has only just begun, this recent success in reducing infantile allergy colitis symptoms suggests that fecal microbiota transplantation can be a feasible strategy to arrest food allergy responses.
Synthetic stool substitute
Considering the strain specific effect of probiotic [103] and limited patient acceptance of fecal microbiota transplantation [113], a synthetic stool substitute was proposed in a pilot study [113]. In this study, a stool substitute with 33 strains was developed based on the fecal microbiota diversity of a healthy donor, also known as RePOOPulate. Clostridium difficile infection symptoms of patients were eradicated after 2–3 days of RePOOPulate treatment and this symptom free state lasted for 6 months. This synthetic stool substitute is so far only tested on two patients with Clostridium difficile infection, yet its potential benefit of reverting normal bowel pattern and restoring immune homeostasis may help to reduce allergic reactions towards food.
Microbiome-based therapy
Microbiome-based therapy can be viewed as a potential way in treating food allergy. In a mouse study, germ free mice were colonized with human feces from infants with cow’s milk allergy and age and gender matched healthy infants [114]. Healthy-colonized mice had a higher abundance of Anaerostipes caccae when compared with cow’s milk allergy colonized mice. Mice were then administered with Anaerostipes caccae in order to further investigate the role of Anaerostipes caccae in regulation of gene expression. Anaerostipes caccae were able to reduce the expression of Th2 dependent, antibody (serum BLG-specific IgE and IgG1), Acot12 expression as well as cytokine responses IL-13 and IL-4, which promoted oral tolerance towards cow’s milk allergy. This opens up a new perspective of food allergy therapy on human.
Conclusion
Studies have shown that the gut microbiota composition is vulnerable to disruptions in early life and that associated changes in host-microbiota homeostasis can cause food allergy. These studies support a regulatory role of the gut microbiota in the manifestation of food allergy, particularly in early life, but many questions remain and the underlying mechanisms are yet to be defined. The majority of our knowledge stems from animal studies and more human studies are required to validate the precise role of the gut microbiota in the development of food allergy. Evidence is emerging to suggest that therapeutic strategies in modifying gut microbiota composition are useful in the prevention, management and treatment of food allergy. Such strategies are apparent future research directions for developing prophylactic and therapeutic approaches against food allergy.
Availability of data and materials
Not applicable.
References
Tamburini S, Shen N, Wu HC, Clemente JC. The microbiome in early life: implications for health outcomes. Nat Med. 2016;22(7):713–22.
Hand TW, Vujkovic-Cvijin I, Ridaura VK, Belkaid Y. Linking the microbiota, chronic disease, and the immune system. Trends Endocrinol Metab. 2016;27(12):831–43.
Penders J, Stobberingh EE, van den Brandt PA, Thijs C. The role of the intestinal microbiota in the development of atopic disorders. Allergy. 2007;62(11):1223–36.
Sender R, Fuchs S, Milo R. Are we really vastly outnumbered? Revisiting the ratio of bacterial to host cells in humans. Cell. 2016;164(3):337–40.
Arrieta M-C, Stiemsma LT, Amenyogbe N, Brown EM, Finlay B. The intestinal microbiome in early life: health and disease. Front Immunol. 2014;5:427.
Abrahamsson TR, Jakobsson HE, Andersson AF, Bjorksten B, Engstrand L, Jenmalm MC. Low diversity of the gut microbiota in infants with atopic eczema. J Allergy Clin Immunol. 2012;129(2):434–40.
Bisgaard H, Li N, Bonnelykke K, Chawes BLK, Skov T, Paludan-Muller G, Stokholm J, Smith B, Krogfelt KA. Reduced diversity of the intestinal microbiota during infancy is associated with increased risk of allergic disease at school age. J Allergy Clin Immunol. 2011;128(3):646–52.
Hooper LV, Littman DR, Macpherson AJ. Interactions between the microbiota and the immune system. Science. 2012;336(6086):1268–73.
Thaiss CA, Levy M, Suez J, Elinav E. The interplay between the innate immune system and the microbiota. Curr Opin Immunol. 2014;26:41–8.
Bridgman SL, Kozyrskyj AL, Scott JA, Becker AB, Azad MB. Gut microbiota and allergic disease in children. Ann Allergy Asthma Immunol. 2016;116(2):99–105.
Petrus NCM, Henneman P, Venema A, Mul A, van Sinderen F, Haagmans M, Mook O, Hennekam RC, Sprikkelman AB, Mannens M. Cow’s milk allergy in Dutch children: an epigenetic pilot survey. Clin Transl Allergy. 2016;6:16.
Bunyavanich S, Shen N, Grishin A, Wood R, Burks W, Dawson P, Jones SM, Leung DYM, Sampson H, Sicherer S, Clemente JC. Early-life gut microbiome composition and milk allergy resolution. J Allergy Clin Immunol. 2016;138(4):1122–30.
Rachid RA, Gerber G, Li N, Umetsu DT, Bry L, Chatila TA. Food allergy in infancy is associated with dysbiosis of the intestinal microbiota. J Allergy Clin Immunol. 2016;137(2):235.
Savage JH, Lee-Sarwar KA, Sordillo J, Bunyavanich S, Zhou Y, O’Connor G, Sandel M, Bacharier LB, Zeiger R, Sodergren E, Weinstock GM, Gold DR, Weiss ST, Litonjua AA. A prospective microbiome-wide association study of food sensitization and food allergy in early childhood. Allergy. 2018;73(1):145–52.
Azad MB, Konya T, Guttman DS, Field CJ, Sears MR, HayGlass KT, Mandhane PJ, Turvey SE, Subbarao P, Becker AB, Scott JA, Kozyrskyj AL, Investigators CS, Allen R, Befus D, Brauer M, Brook J, Cyr M, Chen E, Daley D, Dell S, Denburg J, Elliott S, Grasemann H, Hegele R, Holness L, Kobor M, Kollmann T, Laprise C, Larche M, Lou W, Macri J, Miller G, Moqbel R, Moraes T, Pare P, Ramsey C, Ratjen F, Ritchie B, Sandford A, Scott J, Silverman F, Tebbutt S, Takaro T, Tang P, To T. Infant gut microbiota and food sensitization: associations in the first year of life. Clin Exp Allergy. 2015;45(3):632–43.
Hua X, Goedert JJ, Pu A, Yu G, Shi J. Allergy associations with the adult fecal microbiota: analysis of the American Gut Project. EBioMedicine. 2016;3:172–9.
Inoue R, Sawai T, Sawai C, Nakatani M, Romero-Perez GA, Ozeki M, Nonomura K, Tsukahara T. A preliminary study of gut dysbiosis in children with food allergy. Biosci Biotechnol Biochem. 2017;81(12):2396–9.
Ling Z, Li Z, Liu X, Cheng Y, Luo Y, Tong X, Yuan L, Wang Y, Sun J, Li L, Xiang C. Altered fecal microbiota composition associated with food allergy in infants. Appl Environ Microbiol. 2014;80(8):2546–54.
Chen C-C, Chen K-J, Kong M-S, Chang H-J, Huang J-L. Alterations in the gut microbiotas of children with food sensitization in early life. Pediatr Allergy Immunol. 2016;27(3):254–62.
Fazlollahi M, Chun Y, Grishin A, Wood RA, Burks AW, Dawson P, Jones SM, Leung DYM, Sampson HA, Sicherer SH, Bunyavanich S. Early-life gut microbiome and egg allergy. Allergy. 2018;73(7):1515–24.
Diaz M, Guadamuro L, Espinosa-Martos I, Mancabelli L, Jimenez S, Molinos-Norniella C, Perez-Solis D, Milani C, Rodriguez JM, Ventura M, Bousono C, Gueimonde M, Margolles A, Diaz JJ, Delgado S. Microbiota and derived parameters in fecal samples of infants with non-IgE cow’s milk protein allergy under a restricted diet. Nutrients. 2018;10(10):1481.
Berni Canani R, De Filippis F, Nocerino R, Paparo L, Di Scala C, Cosenza L, Della Gatta G, Calignano A, De Caro C, Laiola M, Gilbert JA, Ercolini D. Gut microbiota composition and butyrate production in children affected by non-IgE-mediated cow’s milk allergy. Sci Rep. 2018;8(1):12500.
Kourosh A, Luna RA, Balderas M, Nance C, Anagnostou A, Devaraj S, Davis CM. Fecal microbiome signatures are different in food-allergic children compared to siblings and healthy children. Pediatr Allergy Immunol. 2018;29(5):545–54.
Shi N, Li N, Duan X, Niu H. Interaction between the gut microbiome and mucosal immune system. Mil Med Res. 2017;4:14.
Lee E, Song E, Nam Y. Dysbiosis of gut microbiome and its impact on epigenetic regulation. Clin Epigenet. 2017;3(2):14.
Ipci K, Altintoprak N, Muluk NB, Senturk M, Cingi C. The possible mechanisms of the human microbiome in allergic diseases. Eur Arch Otorhinolaryngol. 2017;274(2):617–26.
Wu HJ, Wu E. The role of gut microbiota in immune homeostasis and autoimmunity. Gut Microbes. 2012;3(1):4–14.
Muir AB, Benitez AJ, Dods K, Spergel JM, Fillon SA. Microbiome and its impact on gastrointestinal atopy. Allergy. 2016;71(9):1256–63.
Rios-Covian D, Ruas-Madiedo P, Margolles A, Gueimonde M, de Los Reyes-Gavilan CG, Salazar N. Intestinal short chain fatty acids and their link with diet and human health. Front Microbiol. 2016;7:185.
den Besten G, van Eunen K, Groen AK, Venema K, Reijngoud D-J, Bakker BM. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J Lipid Res. 2013;54(9):2325–40.
Louis P, Duncan SH, McCrae SI, Millar J, Jackson MS, Flint HJ. Restricted distribution of the butyrate kinase pathway among butyrate-producing bacteria from the human colon. J Bacteriol. 2004;186(7):2099–106.
Reichardt N, Duncan SH, Young P, Belenguer A, McWilliam Leitch C, Scott KP, Flint HJ, Louis P. Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 2014;8(6):1323–35.
Goverse G, Molenaar R, Macia L, Tan J, Erkelens MN, Konijn T, Knippenberg M, Cook EC, Hanekamp D, Veldhoen M, Hartog A, Roeselers G, Mackay CR, Mebius RE. Diet-derived short chain fatty acids stimulate intestinal epithelial cells to induce mucosal tolerogenic dendritic cells. J Immunol. 2017;198(5):2172–81.
Cait A, Cardenas E, Dimitriu P, Amenyogbe N, Dai D, Cait J, Sbihi H, Stiemsma L, Subbarao P, Mandhane PJ, Becker AB, Moraes TJ, Sears MR, Lefebvre DL, Azad MB, Kollmann T, Turvey SE, Mohn WW. Reduced genetic potential for butyrate fermentation in the gut microbiome of infants who develop allergic sensitization. J Allergy Clin Immunol. 2019;144(6):1638–47.
Tan J, McKenzie C, Vuillermin PJ, Goverse G, Vinuesa CG, Mebius RE, Macia L, Mackay CR. Dietary fiber and bacterial SCFA enhance oral tolerance and protect against food allergy through diverse cellular pathways. Cell Rep. 2016;15(12):2809–24.
Asarat M, Apostolopoulos V, Vasiljevic T, Donkor O. Short-chain fatty acids regulate cytokines and Th17/Treg cells in human peripheral blood mononuclear cells in vitro. Immunol Invest. 2016;45(3):205–22.
Zhu Z, Zhu B, Hu C, Liu Y, Wang X, Zhang J, Wang F, Zhu M. Short-chain fatty acids as a target for prevention against food allergy by regulatory T cells. JGH Open. 2019;3(3):190–5.
Khan AR, Hams E, Floudas A, Sparwasser T, Weaver CT, Fallon PG. PD-L1hi B cells are critical regulators of humoral immunity. Nat Commun. 2015;6:5997.
van de Veen W, Stanic B, Wirz OF, Jansen K, Globinska A, Akdis M. Role of regulatory B cells in immune tolerance to allergens and beyond. J Allergy Clin Immunol. 2016;138(3):654–65.
Noval Rivas M, Burton OT, Wise P, Charbonnier L-M, Georgiev P, Oettgen HC, Rachid R, Chatila TA. Regulatory T cell reprogramming toward a Th2-cell-like lineage impairs oral tolerance and promotes food allergy. Immunity. 2015;42(3):512–23.
Nagata Y, Yamamoto T, Hayashi M, Hayashi S, Kadowaki M. Improvement of therapeutic efficacy of oral immunotherapy in combination with regulatory T cell-inducer kakkonto in a murine food allergy model. PLoS ONE. 2017;12(1):e0170577.
Smaldini PL, Orsini Delgado ML, Fossati CA, Docena GH. Orally-induced intestinal CD4+CD25+FoxP3+ Treg controlled undesired responses towards oral antigens and effectively dampened food allergic reactions. PLoS ONE. 2015;10(10):e0141116.
Dang TD, Allen KJ, Martino DJ, Koplin JJ, Licciardi PV, Tang ML. Food-allergic infants have impaired regulatory T-cell responses following in vivo allergen exposure. Pediatr Allergy Immunol. 2016;27(1):35–43.
Atarashi K, Tanoue T, Oshima K, Suda W, Nagano Y, Nishikawa H, Fukuda S, Saito T, Narushima S, Hase K, Kim S, Fritz JV, Wilmes P, Ueha S, Matsushima K, Ohno H, Olle B, Sakaguchi S, Taniguchi T, Morita H, Hattori M, Honda K. Treg induction by a rationally selected mixture of Clostridia strains from the human microbiota. Nature. 2013;500(7461):232–6.
Atarashi K, Tanoue T, Shima T, Imaoka A, Kuwahara T, Momose Y, Cheng G, Yamasaki S, Saito T, Ohba Y, Taniguchi T, Takeda K, Hori S, Ivanov II, Umesaki Y, Itoh K, Honda K. Induction of colonic regulatory T cells by indigenous Clostridium species. Science. 2011;331(6015):337–41.
Zhang J, Su H, Li Q, Wu H, Liu M, Huang J, Zeng M, Zheng Y, Sun X. Oral administration of Clostridium butyricum CGMCC0313-1 inhibits beta-lactoglobulin-induced intestinal anaphylaxis in a mouse model of food allergy. Gut Pathog. 2017;9:11.
Abdel-Gadir A, Stephen-Victor E, Gerber GK, Noval Rivas M, Wang S, Harb H, Wang L, Li N, Crestani E, Spielman S, Secor W, Biehl H, Dibendetto N, Dong X, Umetsu DT, Bry L, Rachid R, Chatila TA. Microbiota therapy acts via a regulatory T cell MyD88/RORgammat pathway to suppress food allergy. Nat Med. 2019;25(7):1164–74.
Ma JY, Zhang J, Li QH, Shi ZL, Wu HJ, Zhang HQ, Tang LP, Yi R, Su H, Sun X. Oral administration of a mixture of probiotics protects against food allergy via induction of CD103(+) dendritic cells and modulates the intestinal microbiota. J Funct Foods. 2019;55:65–75.
van den Elsen LW, Poyntz HC, Weyrich LS, Young W, Forbes-Blom EE. Embracing the gut microbiota: the new frontier for inflammatory and infectious diseases. Clin Transl Immunol. 2017;6(1):125.
Molloy J, Allen K, Collier F, Tang MLK, Ward AC, Vuillermin P. The potential link between gut microbiota and IgE-mediated food allergy in early life. Int J Environ Res Public Health. 2013;10(12):7235–56.
Round JL, Mazmanian SK. Inducible Foxp3+ regulatory T-cell development by a commensal bacterium of the intestinal microbiota. Proc Natl Acad Sci USA. 2010;107(27):12204–9.
Remely M, Haslberger AG. The microbial epigenome in metabolic syndrome. Mol Aspects Med. 2017;54:71–7.
Savage J, Sordillo J, Sodergren E, Weinstock G, Gold D, Weiss S, Litonjua A. A prospective microbiome-wide association study of childhood food sensitization and allergy. J Allergy Clin Immunol. 2016;137(2):AB96.
Diesner SC, Bergmayr C, Pfitzner B, Assmann V, Krishnamurthy D, Starkl P, Endesfelder D, Rothballer M, Welzl G, Rattei T, Eiwegger T, Szepfalusi Z, Fehrenbach H, Jensen-Jarolim E, Hartmann A, Pali-Scholl I, Untersmayr E. A distinct microbiota composition is associated with protection from food allergy in an oral mouse immunization model. Clin Immunol. 2016;173:10–8.
Stefka AT, Feehley T, Tripathi P, Qiu J, McCoy K, Mazmanian SK, Tjota MY, Seo GY, Cao S, Theriault BR, Antonopoulos DA, Zhou L, Chang EB, Fu YX, Nagler CR. Commensal bacteria protect against food allergen sensitization. Proc Natl Acad Sci USA. 2014;111(36):13145–50.
Guo L, Bai H, Dong Y, Huang D, Zhang X, Gong S, Zhao X, Fei P. Comparative analysis of fecal microbiota in 5–8-year-old children with and without cow milk protein allergy. Iran J Pediatr. 2016;26(6):6397.
Di Costanzo M, Amoroso A, Canani RB. Gut microbiota as a target for food allergy. J Pediatr Gastroenterol Nutr. 2016;63(1):11–3.
Strachan DP. Hay fever, hygiene, and household size. BMJ. 1989;299(6710):1259–60.
Bloomfield SF, Rook GA, Scott EA, Shanahan F, Stanwell-Smith R, Turner P. Time to abandon the hygiene hypothesis: new perspectives on allergic disease, the human microbiome, infectious disease prevention and the role of targeted hygiene. Perspect Public Health. 2016;136(4):213–24.
Sordillo JE, Zhou Y, McGeachie MJ, Ziniti J, Lange N, Laranjo N, Savage JR, Carey V, O’Connor G, Sandel M, Strunk R, Bacharier L, Zeiger R, Weiss ST, Weinstock G, Gold DR, Litonjua AA. Factors influencing the infant gut microbiome at age 3–6 months: findings from the ethnically diverse Vitamin D Antenatal Asthma Reduction Trial (VDAART). J Allergy Clin Immunol. 2017;139(2):482.e14–91.e14.
Papathoma E, Triga M, Fouzas S, Dimitriou G. Cesarean section delivery and development of food allergy and atopic dermatitis in early childhood. Pediatr Allergy Immunol. 2016;27(4):419–24.
Rutayisire E, Huang K, Liu Y, Tao F. The mode of delivery affects the diversity and colonization pattern of the gut microbiota during the first year of infants’ life: a systematic review. BMC Gastroenterol. 2016;16(1):86.
Clemente JC, Ursell LK, Parfrey LW, Knight R. The impact of the gut microbiota on human health: an integrative view. Cell. 2012;148(6):1258–70.
Bashir MEH, Louie S, Shi HN, Nagler-Anderson C. Toll-like receptor 4 signaling by intestinal microbes influences susceptibility to food allergy. J Immunol. 2004;172(11):6978–87.
Love BL, Mann JR, Hardin JW, Lu ZK, Cox C, Amrol DJ. Antibiotic prescription and food allergy in young children. Allergy Asthma Clin Immunol. 2016;12:41.
Hirsch AG, Pollak J, Glass TA, Poulsen MN, Bailey-Davis L, Mowery J, Schwartz BS. Early-life antibiotic use and subsequent diagnosis of food allergy and allergic diseases. Clin Exp Allergy. 2017;47(2):236–44.
Gupta RS, Singh AM, Walkner M, Caruso D, Bryce PJ, Wang X, Pongracic JA, Smith BM. Hygiene factors associated with childhood food allergy and asthma. Allergy Asthma Proc. 2016;37(6):140–6.
Brown K, DeCoffe D, Molcan E, Gibson D. Diet-induced dysbiosis of the intestinal microbiota and the effects on immunity and disease. Nutrients. 2012;4(8):1095–119.
Roduit C, Frei R, Depner M, Schaub B, Loss G, Genuneit J, Pfefferle P, Hyvärinen A, Karvonen A, Riedler J, Dalphin J, Pekkanen J, von Mutius R, Braun-Fahrländer C, Lauener R. Increased food diversity in the first year of life is inversely associated with allergic diseases. J Allergy Clin Immunol. 2014;133(4):1056–64.
Grummer-Strawn LM, Scanlon KS, Fein SB. Infant feeding and feeding transitions during the first year of life. Pediatrics. 2008;122:36–42.
Bode L. Human milk oligosaccharides: every baby needs a sugar mama. Glycobiology. 2012;22(9):1147–62.
Yoshioka H, Iseki K, Fujita K. Development and differences of intestinal flora in the neonatal period in breast-fed and bottle-fed infants. Pediatrics. 1983;72(3):317–21.
Walker WA, Iyengar RS. Breast milk, microbiota, and intestinal immune homeostasis. Pediatr Res. 2015;77(1–2):220–8.
Harmsen HJ, Wildeboer-Veloo AC, Raangs GC, Wagendorp AA, Klijn N, Bindels JG, Welling GW. Analysis of intestinal flora development in breast-fed and formula-fed infants by using molecular identification and detection methods. J Pediatr Gastroenterol Nutr. 2000;30(1):61–7.
Gil F, Amezqueta A, Martinez D, Aznal E, Etayo V, Dura T, Sanchez-Valverde F. Association between caesarean delivery and isolated doses of formula feeding in cow milk allergy. Int Arch Allergy Immunol. 2017;173(3):147–52.
Richards JL, Yap YA, McLeod KH, Mackay CR, Marino E. Dietary metabolites and the gut microbiota: an alternative approach to control inflammatory and autoimmune diseases. Clin Transl Immunol. 2016;5(5):e82.
Roager HM, Licht TR. Microbial tryptophan catabolites in health and disease. Nat Commun. 2018;9(1):3294.
Lanis JM, Alexeev EE, Curtis VF, Kitzenberg DA, Kao DJ, Battista KD, Gerich ME, Glover LE, Kominsky DJ, Colgan SP. Tryptophan metabolite activation of the aryl hydrocarbon receptor regulates IL-10 receptor expression on intestinal epithelia. Mucosal Immunol. 2017;10(5):1133–44.
David LA, Maurice CF, Carmody RN, Gootenberg DB, Button JE, Wolfe BE, Ling AV, Devlin AS, Varma Y, Fischbach MA, Biddinger SB, Dutton RJ, Turnbaugh PJ. Diet rapidly and reproducibly alters the human gut microbiome. Nature. 2014;505(7484):559–63.
Miyazato S, Tsuda M, Kishimoto Y, Hosono A. Suppressive effect of dietary resistant maltodextrin on systemic immunity in a mouse model of food allergy. Biosci Microbiota Food Health. 2018;38:89–95.
Smith PK, Masilamani M, Li X-M, Sampson HA. The false alarm hypothesis: food allergy is associated with high dietary advanced glycation end-products and proglycating dietary sugars that mimic alarmins. J Allergy Clin Immunol. 2017;139(2):429–37.
Velmurugan G. Gut microbiota in toxicological risk assessment of drugs and chemicals: the need of hour. Gut Microbes. 2018;9(5):465–8.
Yamashita H, Matsuhara H, Miotani S, Sako Y, Matsui T, Tanaka H, Inagaki N. Artificial sweeteners and mixture of food additives cause to break oral tolerance and induce food allergy in murine oral tolerance model for food allergy. Clin Exp Allergy. 2017;47(9):1204–13.
Jin YN, Bursley J, Dover H, Gangur V, Rockwell CE. The effect of the common food additive tBHQ in OVA-elicited food allergy. J Immunol. 2018;200(1):104.4.
Gruzieva O, Bellander T, Eneroth K, Kull I, Melen E, Nordling E, van Hage M, Wickman M, Moskalenko V, Hulchiy O, Pershagen G. Traffic-related air pollution and development of allergic sensitization in children during the first 8 years of life. J Allergy Clin Immunol. 2012;129(1):240–6.
Sbihi H, Allen RW, Becker A, Brook JR, Mandhane P, Scott JA, Sears MR, Subbarao P, Takaro TK, Turvey SE, Brauer M. Perinatal exposure to traffic-related air pollution and atopy at 1 year of age in a multi-center Canadian birth cohort study. Environ Health Perspect. 2015;123(9):902–8.
Wang IJ, Tung TH, Tang CS, Zhao ZH. Allergens, air pollutants, and childhood allergic diseases. Int J Hyg Environ Health. 2016;219(1):66–71.
Mutlu EA, Comba IY, Cho T, Engen PA, Yazici C, Soberanes S, Hamanaka RB, Nigdelioglu R, Meliton AY, Ghio AJ, Budinger GRS, Mutlu GM. Inhalational exposure to particulate matter air pollution alters the composition of the gut microbiome. Environ Pollut. 2018;240:817–30.
Barcik W, Untersmayr E, Pati-Scholl I, O’Mahony L, Frei R. Influence of microbiome and diet on immune responses in food allergy models. Drug Discov Today Ther Strateg. 2015;17–18:71–80.
Rachid R, Chatila TA. The role of the gut microbiota in food allergy. Curr Opin Pediatr. 2016;28(6):748–53.
Sidhu M, van der Poorten D. The gut microbiome. Aust Fam Physician. 2017;46(4):206–11.
Berti C, Agostoni C, Davanzo R, Hypponen E, Isolauri E, Meltzer HM, Steegers-Theunissen RP, Cetin I. Early-life nutritional exposures and lifelong health: immediate and long-lasting impacts of probiotics, vitamin D, and breastfeeding. Nutr Rev. 2017;75(2):83–97.
Berni Canani R, Sangwan N, Stefka AT, Nocerino R, Paparo L, Aitoro R, Calignano A, Khan AA, Gilbert JA, Nagler CR. Lactobacillus rhamnosus GG-supplemented formula expands butyrate-producing bacterial strains in food allergic infants. ISME J. 2016;10(3):742–50.
Thang CL, Baurhoo B, Boye JI, Simpson BK, Zhao X. Effects of Lactobacillus rhamnosus GG supplementation on cow’s milk allergy in a mouse model. Allergy Asthma Clin Immunol. 2011;7:20.
Neau E, Delannoy J, Marion C, Cottart C-H, Labellie C, Holowacz S, Butel M-J, Kapel N, Waligora-Dupriet A-J. Three novel candidate probiotic strains with prophylactic properties in a murine model of cow’s milk allergy. Appl Environ Microbiol. 2016;82(6):1722–33.
Berni Canani R, Di Costanzo M, Bedogni G, Amoroso A, Cosenza L, Di Scala C, Granata V, Nocerino R. Extensively hydrolyzed casein formula containing Lactobacillus rhamnosus GG reduces the occurrence of other allergic manifestations in children with cow’s milk allergy: 3-year randomized controlled trial. J Allergy Clin Immunol. 2017;139(6):1906–13.
Paparo L, Nocerino R, Bruno C, Di Scala C, Cosenza L, Bedogni G, Di Costanzo M, Mennini M, D’Argenio V, Salvatore F, Berni Canani R. Publisher correction: randomized controlled trial on the influence of dietary intervention on epigenetic mechanisms in children with cow’s milk allergy: the EPICMA study. Sci Rep. 2019;9(1):9504.
Jensen H, Dromtorp SM, Axelsson L, Grimmer S. Immunomodulation of monocytes by probiotic and selected lactic acid bacteria. Probiotics Antimicrob Proteins. 2015;7(1):14–23.
Tanoue T, Atarashi K, Honda K. Development and maintenance of intestinal regulatory T cells. Nat Rev Immunol. 2016;16(5):295–309.
Tang ML, Ponsonby AL, Orsini F, Tey D, Robinson M, Su EL, Licciardi P, Burks W, Donath S. Administration of a probiotic with peanut oral immunotherapy: a randomized trial. J Allergy Clin Immunol. 2015;135(3):737–44.
Fu L, Song J, Wang C, Fu S, Wang Y. Bifidobacterium infantis potentially alleviates shrimp tropomyosin-induced allergy by tolerogenic dendritic cell-dependent induction of regulatory T cells and alterations in gut microbiota. Front Immunol. 2017;8:1536.
Shi Y, Xu LZ, Peng K, Wu W, Wu R, Liu ZQ, Yang G, Geng XR, Liu J, Liu ZG, Liu Z, Yang PC. Specific immunotherapy in combination with Clostridium butyricum inhibits allergic inflammation in the mouse intestine. Sci Rep. 2015;5:17651.
Mileti E, Matteoli G, Iliev ID, Rescigno M. Comparison of the immunomodulatory properties of three probiotic strains of Lactobacilli using complex culture systems: prediction for in vivo efficacy. PLoS ONE. 2009;4(9):7056.
Scourboutakos MJ, Franco-Arellano B, Murphy SA, Norsen S, Comelli EM, L’Abbe MR. Mismatch between probiotic benefits in trials versus food products. Nutrients. 2017;9(4):400.
Liu Z, Liu W, Ran C, Hu J, Zhou Z. Abrupt suspension of probiotics administration may increase host pathogen susceptibility by inducing gut dysbiosis. Sci Rep. 2016;6:23214.
Markowiak P, Slizewska K. Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients. 2017;9(9):1021.
Camps-Bossacoma M, Perez-Cano FJ, Franch A, Castell M. Gut microbiota in a rat oral sensitization model: effect of a cocoa-enriched diet. Oxid Med Cell Longev. 2017;2017:7417505.
Camps-Bossacoma M, Abril-Gil M, Saldana-Ruiz S, Franch A, Perez-Cano FJ, Castell M. Cocoa diet prevents antibody synthesis and modifies lymph node composition and functionality in a rat oral sensitization model. Nutrients. 2016;8(4):242.
Candy DCA, Van Ampting MTJ, Oude Nijhuis MM, Wopereis H, Butt AM, Peroni DG, Vandenplas Y, Fox AT, Shah N, West CE, Garssen J, Harthoorn LF, Knol J, Michaelis LJ. A synbiotic-containing amino-acid-based formula improves gut microbiota in non-IgE-mediated allergic infants. Pediatr Res. 2018;83(3):677–86.
Borody TJ, Khoruts A. Fecal microbiota transplantation and emerging applications. Nat Rev Gastroenterol Hepatol. 2011;9(2):88–96.
Gupta S, Allen-Vercoe E, Petrof EO. Fecal microbiota transplantation: in perspective. Ther Adv Gastroenterol. 2016;9(2):229–39.
Liu S-X, Li Y-H, Dai W-K, Li X-S, Qiu C-Z, Ruan M-L, Zou B, Dong C, Liu Y-H, He J-Y, Huang Z-H, Shu S-N. Fecal microbiota transplantation induces remission of infantile allergic colitis through gut microbiota re-establishment. World J Gastroenterol. 2017;23(48):8570–81.
Petrof EO, Gloor GB, Vanner SJ, Weese SJ, Carter D, Daigneault MC, Brown EM, Schroeter K, Allen-Vercoe E. Stool substitute transplant therapy for the eradication of Clostridium difficile infection: ‘RePOOPulating’ the gut. Microbiome. 2013;1(1):3.
Feehley T, Plunkett CH, Bao R, Choi Hong SM, Culleen E, Belda-Ferre P, Campbell E, Aitoro R, Nocerino R, Paparo L, Andrade J, Antonopoulos DA, Berni Canani R, Nagler CR. Healthy infants harbor intestinal bacteria that protect against food allergy. Nat Med. 2019;25(3):448–53.
Dong P, Feng JJ, Yan DY, Lyu YJ, Xu X. Early-life gut microbiome and cow’s milk allergy—a prospective case–control 6-month follow-up study. Saudi J Biol Sci. 2018;25(5):875–80.
Nagano Y, Itoh K, Honda K. The induction of Treg cells by gut-indigenous Clostridium. Curr Opin Immunol. 2012;24(4):392–7.
Acknowledgements
We thank Professor Jane Scott at Curtin University for her careful review and suggestions for the manuscript. We would also like to acknowledge the infrastructure support of the Royal Perth Hospital Medical Research Foundation.
Funding
This work was supported by the National Key Research and Development Program of China (2017YFD0400301, 2016YFC0900803), the National Natural Science Foundation of China (81573112), Telethon Perth Children’s Hospital Research Funds and Australian Government Research Training Program (RTP) Stipend Scholarship.
Author information
Authors and Affiliations
Contributions
KHL contributed to the preparation of the manuscript and the figure. YS, WW, KY and GZ conceived the review and revised the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that no competing interests exist.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Lee, K.H., Song, Y., Wu, W. et al. The gut microbiota, environmental factors, and links to the development of food allergy. Clin Mol Allergy 18, 5 (2020). https://doi.org/10.1186/s12948-020-00120-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12948-020-00120-x