
Abstract
Colorectal cancer (CRC) is the third most common cause of cancer globally and the fourth attributable cause of mortality and morbidity due to cancer. An emerging factor contributing to CRC is the gut microbiota and the cellular changes associated with it. Further insights on this may help in the prevention, diagnosis and new therapeutic approaches to colorectal cancer. In most cases of CRC, genetic factors appear to contribute less to its aetiology than environmental and epigenetic factors; therefore, it may be important to investigate these environmental factors, their effects, and the mechanisms that may contribute to this cancer. The gut microbiota has recently been highlighted as a potential risk factor that may affect the structural components of the tumor microenvironment, as well as free radical and enzymatic metabolites directly, or indirectly. Many studies have reported changes in the gut microbiota of patients with colorectal cancer. What is controversial is whether the cancer is the cause or consequence of the change in the microbiota. There is strong evidence supporting both possibilities. The presence of Fusobacterium nucleatum in human colorectal specimens has been demonstrated by RNA-sequencing. F. nucleatum has been shown to express high levels of virulence factors such as FadA, Fap2 and MORN2 proteins. Our review of the published data suggest that F. nucleatum may be a prognostic biomarker of CRC risk, and hence raises the potential of antibiotic treatment of F. nucleatum for the prevention of CRC.
Similar content being viewed by others


Introduction
CRC is one of the most common malignancies of men and women in most countries [1, 2]. The third most common cause of cancer and the fourth leading cause of cancer-related deaths [3]. More than 1.2 million new cases of CRC are reported annually throughout the world [4]. Identification of the microbial mechanisms involved in the etiology of CRC and the recognition of the associated cellular changes as one of the factors in the development of cancer may contribute to cancer prevention, its early diagnosis and potentially new therapies for CRC. The trend is for a projected increase in CRC by 60% to more than 2.2 million new cases, and 1.1 million cancer deaths by the year 2030 [3]. This increase in prevalence has caused considerable debate about the most appropriate prevention approaches. These predictions represent a major problem in developing and developed countries in the public health sector [5]. There has been a global increase in the standardised rate of the age of CRC from 1990 to 2017 with considerable heterogeneity at regional and national level. There has been a decline in the age-standardized death and disability-adjusted life-years rates (DALY) [6]. According to estimates by the DALY criteria, CRC is the world's 36th leading cause of death from disease in 2017 and the fourth most common cause of cancer. The gut microbiota may be one environmental risk factor predisposing to CRC [7]. Bacteria are found throughout the human body, but especially in the digestive tract [8]. The gut at birth is sterile, but some organisms enter it soon after birth. In breastfeed infants, the gut contains a large number of Lactic acid bacteria and Lactobacilli, streptococci, for example bifidobacterium. The gut microflora changes with changes in dietary habits and the selection of adult dietary patterns [9]. Gut bacteria are important for the synthesis of vitamin K, and for the conversion of bile pigments and bile acids to secondary bile acids [10]. In addition, these bacteria are involved in the uptake of food and metabolic products and have antagonistic effects with microbial pathogens. The microbial flora of the gut produces ammonium and other metabolic products absorbed from the intestinal mucosa and can participate in the occurrence of hepatic coma. Anaerobic colonic bacteria, such as Bacteroides fragilis, Clostridium and Peptostreptococcus play a role in the progress of intra-abdominal abscesses [11]. Therefore, intestinal microbes appear to play a crucial role in digestive function and health [12, 13]. It has been proposed that commensal bacteria in the colon play a vital role in the development of CRC [14]. Various studies have shown that chronic infections can be important factors in the development of cancer. Gastric, liver, and cervical cancers are caused by Helicobacter, Hepatitis B and C and human Papillomavirus, respectively [15, 16]. These pathogens activate tumor signaling pathways like NF-kB, STAT3 [15,16,17]. There is good evidence for a relationship between gut microbiota and CRC [18]. This is proposed to be due to the expression of proteins that have antiapoptotic, growth factor or cytokine that enhance cancer cell growth, metastasis or resistance to therapy [17]. However, F. nucleatum has been shown to also express high levels of virulence factors such as FadA, Fap2 and MORN2 proteins [19]. Studies have demonstrated that the dominant microbiome is very similar in primary and metastatic tumors [20]. It is assumed that Fusobacterium moves to distant sites with primary tumor cells as a part of metastatic tissue colonization. This indicates that the tumor microbiomes are the essential components of the cancerous microenvironment [20, 21].
Objectives
In this paper, we aimed to examine the potential role of the gut microbiome, especially F. nucleatum, in inhibiting the immune system in CRC and the stimulatory effects of its surface proteins on the establishment or dissemination of CRC and stimulation of its tumorigenic signals. Also, the role of F. nucleatum and its virulence factors in the development of CRC in particular are systematic reviewed. The cellular signals associated with the creation of tumors activated by bacteria will also be explained.
Search strategy
The protocol was performed in accordance with the preferred reporting items for Systematic reviews and Meta-Analyses (PRISMA) guidelines [22], outlined in Table 1. The following databases were searched: MEDLINE, Embase via PubMed, Scopus, Web of Science database and Google Scholar. A manual search was used to find reference lists of related articles and reviews. In order to locate reference lists of relevant publications and reviews, a manual search was used. The above manual search was made in order to find articles that were not identified by internet searches. The authors were consulted to collect further information in situations where it was needed. Language constraints have been imposed for the search or collection of English publications written in December 2020. The following key- words were used in this search: [(Colorectal[Title/Abstract] OR Intestinal[Title/Abstract]) AND (Neoplasm*[Title/Abstract] OR Carcinoma*[Title/Abstract] OR Cancer*[Title/Abstract] OR Tumor*[Title/Abstract] OR Malignanc*[Title/Abstract])] OR [Adenoma*[Title/Abstract] AND (Colon [Title/Abstract] OR Intestin* [Title/Abstract] OR colonic [Title/Abstract] OR Polypos* [Title/Abstract])] AND [(fecal[Title/Abstract] OR faecal[Title/Abstract] OR feces[Title/Abstract]) AND (Fusobacteri*[Title/Abstract] OR F. nucleatum[Title/Abstract])] AND [Microbio*[Title/Abstract] OR Microbial[Title/Abstract] OR Diet[Title/Abstract] OR Dysbios*[Title/Abstract] OR Dysbacterios*[Title/Abstract] OR MicroRNA[Title/Abstract] OR miRNAs[Title/Abstract] OR “Micro RNA” [Title/Abstract] OR miRNA[Title/Abstract]] OR [Marker*[Title/Abstract] AND (Tumor[Title/Abstract] OR Carcinogen*[Title/Abstract] OR Neoplasm*[Title/Abstract] OR Cancer[Title/Abstract])) OR (Biomarker*[Title/Abstract] AND (Tumor[Title/Abstract] OR Carcinogen*[Title/Abstract] OR Neoplasm*[Title/Abstract] OR Cancer[Title/Abstract])) OR immunomodulator*[Title/Abstract]].
Study selection
Three hundred and ninety one unique records were checked by title and abstract to assess their eligibility for inclusion in the project after finding a total of 497 papers and deleting the duplicate records. The full texts of 202 publications were then checked and the related articles were chosen according to the study inclusion criteria (Fig. 1). The inclusion criteria were: Studies measuring the association of Fusobacterium nucleatum with colorectal cancer in patients and the published studies in English language. The exclusion criteria were: 1—No access to full-text articles 2—Case reports, randomized clinical trials and review articles 3—Studies on teenagers and 4—Duplicate records were excluded. Figure 1 shows the selection process for articles. Data collected using EndNote software. The main characteristics are summarized in Table 1.

PRISMA flowchart of study selection
Colorectal cancer
Cancer usually arises due to the failure of mechanisms controlling cell growth and proliferation. This control system responds to growth inhibition, growth and death signals. Colorectal carcinogenesis involves a series of well-defined changes that begin with a benign mucosal lesion called a polyp and can progress to malignancy leading to cancerous changes such as hyperplasia, adenoma, carcinoma, and metastasis [60]. The molecular mechanisms involved in these changes include activation of specific oncogenes and inactivation of tumor suppressor genes [61, 62]. Cancer is a multifactorial disease due to genetic, epigenetic, and environmental factors [63]. CRC can be asymptomatic for many years. Malignancies on the right-side of colon, including the cecum, ascending, and descending colon are associated with fatigue, weakness and iron deficiency anemia; however, left colon neoplasms are associated with concealed bleeding, alterations in bowel movement, and lower left quadrant cramp. Diagnosis is often made by the detection of fecal occult blood testing followed by endoscopy; this is then followed by biopsy and MRI [64]. One of the changes involved in the development of some cancers, including CRC, is the role of infections on tumor suppressor genes in the initiation, progression, and metastasis of cancer [65].
The microbiota and colorectal cancer
Gut dysbiosis can promote CRC through various processes that include: the induction of a chronic inflammatory disease or immune response, biosynthesis of toxic metabolites and genotoxin and effect of host metabolism [66, 67]. Alternatively, Microbiota can prevent cancer by producing metabolites and enzymes. Although, some bacterial metabolites secreted from Enterococcus faecalis, enterotoxin Bacteroides fragilis or FadA in F. nucleatum are capable of damaging DNA, they can induce proliferation of colon cells in studies on gut microbiota in cancer patients [68]. Gut bacterial composition can be affected by environmental factors and tumour genomics [69]. Most cases of CRC are treatable if a diagnosis is made early enough. The survival rate in patients in whom an early detection is made is approximately 5 times greater than for patients diagnosed with advanced malignancies [70]. Consequently, it is necessary to evaluate valuable early diagnosis markers for CRC cases [70]. In the following, we will discuss the role of F. nucleatum as a parameter in the development and diagnosis of colorectal cancer.
Tumorigenic potential of Fusobacterium nucleatum
Sequences of Fusobacterium species were found to be enriched in colorectal carcinomas [71]. The results were confirmed with the use of quantitative PCR and sequence analysis of 16S rDNA performed on 95 normal-tumor pairs of DNA. In addition, Fusobacteria were observed in colorectal tumors by FISH. According to the obtained results, there are some changes in the microbiota in CRC [71]. F. nucleatum and some common bacteria were found in the primary tumors but also in distant metastases [72]. Preliminary evidence indicates that this bacterium is initially found in cancer cells of metastasis type instead of the stroma. The tumor growth in mice with xenografts from of CRC containing F. nucleatum was reduced following treatment with antibiotics, consistent with the causal role played by bacteria in the development of tumors [20]. Preclinical rodent studies have recently shown that antibiotic therapy or the absence of the gut microbiota reduces the incidence of tumors in several murine colitis-associated CRC models [18]. The frequency of Fusobacterium in human tumors by the RNA-seq method was similar to the one obtained from mice tumors using flow cytometry [18]. In most cases, Fusobacteria are not part of the natural bacteria of the large intestinal flora. Studies show that cancerous tissues contain significantly more Fusobacteria [73]. Previous research has indicated that infection with this bacterium increases the incidence of ulcerative colitis in which inflammation of the intestinal lining destroys the intestinal cells and consequently is a risk factor for colorectal cancer [27].
The impact of diet on the microbiota and colorectal cancer
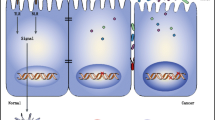
At birth, four main bacterial species are present in the gut: Firmicutes, Bacteriodetes, Proteobacteria and Actinobacteria [74]. They vary greatly among healthy individuals depending on environmental, genetic, host immune system, diet, and exposure to infection or antibiotics [20, 74]. Despite the considerable variation among individuals, it has been found that there are similar microbial populations in colorectum, including anaerobic bacteria such as Bacteroides, Eubacterium, Bifidobacterium, Fusobacterium, Peptostreptococcus, Atopobium and optional anaerobes, including Lactobacilli, Enterococci and Enterobacteria. However, diet, age, gender, and ethnicity affect individual microbes, making its dynamic nature difficult investigate [8, 75]. From the 1990s onward, studies have shown an association between CRC and certain bacterial species [76]. Shen et al., evaluated 21 adenomas and 23 non-adenomas. In cancerous tissues, Proteobacteria is increased, and Bacteroidetes decreased [77]. It is possible that some probiotics facilitate immunomodulatory and anticancer activities in different contexts [10]. For example, lactobacillus in the lactic acid bacteria group is the main probiotic organisms. Various reports have indicated that isolates of Lactobacillus spp. [10] like Lactobacillus acidophilus in different forms may increase the anticancer effects by different mechanisms such as downregulation of ErbB-2, activation of natural killer cells, dendritic cell maturation, and release of probiotic-derived ferrichrome (iron-scavenging peptide) [78, 79]. The microbiome has been called “The forgotten organ” [80, 81]. Microbiota can play a key role in the development of CRC by altering the bacterial composition of the intestine (dysbiosis), high production of some bacterial enzymes, changes in the distribution of bacterial communities and alteration in bacterial metabolic activity [82, 83]. On the other hand, some of the components of the microbiota control the differentiation of intestinal epithelial cells and their proliferation, growth and development of the epithelial barrier, make strong apical bonds, protect against strains of pathogens, fermentation of carbohydrates indigestible for the production of short chain fatty acids (SCFA), bile acid metabolism and destruction of carcinogens in the diet in protection against cancer [84,85,86,87]. Many factors can alter the microenvironments of the digestive tract and consequently the bacterial flora, such as consumption of antibiotics, mental and physical stressors, radiation, and diet [88]. The microbiota play a significant role in the regulation of inflammation, immune response or hematopoiesis among others [89]. Modification of the microbiota may lead to some pathologies such as depression and cancer [90, 91]. Prevention of carcinogenesis by modulating tumor or host cell microenvironment may be possible. Moreover, the microbiota has been found to influence chemotherapy, radiotherapy and immunotherapy efficacy and toxicity [80]. L. casei probiotic-derived ferrichrome has its anti-tumor effect through the pathway contributing to JNK-mediated apoptosis [92]. They are also associated with decreased polarization of pro-inflammatory TH17 cells and consequently a reduction in anti-inflammatory Treg cells differentiation (regulatory T cells) and/or gut Tr1 cells (T regulatory type 1 cells) in addition to anti-inflammatory metabolites production [10, 93]. There is a special association between the microbiome profiles and cancer growth and progression. Consequently, interventions altering microbiome composition are likely to affect oncogenesis (Fig. 1). The microbiome may remain unchanged for many years. However, factors such as response to antibiotic therapy, exposure to pathogens, fasting, changes in daily diet composition and other causes such as stress, cold and diurnal rhythm disruption can cause permanent changes in it [10]. Moreover, according to reports, microbiomes affect various traits ranging from metabolism to mood [10]. The microenvironment of CRC is a complex community of genomically changed tumor cells, non-neoplastic cells, and a varied group of microorganisms [71]. Many genetic and epigenetic factors affect the reported recurrence of the disease; in many studies, the gut microbiome has not been identified as an important factor in the disease occurrence. With the progress of advanced bowel sampling techniques and analysis of both nucleic acid (RNA sequences) and protein (Proteome) products, it has been identified that the gut microbial community is a key component in not only in tumorigenesis but also the non-recurrence of disease after surgery [74, 94]. Most studies on the role of the microbiome on CRC recurrence have been investigated in clinical studies where local recurrence has occurred [74]. F. nucleatum can cause cancer by activating cellular signals through various mechanisms. These mechanisms are important for causing cancer in terms of cell surface receptors and their effects on the immune system (Fig. 2).

The main mechanism of F. nucleatum pathogenesis in CRC is illustrated. The adhesion and invasion of FadA from F. nucleatum to epithelialand endothelial cells of human in pathway 1 can be observed while levels of inflammatory cytokine (IL-6, IL-8,IL-10, IL-18, TNF-α, and NF- κB) grow in a proinflammatory microenvironment which in turn leads to colorectal tumor progression; FadA interaction with E-cadherin in pathway 2 in epithelial cells leads to activating of β-catenin signaling, increasing NF-κB inflammatory gene expression and enhancing tumor cell proliferation. F.nucleatum-infected cells, on the other hand, enhance miRNA expression by Toll-like receptor activation and therefore miRNA release development. F.nucleatum in pathways 3 and 4 reduces the activity of human T cells in a micro-suppressor of the tumor immune system. The interaction between Fap2 from F.nucleatum and the human inhibitor receptor TIGIT in pathway 5 leads to the death of lymphocyte cells of human, resulting in a microenvironment of immunosuppression that increases the progression of CRC
Doll and Peto have previously argued that 30% of the risk of cancer might be attributed to diet. Since then, much available evidence has pointed out that several nutrients can change cancer growth and progression [95]. Long-term dietary habits can shape the gut microbiota [87]. The International Agency for Research on Cancer (IARC), as part of the World Health Organization (WHO), has suggested that there is enough evidence to consider consumption of processed meat (Group 1) and red meat (Group 2A) in humans as having possible carcinogenic effects. IARC analyzed a group of half a million English men and women. In their analysis, they concluded that processed meat and red alcohol were related to increased risk of colorectal cancer. They also demonstrated an association between reduced risk of cancer and fiber in bread and breakfast cereals [96]. The incidence and mortality of polygenic diseases like cancer vary depending on genetic susceptibility and environmental factors. Interaction of specific nutrients on genetic code exists in all nucleated cells [93]. For example, high consumption of refined starches and sugar increases the production of superoxide anion in leukocytes, mononuclear cells and free fatty acids (FFA) [97]. It also increases the levels and activity of the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-kB), a transcriptional regulator activating at least 125 genes, most of which are pro-inflammatory. Glucose intake also increases the two pro-inflammatory transcription factors; activated protein 1 (AP-1) and early growth response protein 1 (Egr-1) [98]. AP-1as a transcription factor induces regulation of the transcription of inflammatory cytokines, matrix metalloproteinase, as well as the transformation of apoptosis and cell proliferation [99]. Egr-1 induced endothelial gene expression [100], and modulation of transcription of tissue factor and plasminogen activator inhibitor-1 (PAI-1) [98, 101]. Increased absorption of linoleic acid, saturated fat, trans fats, refined starches and sugars can increase the production of free radicals and NF-kB activation leading to rapid expression of pro-inflammatory genes [98]. Nutrients, antioxidants, micronutrients, minerals, vitamins, coenzyme Q10 and ω3 fatty acids may inhibit NF-kB superoxide production, AP-1, and Egr-1 [98]. The evidence indicates that dietary fiber, especially starch that is resistant to digestion, enhances intestinal health. One of the issues causing the starch to be the focus of empirical research is its potential protection against CRC development [102]. Other studies have indicated that butyrate (main short-chain fatty acids) from resistant starch fermentation through the bacteria in the gut causes physiological changes in humans [103] and plays a significant role over the lifestyle in protecting the body against deteriorating metabolic control and inflammatory status associated with western lifestyles [104]. Although there is evidence of the cellular effects of butyrate, much research has been conducted to determine which mechanisms of butyrate can be used for antitumor applications [105]. Statistical and bioinformatics analysis was then performed to determine which potentially important genes and proteins are involved in inducing apoptosis of colon cancer cells. Furthermore, 1347 proteins such as isoforms of protein and modifications were detected using proteomics (2D-DIGE and mass spectrometry). Moreover, 139 proteins were identified. These proteins were likely to play a role in the apoptotic response to butyrate [93]. These reactions, along with the microbial population in the gastrointestinal tract, particularly the large intestine, cause the formation of microbiomes, including all microorganisms, their genes and metabolites. Extensive investigations to find out the genetic map of microorganisms are in progress, since microbial genes and their interactions with body cells exist before, during, and after illnesses.
Factors affecting the intestinal microbial population
Most studies indicate that the composition of the intestinal microbiota is formed before the age of three and then has a constant composition throughout life [106, 107]. Some factors such as the use of antibiotics, special diet, and chemotherapy can interfere with the structure of the gut microbiota [108]. Since the microbiota plays an important role in the normal functioning of the body, today it is considered an organ created at birth and evolves with us. The role of the microbiota in the development of some gastrointestinal diseases has been demonstrated [109]. These diseases can range from causing inflammation to colon cancer [110]. The gut microbiota may also sometimes be involved in the development of extra-intestinal immunological diseases [110, 111]. Probiotics such as bifidobacteria, lactobacillus, bacteroids are mainly found in the colorectal and are beneficial for human health. They control the population of pathogenic bacteria by producing short-chain fatty acids such as acetic, butyric and propionic acid. Prebiotics are also a substrate choice for the growth of beneficial bacteria like bifidobacteria. Prebiotic foods include sugars such as inulin and oligofructose (FOS). During breastfeeding, the major bacterium in neonatal feces is bifidobacteria, but during weaning, the level of bifidobacteria is decreased and other bacteria such as coliform, Clostridium and Streptococcus are increased [112]. As more molecular techniques and bioinformatics analyses were developed, a better understanding of a healthy microbiome or disruption of the microbial community, including loss of beneficial bacteria or loss of diversity among them, was achieved [74]. The disorder produces a specific condition called dysbiosis, which means the loss of the health-promoting microbiome known as disease-producing pathogens. Numerous studies have shown that Fusobacterium, Alistipes, Porphyromonadaceae, Coriobacteridae, Staphylococca-ceae, Akkermansia, Lactobacillus, Faecalibacterium, Roseburia and Treponema are present in patients with CRC [58, 113,114,115,116,117,118]. The present findings emphasize the importance of cell-bacterial interactions in a network. Various mechanisms such as aberrant activation of immune cells, induction of DNA damage through production of oxygen and nitrogen species, and increased levels of immunocyte-derived bioactive molecules facilitate tumor progression [16]. Using an antibody neutralization assay, an important role for epithelial expression of TLR2 was identified in this process [119]. These findings are consistent with the recent role of TLRin the development of colorectal cancer.
Fusobacterium as a biomarker in CRC
Fusobacterium is a genus of anaerobic, Gram-negative, non-spore forming bacteria, similar to Bacteroides. F. nucleatum and F. necrophorum are usually members of the Fusobacterium species. They usually reside in the oral cavity and sometimes cause periodontal and gum infections [9]. Fusobacterium is considered a risk factor involved in CRC start and improvement. Immune modulation is considered to be the most important mechanism of Fusobacterium playing a role in CRC carcinogenesis (Fig. 1). It includes increased cells of myeloid-derived suppressor and natural killer cell inhibitors, FadA and Fap2 virulence factors, microRNAs and bacterial metabolism [120, 121].
Carcinogenesis mechanisms of F. nucleatum
Tumorigenesis mechanism of Fusobacteria includes receptors of pattern recognition and downstream inflammation, but these bacteria with the recruitment of myeloid cells lead to infiltration of adenomas and carcinomas, thereby resulting in NF-κB-dependent TLR4 signaling [122]. Recently, it has also been demonstrated that F. nucleatum leads to increased expression of inflammatory mediators (IL1B, IL6 and IL8) [40, 119]. This is possibly due to miRNA-mediated activation of TLR2/TLR4 [75, 119]. In the immune response to bacterial infection, TLRs are highly important. Among them, TLR4 is considered a representative receptor for LPS. When TLR4 is activated by LPS, a series of intracellular events are triggered. This leads to nuclear translocation of NF-κB, thereby increasing the expression of IL-8 gene [123]. However, F. nucleatum doesnot encode any known toxins, while only few common virulence factors are encoded by it. Adhesion protein FadA is a known virulence factor in F. nucleatum contributing to easier attachment and invasion of bacteria [124, 125]. FadA binds with an E-cadherin receptor and increases carcinogenesis. It causes activation of β-catenin and stimulates expression of transcriptional factors, Wnt genes, inflammatory genes, and related oncogenes (Fig. 1) [126]. In this adhesion process, MORN2 may also be involved. However, the exact function of MORN2 is unknown [126]. When FadA adhesin from F. nucleatum binds with CDH1, it causes an increase in the proliferation of Fusobacterium/WNT [126]. FadA and MORN2 proteins of F. nucleatum play a key role in cell invasion [127]. FadA is a small ligand (125 AA) present on the surface of Fusobacterium, which has been shown to bind to E-Cadherin and activates β-catenin signaling in human cancer xenografts of mice models [112]. Thus, FadA binding is directly involved in host cell binding and invasion of Fusobacterium. MORN2 proteins further contain a signal sequence allowing the transfer of small peptides into the periplasmic space and from the outer membrane to the extracellular environment [112]. Among the empirically identified proteins associated with disease severity is Fap2 lectin, a galactose adhesion protein, which binds with the NK Cell Receptor TIGIT and inhibits the destruction of tumor cells by inhibiting NK cells. Fusobacterium binds with the Gal-GalNAc receptor on the surface of colon cancer cells, thereby producing proinflammatory cytokines and proliferating cancer cells [128]. Metagenomic analyses have indicated increased Fusobacterium species in CRC compared to adjacent normal tissue by total genome sequencing, transcriptome sequencing or by 16S rRNA gene sequencing used as a tool to identify bacteria [120, 121, 129]. There has been a correlation between F. nucleatum in CRC, chemo-resistance and poor prognosis. According to what mentioned before, binding of the Fap2 protein of F. nucleatum with the inhibitory receptor TIGIT of human protects tumors from immune cell attack expressed in natural killer (NK) cells. F. nucleatum would also inhibit T and NK cell activities [18]. The following section describes the other five extensive families of pattern recognition receptors (PRRs). They include CLRs (C-type lectin receptors), LRR (nucleotide-binding domain leucine-rich repeat) containing (NOD-like) receptors (NLRs), RLRs (RIG-I-like receptors), ALRs (AIM2-like receptors) and cytoplasmic DNA sensors [122]. Recent research activities emphasize the importance of pathogen–host signaling, by PRRs in the whole range of inflammatory responses, including cancer development and inhibition [130]. PRRs signaling impacts all stages of intestinal cancer, from the early stages of cancer to the metastatic stage and appearance of different cells in the tumor microenvironment, and from neoplastic cells to tumor and stromal cells [122, 130]. Small secretory peptides bind to myeloid-derived suppressor cells (MDSCs), thereby inhibiting CD4+ T cell. Inhibition of immune cells would be desirable for tumor cells, since it will spread the tumor to other parts of the body [112]. Investigation of 16 s rRNA sequencing of increased F. nucleatum levels in mucosal or fecal samples of CRC patients has shown that F. nucleatum levels in CRC tissue is associated with the tumor site of right-sided proximal colorectum and CpG island methylator phenotype (CIMP) status, microsatellite instability (MSI) and mutations in BRAF, KRAS, CHD7, CHD8 and TP53 genes [131]. Increased inflammatory cytokines such as NF-KB, TNF-α, IL10, IL8, IL6, and increased levels of E. cadherin on epithelial cells activates B-catenin signaling, increases NF-κB, C-myc expression and proliferates tumor cells [70]. Cells infected with F. nucleatum, due to activation of Toll-like receptors (TLR), cause more mRNA expression and release. F. nucleatum induces lymphocyte cell death and tumor progression by blocking G1 phase cell myeloid derivative suppressor cells (MDSCs) and TIGIT receptor inhibition [52, 132]. F. nucleatum also affects the IL-6-STAT3 axis signaling and induces tumorigenesis by directly interacting with epithelial cells through activating TLRs. The key molecules stimulating tumor growth and invasion induced by these bacteria include IL-6, cyclin D1, TNFα, MMP9 and heparanase [127].
F. nucleatum, immunomodulatory of the tumor microenvironment
Cancer in its simplest form of uncontrolled cell growth in association with F. nucleatum is likely to affect the proliferation of cancer cells in the colorectal. According to epidemiological associations, F. nucleatum can improve instability and mutation of genes [56]. In the stool of mice with colon cancer, there was a correlation between immunotherapy by antibodies for IL.10 receptor (antiIL10R) and CpG oligodeoxynucleotides with the increased Alistipes shahii. In this model, A. shahii caused an increase in the production of the tumor necrosis factor (TNF) by intrauterine myeloid cells, while TNF neutralization abolished the therapeutic effect [10]. It has been reported that enterotoxigenic B. fragilis stimulate pro-inflammatory Th17 cells that accelerate carcinogenesis in mice prone to the tumor [10, 79]. Compared to different bacterial strains, F. nucleatum can correctly identify patients with CRC. Recent studies have shown that F. nucleatum DNA in the early stages of the disease has the diagnostic potential as a non-invasive primary biomarker for CRC from fecal samples [52]. Fusobacterium is associated with the signature of human CRC gene expression. A correlation of immune cell marker genes, including tumor-associated macrophages (TAMs) (CD209, CD206/MRC1, IL6, IL8, and CXCL10), MDSCs (CD33 and IL6), dendritic cells (DCs) (CD11c/ITGAX, CD209, TNF, and CD80) and Fusobacterium has been found in humans [18]. Some T cell subsets are associated with CRC prognosis. For example, Th1 subsets detected by interferon-gamma secretion (IFNγ) with better prognosis and Th17 identified due to IL-17 production are accompanied by a worse prognosis. Several studies have shown that Fusobacteria, in particular Fusobacterium, is also prevalent in CRC tissues despite being predominant in the oral microbiome. Using tissues from CRC patients that were positive for the 16S ribosomal RNA gene Fuso sequence and Th1 and Th17 cell populations in CRC patients by flow cytometer, there was a positive relation with both IL-17+ and IFNγ+ cytokines. These findings suggest that immune responses in CRC patients (Th1 and Th17) correlate with the frequency of Fusobacterium, especially the nucleatum [56]. Fusobacterium-related genes, including PTGS2 (COX-2), IL1b, IL6, IL8 and TNF, are expressed not only in colon cancer but also in cultures of human and mouse cell lines in vitro known as the central link between inflammation and cancer [18]. In general, the expression of human Fusobacterium-dependent proinflammatory genes is higher in colorectal tumors than in small bowel tumors. This may be due to the anatomical location related to the fact that the listed genes are derived from human CRC [18]. Mouse studies have indicated that the gut microbiome may regulate local immune responses and affect chemotherapy and immunotherapy [74]. In patients with colorectal cancer, autophagy pathways are rich and active and high levels of F. nucleatum cause resistance to chemotherapy [45]. F. nucleatum binds with the host epithelial E-cadherin and stimulates colorectal carcinogenesis through Fusobacterial adhesion FadA [74]. It has also been found that F. nucleatum targets micro-RNA and autophagy Signaling via upregulation of CARD3 expression causing resistance to chemotherapy [26, 45]. The direct association of Fusobacterium with recurrent CRC has even been postulated as a way to predict disease outcomes or change chemotherapy regimens such as inclusion of capecitabine and oxaliplatin for patients with a high burden of F. nucleatum [45]. These observations suggest further consideration of antimicrobial interventions as a potential treatment for patients with CRC related to Fusobacterium [131]. One concern is the negative effect of broad-spectrum antibiotics on the intestinal microbiome [20]. Metronidazole is ideal, since it targets various anaerobic bacteria, including Fusobacterium anaerobic bacteria. Fusobacteria are highly sensitive to metronidazole. Finally, oral administration of metronidazole to mice that were Fusobacterium- positive resulted in a significant decrease in tumor growth pathways. Treatment with metronidazole resulted in a significant reduction in the Fusobacterium burden [20]. However, antibiotics are somewhat similar to cytotoxic chemotherapy, and their treatment is relatively non-targeted. Enterotoxigenic Bacteroides fragilis (ETBF) is a toxin-producing bacterium that can activate TH17-mediated colitis, with simultaneous colon-specific STAT3 activation and tumor stimulation in susceptible ApcMin (Multiple Intestinal Neoplasia) mice, which is reversed by IL-17 antibody blockade [133]. This issue is also considered a limitation for the treatment method. Other bacteria involved in CRC may also respond to tumor progression even beyond antibiotics. Nevertheless, as shown with metronidazole treatment, even in the late stages of the disease, it may response to clear Fusobacterial colonization of carcinomas in experimental mice models [134]. A recent study has shown that colorectal tumors with a high Fusobacterium burden are likely to recur, implying that Fusobacterium-positive tumors may benefit from anti-Fusobacterial treatment [20].
Fusobacterium-associated microRNAs
MicroRNAs (MiRNAs) are non-coding molecules of RNA with approximately a length of 19–25 nucleotides. At the post-transcriptional level, they regulate target genes expression negatively. It has been shown that oncogenic miRNAs (clusters of miR-17-92a and miR-25-106b [13]) play an active role in CRC progression [135]. Moreover, it has been shown that different miRNAs such asmiR-21, miR-224, miR-200c miR-96, miR-135, miR-31, and miR-155 are related to pathogenesis of CRC [136, 137]. The microarray analysis results showed the active role of 49 miRNAs in F. nucleatum induced CRC, while in a Multi-Class-Dif analysis, there was a significant expression of 96 miRNAs in early and advanced stages of CRC with positive infection of F. nucleatum [13]. Among different expressions of miRNAs, miR-4474 and miR-4717 expressions were upregulated in CRC with positive infection of F. nucleatum [13]. Other genes, including CREBBP (CREB-binding protein), STAT1, CAMK2B, PRKACB, JUN, TP53 and EWSR1, which were involved in cancer signaling pathways were dysregulated [13]. MiR-4802 and miR-18a* are abnormally reduced in expression by F. nucleatum that has been also known to induce chemoresistance to oxaliplatin and 5-FU by reduction of apoptosis through the activation of autophagy and TLR4/MYD88 signaling [138]. Enrichment of Fusobacterium species is observed in the microbiota in carcinomas near healthy colonic tissue. They are observed in stool samples obtained from CRC patients at a higher degree compared to healthy controls. In the early stages of tumorigenesis, F. nucleatum that is usually present in the oropharynx [139], is in not only carcinomas but also colorectal adenomas [21].
Perspective and conclusion
The gut microbiota is the largest reservoir of human microbiota. They consist of species of microorganisms living in the gastrointestinal tract in coexistence with the host, reaching a population of tens of 1014 [140]. They include at least 1000 different species of known bacteria containing more than 3 million genes (150 times more than human genes) [140,141,142]. Although more than a thousand different species of bacteria are found in the human gut, only 150 to 170 of them are common in different individuals [141]. Microbiota of each person is distinctive. Therefore, the identification and determination of normal microbiota in different societies and ages are an important factor and a prerequisite for further identification of the influencing factors. A healthy and balanced gut microbiota is the key to ensuring proper digestive function [143]. They also play a crucial role in the immune system and play a vital role in a mucosal barrier [144]. Other important roles of the gut microbiota are to help digest certain foods that the stomach and small intestine cannot digest, produce some vitamins (B and K), help protect against other microorganisms and maintain intestinal integrity. In some cases, a change in the composition of the microbiota can interfere with its balance, called dysbiosis. Intestinal microbiota dysbiosis can cause intestinal diseases such as inflammatory gut disease, irritable gut syndrome, CRC and extra-intestinal diseases such as diabetes, obesity, cardiovascular disease, non-alcoholic fatty liver disease, liver cells and decreased mental health [145,146,147,148]. Researchers at the Wyss Institute at Harvard, engineered the E. coli strain as a probiotic-gut bacterium producing a network of nanofibers that were directly attached to the mucosa [149]. This strain fills the inflamed areas like a patch and protects these areas from environmental factors and gut microbes. This probiotic-based treatment improved rats with chemical agents and increased mucosal healing. Although many studies have focused on the local delivery of anti-inflammatory drugs to fistulae, ulcers, and intestinal inflammation, fewer studies have been conducted on mucosal healing that plays an important role in suppressing these diseases. Matrix contributes to fibrosis to repair intestinal epithelial [149]. This matrix contains Curli nanofibers, known as an intestinal barrier enhancer and an epithelial enhancer. The researchers maintain that this method can produce engineered bacteria that will settle in the gut and secrete the desired biomaterials [149]. The consumption of yogurt, kefir (a kind of yogurt drink), cheese, fruits and vegetables, seafood, avoiding foods made with preservatives and taking probiotic supplements can be effective in maintaining the normal gut microbiota balance [150]. It is still unknown whether F. nucleatum colonization is the outcome or a cause of carcinogenesis or inflammation in colorectal tissue. The results produced some intriguing results representing Fusobacterium sp. as a potential biomarker for colorectal carcinogenesis. Above all, these results provide a mechanistic insight indicative of the mediation of Fusobacterium sp. actions through FadA binding to host epithelial cells’ receptors in order to change the function of barrier, to increase inflammation through modulation of the microenvironment of tumor, and to activate pro-oncogenic signals for CRC promotion. These findings can affect the prevention, diagnosis and treatment of CRC. However, further studies are required for evaluation of the FadA diagnostic potentiality. More certain answers on the temporal order between F. nucleatum and CRC can be found in prospective studies. Even though F. nucleatum colonization may result from colorectal cancer, nevertheless it may play a key role in tumor malignancy increasing, metastasis promoting and antitumor immunity evasion. Some interesting questions have been raised on cancer causes based on the role of F. nucleatum in tumorigenesis. It is possible to reduce the development of cancer through manipulation of bacterial microbiota by fecal microbial implants, probiotics and using antibiotic treatments or vaccination. Although fecal F. nucleatum may be a considered as a proper measurable biological marker for detection of CRC; further research is required to make it clear how it changes in different stages of colon cancer. A combination of microbiome modulation and its products with simpler immunotherapy approaches directly targeting malignant cells could be used in the future for antineoplastic therapy. The importance of this method is in the new anticancer method or enhanced therapeutic drugs against cancer. This method will have positive clinical results for patients with cancer. Oncomicrobiotics (cocktail of bacteria or bacterial products) is a new supportive treatment for cancer improving immune responses by enhanced gut function. Therefore, much more research is needed to be conducted on cross talk between host–bacteria and their virulence proteins that play a role in colorectal carcinogenesis.
Availability of data and materials
Not applicable.
Change history
26 March 2022
A Correction to this paper has been published: https://doi.org/10.1186/s12935-022-02549-3
Abbreviations
- CRC:
-
Colorectal cancer
- DALY:
-
Disability-adjusted life-years rates
- SCFA:
-
Short chain fatty acids
- Tr1 cells:
-
T regulatory type 1 cells
- WHO:
-
World Health Organization
- FFA:
-
Free fatty acids
- NF-Kb:
-
Nuclear factor kappa-light-chain-enhancer of activated B
- AP-1:
-
Activated protein 1
- Egr-1:
-
Early growth response protein 1
- CIMP:
-
CpGIsland methylator phenotype
- MSI:
-
Microsatellite instability
- MDSCs:
-
Myeloid derivative suppressor cells
- antiIL10R:
-
Antibodies for IL.10 receptor
- TNF:
-
Tumor necrosis factor
- TAMs:
-
Tumor-associated macrophages
- IFNγ:
-
Interferon-gamma
References
Society AC. Key statistics for colorectal cancer. Information and resources for cancer: breast, colon, prostate, lung and other forms. 2016.
Fatemi SR, Pourhoseingholi MA, Asadi F, Vahedi M, Pasha S, Alizadeh L, Zali MR. Recurrence and five-year survival in colorectal cancer patients after surgery. Int J Cancer Manag. 2015;8(4):e3439.
Wong MCS, Huang J, Huang JLW, Pang TWY, Choi P, Wang J, Chiang JI, Jiang JY. Global prevalence of colorectal neoplasia: a systematic review and meta-analysis. Clin Gastroenterol Hepatol. 2020;18(3):553-561.e510.
Hasanpour-Heidari S, Fazel A, Semnani S, Khandoozi S-R, Amiriani T, Sedaghat S, Hosseinpoor R, Azarhoush R, Poorabbasi M, Naeimi-Tabiei M. Temporal and geographical variations in colorectal cancer incidence in Northern Iran 2004–2013. Cancer Epidemiol. 2019;59:143–7.
World Health Organization. The world health report 2002: reducing risks, promoting healthy life. Geneva: World Health Organization; 2002.
Safiri S, Sepanlou SG, Ikuta KS, Bisignano C, Salimzadeh H, Delavari A, Ansari R, Roshandel G, Merat S, Fitzmaurice C. The global, regional, and national burden of colorectal cancer and its attributable risk factors in 195 countries and territories, 1990–2017: a systematic analysis for the Global Burden of Disease Study 2017. Lancet Gastroenterol Hepatol. 2019;4:913–33.
Armstrong D, Dregan A, Ashworth M, White P, McGee C, de Lusignan S. The association between colorectal cancer and prior antibiotic prescriptions: case control study. Br J Cancer. 2020;122(912):917.
Kelly D, Yang L, Pei Z. Gut microbiota, fusobacteria, and colorectal cancer. Diseases. 2018;6(4):109.
Bolstad A, Jensen HB, Bakken V. Taxonomy, biology, and periodontal aspects of Fusobacterium nucleatum. Clin Microbiol Rev. 1996;9(1):55–71.
Zitvogel L, Daillère R, Roberti MP, Routy B, Kroemer G. Anticancer effects of the microbiome and its products. Nat Rev Microbiol. 2017;15(8):465–78.
Kozlov A, Bean L, Hill EV, Zhao L, Li E, Wang GP. Molecular identification of bacteria in intra-abdominal abscesses using deep sequencing. Open Forum Infect Dis. 2018;5(2):ofy025.
Brooks GF. Jawetz, Melnick, & Adelberg’s medical microbiology/Geo, F. Brooks…[et al.]. New York: McGraw Hill Medical; 2010.
Feng Y-Y, Zeng D-Z, Tong Y-N, Lu X-X, Dun G-D, Tang B, Zhang Z-J, Ye X-L, Li Q, Xie J-P. Alteration of microRNA-4474/4717 expression and CREB-binding protein in human colorectal cancer tissues infected with Fusobacterium nucleatum. PLoS ONE. 2019;14(4):e0215088.
Wang H-F, Li L-F, Guo S-H, Zeng Q-Y, Ning F, Liu W-L, Zhang G. Evaluation of antibody level against Fusobacterium nucleatum in the serological diagnosis of colorectal cancer. Sci Rep. 2016;6(1):33440.
Chen B, Du G, Guo J, Zhang Y. Bugs, drugs, and cancer: can the microbiome be a potential therapeutic target for cancer management? Drug Discov Today. 2019;24(4):1000–9.
Binder Gallimidi A, Fischman S, Revach B, Bulvik R, Maliutina A, Rubinstein AM, Nussbaum G, Elkin M. Periodontal pathogens Porphyromonas gingivalis and Fusobacterium nucleatum promote tumor progression in an oral-specific chemical carcinogenesis model. Oncotarget. 2015;6(26):22613–23.
Gallimidi AB, Fischman S, Revach B, Bulvik R, Maliutina A, Rubinstein AM, Nussbaum G, Elkin M. Periodontal pathogens Porphyromonas gingivalis and Fusobacterium nucleatum promote tumor progression in an oral-specific chemical carcinogenesis model. Oncotarget. 2015;6(26):22613.
Kostic AD, Chun E, Robertson L, Glickman JN, Gallini CA, Michaud M, Clancy TE, Chung DC, Lochhead P, Hold GL. Fusobacterium nucleatum potentiates intestinal tumorigenesis and modulates the tumor-immune microenvironment. Cell Host Microbe. 2013;14(2):207–15.
Guo P, Tian Z, Kong X, Yang L, Shan X, Dong B, Ding X, Jing X, Jiang C, Jiang N, et al. FadA promotes DNA damage and progression of Fusobacterium nucleatum-induced colorectal cancer through up-regulation of chk2. J Exp Clin Cancer Res. 2020;39(1):202.
Bullman S, Pedamallu CS, Sicinska E, Clancy TE, Zhang X, Cai D, Neuberg D, Huang K, Guevara F, Nelson T. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science. 2017;358(6369):1443–8.
Zhou Z, Chen J, Yao H, Hu H. Fusobacterium and colorectal cancer. Front Oncol. 2018;8:371–371.
Liberati A, Altman DG, Tetzlaff J, Mulrow C, Gøtzsche PC, Ioannidis JPA, Clarke M, Devereaux PJ, Kleijnen J, Moher D. The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate healthcare interventions: explanation and elaboration. BMJ. 2009;339:b2700.
Haruki K, Kosumi K, Hamada T, Twombly TS, Väyrynen JP, Kim SA, Masugi Y, Qian ZR, Mima K, Baba Y, et al. Association of autophagy status with amount of Fusobacterium nucleatum in colorectal cancer. J Pathol. 2020;250(4):397–408.
Okita Y, Koi M, Takeda K, Ross R, Mukherjee B, Koeppe E, Stoffel EM, Galanko JA, McCoy AN, Keku TO, et al. Fusobacterium nucleatum infection correlates with two types of microsatellite alterations in colorectal cancer and triggers DNA damage. Gut Pathog. 2020;12:46.
Chen S, Su T, Zhang Y, Lee A, He J, Ge Q, Wang L, Si J, Zhuo W, Wang L. Fusobacterium nucleatum promotes colorectal cancer metastasis by modulating KRT7-AS/KRT7. Gut Microbes. 2020;11(3):511–25.
Chen Y, Chen Y, Zhang J, Cao P, Su W, Deng Y, Zhan N, Fu X, Huang Y, Dong W. Fusobacterium nucleatum promotes metastasis in colorectal cancer by activating autophagy signaling via the upregulation of CARD3 expression. Theranostics. 2020;10(1):323–39.
Abed J, Maalouf N, Manson AL, Earl AM, Parhi L, Emgård JEM, Klutstein M, Tayeb S, Almogy G, Atlan KA, et al. Colon cancer-associated Fusobacterium nucleatum may originate from the oral cavity and reach colon tumors via the circulatory system. Front Cell Infect Microbiol. 2020;10:400.
Chen Y, Lu Y, Ke Y, Li Y. Prognostic impact of the Fusobacterium nucleatum status in colorectal cancers. Medicine (Baltimore). 2019;98(39):e17221.
Butt J, Jenab M, Pawlita M, Overvad K, Tjonneland A, Olsen A, Boutron-Ruault MC, Carbonnel F, Mancini FR, Kaaks R, et al. Antibody responses to Fusobacterium nucleatum proteins in prediagnostic blood samples are not associated with risk of developing colorectal cancer. Cancer Epidemiol Biomarkers Prev. 2019;28(9):1552–5.
Guven DC, Dizdar O, Alp A, Akdoğan Kittana FN, Karakoc D, Hamaloglu E, Lacin S, Karakas Y, Kilickap S, Hayran M, et al. Analysis of Fusobacterium nucleatum and Streptococcus gallolyticus in saliva of colorectal cancer patients. Biomark Med. 2019;13(9):725–35.
Tunsjø HS, Gundersen G, Rangnes F, Noone JC, Endres A, Bemanian V. Detection of Fusobacterium nucleatum in stool and colonic tissues from Norwegian colorectal cancer patients. Eur J Clin Microbiol Infect Dis. 2019;38(7):1367–76.
Kunzmann AT, Proença MA, Jordao HW, Jiraskova K, Schneiderova M, Levy M, Liska V, Buchler T, Vodickova L, Vymetalkova V, et al. Fusobacterium nucleatum tumor DNA levels are associated with survival in colorectal cancer patients. Eur J Clin Microbiol Infect Dis. 2019;38(10):1891–9.
Zhang S, Yang Y, Weng W, Guo B, Cai G, Ma Y, Cai S. Fusobacterium nucleatum promotes chemoresistance to 5-fluorouracil by upregulation of BIRC3 expression in colorectal cancer. J Exp Clin Cancer Res. 2019;38(1):14.
Lee DW, Han SW, Kang JK, Bae JM, Kim HP, Won JK, Jeong SY, Park KJ, Kang GH, Kim TY. Association between Fusobacterium nucleatum, pathway mutation, and patient prognosis in colorectal cancer. Ann Surg Oncol. 2018;25(11):3389–95.
Yamaoka Y, Suehiro Y, Hashimoto S, Hoshida T, Fujimoto M, Watanabe M, Imanaga D, Sakai K, Matsumoto T, Nishioka M, et al. Fusobacterium nucleatum as a prognostic marker of colorectal cancer in a Japanese population. J Gastroenterol. 2018;53(4):517–24.
Hamada T, Zhang X, Mima K, Bullman S, Sukawa Y, Nowak JA, Kosumi K, Masugi Y, Twombly TS, Cao Y, et al. Fusobacterium nucleatum in colorectal cancer relates to immune response differentially by tumor microsatellite instability status. Cancer Immunol Res. 2018;6(11):1327–36.
Chen T, Li Q, Zhang X, Long R, Wu Y, Wu J, Fu X. TOX expression decreases with progression of colorectal cancers and is associated with CD4 T-cell density and Fusobacterium nucleatum infection. Hum Pathol. 2018;79:93–101.
Liu L, Tabung FK, Zhang X, Nowak JA, Qian ZR, Hamada T, Nevo D, Bullman S, Mima K, Kosumi K, et al. Diets that promote colon inflammation associate with risk of colorectal carcinomas that contain Fusobacterium nucleatum. Clin Gastroenterol Hepatol. 2018;16(10):1622-1631.e1623.
Guo S, Li L, Xu B, Li M, Zeng Q, Xiao H, Xue Y, Wu Y, Wang Y, Liu W, et al. A simple and novel fecal biomarker for colorectal cancer: ratio of Fusobacterium nucleatum to probiotics populations, based on their antagonistic effect. Clin Chem. 2018;64(9):1327–37.
Proença MA, Biselli JM, Succi M, Severino FE, Berardinelli GN, Caetano A, Reis RM, Hughes DJ, Silva AE. Relationship between Fusobacterium nucleatum, inflammatory mediators and microRNAs in colorectal carcinogenesis. World J Gastroenterol. 2018;24(47):5351–65.
Chen Y, Peng Y, Yu J, Chen T, Wu Y, Shi L, Li Q, Wu J, Fu X. Invasive Fusobacterium nucleatum activates beta-catenin signaling in colorectal cancer via a TLR4/P-PAK1 cascade. Oncotarget. 2017;8(19):31802–14.
Yan X, Liu L, Li H, Qin H, Sun Z. Clinical significance of Fusobacterium nucleatum, epithelial–mesenchymal transition, and cancer stem cell markers in stage III/IV colorectal cancer patients. Onco Targets Ther. 2017;10:5031–46.
Suehiro Y, Sakai K, Nishioka M, Hashimoto S, Takami T, Higaki S, Shindo Y, Hazama S, Oka M, Nagano H, et al. Highly sensitive stool DNA testing of Fusobacterium nucleatum as a marker for detection of colorectal tumours in a Japanese population. Ann Clin Biochem. 2017;54(1):86–91.
Ye X, Wang R, Bhattacharya R, Boulbes DR, Fan F, Xia L, Adoni H, Ajami NJ, Wong MC, Smith DP, et al. Fusobacteriumnucleatum subspecies animalis influences proinflammatory cytokine expression and monocyte activation in human colorectal tumors. Cancer Prev Res (Phila). 2017;10(7):398–409.
Yu T, Guo F, Yu Y, Sun T, Ma D, Han J, Qian Y, Kryczek I, Sun D, Nagarsheth N, et al. Fusobacterium nucleatum promotes chemoresistance to colorectal cancer by modulating autophagy. Cell. 2017;170(3):548-563.e516.
Mehta RS, Nishihara R, Cao Y, Song M, Mima K, Qian ZR, Nowak JA, Kosumi K, Hamada T, Masugi Y, et al. Association of dietary patterns with risk of colorectal cancer subtypes classified by Fusobacteriumnucleatum in tumor tissue. JAMA Oncol. 2017;3(7):921–7.
Amitay EL, Werner S, Vital M, Pieper DH, Hofler D, Gierse IJ, Butt J, Balavarca Y, Cuk K, Brenner H. Fusobacterium and colorectal cancer: causal factor or passenger? Results from a large colorectal cancer screening study. Carcinogenesis. 2017;38(8):781–8.
Mima K, Cao Y, Chan AT, Qian ZR, Nowak JA, Masugi Y, Shi Y, Song M, da Silva A, Gu M, et al. Fusobacterium nucleatum in colorectal carcinoma tissue according to tumor location. Clin Transl Gastroenterol. 2016;7(11):e200.
Nosho K, Sukawa Y, Adachi Y, Ito M, Mitsuhashi K, Kurihara H, Kanno S, Yamamoto I, Ishigami K, Igarashi H, et al. Association of Fusobacterium nucleatum with immunity and molecular alterations in colorectal cancer. World J Gastroenterol. 2016;22(2):557–66.
Li YY, Ge QX, Cao J, Zhou YJ, Du YL, Shen B, Wan YJ, Nie YQ. Association of Fusobacterium nucleatum infection with colorectal cancer in Chinese patients. World J Gastroenterol. 2016;22(11):3227–33.
Mima K, Nishihara R, Qian ZR, Cao Y, Sukawa Y, Nowak JA, Yang J, Dou R, Masugi Y, Song M, et al. Fusobacterium nucleatum in colorectal carcinoma tissue and patient prognosis. Gut. 2016;65(12):1973–80.
Wang HF, Li LF, Guo SH, Zeng QY, Ning F, Liu WL, Zhang G. Evaluation of antibody level against Fusobacterium nucleatum in the serological diagnosis of colorectal cancer. Sci Rep. 2016;6:33440.
Fukugaiti MH, Ignacio A, Fernandes MR, Ribeiro Júnior U, Nakano V, Avila-Campos MJ. High occurrence of Fusobacterium nucleatum and Clostridium difficile in the intestinal microbiota of colorectal carcinoma patients. Braz J Microbiol. 2015;46(4):1135–40.
Mima K, Sukawa Y, Nishihara R, Qian ZR, Yamauchi M, Inamura K, Kim SA, Masuda A, Nowak JA, Nosho K, et al. Fusobacterium nucleatum and T cells in colorectal carcinoma. JAMA Oncol. 2015;1(5):653–61.
Ito M, Kanno S, Nosho K, Sukawa Y, Mitsuhashi K, Kurihara H, Igarashi H, Takahashi T, Tachibana M, Takahashi H, et al. Association of Fusobacterium nucleatum with clinical and molecular features in colorectal serrated pathway. Int J Cancer. 2015;137(6):1258–68.
Tahara T, Yamamoto E, Suzuki H, Maruyama R, Chung W, Garriga J, Jelinek J, Yamano HO, Sugai T, An B, et al. Fusobacterium in colonic flora and molecular features of colorectal carcinoma. Cancer Res. 2014;74(5):1311–8.
Flanagan L, Schmid J, Ebert M, Soucek P, Kunicka T, Liska V, Bruha J, Neary P, Dezeeuw N, Tommasino M, et al. Fusobacteriumnucleatum associates with stages of colorectal neoplasia development, colorectal cancer and disease outcome. Eur J Clin Microbiol Infect Dis. 2014;33(8):1381–90.
McCoy AN, Araújo-Pérez F, Azcárate-Peril A, Yeh JJ, Sandler RS, Keku TO. Fusobacterium is associated with colorectal adenomas. PLoS ONE. 2013;8(1):e53653.
Castellarin M, Warren RL, Freeman JD, Dreolini L, Krzywinski M, Strauss J, Barnes R, Watson P, Allen-Vercoe E, Moore RA, et al. Fusobacterium nucleatum infection is prevalent in human colorectal carcinoma. Genome Res. 2012;22(2):299–306.
Tanaka T. Colorectal carcinogenesis: review of human and experimental animal studies. J Carcinog. 2009;8:5–5.
Lodish H, Berk A, Kaiser CA, Krieger M, Scott MP, Bretscher A, Ploegh H, Matsudaira P. Molecular cell biology. New York: Macmillan; 2008.
Villa TG, Vinas M. New weapons to control bacterial growth. Berlin: Springer; 2016.
Vatandoost N, Ghanbari J, Mojaver M, Avan A, Ghayour-Mobarhan M, Nedaeinia R, Salehi R. Early detection of colorectal cancer: from conventional methods to novel biomarkers. J Cancer Res Clin Oncol. 2015;42:341–51.
Group COoSTS. A comparison of laparoscopically assisted and open colectomy for colon cancer. N Engl J Med. 2004;350(20):2050–9.
Guzman-Pruneda FA, Husain SG, Jones CD, Beal EW, Porter E, Grove M, Moffatt-Bruce S, Schmidt CR. Compliance with preoperative care measures reduces surgical site infection after colorectal operation. J Surg Oncol. 2019;119(4):497–502.
Zou S, Fang L, Lee M-H. Dysbiosis of gut microbiota in promoting the development of colorectal cancer. Gastroenterol Rep (Oxf). 2018;6(1):1–12.
Wieczorska K, Stolarek M, Stec R. The role of the gut microbiome in colorectal cancer: where are we? Where are we going? Clin Colorectal Cancer. 2020;19(1):5–12.
Baliou S, Adamaki M, Spandidos DA, Kyriakopoulos AM, Christodoulou I, Zoumpourlis V. The microbiome, its molecular mechanisms and its potential as a therapeutic strategy against colorectal carcinogenesis. World Acad Sci J. 2019;1(1):3–19.
Sarhadi V, Lahti L, Saberi F, Youssef O, Kokkola A, Karla T, Tikkanen M, Rautelin H, Puolakkainen P, Salehi R, et al. Gut microbiota and host gene mutations in colorectal cancer patients and controls of Iranian and finnish origin. Anticancer Res. 2020;40(3):1325–34.
Shang F-M, Liu H-L. Fusobacterium nucleatum and colorectal cancer: a review. World J Gastrointest Oncol. 2018;10(3):71.
Kostic AD, Gevers D, Pedamallu CS, Michaud M, Duke F, Earl AM, Ojesina AI, Jung J, Bass AJ, Tabernero J, et al. Genomic analysis identifies association of Fusobacterium with colorectal carcinoma. Genome Res. 2012;22(2):292–8.
Okita Y, Koi M, Takeda K, Ross R, Mukherjee B, Koeppe E, Stoffel EM, Galanko JA, McCoy AN, Keku TO, et al. Fusobacteriumnucleatum infection correlates with two types of microsatellite alterations in colorectal cancer and triggers DNA damage. Gut Pathog. 2020;12(1):46.
Lee SA, Liu F, Riordan SM, Lee CS, Zhang L. Global investigations of Fusobacterium nucleatum in human colorectal cancer. Front Oncol. 2019;9:566–566.
Gaines S, Shao C, Hyman N, Alverdy JC. Gut microbiome influences on anastomotic leak and recurrence rates following colorectal cancer surgery. Br J Surg. 2018;105(2):e131–41.
Nie Y, Luo F, Lin Q. Dietary nutrition and gut microflora: a promising target for treating diseases. Trends Food Sci Technol. 2018;75:72–80.
Gorbach SL, Goldin BR. The intestinal microflora and tbe colon cancer connection. Rev Infect Dis. 1990;12(Supplement_2):S252–61.
Shen XJ, Rawls JF, Randall TA, Burcall L, Mpande C, Jenkins N, Jovov B, Abdo Z, Sandler RS, Keku TO. Molecular characterization of mucosal adherent bacteria and associations with colorectal adenomas. Gut Microbes. 2010;1(3):138–47.
Rahbar Saadat Y, Yari Khosroushahi A, Pourghassem Gargari B. A comprehensive review of anticancer, immunomodulatory and health beneficial effects of the lactic acid bacteria exopolysaccharides. Carbohyd Polym. 2019;217:79–89.
Zitvogel L, Daillère R, Roberti MP, Routy B, Kroemer G. Anticancer effects of the microbiome and its products. Nat Rev Microbiol. 2017;15:465.
Bosch TC, Miller DJ. The holobiont imperative, vol. 10. Vienna: Springer; 2016. p. 978–973.
Saei AA, Barzegari A. The microbiome: the forgotten organ of the astronaut’s body–probiotics beyond terrestrial limits. Future Microbiol. 2012;7(9):1037–46.
Lin C, Cai X, Zhang J, Wang W, Sheng Q, Hua H, Zhou X. Role of gut microbiota in the development and treatment of colorectal cancer. Digestion. 2019;100(1):72–8.
Boulangé CL, Neves AL, Chilloux J, Nicholson JK, Dumas M-E. Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 2016;8(1):42.
Forgie AJ, Fouhse JM, Willing BP. Diet-microbe-host interactions that affect gut mucosal integrity and infection resistance. Front Immunol. 2019;10:1802.
Zeng H, Lazarova DL, Bordonaro M. Mechanisms linking dietary fiber, gut microbiota and colon cancer prevention. World J Gastrointest Oncol. 2014;6(2):41–51.
Zeng H, Umar S, Rust B, Lazarova D, Bordonaro M. Secondary bile acids and short chain fatty acids in the colon: a focus on colonic microbiome, cell proliferation, inflammation, and cancer. Int J Mol Sci. 2019;20(5):1214.
Cai R, Cheng C, Chen J, Xu X, Ding C, Gu B. Interactions of commensal and pathogenic microorganisms with the mucus layer in the colon. Gut Microbes. 2020;11:680–90.
Dudek-Wicher RK, Junka A, Bartoszewicz M. The influence of antibiotics and dietary components on gut microbiota. Prz Gastroenterol. 2018;13(2):85–92.
Xu D. Regulation of inflammatory signaling in health and disease, vol. 1024. Berlin: Springer; 2017.
Clapp M, Aurora N, Herrera L, Bhatia M, Wilen E, Wakefield S. Gut microbiota’s effect on mental health: the gut-brain axis. Clin Pract. 2017;7(4):987–987.
Vivarelli S, Salemi R, Candido S, Falzone L, Santagati M, Stefani S, Torino F, Banna GL, Tonini G, Libra M. Gut microbiota and cancer: from pathogenesis to therapy. Cancers. 2019;11(1):38.
Konishi H, Fujiya M, Tanaka H, Ueno N, Moriichi K, Sasajima J, Ikuta K, Akutsu H, Tanabe H, Kohgo Y. Probiotic-derived ferrichrome inhibits colon cancer progression via JNK-mediated apoptosis. Nat Commun. 2016;7:12365.
Fenech M, El-Sohemy A, Cahill L, Ferguson LR, French T-AC, Tai ES, Milner J, Koh W-P, Xie L, Zucker M. Nutrigenetics and nutrigenomics: viewpoints on the current status and applications in nutrition research and practice. Lifestyle Genom. 2011;4(2):69–89.
Gaines S, Williamson AJ, Hyman N, Kandel J. How the microbiome is shaping our understanding of cancer biology and its treatment. Semin Colon Rectal Surg. 2018;29(1):12–6.
Doll R, Peto R. The causes of cancer: quantitative estimates of avoidable risks of cancer in the United States today. J Natl Cancer Inst. 1981;66(6):1192–308.
Bradbury KE, Murphy N, Key TJ. Diet and colorectal cancer in UK Biobank: a prospective study. Int J Epidemiol. 2019;49:246–58.
Esposito K, Giugliano D. Diet and inflammation: a link to metabolic and cardiovascular diseases. Eur Heart J. 2005;27(1):15–20.
Mishra S, Singh R, Dwivedi S, De Meester F, Rybar R, Pella D, Fedacko J, Juneja LR. Effects of nutraceuticals on genetic expressions. Open Nutra J. 2009;2:70–80.
Gazon H, Barbeau B, Mesnard J-M, Peloponese J-M Jr. Hijacking of the AP-1 signaling pathway during development of ATL. Front Microbiol. 2018;8:2686.
Khachigian LM, Lindner V, Williams AJ, Collins T. Egr-1-induced endothelial gene expression: a common theme in vascular injury. Science. 1996;271(5254):1427–31.
Ghanim H, Aljada A, Hofmeyer D, Dandona P. Increased Egr-1, tissue factor and plasminogen activator inhibitor-1 in circulating mononuclear cells of the obese. Diabetes. 2004;53
Cassidy A, Bingham SA, Cummings JH. Starch intake and colorectal cancer risk: an international comparison. Br J Cancer. 1994;69(5):937–42.
Brouns F, Kettlitz B, Arrigoni E. Resistant starch and “the butyrate revolution.” Trends Food Sci Technol. 2002;13(8):251–61.
Morrison DJ, Preston T. Formation of short chain fatty acids by the gut microbiota and their impact on human metabolism. Gut Microbes. 2016;7(3):189–200.
Chen D, Jin D, Huang S, Wu J, Xu M, Liu T, Dong W, Liu X, Wang S, Zhong W, et al. Clostridium butyricum, a butyrate-producing probiotic, inhibits intestinal tumor development through modulating Wnt signaling and gut microbiota. Cancer Lett. 2020;469:456–67.
Derrien M, Alvarez A-S, de Vos WM. The gut microbiota in the first decade of life. Trends Microbiol. 2019;27(12):997–1010.
Odamaki T, Kato K, Sugahara H, Hashikura N, Takahashi S, Xiao J-Z, Abe F, Osawa R. Age-related changes in gut microbiota composition from newborn to centenarian: a cross-sectional study. BMC Microbiol. 2016;16:90–90.
Rodríguez JM, Murphy K, Stanton C, Ross RP, Kober OI, Juge N, Avershina E, Rudi K, Narbad A, Jenmalm MC, et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb Ecol Health Dis. 2015;26:26050–26050.
Holscher HD. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes. 2017;8(2):172–84.
Deehan EC, Walter J. The fiber gap and the disappearing gut microbiome: implications for human nutrition. Trends Endocrinol Metab. 2016;27(5):239–42.
Maynard CL, Elson CO, Hatton RD, Weaver CT. Reciprocal interactions of the intestinal microbiota and immune system. Nature. 2012;489(7415):231–41.
Umana A, Sanders BE, Yoo CC, Casasanta MA, Udayasuryan B, Verbridge SS, Slade DJ. Reevaluating the Fusobacterium virulence factor landscape. bioRxiv. 2019:534297
Yang Y, Jobin C. Novel insights into microbiome in colitis and colorectal cancer. Curr Opin Gastroenterol. 2017;33(6):422–7.
Chen W, Liu F, Ling Z, Tong X, Xiang C. Human intestinal lumen and mucosa-associated microbiota in patients with colorectal cancer. PLoS ONE. 2012;7(6):e39743–e39743.
Zhu Q, Jin Z, Wu W, Gao R, Guo B, Gao Z, Yang Y, Qin H. Analysis of the intestinal lumen microbiota in an animal model of colorectal cancer. PLoS ONE. 2014;9(6):e90849–e90849.
Everard A, Belzer C, Geurts L, Ouwerkerk JP, Druart C, Bindels LB, Guiot Y, Derrien M, Muccioli GG, Delzenne NM, et al. Cross-talk between Akkermansiamuciniphila and intestinal epithelium controls diet-induced obesity. Proc Natl Acad Sci. 2013;110(22):9066–71.
Mandal P. Potential biomarkers associated with oxidative stress for risk assessment of colorectal cancer. Naunyn Schmiedebergs Arch Pharmacol. 2017;390(6):557–65.
Sobhani I, Tap J, Roudot-Thoraval F, Roperch JP, Letulle S, Langella P, Corthier G, Tran Van Nhieu J, Furet JP. Microbial dysbiosis in colorectal cancer (CRC) patients. PLoS ONE. 2011;6(1):e16393–e16393.
Temraz S, Nassar F, Nasr R, Charafeddine M, Mukherji D, Shamseddine A. Gut microbiome: a promising biomarker for immunotherapy in colorectal cancer. Int J Mol Sci. 2019;20(17):4155.
Han YW. Fusobacterium nucleatum: a commensal-turned pathogen. Curr Opin Microbiol. 2015;23:141–7.
Huang S, Yang Z, Zou D, Dong D, Liu A, Liu W, Huang L. Rapid detection of nusG and fadA in Fusobacterium nucleatum by loop-mediated isothermal amplification. J Med Microbiol. 2016;65(8):760–9.
Siegel SJ, Rakoff-Nahoum S. Innate immune pattern recognition and the development of intestinal cancer. In: Microbiome and cancer. Springer; 2019:299–316.
Miyata N, Hayashi Y, Hayashi S, Sato K, Hirai Y, Yamamoto H, Sugano K. Lipopolysaccharides from non-Helicobacter pylori gastric bacteria potently stimulate interleukin-8 production in gastric epithelial cells. Clin Transl Gastroenterol. 2019;10(3):e00024.
Xu M, Yamada M, Li M, Liu H, Chen SG, Han YW. FadA from Fusobacterium nucleatum utilizes both secreted and nonsecreted forms for functional oligomerization for attachment and invasion of host cells. J Biol Chem. 2007;282(34):25000–9.
Ding Q, Tan KS. The danger signal extracellular ATP is an inducer of Fusobacterium nucleatum biofilm dispersal. Front Cell Infect Microbiol. 2016;6:155.
Rubinstein MR, Wang X, Liu W, Hao Y, Cai G, Han YW. Fusobacterium nucleatum promotes colorectal carcinogenesis by modulating E-cadherin/β-catenin signaling via its FadA adhesin. Cell Host Microbe. 2013;14(2):195–206.
Liu J, Hsieh C-L, Gelincik O, Devolder B, Sei S, Zhang S, Lipkin SM, Chang Y-F. Proteomic characterization of outer membrane vesicles from gut mucosa-derived Fusobacterium nucleatum. J Proteomics. 2019;195:125–37.
Ganesan K, Guo S, Fayyaz S, Zhang G, Xu B. Targeting programmed Fusobacterium nucleatum Fap2 for colorectal cancer therapy. Cancers. 2019;11(10):1592.
Sun T, Liu S, Zhou Y, Yao Z, Zhang D, Cao S, Wei Z, Tan B, Li Y, Lian Z, et al. Evolutionary biologic changes of gut microbiota in an “adenoma-carcinoma sequence” mouse colorectal cancer model induced by 1, 2-dimethylhydrazine. Oncotarget. 2017;8(1):444–57.
Bahia D, Satoskar AR, Dussurget O. Editorial: Cell signaling in host-pathogen interactions: the host point of view. Front Immunol. 2018;9:221–221.
Saito K, Koido S, Odamaki T, Kajihara M, Kato K, Horiuchi S, Adachi S, Arakawa H, Yoshida S, Akasu T. Metagenomic analyses of the gut microbiota associated with colorectal adenoma. PLoS ONE. 2019;14(2):e0212406.
Sun C-H, Li B-B, Wang B, Zhao J, Li T-T, Li W-B, Tang D, Qiu M-J, Wang X-C, Zhu C-M. The role of Fusobacterium nucleatum in colorectal cancer: from carcinogenesis to clinical management. Chron Dis Transl Med. 2019;5:178–87.
Bhatt AP, Redinbo MR, Bultman SJ. The role of the microbiome in cancer development and therapy. CA Cancer J Clin. 2017;67(4):326–44.
Brennan CA, Garrett WS. Fusobacterium nucleatum—symbiont, opportunist and oncobacterium. Nat Rev Microbiol. 2019;17(3):156–66.
Nedaeinia R, Sharifi M, Avan A, Kazemi M, Nabinejad A, Ferns GA, Ghayour-Mobarhan M, Salehi R. Inhibition of microRNA-21 via locked nucleic acid-anti-miR suppressed metastatic features of colorectal cancer cells through modulation of programmed cell death 4. Tumor Biol. 2017;39(3):1010428317692261.
Ding L, Lan Z, Xiong X, Ao H, Feng Y, Gu H, Yu M, Cui Q. The dual role of microRNAs in colorectal cancer progression. Int J Mol Sci. 2018;19(9):2791.
Sun Y, Liu Y, Cogdell D, Calin GA, Sun B, Kopetz S, Hamilton SR, Zhang W. Examining plasma microRNA markers for colorectal cancer at different stages. Oncotarget. 2016;7(10):11434–49.
Anfossi S, Calin GA. Gut microbiota: a new player in regulating immune-and chemo-therapy efficacy. Cancer Drug Resist. 2020;3:356.
Riordan T. Human infection with Fusobacterium necrophorum (Necrobacillosis), with a focus on Lemierre’s syndrome. Clin Microbiol Rev. 2007;20(4):622–59.
Kaser A, Zeissig S, Blumberg RS. Inflammatory bowel disease. Annu Rev Immunol. 2010;28(1):573–621.
Qin J, Li R, Raes J, Arumugam M, Burgdorf KS, Manichanh C, Nielsen T, Pons N, Levenez F, Yamada T, et al. A human gut microbial gene catalogue established by metagenomic sequencing. Nature. 2010;464(7285):59–65.
Haque SZ, Haque M. The ecological community of commensal, symbiotic, and pathogenic gastrointestinal microorganisms—an appraisal. Clin Exp Gastroenterol. 2017;10:91–103.
Lazar V, Ditu L-M, Pircalabioru GG, Gheorghe I, Curutiu C, Holban AM, Picu A, Petcu L, Chifiriuc MC. Aspects of gut microbiota and immune system interactions in infectious diseases, immunopathology, and cancer. Front Immunol. 2018;9:1830–1830.
Mezouar S, Chantran Y, Michel J, Fabre A, Dubus J-C, Leone M, Sereme Y, Mège J-L, Ranque S, Desnues B, et al. Microbiome and the immune system: from a healthy steady-state to allergy associated disruption. Hum Microbiome J. 2018;10:11–20.
Belkaid Y, Hand TW. Role of the microbiota in immunity and inflammation. Cell. 2014;157(1):121–41.
Jin M, Qian Z, Yin J, Xu W, Zhou X. The role of intestinal microbiota in cardiovascular disease. J Cell Mol Med. 2019;23(4):2343–50.
Butler MI, Morkl S, Sandhu KV, Cryan JF, Dinan TG. The gut microbiome and mental health: what should we tell our patients?: Le microbiote Intestinal et la Sante Mentale : que Devrions-Nous dire a nos Patients? Can J Psychiatry. 2019;64(11):747–60.
Jasirwan COM, Lesmana CRA, Hasan I, Sulaiman AS, Gani RA. The role of gut microbiota in non-alcoholic fatty liver disease: pathways of mechanisms. Biosci Microbiota Food Health. 2019;38(3):81–8.
Praveschotinunt P, Duraj-Thatte AM, Gelfat I, Bahl F, Chou DB, Joshi NS. Engineered E. coli Nissle 1917 for the delivery of matrix-tethered therapeutic domains to the gut. Nat Commun. 2019;10(1):5580.
Fernandez MA, Marette A. Potential health benefits of combining yogurt and fruits based on their probiotic and prebiotic properties. Adv Nutr. 2017;8(1):155S-164S.
Acknowledgements
We would like to express our gratitude to Research Child Growth and Development Research Center, Research Institute for Primordial Prevention of Non-Communicable Disease, Isfahan University of Medical Science. Isfahan, Iran.
Funding
This study was supported by a research grant, No 299083, from Vice-chancellery for research, Isfahan University of Medical Science, Isfahan, Iran.
Author information
Authors and Affiliations
Contributions
RN and RS designed research; MR and RN wrote the manuscript; SHJ, LR, HF, MR, SJ and MM, contributed equal time and effort in the investigation, research of the manuscript and designing of the figure. GAF and MGM revising the manuscript critically for important intellectual content. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no conflicts of interest for this article.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ranjbar, M., Salehi, R., Haghjooy Javanmard, S. et al. The dysbiosis signature of Fusobacterium nucleatum in colorectal cancer-cause or consequences? A systematic review. Cancer Cell Int 21, 194 (2021). https://doi.org/10.1186/s12935-021-01886-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-021-01886-z