
Abstract
Background
Resistance to anti-malarial drugs is associated with polymorphisms in target genes and surveillance for these molecular markers is important to detect the emergence of mutations associated with drug resistance and signal recovering sensitivity to anti-malarials previously used.
Methods
The presence of polymorphisms in genes associated with Plasmodium falciparum resistance to chloroquine and sulfadoxine-pyrimethamine was evaluated by Sanger sequencing, in 85 P. falciparum day of enrollment samples from a therapeutic efficacy study of artemether–lumefantrine conducted in 2018–2019 in Quibdo, Colombia. Samples were genotyped to assess mutations in pfcrt (codons 72–76), pfdhfr (codons 51, 59, 108, and 164), and pfdhps genes (codons 436, 437, 540, and 581). Further, the genetic diversity of infections using seven neutral microsatellites (NMSs) (C2M34, C3M69, Poly α, TA1, TA109, 2490, and PfPK2) was assessed.
Results
All isolates carried mutant alleles for pfcrt (K76T and N75E), and for pfdhfr (N51I and S108N), while for pfdhps, mutations were observed only for codon A437G (32/73, 43.8%). Fifty samples (58.8%) showed a complete neutral microsatellites (NMS) profile. The low mean number of alleles (2 ± 0.57) per locus and mean expected heterozygosity (0.17 ± 0.03) showed a reduced genetic diversity. NMS multilocus genotypes (MMG) were built and nine MMG were identified.
Conclusions
Overall, these findings confirm the fixation of chloroquine and pyrimethamine-resistant alleles already described in the literature, implying that these drugs are not currently appropriate for use in Colombia. In contrast, mutations in the pfdhps gene were only observed at codon 437, an indication that full resistance to sulfadoxine has not been achieved in Choco. MMGs found matched the clonal lineage E variant 1 previously reported in northwestern Colombia.
Similar content being viewed by others

Background
Malaria transmission in the Americas is characterized as unstable, heterogeneous, and predominantly of low intensity [1]. In addition, a downward trend of malaria cases was observed between 2000 and 2014, which changed in 2015, when a significant resurgence of epidemic transmission in Venezuela led to increase in number of cases along with increased transmission in endemic areas of Brazil, Colombia, Guyana, Nicaragua, and Panama; as well as outbreaks in countries that were moving towards elimination [2]. In 2019, more than 86% of malaria cases in the Americas, were concentrated in Venezuela, Brazil, and Colombia [3].
Colombia is endemic for Plasmodium falciparum and Plasmodium vivax and approximately 70,000 malaria cases are reported annually by the public health services, mainly in departments of Choco, and Nariño [4]. Despite the relatively large number of malaria cases reported each year and the use of different anti-malarial treatments in recent decades, little is known about the molecular markers of drug resistance and genetic diversity of P. falciparum in the country. Since 2010, the first-line regimen for treating uncomplicated P. falciparum malaria in Colombia has been artemether–lumefantrine (AL). Prior to the use of this artemisinin-based combination therapy (ACT), chloroquine (CQ), sulfadoxine-pyrimethamine (SP), and amodiaquine (AQ) were widely used, until high rates of therapeutic failure were reached [5,6,7]. CQ monotherapy was employed until the early 1980s, but when resistance to this anti-malarial emerged, SP in combination with CQ or AQ became the therapeutic regimens utilized. In 1999, AQ-SP was implemented as the first-line treatment for uncomplicated P. falciparum malaria and from 2006, two artemisinin-based combinations, artesunate-mefloquine and AL, were introduced.
Plasmodium falciparum resistance to CQ and SP has been associated with point mutations in the chloroquine-resistance transporter (pfcrt gene) [8], and the dihydrofolate reductase (pfdhfr) and dihydropteroate synthase (pfdhps) genes [9], respectively. The pfcrt point mutation K76T is critical to confering CQ resistance, but mutations in other codons are also associated with resistance [8]. Resistance to sulfadoxine-pyrimethamine is associated with the accumulation of mutations at codons 50, 51, 59, 108, and 164 in the pfdhfr gene (codon 50 being more relevant in South America) and 436, 437, 540, and 581 of the pfdhps gene, respectively [9].
In 2018, a therapeutic efficacy study (TES) was carried out to evaluate the efficacy of AL for the treatment of uncomplicated P. falciparum malaria in Quibdo, Choco, together with the evaluation of the confirmed molecular marker of resistance to artemisinin in the pfk13 gene, and to unconfirmed marker for lumefantrine resistance, pfmdr1 gene [10]. Although TESs are indispensable to assess the efficacy of anti-malarial drugs, molecular surveillance is helpful to detect the emergence of mutations associated with drug resistance and signal recovering sensitivity to an anti-malarial used in the past. To evaluate if the prevalence of wild-type genotypes of pfcrt, pfdhfr, and pfdhps genes has changed over time since their withdrawal in Colombia, molecular testing of samples collected during the 2018 TES was conducted. In addition, the genetic diversity of infections using seven neutral microsatellites (NMSs) loci was assessed.
Methods
Ethics statement
Samples for this study were obtained from a TES conducted in Quibdo in 2018 [10]; as part of the TES, written informed consent for secondary use of remnant samples were obtained. The TES protocol was reviewed and approved by the Institutional Ethics Committee of the Colombian National Institute of Health (Protocol CEMIN 2-2018, Minute #5 of March 22, 2018) and the PAHO Ethics Review Committee (PAHOERC) (PAHO 2018-04-0029). CDC considered the study a public health evaluation (reference number: 2018-063).
Study site, sample collection, DNA extraction, and PET-PCR
Quibdo is the capital of Choco department, located in northwestern Colombia (lat. 5° 41′ 32″ N, long. 76° 39′ 29″ W) on the banks of the Atrato river in the Pacific region, at an altitude of 43 m above sea level, the average temperature is 28 °C and annual humidity oscillates between 86 and 88%. The Pacific region is separated from Amazon and Orinoco region by the Andes Mountain range. Quibdo has a population of 116,087, mainly consisting of Afro Colombians (87.5%), of which 65% is settled in the urban area. Quibdo contributes 6 to 11% of the total cases of malaria reported in Colombia.
Samples collected at patient enrollment in the TES were used in this study. Parasite DNA was extracted from dried blood spots collected on filter paper using the QIAamp® DNA Micro Kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. The Plasmodium species was confirmed by a real-time PCR, photo-induced electron transfer polymerase chain reaction (PET-PCR) as previously described [11].
Sanger sequencing for molecular markers of resistance
Plasmodium falciparum samples were genotyped using PCR or nested PCR followed by Sanger sequencing to evaluate mutations at pfcrt gene (codons 72–76), pfdhfr gene (codons 51, 59, 108, and 164), and pfdhps gene (codons 436, 437, 540, and 581). Primer sequences and PCR cycling conditions for each gene are reported elsewhere [12,13,14]. Briefly, primary PCR was performed with 1 μL genomic DNA, 7.5 μL of 2X master mix (Promega, Madison, WI, USA), and 0.27 μM each primer in 15 μL final volume for pfdhfr and dhps genes. Conditions for pfcrt gene were the same, except 1.2 μL of DNA and a final concentration of 0.33 μM each primer. Nested PCR was done for pfdhps and pfcrt genes. PCR products were purified with ExoSAP-it reagent (New England Biolabs, Ipswich, MA, USA) and sequenced using the Sanger method (Genewiz INC, NJ, USA). Laboratory P. falciparum strains, 3D7 and 7G8, were used as wild-type and mutant-type controls, respectively. Sequence analysis was performed using Geneious® 7.1.7 software (Biomatters, Auckland, New Zealand) using the P. falciparum 3D7 reference strain for pfcrt (PF3D7_0709000), pfdhfr (PF3D7_0417200), and pfdhps (PF3D7_0810800) genes. Mutant alleles were identified, and the frequency of each allele was determined.
Neutral microsatellites
All samples were genotyped using a set of seven NMSs to identify polyclonal infections. The microsatellite loci amplified were on chromosome (chr) 2 (C2M34), chr 3 (C3M69), chr 4 (Polyα), chr 6 (TA1 and TA109), chr 10 (2490), and chr 12 (PfPK2) [15,16,17]. Amplification products labeled fluorescently with FAM or HEX of the 7 loci were followed by fragment electrophoresis on a capillary sequencer ABI 3130 xl Genetic Analyzer (Applied Biosystems, Foster city, CA, USA) and then fragment sizes were scored using GeneMapper®V2.7.0 software (Applied Biosystems, Foster city, CA, USA). PCR cycling conditions for each locus [15,16,17] are registered elsewhere. Samples that carried a single allele at each locus were considered monoclonal infections and, when more than one allele was found at any ≥ 1 locus, they were considered polyclonal infections. Presence of alleles (multiple peaks) was evaluated by locus and minor alleles were scored if they were more than 33% the height of the peaks corresponding to the predominant alleles. Genetic variation at each locus was quantified by the allelic diversity, which was calculated in terms of expected heterozygosity (He) using the formula \({H}_{e}=\left[\frac{n}{n-1}\right]\left[1-\sum {p}_{i}^{2}\right]\), where n is the number of P. falciparum isolates genotyped for a particular locus and pi is the frequency of the ith allele. The sampling variance of He was estimated as \(\left[\frac{2\left(n-1\right)}{{n}^{3}}\right]\left(2\left(n-2\right)\left[\sum {p}_{i}^{3}-{\left(\sum {p}_{i}^{2}\right)}^{2}\right]\right)\) [18]. Infections containing more than one allele at any of the seven NMSs were not included in the analysis.
Results
Molecular markers of resistance
A total of 88 samples were processed of which 85 could be confirmed as P. falciparum by PET-PCR. Among these 85 samples, sequencing of the pfcrt, pfdhfr, and pfdhps genes was successful in 72 (84.7%), 79 (92.9%), and 73 (85.9%) samples, respectively. For the pfcrt gene, all isolates carried the K76T and N75E mutant alleles. Only two mutant alleles, N51I and S108N, were observed in the pfdhfr gene, and for the pfdhps gene wild-type alleles prevailed in all codons investigated except in codon 437, where 32 (43.8%) isolates carried the mutant allele A437G (Table 1).
Genetic diversity
A complete NMS profile was obtained for 58.8% (50/85) of the samples, 22.4% (19/85) lacked allele amplification in one locus and 18.8% (16/85) in ≥ 2 loci. Monoclonal infections were predominant (96%) and only 2 samples showed evidence of polyclonal infections. Fifteen alleles were found and of these, one had not been previously reported in Colombia [19,20,21]; allele size 193 bp in TA109 was found in a polyclonal infection and was different from those previously reported in Choco. Mean number of alleles per locus (2.0 ± 0.57) and mean He (0.17 ± 0.03) were low (Table 2).
NMS multilocus genotypes (MMGs) were built from the samples with monoclonal infection by combining the results of the seven microsatellites and coded with numbers. Nine MMGs were identified from samples with the complete profile (Table 3) revealing the existence of related multilocus genotypes. MMG 1 and MMG 2 were the most prevalent genotypes and between each MMG there were minor variations in NMS composition, for example, MMG 1 and MMG 2 shared a similar genetic profile differing only in one allele size found in C2M34 (226 vs. 236 bp). The remaining seven MMGs were found in one or three samples and were related to MMG 1, either by sharing some of the alleles or by their close allele sizes. The allele sizes detected in 37 (77.1%) isolates (MMGs 1, 2, and 3) matched the clonal lineage E variant 1 (Ev1) previously reported [21], which exhibits the following profile TA1: 172, Polyα: 148, PfPK2: 175, TA109: 160, 2490: 74/75, C2M34: 236/226, C3M69: 124. Ev1 is a variant of the clonal lineage E, which was described earlier in Peru [22] and Ecuador [23]. Eleven (22.9%) isolates had different alleles from those described for Ev1 such as 154 in Poly-α, 160 and 163 in PfPK2, and 80 in 2490. This finding suggests that in Quibdo another variant of the clonal line E circulates, which was named variant 2 (Ev2). This variant was found in 8 isolates (MMGs 5, 6, 7, and 8) and its microsatellite profile was TA1: 172, Polyα: 148, PfPK2: 160/163, TA109: 160, 2490: 74/80, C2M34: 226, and C3M69: 124/140 (Fig. 1).

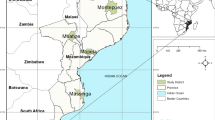
Variants of P. falciparum genetic lineage E found in Quibdo, Choco, 2018. Map of Colombia showing P. falciparum genetic lineages E (Ev1 and Ev2) found in Quibdo, Choco and genetic lineages found in other departments of Colombia and in neighbouring countries. Turbo in Antioquia (20), Antioquia (21), Nariño (21), Amazonas (21), Perú (22), and Ecuador (23)
No association was observed between the two resistance genotypes (combination of pfcrt, pfdhfr, and pfdhps resistance alleles) and MMGs found in this study; however, parasites with MMGs 2, 3, and 8 carried the CVMET/ICNI/AKA genotype and parasites with MMGs 4, 7, and 9 harbored CVMET/ICNI/GKA. Parasites with MMGs 1 and 6 showed both genotypes (see Additional file 1). When comparing the isolates belonging to Ev1 with the rest of the isolates found in Quibdo, samples shared the same drug resistance genotypes CVMET/ICNI/AKA or CVMET/ICNI/GKA, being more common the genotype CVMET/ICNI/AKA within the variant Ev1 (68,8%). Considering that the samples analysed in a previous report [10] are the same as in this study, Pfmdr1 genotypes NFSDD/NFSDY previously reported [10] were found in both the Ev1 and the remaining isolates; however, genotype NFSDD was detected mostly in Ev1 (25%) and only in isolates with an allele size of 226 bp in C2M34 (see Additional file 1).
Discussion
The monitoring of molecular markers of resistance is useful for detecting changes in the parasite genome associated with anti-malarial drugs resistance over time. In this study, only one CQ-resistant genotype (CVMET) was found, in contrast to the three genotypes (CVMET/CVMNT/CVIET) previously reported in samples from the Colombian Pacific [24]. Field isolates carried mutant K76T in pfcrt gene, indicating that, despite the withdrawal of CQ from national treatment guidelines for P. falciparum in 1998, resurgence and re-expansion of wild-type pfcrt P. falciparum has not occurred. A similar situation was reported in a previous study carried out in three Colombian malaria-endemic areas [25] and other countries in the Americas [26, 27]. This finding contrasts with what was observed in some countries of Africa, like Malawi [28, 29], where the recovery of CQ-sensitive wild-type pfcrt P. falciparum occurred in as little as 10 years after the use of CQ was discontinued.
As a result of CQ resistance in Colombia, SP was adopted in 1981; high and moderate levels of therapeutic failure were quickly observed in the Amazon basin [7] and northwestern regions [6], respectively. The analysis of molecular markers of resistance to SP in this study shows a predominance of double mutant (ICNI) for pfdhfr, while mutations in the pfdhps gene were only observed at codon 437 (43.8%), an indication that full resistance to sulfadoxine was not achieved in circulating parasites in Choco. In addition, ACT replaced the combined therapy SP with AQ for uncomplicated P. falciparum malaria in 2008, and since then there has been no selective pressure with SP. Similar resistance profiles were observed in northwest Colombia in previous studies [19, 30, 31]. This likely slow emergence of mutations in pfdhps in parasites from Quibdo could be explained, in part, by the use of multiple drug regimens (SP in combination with CQ or AQ plus primaquine as a gametocytocidal agent) unlike Brazil and the Peruvian Amazon region, where SP monotherapy was used and parasites with double and triple mutations predominate.
The frequency of mutations associated with resistance to CQ (K76T 100%) [25, 32, 33] and SP (N51I/S108N 93–100% and A437G 65–70%) [19, 30, 31] has remained constant in northwest Colombia for at least 10 years since introduction of ACT, suggesting the fixation of resistant alleles [12, 34] in Choco. This situation could be explained by the strong selective pressure exerted by the drugs that fixed the mutant allele in this population, considering that the frequency of these mutant alleles is 100% and it has prevailed unchanged over the last decade in this part of the country. Other possible reasons for fixation of these mutant alleles include CQ and SP drug pressure not allowing for expansion of sensitive strains. Although CQ and SP are no longer used to treat P. falciparum malaria in Colombia, CQ constitutes the first-line regimen for vivax malaria and self-medication with CQ for non-laboratory confirm malaria has also been observed in 32% of symptomatic individuals [35]. In addition, the prevalence of CVMET over time may be because this haplotype, like SVMNT found in Papua New Guinea and Brazil, has no fitness disadvantage over the sensitive one even in the absence of drug pressure [36]. Finally, it is likely that no wild-type populations, able to replace the resistant populations, were left in Colombia after the development of resistance.
When comparing the number of alleles per microsatellite locus against collected samples in Quibdo during 2001–2007 [19] and in Turbo during 2002–2008 [20], a town located in Antioquia department in northwestern Colombia, more alleles were detected at that time (six and seven for Polyα and six and nine for PfPK2). Higher diversity values (expected heterozygosities) were found in both studies, 0.915 for the first, and 0.35 to 0.18 for the second, compared to 0.17 in the present study. These results points to a progressive reduction in the level of genetic diversity in P. falciparum from Quibdo, as well as a decrease in the frequency of multiple infections. These findings suggest the circulating P. falciparum population have a more clonal composition because these populations have undergone bottlenecks possibly caused by actions of national malaria control programme or interventions focused on improving access to timely diagnosis and adequate treatment, and the coverage in the use of insecticide-treated bed nets such as World Fund Malaria Project implemented in Colombia between 2010 and 2015. Additionally, alleles 172 bp in TA1 and 160 bp in TA109 seem to be fixed in parasites from Quibdo.
Of the 48 isolates analysed for NMS, 77.1% shared the same microsatellite profile of the clonal lineage EV1, which was described in the neighbouring department of Antioquia [21], and 16.7% showed a new variant of the clonal lineage E, named Ev2 (Fig. 1). This variant is characterized by having a genetically identical set of markers, but variable at others, with common ancestors occurring more than 15 years ago, according to allele sizes reported in a study carried out in Quibdo with samples collected in 2001 and 2006 [19].
When comparing the haplotypes found in Quibdo with other haplotypes reported in Colombia, it is evident that the clonal lineage E variant 1 predominates in this town and it is characteristic in this area of the country; this same variant was reported in samples from Antioquia collected in 2012, indicating that in the northwestern zone of Colombia there is a predominance of variants derived from genetic lineage E. The genetic lineage Bv1 predominates in the Amazonas department 21, variant previously observed in Peru 22, and the genetic lineage F 21 in Nariño department (southwestern Colombia) (Fig. 1). The foregoing allows us to suggest that specific genetic lineages predominate in the different endemic areas of malaria in Colombia.
Conclusions
Overall, the frequency of anti-malarial drug resistance genotypes and NMS multilocus genotypes was studied in northwestern Colombia to explore possible genomic adaptations in the parasite in response to changes in the anti-malarial policy and genetically characterize P. falciparum populations. Monitoring of molecular markers of anti-malarial drugs resistance should be considered as part of regular in vivo efficacy trials in malaria-endemic countries to timely detect emergence of polymorphisms conferring decreased susceptibility to anti-malarials, especially since self-medication, the use of subtherapeutic doses, gold mining activities associated with malaria outbreaks, and human mobility and migration are common in Colombia.
Availability of data and materials
Database is available in supplementary information.
Abbreviations
- ACT:
-
Artemisinin-based combination therapy
- AL:
-
Artemether–lumefantrine
- AQ:
-
Amodiaquine
- CQ:
-
Chloroquine
- DNA:
-
Deoxyribonucleic acid
- He:
-
Heterozygosity
- MMG:
-
NMS multilocus genotypes
- NMS:
-
Neutral microsatellites
- PCR:
-
Polymerase chain reaction
- PET-PCR:
-
Photo-induced electron transfer-PCR
- SP:
-
Sulfadoxine-pyrimethamine
- TES:
-
Therapeutic efficacy study
References
Padilla-Rodríguez JC, Olivera MJ, Ahumada-Franco ML, Paredes-Medina AE. Malaria risk stratification in Colombia 2010 to 2019. PLoS ONE. 2021;16: e0247811.
WHO. World malaria report 2019. Geneva, World Health Organization, 2019. https://www.who.int/publications/i/item/world-malaria-report-2019. Accessed 3 Nov 2021.
WHO. World malaria report 2020. Geneva, World Health Organization, 2020. https://www.who.int/publications/i/item/9789240015791. Accessed 3 Nov 2021.
Ferro C, Sastoque LA. Informe del evento malaria, Colombia 2019. Instituto Nacional de Salud. http://www.ins.gov.co/buscador-eventos/Informesdeevento/MALARIA%20PE%20XIII%202019.pdf. Accessed 9 Nov 2021.
Osorio LE, Giraldo LE, Grajales LF, Arriaga AL, Andrade AL, Ruebush TK, et al. Assessment of therapeutic response of Plasmodium falciparum to chloroquine and sulfadoxine-pyrimethamine in an area of low malaria transmission in Colombia. Am J Trop Med Hyg. 1999;61:968–72.
Blair S, Carmona-Fonseca J, Piñeros JG, Ríos A, Alvarez T, Alvarez G, et al. Therapeutic efficacy test in malaria falciparum in Antioquia, Colombia. Malar J. 2006;5:14.
Osorio L, Perez LDP, Gonzalez IJ. Assessment of the efficacy of antimalarial drugs in Tarapaca, in the Colombian Amazon basin. Biomedica. 2007;27:133–40.
Bray PG, Martin RE, Tilley L, Ward SA, Kirk K, Fidock DA. Defining the role of PfCRT in Plasmodium falciparum chloroquine resistance. Mol Microbiol. 2005;56:323–33.
Gregson A, Plowe CV. Mechanisms of resistance of malaria parasites to antifolates. Pharmacol Rev. 2005;57:117–45.
Olivera MJ, Guerra AP, Cortes LJ, Horth RZ, Padilla J, Novoa J, et al. Artemether–lumefantrine efficacy for the treatment of uncomplicated Plasmodium falciparum malaria in Choco, Colombia after 8 years as first-line treatment. Am J Trop Med Hyg. 2020;102:1056–63.
Lucchi NW, Narayanan J, Karell MA, Xayavong M, Kariuki S, DaSilva AJ, et al. Molecular diagnosis of malaria by photo-induced electron transfer fluorogenic primers: PET-PCR. PLoS ONE. 2013;8: e56677.
Griffing S, Syphard L, Sridaran S, McCollum AM, Mixson-Hayden T, Vinayak S, et al. pfmdr1 amplification and fixation of pfcrt chloroquine resistance alleles in Plasmodium falciparum in Venezuela. Antimicrob Agents Chemother. 2010;54:1572–9.
McCollum AM, Poe AC, Hamel M, Huber C, Zhou Z, Shi YP, et al. Antifolate resistance in Plasmodium falciparum: multiple origins and identification of novel dhfr alleles. J Infect Dis. 2006;194:189–97.
Vinayak S, Alam MT, Mixson-Hayden T, McCollum AM, Sem R, Shah NK, et al. Origin and evolution of sulfadoxine resistant Plasmodium falciparum. PLoS Pathog. 2010;6: e1000830.
Anderson TJ, Su XZ, Bockarie M, Lagog M, Day KP. Twelve microsatellite markers for characterization of Plasmodium falciparum from finger-prick blood samples. Parasitology. 1999;119:113–25.
Nair S, Williams JT, Brockman A, Paiphun L, Mayxay M, Newton PN, et al. A selective sweep driven by pyrimethamine treatment in Southeast Asian malaria parasites. Mol Biol Evol. 2003;20:1526–36.
Roper C, Pearce R, Bredenkamp B, Gumede J, Drakeley C, Mosha F, et al. Antifolate antimalarial resistance in southeast Africa: a population-based analysis. Lancet. 2003;361:1174–81.
Nash D, Nair S, Mayxay M, Newton PN, Guthmann JP, Nosten F, et al. Selection strength and hitchhiking around two anti-malarial resistance genes. Proc Biol Sci. 2005;272:1153–61.
Corredor V, Murillo C, Echeverry DF, Benavides J, Pearce RJ, Roper C, et al. Origin and dissemination across the Colombian Andes Mountain range of sulfadoxine-pyrimethamine resistance in Plasmodium falciparum. Antimicrob Agents Chemother. 2010;54:3121–5.
Chenet SM, Taylor JE, Blair S, Zuluaga L, Escalante AA. Longitudinal analysis of Plasmodium falciparum genetic variation in Turbo, Colombia: implications for malaria control and elimination. Malar J. 2015;14:363.
Dorado EJ, Okoth SA, Montenegro LM, Diaz G, Barnwell JW, Udhayakumar V, et al. Genetic characterisation of Plasmodium falciparum isolates with deletion of the pfhrp2 and/or pfhrp3 genes in Colombia: the Amazon region, a challenge for malaria diagnosis and control. PLoS ONE. 2016;11: e0163137.
Griffing SM, Mixson-Hayden T, Sridaran S, Alam MT, McCollum AM, Cabezas C, et al. South American Plasmodium falciparum after the malaria eradication era: clonal population expansion and survival of the fittest hybrids. PLoS ONE. 2011;6: e23486.
Sáenz FE, Morton LC, Okoth SA, Valenzuela G, Vera-Arias CA, Vélez-Álvarez E, et al. Clonal population expansion in an outbreak of Plasmodium falciparum on the northwest coast of Ecuador. Malar J. 2015;14:497.
Echeverry DF, Holmgren G, Murillo C, Higuita JC, Björkman A, Gil JP, et al. Polymorphisms in the pfcrt and pfmdr1 genes of Plasmodium falciparum and in vitro susceptibility to amodiaquine and desethylamodiaquine. Am J Trop Med Hyg. 2007;77:1034–8.
Aponte S, Guerra AP, Álvarez-Larrotta C, Bernal SD, Restrepo C, González C, et al. Baseline in vivo, ex vivo and molecular responses of Plasmodium falciparum to artemether and lumefantrine in three endemic zones for malaria in Colombia. Trans R Soc Trop Med Hyg. 2017;111:71–80.
Arróspide N, Hijar-Guerra G, de Mora D, Diaz-Cortéz CE, Veloz-Pérez R, Gutiérrez S, et al. Mutant alleles associated to chloroquine and sulfadoxine-pyrimethanime resistance in Plasmodium falciparum of the Ecuador-Peru and Ecuador-Colombia borders. Rev Peru Med Exp Salud Publica. 2014;31:282–7.
Valenzuela G, Castro LE, Valencia-Zamora J, Vera-Arias CA, Rohrbach P, Sáenz FE. Genotypes and phenotypes of resistance in Ecuadorian Plasmodium falciparum. Malar J. 2019;18:415.
Mita T, Kaneko A, Lum JK, Bwijo B, Takechi M, Zungu IL, et al. Recovery of chloroquine sensitivity and low prevalence of the Plasmodium falciparum chloroquine resistance transporter gene mutation K76T following the discontinuance of chloroquine use in Malawi. Am J Trop Med Hyg. 2003;68:413–5.
Laufer MK, Takala-Harrison S, Dzinjalamala FK, Stine OC, Taylor TE, Plowe CV. Return of chloroquine-susceptible falciparum malaria in Malawi was a reexpansion of diverse susceptible parasites. J Infect Dis. 2010;202:801–8.
Galindo JA, Knudson A, Nicholls RS, Guerra AP. Point mutations in dihydrofolate reductase and dihydropteroate synthase genes of Plasmodium falciparum from three endemic malaria regions in Colombia. Biomedica. 2010;30:56–64.
Hernández DC, Guerra AP, Cucunubá ZM, Nicholls RS, Barrera SM. Haplotypes associated with resistance to sulfadoxine-pyrimethamine in Plasmodium falciparum in two malaria endemic locations in Colombia. Infect Genet Evol. 2013;18:183–90.
Restrepo-Pineda E, Arango E, Maestre A, Do Rosário VE, Cravo P. Studies on antimalarial drug susceptibility in Colombia, in relation to Pfmdr1 and Pfcrt. Parasitology. 2008;135:547–53.
Restrepo E, Carmona-Fonseca J, Maestre A. Plasmodium falciparum: high frequency of pfcrt point mutations and emergence of new mutant haplotypes in Colombia. Biomedica. 2008;28:523–30.
Pacheco MA, Forero-Peña DA, Schneider KA, Chavero M, Gamardo A, Figuera L, et al. Malaria in Venezuela: changes in the complexity of infection reflects the increment in transmission intensity. Malar J. 2020;19:176.
Diaz G, Lasso AM, Murillo C, Montenegro LM, Echeverry DF. Evidence of self-medication with chloroquine before consultation for malaria in the Southern Pacific Coast Region of Colombia. Am J Trop Med Hyg. 2019;100:66–71.
Mehlotra RK, Fujioka H, Roepe PD, Janneh O, Ursos LM, Jacobs-Lorena V, et al. Evolution of a unique Plasmodium falciparum chloroquine-resistance phenotype in association with pfcrt polymorphism in Papua New Guinea and South America. Proc Natl Acad Sci USA. 2001;98:12689–94.
Acknowledgements
We would like to thank the managers of the malaria control programme in Choco, and staff and participants for their collaboration in this study.
Funding
Funding to carry out the molecular study was partially provided by the U.S. Agency for International Development (USAID) through the Latin American and the Caribbean Regional Malaria Programme, and by the Colombian National Institute of Health. The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
APG performed molecular assays, conceived and wrote the first draft of the article. APG and NWL analysed and interpreted the microsatellites and molecular markers data. . SMCh, AMO and NWL made important contributions on the initial draft and assisted in development of the final manuscript. SMCh, AMO, MJO, NWL, LJC contributed to the analysis, content, and writing of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The TES protocol was reviewed and approved by the Institutional Ethics Committee of the Colombian National Institute of Health (Protocol CEMIN 2-2018, Minute #5 of March 22, 2018) and the PAHO Ethics Review Committee (PAHOERC) (PAHO 2018-04-0029). CDC considered the study a public health evaluation (reference number: 2018-063).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Table S1.
Alleles found in samples from Quibdo, 2018. *Pfmdr1 data is already published (10). Tabular data with the alleles found in crt, dhfr, dphs, and mdr1 genes and 7 neutral microsatellites in 42 samples from Quibdo, Colombia, 2018. Red letter indicates a mutant allele. N/A indicates no allele amplification at that locus.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Guerra, A.P., Olivera, M.J., Cortés, L.J. et al. Molecular surveillance for anti-malarial drug resistance and genetic diversity of Plasmodium falciparum after chloroquine and sulfadoxine-pyrimethamine withdrawal in Quibdo, Colombia, 2018. Malar J 21, 306 (2022). https://doi.org/10.1186/s12936-022-04328-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-022-04328-x