
Abstract
Background
Acinetobacter baumannii is a nosocomial pathogen which is establishing as a major cause of morbidity and mortality within the healthcare community. The success of this pathogen is largely due to its ability to rapidly gain resistance to antimicrobial therapies and its capability to persist in an abiotic environment through the production of a biofilm. Our tertiary-care hospital has showed high incidence of carbapenem-resistant Acinetobacter baumannii (CRAB) isolates.
Methods
In this study we explore both genotypic and phenotypic properties of 26 CRAB isolates: 16 isolates were collected from January 2010 to March 2011, and 10 were collected between February and May 2015.
Results
We determined that all 26 CRAB isolates possessed multiple β-lactamase genes, including genes from Groups A, C, and D. Specifically, 42% of the isolates possesses the potentially plasmid-borne genes of OXA-23-like or OXA-40-like β-lactamase. The presence of mobile gene element integron cassettes and/or integrases in 88% of the isolates suggests a possible mechanism of dissemination of antibiotic resistance genes. Additionally, the location of insertion sequence (IS) ISAba1 in promotor region of of the OXA-51-like, ADC-7, and ampC genes was confirmed. Multilocus sequence typing (MLST) demonstrated that all 26 CRAB isolates were either sequence type (ST)-229 or ST-2. Interestingly, ST-2 went from being the minority CRAB strain in the 2010–2011 isolates to the predominant strain in the 2015 isolates (from 32 to 90%). We show that the ST-2 strains have an enhanced ability to produce biofilms in comparison to the ST-229 strains, and this fact has potentially led to more successful colonization of the clinical environment over time.
Conclusions
This study provides a longitudinal genetic and phenotypic survey of two CRAB sequence types, and suggests how their differing phenotypes may interact with the selective pressures of a hospital setting effecting strain dominance over a 5-year period.
Similar content being viewed by others


Background
Acinetobacter baumannii is an opportunistic pathogen that causes a variety of nosocomial infections resulting in considerable morbidity and mortality, and presents a particular threat to intensive care unit (ICU) patients [1,2,3,4]. The number of A. baumannii infections has been increasing globally over the past three decades; particularly with regard to infections involving multi-drug resistant (MDR) isolates. The rise of MDR bacterial strains is a major cause for concern among healthcare professionals and is prompting changes in both antibiotic regiments and hospital disinfection techniques [5,6,7,8,9,10]. Genomic studies exploring outbreaks of MDR A. baumannii from geographic regions distant from one another shows a high degree of similarity among strains, indicating that most outbreaks may be due to a limited number of clonal lines [11].
The ability of A. baumannii to rapidly gain resistance to antimicrobial compounds is an important population-level virulence factor [12] for the successful dissemination of the organism within and among health-care facilities [13, 14]. Most strains of A. baumannii are innately resistant to several classes of antibiotics such as first- and second-generation cephalosporins, chloramphenicol, and aminopenicillins; in addition, the ability to both accept exogenous genetic material and overexpress endogenous resistance genes has quickly resulted in the appearance of the MDR phenotype within multiple clonal lineages [6, 15]. Historically the drug of choice to treat severe A. baumannii infections has been carbapenems, the clinical use of both meropenem and imipenem increased dramatically in the last 15 years to treat newly emerging MDR strains [1]. Likely as a result of this selective pressure carbapenem-resistant Acinetobacter baumannii (CRAB) strains are being reported with greater frequency.
In addition to antibiotic resistance, the ability to adhere and produce biofilm on both biotic and abiotic surfaces has been shown to be a virulence factor in many clinical isolates of A. baumannii and other bacterial pathogens [16,17,18,19]. The ability of clinical isolates to adhere to lung epithelial cells which is a critical step in the establishment of lung infections, and have also been documented [20,21,22]. The organism survives even on abiotic surfaces for months, and numerous retrospective epidemiological studies have shown its ability to colonize medical equipment, furniture, and healthcare personnel; this phenomenon provides the organism the capability of causing outbreaks both within and among medical institutions [23,24,25,26,27,28].
In the present study, we characterize the carbapenem resistance genotypic profile, as well as the colonization-associated phenotypes of 26 CRAB isolates, collected between 2010 and 2015, from a tertiary care hospital in Philadelphia, Pennsylvania. By analyzing these data, we aimed to determine if there was a temporal trend in resistance or colonization among these clinical isolates.
Methods
Bacterial isolates and reference strains
This study included 26 clinical isolates of carbapenem-resistant [minimum inhibitory concentration of meropenem (MIC) ≥ 8 mg/L] isolated from Hahnemann Hospital of Drexel University College of Medicine, Philadelphia, PA. Sixteen of the isolates were collected between 2010 and 2011 and were previously partially described by Sen and Joshi [29], (old isolates) and 10 isolates were collected in 2015 (New Isolates) which have not been described to date. The isolates were randomly selected, based on their MIC to meropenem. A. baumannii isolates were identified using the Vitek2 system (BioMerieux Vitek Systems Inc., USA), and confirmed by a molecular technique of OXA-51 PCR. Clinical Laboratory Standards Institute (CLSI)—recommended quality control (QC) strains were used as reference strains when necessary. QC strains included: A. baumannii ATCC 19606, A. baumannii ATCC 17978; Escherichia coli ATCC-BAA-2452, NCTC 13476; Klebsiella pneumoniae ATCC-BAA-2146, K. pneumoniae ATCC-BAA-1705, K. pneumoniae NCTC13440, and Enterobacter cloacae NCTC 13464. Additional file 1: Table S1 shows the QC strains and the reference gene for which they were used. The study was approved by the Institutional Review Board (IRB) of Drexel University, and a waiver granted.
Determination of minimum inhibitory concentration (MIC)
Carbapenem MIC results were obtained for each CRAB isolate using the VITEK 2 system and the results were confirmed by broth microdilution. All clinical breakpoints recommendations and broth microdilution protocols were followed as per Clinical Laboratory Standards Institute (CLSI) guidelines [30]. The antimicrobial agents tested were ampicillin/sulbactam, imipenem, meropenem, amikacin, levofloxacin, gentamicin, cefepime, tobramycin, trimethoprim/sulfamethoxazole and piperacillin. In routine, our hospital laboratory does not include and report polymyxin and tigecycline for this organism, and hence these agents were not included to study panel.
Polymerase chain reaction (PCR) Amplification and DNA Sequencing
Using DNeasy Blood and Tissue kit (Qiagen, Valencia, CA), genomic DNA was isolated from bacterial cells. All PCR primers targeting resistance genes and mobile elements used in this study are listed in Additional file 2: Table S2. PCR was performed using Thermo Scientific Dream Taq Green PCR Master Mix (Thermo-Fisher Scientific, Inc, Waltham, MA) using conditions specified by the reference sources. The isolates were tested for each target gene two–four times. Negative controls were included with every PCR setup to monitor for carryover [31] and rule out nonspecific amplification. Appropriate positive control strains were used to confirm the validity of each PCR primer pair (Additional file 1: Table S1). Wherever an appropriate QC strain for a particular gene was unavailable the amplicon present with the approximate size of the predicted product was purified using described method and sequences analyzed, and gene identifications were made using NCBI nucleotide BLAST tool (https://blast.ncbi.nlm.nih.gov/Blast/). The presence of IS elements (ISAba1) upstream of target genes following the method described by Sen and Joshi [29].
Meropenem-EDTA double disk synergy test (DDS)
DDS was performed as described previously by Yong et al. using 0.5 McFarland standards, and disks containing Meropenem (10 µg) alone; Meropenem (10 µg) plus 10 µL of 0.5 M EDTA; and 0.5 M EDTA alone. Appropriate control strains were included and synergistic results were interpreted as described [29, 32].
Multi-locus sequence typing (MLST)
MLST assignments were determined using the scheme provided by Pasteur Institute MLST and Whole Genome MLST Database (https://www.pasteur.fr/mlst). MLST target genes, PCR primer sequences, and annealing temperatures were all adopted from the database and optimized. The resulting PCR amplicons were run on a 1% agarose gel to determine the amplicon size and quality, and the PCR product was purified using GeneJet PCR purification kit (ThermoFisher), and sent to GeneWiz, Inc. (GeneWiz) for Sanger sequencing. The allelic identification of the sequences and strain determination was performed using the Pasteur MLST database (http://www.pubmlst.org/abaumannii/).
Biofilm formation assay
Biofilm production was determined using 96-well polystyrene plates as described by O’Toole et al., with minor modifications [33]. Briefly, overnight cultures were grown in LB broth at 37 °C in an aerobic incubator. The following day, 200 µL LB broth diluted cultures (1:100) were added to multiple wells and the plates incubated at 37 °C in a static aerobic incubator for either 24 or 48 h. At the appropriate time the optical density was read at 595 nm, then each well received 30 µL of 0.1% Crystal Violet solution to incubate for 1 h at room temperature. After incubation, the plates were washed with tap water three times and allowed to dry at room temperature for 1 h. Then 250 µL of ethanol:acetone (4:1) solution was added to each well to solubilize the remaining stain. The optical density was then read at 570 nm, and the biofilm production was quantified by taking the OD570:OD590 ratio. The ratio was corrected with negative control (LB alone), and biofilm forming capability of the isolates considered if the OD570:OD590 ratio was a positive value. Experiments were run in two separate assays with a total of sixteen replicates each.
Results
Characterization of isolates
A total of 26 CRAB isolates were analyzed in this study, all collected from Hahnemann Hospital. Sixteen isolates were collected during 2010–2011 (old isolates), and the additional 10 isolates were collected in 2015 (new isolates). The relevant descriptive features are summarized in Table 1. The average age of all the patients from which the isolates were collected was 50.1 (± 17.7) years, with a range of 19–84 years. Of the 26 total patients 15 (57%) were male. A total of 14 (54%) patients were admitted in the ICU during the time of collection, and the other 12 (46%) patients were being treated for various conditions in non-ICU in-patient rooms in the hospital. A total of 12 (43%) patients’ infections were considered nosocomial in origin, in which the first positive A. baumannii culture was reported ≥ 48 h post-admission [34].
Antibiotic susceptibility
The findings of the antibiotic susceptibility testing for all isolates are showed in Additional file 3: Table S3. The antibiogram patterns of all 26 CRAB isolates by isolate groups (2010–2011 versus 2015), clinical source, and MLST sequence types are presented in Table 2. All 26 (100%) of the CRAB isolates met the clinical definition of the MDR A. baumannii phenotype, defined as being non-susceptible to at least 3 classes of antibiotics [34]. All isolates (100%) were resistant or intermediately resistant to carbapenems (meropenem and imipenem), as well as Levofloxacin (fluoroquinolone). More than 80% of the isolates were resistant or intermediately resistant to aminoglycosides, cephalosporins, extended-spectrum penicillins, and folate inhibitors.
Multi-locus sequence typing (MLST)
MLST was performed for all 26 CRAB isolates as described in Methods. A total of 12 (46%) isolates were determined to be ST-229 and 14 (54%) were determined to be ST-2. In the 2010–2011 isolate group, 11 (68%) of the 16 isolates were determined to be members of ST-229, while the remaining 5 (32%) isolates were identified as ST-2. In the 2015 isolate group, 9 (90%) of the isolates were identified as strain 2, and only 1 (10%) were determined to be ST 229 (Additional file 4: Figure S1). The isolates and their MLST sequence types are summarized in Table 3.
Extended-spectrum β-lactamase (ESBL) genotypic analysis
Table 3 summarizes all the ESBL genes explored in this study. All CRAB isolates, and A. baumannii controls carried the Group D oxacillinase bla OXA-51-like gene (Additional file 4: Figure S1), while no isolates contained the bla OXA-24 or bla OXA-58-like gene. Two isolates, one from the 2010–2011 isolate group and one from the 2015 isolate group, possessed the bla OXA-40-like gene (Fig. 1). Both bla OXA-40-like positive isolates had the same MLST sequence type with a similar genotypic profile. Similarly, 9 (34%) isolates had bla OXA-23-like genes (one from the 2010–2011 isolates and eight from the 2015 isolates) had the similar genotypic profiles. All of the CRAB isolates contained the Group A β-lactamase gene bla TEM, and 11 (42%) of the isolates had bla SHV. The bla SHV positive isolates were all ST-229, suggesting a similar clonal linage. All of the isolates were negative for other Group A β-lactamase genes including bla PER, bla VEB, as well as the most commonly reported clinical subtypes of bla KPC, and bla CTX-M. Additionally all isolates were negative for all of the Group B metallo-β-lactamases (MBLs) genes tested which included, bla Imp, bla VIM, bla GIM, bla SPM, bla SIM, bla DIM, and bla NDM-1. All of the CRAB isolates tested contained the Group C cephalosporinase bla ampC gene, while 20 (76%) also possessed the cephalosporinase bla ADC-7 gene.

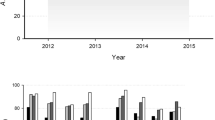
Biofilm production of CRAB isolates at 24 h (48 h data not shown) in 96-well polystyrene plate using Crystal Violet staining method. a Biofilm production with isolates grouped as old and new isolates (*p = 0.03). b Isolates grouped by clinical source exhibited non-significant value, may be due to lack of clonal heterogeneity. c CRAB isolates showing biofilm production, grouped by MLST sequence type/strain type (*p = 0.04)
Mobile genes and insertion sequences
As described by Koeleman et al., the presence of mobile genetic elements is of particular interest as they provide strong evidence for the horizontal dissemination of antibiotic resistance genes [35]. The two mobile gene elements explored in this study were the class 1 integron cassette (ICA1) and three integrase coding genes (int123) as described in Hujer et al. [36]. A total of 11 (42%) of the CRAB isolates contained ICA1, and 21 (80%) of the isolates had int123. Only two isolates #58 (bla OXA-40-like positive) and #64 (bla OXA-23-like positive) from the 2010–2011 isolates were negative for both mobile gene elements, while all 10 (100%) of the 2015 CRAB isolates were positive for one or both of the mobile gene elements (Table 3). These data suggest that the majority of the CRAB isolates in present study may have ability to transfer genetic elements through integrons, and that these mobile elements may contain antibiotic resistance genes.
In addition, we detected IS elements in the promoter regions of several antibiotic resistance genes. A total of 25 (96%) of the CRAB isolates examined in this study were found to have the ISAba1 element upstream of OXA-51 (Additional file 5: Figure S2; Table 3), and one negative isolate #58 also carried ISAba1 in the non-promoter region of OXA-51 (this isolate also processes the bla OXA-40-like gene). In addition, ISAba1 was found upstream of the ampC cephalosporinases in 92% of the isolates, and upstream of the ADC-7 gene in 76% of the isolates.
Biofilm production
In vitro biofilm production assays were performed on all CRAB isolates (Fig. 1) with 9 of 26 (34%) showing varying degrees of biofilm production. Figure 1a shows the 2015 isolates includes more biofilm producers than the 2010–2011 isolates (p = 0.03). There was no significant association found between biofilm production and the source of the isolate (Fig. 1b). Evaluation of biofilm production between the two MLST strain types demonstrated that the ST-2 strains were significantly better biofilm producers than the ST-229 strains (p = 0.04) (Fig. 1c).
Discussion
The present study was undertaken to determine whether any patterns of resistance and fitness within our clinical CRAB isolate groups evolved or expanded over time. The unique aspect to this study is the two isolate groups we used were collected 4–5 years apart. This fact allowed us to have a look longitudinally at the genotypes and phenotypes of antibiotic-resistant A. baumannii isolates, and to potentially identify traits that make certain isolates successful pathogens under the selective pressures of a hospital setting.
From a clinical perspective, all the isolates examined in this study displayed the MDR phenotype [34]. We aimed to find the genetic mechanisms conferring these resistances in our isolates by exploring the presence of a large number of β-lactamase encoding genes. Metallo-β-lactamase positive A. baumannii isolates are rare in the United States, and we did not find isolate carrying MBL gene; however globally, reports detecting their appearance are increasing in frequency [37,38,39]. The Group A β-lactamases encoded by bla TEM and bla SHV genes has been shown to hydrolyze both penicillins, as well as first- and second- generation cephalosporins [40]. Interestingly there seems to be a distinction with regard to ST type with regard to possession of the bla SHV gene wherein the bla SHV gene was found in 10 (83%) of ST-229 isolates, and 1 (7%) of the ST-2 isolates. Although bla ampC and bla ADC-7 genes encode cephalosporinases were present, the later (bla ADC-7 was predominant (in 76% of isolates), which have been shown to be inducible [41, 42].
The bla OXA-51-like allele is chromosomal and has been reported as an intrinsic gene of the A. baumannii species [43]. Both the bla OXA-23-like and bla OXA-40-like genes can be either plasmid or chromosome borne, and have an increased risk of horizontal dissemination, resulting in increased rates of resistance in healthcare settings [44, 45]. In our isolate groups, only 2 (7%) isolates contained the bla OXA-40-like gene; interestingly these two isolates display the greatest phenotypic resistance to meropenem (≥ 256 μg/mL), while 9 (34%) of our CRAB isolates possess the bla OXA-23-like gene, and displayed MICs between 32–64 μg/mL. These two oxacillinase genes were only found in ST-2 isolates in among the 2010–2011 isolate group; but seem to have been successfully disseminated into the hospital setting overtime as they were found in 9 (90%) of the 2015 isolate group.
A major concern in the healthcare community is the acquisition of genes such OXA-23-like and OXA-40-like through mobile genetic element transfers [46]. We did not determine whether these mobile elements such as integron-encoded integrase genes flank resistance genes, but their presence in majority (80%) of proposed CRAB suggests that those isolates are capable of acquiring and donating virulence genes by recombination.
An interesting finding from this study was the presence of IS elements upstream of several β-lactamase genes. It has been previously shown that the insertion of the IS element ISAba1 upstream of the OXA-51 gene changes the expression level leading to increased antimicrobial resistance [43]. Except isolate #58, which was co-carrying blaOXA-40 like gene), every isolate examined in this study was found to have ISAba1 upstream of the bla OXA-51-like gene (Additional file 5: Figure S2), and suggesting that the isolates may have additional mechanisms resistance against carbopenem. The ISAba1 was also present in promoter region of bla ampC gene (92%) and bla ADC-7 gene (76%) of the isolates, and suggest the overexpression of the downstream cephalosporinase genes resulting in an enhanced resistance phenotype.
MLST typing showed differing prevalence of the two STs of resistant strains of A. baumannii throughout the hospital over time. A comparison suggests that the majority of the isolates of 2010–2011 group were the members of ST-229, and only 5 (32%) isolates were ST-2, whereas interestingly, 90% of the isolates of 2015 group were ST-2, and only (10%) determined as ST-229. These data indicate over the course of 4–5 years that the ST-2 has successfully replaced most of the ST-229 strains to become the predominant resistant type within our hospital.
Why is one strain type more successful than the other when it appears the two strain types share similar genetic resistance mechanisms? It has been previously reported that biofilm production is an important virulence factor for A. baumannii as it may aid colonization and survival on abiotic hospital environments [47]. Thus, we explored the ability of our CRAB isolates to produce biofilm on an abiotic (polystyrene) surface, to give an indication of how well these strains might colonize both medical equipment and objects in the clinical environment. We found that there were no significant differences between the clinical source of the isolates or their ability to produce biofilms (Fig. 1b). Other studies using more clonally diverse strains have shown enhanced biofilm producing isolates derived from urine and sputum cultures [48, 49]. Therefore, the non-significant result in our isolate group may be due to a lack of clonal heterogeneity. When we grouped our isolates by isolate groups (2010–2011 versus 2015) we found the 2015 isolate group had an enhanced ability to produce biofilm over the old isolates (Fig. 1a). Due to the predominance of ST-2 in the 2015 isolate group we explored biofilm production by MLST strain type, wherein the ST-2 showed an increased biofilm production compared to ST-229 (Fig. 1c). Therefore, a possible explanation for the success of ST-2 over ST-229 may be its enhanced ability to produce biofilms resulting in a more effective colonization within the hospital, even when the two strains exhibit similar antibiotic resistance phenotypes.
Conclusion
The temporal nature of the isolates used in this study provided a unique opportunity to epidemiologically monitor multi-drug-resistant A. baumannii isolates from the same hospital over time. We have shown that all of the CRAB isolates in our hospital belong to either MLST strain type ST-2 or ST-229 and that these two strain types most likely share similar mechanisms of resistance with possibly ISAba1-dependent overexpression of β-lactamase genes, resulting in an MDR phenotype. Interestingly over a 4–5 years’ timeframe the ST-2 type replaced the ST229 type to become the predominate CRAB strain within the hospital which we believe is due to its enhanced ability to produce biofilms leading to improved colonization and survival on abiotic surfaces. To our knowledge, this is the first study to comparatively characterize the genetic and phenotypic properties of clinical MDR A. baumannii isolates from a US hospital over an extended time frame.
References
Bergogne-Berezin E, Towner KJ. Acinetobacter spp. as nosocomial pathogens: microbiological, clinical, and epidemiological features. Clin Microbiol Rev. 1996;9(2):148–65.
Seifert H, Strate A, Pulverer G. Nosocomial bacteremia due to Acinetobacter baumannii. Clinical features, epidemiology, and predictors of mortality. Medicine (Baltimore). 1995;74(6):340–9.
Falagas ME, Bliziotis IA, Siempos II. Attributable mortality of Acinetobacter baumannii infections in critically ill patients: a systematic review of matched cohort and case-control studies. Crit Care. 2006;10(2):R48.
Peleg AY, Seifert H, Paterson DL. Acinetobacter baumannii: emergence of a successful pathogen. Clin Microbiol Rev. 2008;21(3):538–82.
Vila J, Marcos A, Marco F, Abdalla S, Vergara Y, Reig R, Gomez-Lus R, Jimenez de Anta T. In vitro antimicrobial production of beta-lactamases, aminoglycoside-modifying enzymes, and chloramphenicol acetyltransferase by and susceptibility of clinical isolates of Acinetobacter baumannii. Antimicrob Agents Chemother. 1993;37(1):138–41.
Perez F, Hujer AM, Hujer KM, Decker BK, Rather PN, Bonomo RA. Global challenge of multidrug-resistant Acinetobacter baumannii. Antimicrob Agents Chemother. 2007;51(10):3471–84.
Jones RN, Deshpande L, Fritsche TR, Sader HS. Determination of epidemic clonality among multidrug-resistant strains of Acinetobacter spp. and Pseudomonas aeruginosa in the MYSTIC Programme (USA, 1999–2003). Diagn Microbiol Infect Dis. 2004;49(3):211–6.
Landman D, Quale JM, Mayorga D, Adedeji A, Vangala K, Ravishankar J, Flores C, Brooks S. Citywide clonal outbreak of multiresistant Acinetobacter baumannii and Pseudomonas aeruginosa in Brooklyn, NY: the pre antibiotic era has returned. Arch Intern Med. 2002;162(13):1515–20.
Peleg AY, Franklin C, Bell JM, Spelman DW. Emergence of carbapenem resistance in Acinetobacter baumannii recovered from blood cultures in Australia. Infect Control Hosp Epidemiol. 2006;27(7):759–61.
Yang HY, Lee HJ, Suh JT, Lee KM. Outbreaks of imipenem resistant Acinetobacter baumannii producing OXA-23 beta-lactamase in a tertiary care hospital in Korea. Yonsei Med J. 2009;50(6):764–70.
Nemec A, Dijkshoorn L, van der Reijden TJ. Long-term predominance of two pan-European clones among multi-resistant Acinetobacter baumannii strains in the Czech Republic. J Med Microbiol. 2004;53(Pt 2):147–53.
Hu FZ, Ehrlich GD. Population-level virulence factors amongst pathogenic bacteria: relation to infection outcome. Future Microbiol. 2008;3(1):31–42.
Gaynes R, Edwards JR. Overview of nosocomial infections caused by gram-negative bacilli. Clin Infect Dis. 2005;41(6):848–54.
Urban C, Segal-Maurer S, Rahal JJ. Considerations in control and treatment of nosocomial infections due to multidrug-resistant Acinetobacter baumannii. Clin Infect Dis. 2003;36(10):1268–74.
Seifert H, Baginski R, Schulze A, Pulverer G. Antimicrobial susceptibility of Acinetobacter species. Antimicrob Agents Chemother. 1993;37(4):750–3.
King LB, Swiatlo E, Swiatlo A, McDaniel LS. Serum resistance and biofilm formation in clinical isolates of Acinetobacter baumannii. FEMS Immunol Med Microbiol. 2009;55(3):414–21.
Gurung J, Khyriem AB, Banik A, Lyngdoh WV, Choudhury B, Bhattacharyya P. Association of biofilm production with multidrug resistance among clinical isolates of Acinetobacter baumannii and Pseudomonas aeruginosa from intensive care unit. Indian J Crit Care Med. 2013;17(4):214–8.
Yun HC, Kreft RE, Castillo MA, Ehrlich GD, Guymon CH, Crouch HK, Chung KK, Wenke JC, Hsu JR, Spirk TL, et al. Comparison of PCR/electron spray ionization-time-of-flight-mass spectrometry versus traditional clinical microbiology for active surveillance of organisms contaminating high-use surfaces in a burn intensive care unit, an orthopedic ward and healthcare workers. BMC Infect Dis. 2012;12:252.
Aul JJ, Anderson KW, Wadowsky RM, Doyle WJ, Kingsley LA, Post JC, Ehrlich GD. Comparative evaluation of culture and PCR for the detection and determination of persistence of bacterial strains and DNAs in the Chinchilla lanigera model of otitis media. Ann Otol Rhinol Laryngol. 1998;107(6):508–13.
Lee JC, Koerten H, van den Broek P, Beekhuizen H, Wolterbeek R, van den Barselaar M, van der Reijden T, van der Meer J, van de Gevel J, Dijkshoorn L. Adherence of Acinetobacter baumannii strains to human bronchial epithelial cells. Res Microbiol. 2006;157(4):360–6.
Lee HW, Koh YM, Kim J, Lee JC, Lee YC, Seol SY, Cho DT, Kim J. Capacity of multidrug-resistant clinical isolates of Acinetobacter baumannii to form biofilm and adhere to epithelial cell surfaces. Clin Microbiol Infect. 2008;14(1):49–54.
de Breij A, Gaddy J, van der Meer J, Koning R, Koster A, van den Broek P, Actis L, Nibbering P, Dijkshoorn L. CsuA/BABCDE-dependent pili are not involved in the adherence of Acinetobacter baumannii ATCC19606(T) to human airway epithelial cells and their inflammatory response. Res Microbiol. 2009;160(3):213–8.
van den Broek PJ, Arends J, Bernards AT, De Brauwer E, Mascini EM, van der Reijden TJ, Spanjaard L, Thewessen EA, van der Zee A, van Zeijl JH, et al. Epidemiology of multiple Acinetobacter outbreaks in The Netherlands during the period 1999–2001. Clin Microbiol Infect. 2006;12(9):837–43.
Wilks M, Wilson A, Warwick S, Price E, Kennedy D, Ely A, Millar MR. Control of an outbreak of multidrug-resistant Acinetobacter baumannii–calcoaceticus colonization and infection in an intensive care unit (ICU) without closing the ICU or placing patients in isolation. Infect Control Hosp Epidemiol. 2006;27(7):654–8.
Catalano M, Quelle LS, Jeric PE, Di Martino A, Maimone SM. Survival of Acinetobacter baumannii on bed rails during an outbreak and during sporadic cases. J Hosp Infect. 1999;42(1):27–35.
Simor AE, Lee M, Vearncombe M, Jones-Paul L, Barry C, Gomez M, Fish JS, Cartotto RC, Palmer R, Louie M. An outbreak due to multiresistant Acinetobacter baumannii in a burn unit: risk factors for acquisition and management. Infect Control Hosp Epidemiol. 2002;23(5):261–7.
El Shafie SS, Alishaq M, Leni Garcia M. Investigation of an outbreak of multidrug-resistant Acinetobacter baumannii in trauma intensive care unit. J Hosp Infect. 2004;56(2):101–5.
Jawad A, Heritage J, Snelling AM, Gascoyne-Binzi DM, Hawkey PM. Influence of relative humidity and suspending menstrua on survival of Acinetobacter spp. on dry surfaces. J Clin Microbiol. 1996;34(12):2881–7.
Sen B, Joshi SG. Studies on Acinetobacter baumannii involving multiple mechanisms of carbapenem resistance. J Appl Microbiol. 2016;120(3):619–29.
CLSI. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; approved standard-ninth edition. In., vol. CLSI document M07-A9. 9th ed. Wayne: Clinical and Laboratory Standards Institute; 2012. p. 697.
Ehrlich GD, Greenberg SJ. PCR-based diagnostics in infectious disease. Boston: Blackwell Scientific Publications; 1994. p. 697.
Yong D, Lee K, Yum JH, Shin HB, Rossolini GM, Chong Y. Imipenem-EDTA disk method for differentiation of metallo-beta-lactamase-producing clinical isolates of Pseudomonas spp. and Acinetobacter spp. J Clin Microbiol. 2002;40(10):3798–801.
O’Toole GA. Microtiter dish biofilm formation assay. J Vis Exp. 2011;(47). pii: 2437. http://doi.org/10.3791/2437.
Magiorakos AP, Srinivasan A, Carey RB, Carmeli Y, Falagas ME, Giske CG, Harbarth S, Hindler JF, Kahlmeter G, Olsson-Liljequist B, et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin Microbiol Infect. 2012;18(3):268–81.
Koeleman JG, Stoof J, Van Der Bijl MW, Vandenbroucke-Grauls CM, Savelkoul PH. Identification of epidemic strains of Acinetobacter baumannii by integrase gene PCR. J Clin Microbiol. 2001;39(1):8–13.
Hujer KM, Hujer AM, Hulten EA, Bajaksouzian S, Adams JM, Donskey CJ, Ecker DJ, Massire C, Eshoo MW, Sampath R, et al. Analysis of antibiotic resistance genes in multidrug-resistant Acinetobacter sp. isolates from military and civilian patients treated at the Walter Reed Army Medical Center. Antimicrob Agents Chemother. 2006;50(12):4114–23.
Davies TA, Marie Queenan A, Morrow BJ, Shang W, Amsler K, He W, Lynch AS, Pillar C, Flamm RK. Longitudinal survey of carbapenem resistance and resistance mechanisms in Enterobacteriaceae and non-fermenters from the USA in 2007–09. J Antimicrob Chemother. 2011;66(10):2298–307.
Chen Y, Zhou Z, Jiang Y, Yu Y. Emergence of NDM-1-producing Acinetobacter baumannii in China. J Antimicrob Chemother. 2011;66(6):1255–9.
Wei WJ, Yang HF, Ye Y, Li JB. New Delhi metallo-beta-lactamase-mediated carbapenem resistance: origin, diagnosis, treatment and public health concern. Chin Med J (Engl). 2015;128(14):1969–76.
Bradford PA. Extended-spectrum beta-lactamases in the 21st century: characterization, epidemiology, and detection of this important resistance threat. Clin Microbiol Rev. 2001;14(4):933–51 (table of contents).
Harris PN, Ferguson JK. Antibiotic therapy for inducible AmpC beta-lactamase-producing Gram-negative bacilli: what are the alternatives to carbapenems, quinolones and aminoglycosides? Int J Antimicrob Agents. 2012;40(4):297–305.
Hujer KM, Hamza NS, Hujer AM, Perez F, Helfand MS, Bethel CR, Thomson JM, Anderson VE, Barlow M, Rice LB, et al. Identification of a new allelic variant of the Acinetobacter baumannii cephalosporinase, ADC-7 beta-lactamase: defining a unique family of class C enzymes. Antimicrob Agents Chemother. 2005;49(7):2941–8.
Turton JF, Ward ME, Woodford N, Kaufmann ME, Pike R, Livermore DM, Pitt TL. The role of ISAba1 in expression of OXA carbapenemase genes in Acinetobacter baumannii. FEMS Microbiol Lett. 2006;258(1):72–7.
Turton JF, Kaufmann ME, Glover J, Coelho JM, Warner M, Pike R, Pitt TL. Detection and typing of integrons in epidemic strains of Acinetobacter baumannii found in the United Kingdom. J Clin Microbiol. 2005;43(7):3074–82.
Lolans K, Rice TW, Munoz-Price LS, Quinn JP. Multicity outbreak of carbapenem-resistant Acinetobacter baumannii isolates producing the carbapenemase OXA-40. Antimicrob Agents Chemother. 2006;50(9):2941–5.
Richet H, Fournier PE. Nosocomial infections caused by Acinetobacter baumannii: a major threat worldwide. Infect Control Hosp Epidemiol. 2006;27(7):645–6.
Badave GK, Kulkarni D. Biofilm producing multidrug resistant Acinetobacter baumannii: an emerging challenge. J Clin Diagn Res. 2015;9(1):Dc08–10.
Duarte A, Ferreira S, Almeida S, Domingues FC. Clinical isolates of Acinetobacter baumannii from a Portuguese hospital: PFGE characterization, antibiotic susceptibility and biofilm-forming ability. Comp Immunol Microbiol Infect Dis. 2016;45:29–33.
Zhang D, Xia J, Xu Y, Gong M, Zhou Y, Xie L, Fang X. Biological features of biofilm-forming ability of Acinetobacter baumannii strains derived from 121 elderly patients with hospital-acquired pneumonia. Clin Exp Med. 2016;16(1):73–80.
Authors’ contributions
The authors KMR and SGJ have hypothesized of mechanisms, designed experiments, analyzed data, and written manuscript. BS, NL, TEB and CLE helped in samples collection, phenotypic verification of some of the members of old isolates, and clinical clubbing for isolate group. KMR, BS and SGJ designed primers and evaluated PCR products, and KMR carried out the experiments, GDE and SGJ verified MLST sequencing data and helped in analyses to KMR. GDE participated in active discussions and data interpretations, and critical editing of the manuscript. All authors read and approved the final manuscript.
Acknowledgements
The research has IRB approval from Drexel University College of Medicine. The technical help provided by clinical laboratory of associated hospital and microbiology immunology department is respectfully acknowledged. This research was a part of graduate (PhD) rotation of Kevin M. Raible in Joshi Laboratory, and was partly presented at American Society for Microbiology-2016 Annual Meeting, and International Symposium of Drexel University.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
All relevant data presented in this manuscript.
Consent for publication
All authors have approved this manuscript.
Ethics approval and consent to participate
This work was approved by the Institutional Review Board of Drexel University, College of Medicine, Philadelphia, PA, USA.
Funding
This research was carried out the intramural support, and as a continuation of Surgical Infection Research Program of the Departments of Surgery, and Microbiology and Immunology.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author information
Authors and Affiliations
Corresponding author
Additional files
12941_2017_248_MOESM3_ESM.pdf
Additional file 3: Table S3. Antimicrobial typing of A. baumannii study isolates, showing individual isolates against commonly used drugs.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Raible, K.M., Sen, B., Law, N. et al. Molecular characterization of β-lactamase genes in clinical isolates of carbapenem-resistant Acinetobacter baumannii . Ann Clin Microbiol Antimicrob 16, 75 (2017). https://doi.org/10.1186/s12941-017-0248-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12941-017-0248-3