
Abstract
Background
Staphylococcus aureus (S. aureus) has established itself over the years as a major cause of morbidity and mortality both within the community and in healthcare settings. Methicillin resistant S. aureus (MRSA) in particular has been a major cause of nosocomial infections resulting in significant increase in healthcare costs. In Africa, the MRSA prevalence has been shown to vary across different countries. In order to better understand the epidemiology of MRSA in a setting, it is important to define its population structure using molecular tools as different clones have been found to predominate in certain geographical locations.
Methods
We carried out PFGE, MLST, SCCmec and spa typing of selected S. aureus isolates from a private and public referral hospital in Nairobi, Kenya.
Results
A total of 93 S. aureus isolates were grouped into 19 PFGE clonal complexes (A–S) and 12 singletons. From these, 55 (32 MRSA and 23 MSSA) representative isolates from each PFGE clonal complex and all singletons were spa typed. There were 18 different MRSA spa types and 22 MSSA spa types. The predominant MRSA spa type was t037 comprising 40.6 % (13/32) of all MRSA. In contrast, the MSSA were quite heterogeneous, only 2 out of 23 MSSA shared the same spa type. Two new MRSA spa types (t13149 and t13150) and 3 new MSSA spa types (t13182, t13193 and t13194) were identified. The predominant clonal complex was CC 5 which included multi-locus sequence types 1, 8 and 241.
Conclusion
In contrast to previous studies published from Kenya, there’s marked genetic diversity amongst clinical MRSA isolates in Nairobi including the presence of well-known epidemic MRSA clones. Given that these clones are resident within our referral hospitals, adherence to strict infection control measures needs to be ensured to reduce morbidity and mortality associated with hospital acquired MRSA infections.
Similar content being viewed by others


Background
Staphylococcus aureus (S. aureus) has established itself over the years as a major cause of morbidity and mortality globally both within the community and in healthcare settings [1–3]. Its ability to cause disease is aided not only by its impressive repertoire of virulence factors but also its ability to develop resistance to antibiotics used in its treatment epitomized by the emergence of methicillin resistant S. aureus (MRSA). Methicillin resistance is conferred by the mecA gene that is carried on a staphylococcal cassette chromosome mec (SCCmec) and codes for a modified penicillin binding protein (PBP2a). This binding protein has reduced affinity to all beta-lactam and beta-lactam/beta-lactamase inhibitor combination antibiotics [4, 5]. In Africa, the MRSA prevalence has been shown to vary across different countries with a prevalence as low as 7 % reported in Madagascar and as high as 82 % in Egypt [6]. This marked variation could be due to different environmental determinants or simply due to a difference in the genetic diversity of S. aureus. In Kenya, there is a marked difference in reported MRSA prevalence in clinical isolates within Nairobi with one recent study reporting a prevalence of 3.7 % while another reported 87.2 % [7, 8]. In order to better understand the epidemiology of MRSA, it is important to define its population structure. Molecular characterization helps in identifying clonal populations which can help in surveillance and investigation of outbreaks.
There is a growing interest in the characterization of MRSA isolates and this stems primarily from its role as a major cause of hospital and community acquired infections [1, 9, 10]. There are various molecular methods used, the more common ones include multi-locus sequence typing (MLST), pulse field gel electrophoresis (PFGE), staphylococcal protein A (spa) typing and SCCmec typing [11]. Despite S. aureus having a very diverse clonal population, MLST studies have shown that a small set of clonal complexes (CC) are associated with most of the MRSA epidemics. These include CC5, CC22, CC30, CC45 and CC80 [6, 12, 13]. A clonal complex can have several sequence types, however the multi-locus sequence types that are regarded as the founders in these clonal complexes are ST5, ST22, ST30, ST45 and ST80 respectively [14]. As regards spa types, it has been shown that particular ones are more predominant in certain regions. For example t030 is quite predominant in hospitals in Turkey [15], t042 and t044 are more common in North Africa while t008 is common in the US [16]. Unfortunately, the molecular epidemiology of MRSA in Africa is not very well described. Most of the studies carried out in Africa characterizing MRSA have emanated from a few countries namely Tunisia, Nigeria, South Africa, Algeria and Egypt [6]. There are very few studies from East Africa that have reported on the molecular characterization of S. aureus presumably due to lack of readily available technical expertise and laboratory facilities. A study done in Kenya looking at carriage of S. aureus by inpatients in a government hospital found that only 6 out of 86 (7 %) S. aureus isolates were MRSA and they all belonged to the same clone (MLST ST239; spa type t037) [17]. This clone is a globally distributed hybrid of ST8 and ST30 and is known to be responsible for several outbreaks in different continents [18–21]. The only other study from Kenya did not report on spa or multi locus sequence types [8].
We set out to characterize selected S. aureus isolates from different hospitals in Nairobi, Kenya in order to identify which clonal lineages are present and further shed light on the molecular epidemiology of both MSSA and MRSA in Kenya.
Methods
We obtained archived methicillin susceptible (MSSA) and MRSA isolates from 2 hospitals in Nairobi, Kenya collected between January 2010 and July 2013. The hospitals included a government hospital whose samples we obtained through the Kenya Medical Research Institute (KEMRI) and the Aga Khan University Hospital Nairobi (AKUHN) which is a private referral hospital with a network of satellite clinics and laboratories spread in and around Nairobi as well as different parts of the country. The isolates from the government hospital were part of a previous study done to determine prevalence of MRSA carriage in a paediatric ward and the rest of the isolates were from clinical specimens submitted to the AKUHN laboratory for routine culture and sensitivity. These were convenience isolates that were not collected through a well-structured, formal and documented process. All isolates were stored at −80 °C and grown overnight on sheep blood agar plates at 37 °C.
S. aureus identification
All isolates were confirmed to be S. aureus using routine bench identification methods which included growth characteristics on sheep blood agar, gram stain, catalase, coagulase, deoxyribonuclease (DNase) and mannitol fermentation tests. A cefoxitin screen using a 30 µg disc (Oxoid, United Kingdom) was performed to distinguish MSSA from MRSA. Isolates with a diameter ≤21 mm were classified as MRSA.
Antibiotic susceptibility
Antibiotic susceptibility was only available for the MRSA isolates obtained from AKUHN. These were performed on Vitek 2 (version 4.01, bioMerieux, Marcy-l’Etoile, France) an automated bacterial identification system that performs antibiotic susceptibility using broth dilution and interpretation based on Clinical Laboratory Standards Institute (CLSI) antimicrobial susceptibility guidelines [22]. Multidrug resistance (MDR) was defined as resistance to three or more drug classes.
DNA derivation
Isolates were grown on blood agar plates (National Health Laboratory Services Media Lab, Cape Town, South Africa) at 37 °C overnight. After incubation, 4–5 large colonies were re-suspended in 200 µL nuclease free water. The samples were incubated at 95 °C for 30 min, followed by −80 °C for 30 min and centrifuged for 10 min at 14,000×g when thawed. The supernatant containing DNA was carefully aspirated without disturbing the pellet of cell debris and stored as DNA aliquots at −20° C until further use.
PFGE
PFGE based on SmaI macrorestriction analysis was performed using the CDC laboratory protocol for S. aureus [23]. The PFGE was run on a CHEF DR III system (Bio-Rad, California, United States of America) with optimum settings as follows: initial 5 s, switch 30 s, run time 29 h, voltage 6 V/cm and a SeaKem Gold agarose (Lonza, Rockland, USA) gel concentration of 1.4 %. S. aureus NCTC 8325 was used as a control in each gel run. Gels were visualized an Alliance 2.7 (UVItec, Cambridge, United Kingdom) gel documentation system after staining with 10 mg/mL ethidium bromide. Analysis of PFGE clusters was performed using the BioNumerics software package (Applied Maths, Sint-Martens-Latem, Belgium), using the Dice coefficient, and visualized as a dendrogram by the unweighted-pair group method, using average linkages with 1 % tolerance and 1 % optimization settings. In order to define a cluster, a cutoff of 80 % similarity was used.
SCCmec typing
SCCmec typing was performed using multiplex PCR as described by Milheirico et al. [24]. All assays were performed in a GeneAmp 9600 thermocycler (Applied Biosystems). The optimal cycling conditions were the following: 95 °C for 5 min; 35 cycles of 95 °C for 45 s, 57 °C for 45 s, and 72 °C for 1 min; and a final extension at 72 °C for 10 min. Each PCR mixture contained 0.5 µL of the primers listed in Table 1, KAPA2G Robust HotStart ReadyMix PCR (KAPA biosystems) which contains KAPA2G Robust HotStart DNA Polymerase (1 U/25 µL reaction) in a proprietary reaction buffer containing dNTPs (0.2 mM of each dNTP at 1X), MgCl2 (2 mM at 1X), 0.3 µL (3 mM) additional MgCl2, 10.7 µL of PCR grade water and genomic DNA in a final volume of 25 µL. The following S. aureus isolates were used as controls: BAA-38, BAA-1681, BAA-39, BAA-1680, BAA-1688 and BAA-42 for SCCmec types I–VI respectively. The PCR products were resolved in a 1 % SeaKem Gold Agarose (Lonza, Rockland, USA) gel in 0.5 % Tris–borate-ethylene-diamine-tetra-acetic acid (EDTA) buffer (Bio-Rad, Hercules, CA) at 4 V/cm for 2.5 h and were visualized with ethidium bromide.
spa typing
This was done using the following primers: 1095 F: 5′-AGACGATCCTTCGGTGAGC-3′ and 1517R: 5′-GCTTTTGCAATGTCATTTACTG-3′. PCR reactions consisted of 12.5 uL of KAPA2G Robust HotStart ReadyMix PCR (KAPA biosystems) which contains KAPA2G Robust HotStart DNA Polymerase (1 U per 25 µL reaction) in a proprietary reaction buffer containing dNTPs (0.2 mM of each dNTP at 1X), MgCl2 (2 mM at 1X), 0.5 μM of primers and genomic DNA in a final volume of 25 µL. PCR conditions were 95 °C for 6 min; 30 cycles each of 95 °C for 45 s, 64 °C for 45 s, and 72 °C for 60 s; and a final extension at 72 °C for 6 min. Sequencing was outsourced to inqaba biotec, a biotechnology company based in Pretoria, South Africa. Using the Ridom spa server (http://www.spa.server.ridom.de), spa sequences were automatically assigned to spa types. Sequence types and clonal complexes (spa-CC) were assigned where possible using Based Upon Repeat Patterns (BURP) grouping analysis from the Ridom StaphType software (version 1.4; Ridom GmbH, Würzburg, Germany). For BURP analysis, default parameters were used which allows spa types with maximum 4 genetic differences to be grouped into one cluster resulting in a calculated cost between members of a group being less than or equal to 4.
MLST
MLST was done on representative isolates from each PFGE clonal complex and selected singletons according to the protocol published by Enright et al. [25]. The PCRs were carried out as uniplex reactions consisting of 1 µM of the forward and reverse primers, 12.5 µL of 2× KAPA Taq ReadyMix (KAPA Biosystems), 2.5 mM MgCl2, 1 µL of template DNA and nuclease free water up to 25 µL. The PCR conditions were 95 °C for 5 min, followed by 30 cycles of 95 °C for 45 s, 56 °C for 45 s and 72 °C for 1 min. A final elongation step was carried out at 72 °C for 10 min. 5 µL of the PCR product was visualised with gel electrophoresis at 120 V for 1 h. Sequencing was performed on the remainder of the PCR product by Inqaba Biotechnical Industries (Pty) Ltd (Pretoria, South Africa). Sequences were inspected and trimmed in BioEdit Sequence Alignment Editor using reference sequences for each of the seven loci. A consensus sequence was generated from the forward and reverse sequences and used to generate sequence types (STs) on the S. aureus MLST database (http://www.saureus.beta.mlst.net/#). Isolates that were not typed by MLST were assigned STs using BURP analysis. Isolates with the same PFGE clonal complex and spa type were assigned the same STs. MLST clonal complexes (MLST-CC) were determined using a Java applet found at http://www.eburst.mlst.net that uses the eBURST algorithm. The default setting was used in which STs that share identical alleles at 6 or 7 of MLST loci are put in the same group. Where there was a discrepancy between the CC determined using eBURST and BURP, we considered the MLST-CC as the correct one.
Results
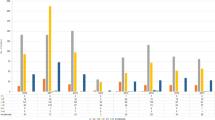
A total of 93 S. aureus isolates underwent PFGE. These were subsequently grouped into 19 PFGE clonal complexes (A–S) and 12 singletons. From these, 55 (32 MRSA and 23 MSSA) representative isolates from each PFGE clonal complex and all singletons were spa typed. This comprised 41 isolates from AKUHN and 14 from KEMRI. In total, there were 18 different MRSA spa types and 22 different spa types amongst the MSSA. The predominant MRSA spa type was t037 comprising 40.6 % (13/32) of all MRSA. In contrast, the MSSA were quite heterogeneous, only 2 out of 23 MSSA shared the same spa type. Two new MRSA spa types (t13149 and t13150) and 3 new MSSA spa types (t13182, t13193, t13194) were identified as shown in Table 2. Three spa types (t005, t318 and t1476) were found in both MSSA and MRSA. BURP analysis for both MSSA and MRSA revealed 7 spa-clonal complexes and 14 singletons as shown in Fig. 1. The predominant spa-CC was spa-CC005 which included the new MRSA spa type 13149. SCCmec type-III [3A] was the predominant type followed by SCCmec-IV [2B]. Only one MRSA isolate was non-typeable using the SCCmec protocol published by Milheirico et al. [24]

Based upon repeat pattern clustering analysis for all identified S. aureus isolates: the clustering analysis resulted in seven spa-clonal complexes and 14 singletons. The blue circle represents the group founder and the circle size is proportional to the frequency of the spa type
MLST STs were determined and extrapolated for 31 isolates. A total of seven different MRSA and MSSA MLST-CC were identified with the predominant one being MLST-CC 5. This clonal complex comprised STs 1, 5, 8 and 241 as shown in Table 2. An isolate belonging to ST241 (t2029) that was detected in a pus sample from AKUHN hospital was found to harbor SCCmec type IV [2B].
Out of the 16 MRSA from AKUHN, 13 were MDR including the two new spa types. Resistance was commonly seen to clindamycin, erythromycin and trimethoprim/sulfamethoxazole (TMP/SMX). A number of isolates had intermediate resistance to levofloxacin. However, two isolates were only resistant to beta lactams but susceptible to all other antibiotics including TMP/SMX as shown in Table 3. None of the MRSA was resistant to vancomycin, linezolid, mupirocin, teicoplanin or tigecycline.
Discussion
This study reveals a markedly heterogeneous population of S. aureus isolates as well as the presence of well described MRSA clonal complexes 5, 22 and 30 that are responsible for several outbreaks worldwide [13, 26]. CC5 has been identified as the major clonal complex causing HA-MRSA in Africa with MRSA ST239/ST241-III [3A] having been identified in several African countries [6]. The main clonal complex in our study was CC5 that included ST 241, a single locus variant of ST 239 also known as the “Brazilian/Hungarian clone”. ST 239 and ST 239 like isolates are well-known epidemic clones responsible for several healthcare associated MRSA outbreaks globally. They have been found to be a cause of hospital acquired infections in other African countries including Algeria, Ghana, Morocco, South Africa and Nigeria [6]. A study done by Aiken et al. [17] in a public hospital that is approximately 40 km from Nairobi identified t037-ST239 as the predominant clone carried by inpatients in a surgical ward. Most of the nasal swabs in our study were obtained from children in a paediatric ward situated in a public referral hospital. The high proportion of t037-ST241 among our MRSA isolates is not necessarily reflective of the true prevalence of this spa type in Nairobi due to a selection bias in the manner in which the isolates were collected. Nevertheless, it is quite concerning that a clone known to be associated with MRSA epidemics is resident within hospitals in Nairobi indicating an urgent need for proper infection control interventions and regular surveillance.
Unlike the study by Aiken et al. [17] that only found one MRSA clone, we identified 18 distinct spa types amongst the MRSA isolates belonging to very diverse sequence types, including 2 MRSA spa types (t13149 and t13150) that have not previously been described. The spa type t13150 was found to belong to ST5-II [2A] which has also been found in Nigeria and Senegal [27, 28]. We identified MRSA belonging to ST22 which in Africa has only previously been found in Algeria, Tunisia and South Africa. This clone has been widely associated with hospital epidemics especially in new born units [29]. The “West Australia MRSA-2 clone” (WA-MRSA-2), ST88-IV [2B] which has been reported in Cameroon and Madagascar was not found and the European MRSA clone ST80-IV that has been found in North African countries was not present in our collection. None of the MRSA in this study belonged to spa type t008, the prevalent spa type associated with the USA300 pulsotype that has been identified as the major cause of community acquired skin and soft tissue infections in North America [30, 31]. Although the isolates included in our study were few, they represent a fairly diverse collection from both a public and private referral hospital and we can therefore conclude that USA300 is not common in Nairobi.
The 23 MSSA belonged to 22 different spa types highlighting their marked genetic diversity in contrast to the MRSA. There were 3 spa types (t005, t318 and t1476) that were found in both MSSA and MRSA suggesting the possibility of local acquisition of an SCCmec element. One of the MSSA spa types belongs to t002 which is associated with the MRSA pulsotype USA100 [16]. The spa type t064 was also found which is associated with one of the major MRSA clones (ST612- SCCmec IV [2B] found in South Africa [32]. ST241 has frequently been associated with SCCmec III [3A], however, one isolate belonging to ST241 harbored SCCmec IV [2B] (this SCCmec element was more common in AKUHN hospital). This particular clone was previously observed in a large university clinic in Nigeria [33]. SCCmec types IV and V are small in size and can be transmitted both in the community and healthcare settings. Potentially, this could result in the emergence of well-known epidemic MRSA clones like the predominant European CA-MRSA clone ST80-IV [2B] whose for bearer is thought to be a PVL-positive MSSA from sub-Saharan Africa that acquired the SCCmec IV [2B] [34]. The MSSA strain t021-ST30 has also been associated with a known PVL positive CA-MRSA clone [35].
The multi-drug resistant patterns for the MRSA in this study are in keeping with what has been described in other countries in Africa [17, 32, 36, 37]. Most of the MRSA were resistant to macrolide–lincosamide, tetracycline and sulphonamide group of antibiotics which is fairly common amongst MRSA especially those that are healthcare associated. However two of the isolates showed resistance to only beta lactam antibiotics suggesting that they may be community acquired (based on their molecular structure) given that they belonged to SCCmec type IV which has been associated with CA-MRSA.
The major limitation of this study is that the isolates characterized were not collected in a structured and consistent manner and as such the proportions reported do not necessarily represent a true picture of the relative distributions of different clones in Nairobi due to a selection bias. The over representation of nasal swab specimens from a paediatric population from one hospital may have exaggerated the prevalence of t037-ST 241. We also did not carry out MLST and spa typing on all isolates due to financial constraints. However, we did ensure that a representative isolate from each PFGE clonal complex was included in the isolates that were further characterized using MLST and spa typing.
Conclusion
To the best of our knowledge, this is the largest study from Kenya that has carried out PFGE, MLST, spa and SCCmec typing on a diverse collection of MRSA isolates. This study highlights the marked genetic diversity of MSSA and MRSA isolates in Nairobi including the presence of well-known epidemic MRSA clones and new MRSA spa types. Given the evolution of S. aureus over the years, there is need for continuous surveillance in order to keep track of emerging clones. The existence of epidemic MRSA clones further justifies the need to strengthen infection control measures within our hospitals so as to avoid nosocomial S. aureus infections.
Abbreviations
- AKUHN:
-
Aga Khan University Hospital Nairobi
- BURP:
-
based upon repeat pattern
- CC:
-
clonal complex
- CLSI:
-
Clinical Laboratory Standards Institute
- DNA:
-
deoxyribonucleic acid
- dNTP:
-
deoxynucleotide triphosphate
- EDTA:
-
ethylene-diamine-tetraacetic acid
- KEMRI:
-
Kenya Medical Research Institute
- MDR:
-
multidrug resistance
- MLST:
-
multi-locus sequence Type
- MSSA:
-
methicillin susceptible Staphylococcus aureus
- MRSA:
-
methicillin resistant Staphylococcus aureus
- ND:
-
not defined
- NF:
-
no founder
- PBP2a:
-
penicillin binding protein 2a
- PCR:
-
polymerase chain reaction
- S. aureus :
-
Staphylococcus aureus
- SCCmec :
-
Staphylococcal cassette chromosome mec
- ST:
-
sequence type
- UT:
-
untypeable
References
Diekema DJ, Pfaller MA, Schmitz FJ, Smayevsky J, Bell J, Jones RN, Beach M. Survey of infections due to Staphylococcus species: frequency of occurrence and antimicrobial susceptibility of isolates collected in the United States, Canada, Latin America, Europe, and the Western Pacific region for the SENTRY Antimicrobial Surveillanc. Clin Infect Dis. 2001;32(Suppl 2):S114–32.
Chambers HF. The changing epidemiology of Staphylococcus aureus? Emerg Infect Dis. 2001;7:178–82.
Grundmann H, Aires-de-Sousa M, Boyce J, Tiemersma E. Emergence and resurgence of meticillin-resistant Staphylococcus aureus as a public-health threat. Lancet. 2006;368:874–85.
Seybold U, Kourbatova EV, Johnson JG, Halvosa SJ, Wang YF, King MD, Ray SM, Blumberg HM. Emergence of community-associated methicillin-resistant Staphylococcus aureus USA300 genotype as a major cause of health care-associated blood stream infections. Clin Infect Dis. 2006;42:647–56.
Chambers HF. Methicillin resistance in staphylococci: molecular and biochemical basis and clinical implications. Clin Microbiol Rev. 1997;10:781–91.
Abdulgader SM, Shittu AO, Nicol MP, Kaba M. Molecular epidemiology of methicillin-resistant Staphylococcus aureus in Africa: a systematic review. Front Microbiol. 2015;6:348.
Omuse G, Kabera B, Revathi G. Low prevalence of methicillin resistant Staphylococcus aureus as determined by an automated identification system in two private hospitals in Nairobi, Kenya: a cross sectional study. BMC Infect Dis. 2014;14:669.
Maina EK, Kiiyukia C, Wamae CN, Waiyaki PG, Kariuki S. Characterization of methicillin-resistant Staphylococcus aureus from skin and soft tissue infections in patients in Nairobi, Kenya. Int J Infect Dis. 2012;17:e115–9.
Zinn CS, Westh H, Rosdahl VT. An international multicenter study of antimicrobial resistance and typing of hospital Staphylococcus aureus isolates from 21 laboratories in 19 countries or states. Microb Drug Resist. 2004;10:160–8.
Cookson BD. Methicillin-resistant Staphylococcus aureus in the community: new battlefronts, or are the battles lost? Infect Control Hosp Epidemiol. 2000;21:398–403.
Falagas ME, Karageorgopoulos DE, Leptidis J, Korbila IP. MRSA in Africa: filling the global map of antimicrobial resistance. PLoS ONE. 2013;8:e68024.
Chatterjee SS, Otto M. Improved understanding of factors driving methicillin-resistant Staphylococcus aureus epidemic waves. Clin Epidemiol. 2013;5:205–17.
Stefani S, Chung DR, Lindsay JA, Friedrich AW, Kearns AM, Westh H, Mackenzie FM. Meticillin-resistant Staphylococcus aureus (MRSA): global epidemiology and harmonisation of typing methods. Int J Antimicrob Agents. 2012;39:273–82.
Enright MC, Robinson DA, Randle G, Feil EJ, Grundmann H, Spratt BG. The evolutionary history of methicillin-resistant Staphylococcus aureus (MRSA). Proc Natl Acad Sci USA. 2002;99:7687–92.
Bozdoğan B, Yıldız O, Oryaşın E, Kırdar S, Gülcü B, Aktepe O, Arslan U, Bayramoğlu G, Coban AY, Coşkuner SA, Güdücüoğlu H, Karabiber N, Oncü S, Tatman Otkun M, Ozkütük N, Ozyurt M, Sener AG. t030 is the most common spa type among methicillin-resistant Staphylococcus aureus strains isolated from Turkish hospitals. Mikrobiyol Bul. 2013;47:571–81.
David MZ, Taylor A, Lynfield R, Boxrud DJ, Short G, Zychowski D, Boyle-Vavra S, Daum RS. Comparing pulsed-field gel electrophoresis with multilocus sequence typing, spa typing, Staphylococcal Cassette Chromosome mec (SCC mec) typing, and PCR for Panton–Valentine Leukocidin, arcA, and opp3 in methicillin-resistant Staphylococcus aureus Isola. J Clin Microbiol. 2013;51:814–9.
Aiken AM, Mutuku IM, Sabat AJ, Akkerboom V, Mwangi J, Scott JAG, Morpeth SC, Friedrich AW, Grundmann H. Carriage of Staphylococcus aureus in Thika level 5 Hospital, Kenya: a cross-sectional study. Antimicrob Resist Infect Control. 2014;3:22.
Robinson DA, Enright MC. Evolution of Staphylococcus aureus by large chromosomal replacements. J Bacteriol. 2004;186:1060–4.
de Aires de Sousa M, Sanches IS, Ferro ML, Vaz MJ, Saraiva Z, Tendeiro T, Serra J, de Lencastre H. Intercontinental spread of a multidrug-resistant methicillin-resistant Staphylococcus aureus clone. J Clin Microbiol. 1998;36:2590–6.
Xu BL, Zhang G, Ye HF, Feil EJ, Chen GR, Zhou XM, Zhan XM, Chen SM, Pan WB. Predominance of the Hungarian clone (ST 239-III) among hospital-acquired meticillin-resistant Staphylococcus aureus isolates recovered throughout mainland China. J Hosp Infect. 2009;71:245–55.
Edgeworth JD, Yadegarfar G, Pathak S, Batra R, Cockfield JD, Wyncoll D, Beale R, Lindsay JA. An outbreak in an intensive care unit of a strain of methicillin-resistant Staphylococcus aureus sequence type 239 associated with an increased rate of vascular access device-related bacteremia. Clin Infect Dis. 2007;44:493–501.
Wayne P. Performance standards for antimicrobial susceptibility testing; twentieth informational supplement. M100-S20. Clin Lab Stand Inst. 2010:29.
CDC. Oxacillin—resistant Staphylococcus aureus on PulseNet (OPN): Laboratory Protocol for Molecular Typing of S. aureus by Pulsed—field gel electrophoresis (pfge) growing cultures : plug preparation . Natl Mol Subtyping Netw foodborne Dis Surveill. 2013:1–24.
Milheirico C, Oliveira DC, de Lencastre H. Update to the multiplex PCR strategy for assignment of mec element types in Staphylococcus aureus. Antimicrob Agents Chemother. 2007;51:4537.
Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol. 2000;38:1008–15.
Deurenberg RH, Stobberingh EE. The evolution of Staphylococcus aureus. Infect Genet Evol. 2008;8:747–63.
Breurec S, Fall C, Pouillot R, Boisier P, Brisse S, Diene-Sarr F, Djibo S, Etienne J, Fonkoua MC, Perrier-Gros-Claude JD, Ramarokoto CE, Randrianirina F, Thiberge JM, Zriouil SB, Garin B, Laurent F. Epidemiology of methicillin-susceptible Staphylococcus aureus lineages in five major African towns: high prevalence of Panton–Valentine leukocidin genes. Clin Microbiol Infect. 2011;17:633–9.
Raji A, Ojemhen O, Umejiburu U, Ogunleye A, Blanc DS, Basset P. High genetic diversity of Staphylococcus aureus in a tertiary care hospital in Southwest Nigeria. Diagn Microbiol Infect Dis. 2013;77:367–9.
Pinto AN, Seth R, Zhou F, Tallon J, Dempsey K, Tracy M, Gilbert GL, O’Sullivan MVN. Emergence and control of an outbreak of infections due to Panton–Valentine leukocidin positive, ST22 methicillin-resistant Staphylococcus aureus in a neonatal intensive care unit. Clin Microbiol Infect. 2013;19:620–7.
Al-Rawahi GN, Reynolds S, Porter SD, Forrester L, Kishi L, Chong T, Bowie WR, Doyle PW. Community-associated CMRSA-10 (USA-300) is the predominant strain among methicillin-resistant Staphylococcus aureus strains causing skin and soft tissue infections in patients presenting to the emergency department of a Canadian tertiary care hospital. J Emerg Med. 2010;38:6–11.
Ray GT, Suaya JA, Baxter R. Incidence, microbiology, and patient characteristics of skin and soft-tissue infections in a U.S. population: a retrospective population-based study. BMC Infect Dis. 2013;13:252.
Shittu A, Nübel U, Udo E, Lin J, Gaogakwe S. Characterization of meticillin-resistant Staphylococcus aureus isolates from hospitals in KwaZulu-Natal province, Republic of South Africa. J Med Microbiol. 2009;58(Pt 9):1219–26.
Ghebremedhin B, Olugbosi MO, Raji AM, Layer F, Bakare RA, Konig B, Konig W. Emergence of a community-associated methicillin-resistant Staphylococcus aureus strain with a unique resistance profile in Southwest Nigeria. J Clin Microbiol. 2009;47:2975–80.
Stegger M, Wirth T, Andersen PS, Skov RL, De Grassi A, Simões PM, Tristan A, Petersen A, Aziz M, Kiil K, Cirković I, Udo EE, del Campo R, Vuopio-Varkila J, Ahmad N, Tokajian S, Peters G, Schaumburg F, Olsson-Liljequist B, Givskov M, Driebe EE, Vigh HE, Shittu A, Ramdani-Bougessa N, Rasigade J-P, Price LB, Vandenesch F, Larsen AR, Laurent F. Origin and evolution of European community-acquired methicillin-resistant Staphylococcus aureus. MBio. 2014;5:e01044.
Monecke S, Coombs G, Shore AC, Coleman DC, Akpaka P, Borg M, Chow H, Ip M, Jatzwauk L, Jonas D, Kadlec K, Kearns A, Laurent F, O’Brien FG, Pearson J, Ruppelt A, Schwarz S, Scicluna E, Slickers P, Tan HL, Weber S, Ehricht R. A field guide to pandemic, epidemic and sporadic clones of methicillin-resistant Staphylococcus aureus. PLoS One. 2011;6:e17936.
Egyir B, Guardabassi L, Sørum M, Nielsen SS, Kolekang A, Frimpong E, Addo KK, Newman MJ, Larsen AR. Molecular Epidemiology and antimicrobial susceptibility of clinical Staphylococcus aureus from Healthcare Institutions in Ghana. PLoS ONE. 2014;9:e89716.
Marais E, Aithma N, Perovic O, Oosthuysen WF, Musenge E, Dusé AG. Antimicrobial susceptibility of methicillin-resistant Staphylococcus aureus isolates from South Africa. S Afr Med J. 2009;99:170–3.
Authors’ contributions
GO conceptualized the study, collected isolates, carried out molecular analysis and drafted the manuscript. KNVZ and KH assisted in molecular analysis of samples and drafting of the manuscript. SA helped in carrying out BURP and eBURST analysis, drafting and critiquing the manuscript. SK, AW and GV assisted in designing the study, getting samples, interpreting of results and drafting the manuscript. All authors read and approved the final manuscript.
Acknowledgements
We acknowledge the use of the S. aureus MLST database which is located at Imperial College London and is funded by the Wellcome Trust.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The datasets analysed during the current study are available from the corresponding author on reasonable request.
Ethics approval and consent to participate
This being a low risk study, a waiver from full ethics review was granted by the AKUHN research ethics committee (2016/REC-33).
Funding
All the molecular work was done in the Microbiology section, Department of Pathology, Tygerberg Hospital, Cape Town, South Africa. This was facilitated through a technology transfer grant awarded to Prof Andrew Whitelaw by Stellenbosch University.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Omuse, G., Van Zyl, K.N., Hoek, K. et al. Molecular characterization of Staphylococcus aureus isolates from various healthcare institutions in Nairobi, Kenya: a cross sectional study. Ann Clin Microbiol Antimicrob 15, 51 (2016). https://doi.org/10.1186/s12941-016-0171-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12941-016-0171-z