
Abstract
Background
The methylotrophic budding yeast Pichia pastoris GS115 is a powerful expression system and hundreds of heterologous proteins have been successfully expressed in this strain. Recently, P. pastoris has also been exploited as an attractive cell factory for the production of high-value biochemicals due to Generally Recognized as Safe (GRAS) status and high growth rate of this yeast strain. However, appropriate regulation of metabolic flux distribution between cell growth and product biosynthesis is still a cumbersome task for achieving efficient biochemical production.
Results
In this study, P. pastoris was exploited for high inositol production using an effective dynamic regulation strategy. Through enhancing native inositol biosynthesis pathway, knocking out inositol transporters, and slowing down carbon flux of glycolysis, an inositol-producing mutant was successfully developed and low inositol production of 0.71 g/L was obtained. The inositol production was further improved by 12.7% through introduction of heterologous inositol-3-phosphate synthase (IPS) and inositol monophosphatase (IMP) which catalyzed the rate-limiting steps for inositol biosynthesis. To control metabolic flux distribution between cell growth and inositol production, the promoters of glucose-6-phosphate dehydrogenase (ZWF), glucose-6-phosphate isomerase (PGI) and 6-phosphofructokinase (PFK1) genes were replaced with a glycerol inducible promoter. Consequently, the mutant strain could be switched from growth mode to production mode by supplementing glycerol and glucose sequentially, leading to an increase of about 4.9-fold in inositol formation. Ultimately, the dissolved oxygen condition in high-cell-density fermentation was optimized, resulting in a high production of 30.71 g/L inositol (~ 40-fold higher than the baseline strain).
Conclusions
The GRAS P. pastoris was engineered as an efficient inositol producer for the first time. Dynamic regulation of cell growth and inositol production was achieved via substrate-dependent modulation of glycolysis and pentose phosphate pathways and the highest inositol titer reported to date by a yeast cell factory was obtained. Results from this study provide valuable guidance for engineering of P. pastoris for the production of other high-value bioproducts.
Similar content being viewed by others


Background
Myo-inositol (inositol) is a carbocyclic sugar widely distributed in plants, animals and microorganisms [1]. Inositol has nine possible structural isomers, which have been extensively characterized and studied [2, 3]. Although the requirements of inositol for monogastric animals and humans are generally met by their cellular biosynthesis, dietary inositol has been found to be beneficial for treating neurological and endocrine diseases such as depression, panic disorder, diabetes and insulin resistance [4, 5]. Recent studies also demonstrated that inositol and its derivatives have an anti-aging effect on mammals including humans [6, 7]. For most of the aquatic animals, inositol is regarded as an essential nutrient and its deficiency could lead to decreased growth rate, fin erosion, dark skin colouration, and fatty liver disease [8,9,10]. Therefore, inositol is broadly applied as aquaculture feed supplement. In addition, due to its positive effect on lipid metabolism, bone formation and skeletal muscle metabolism [11, 12], inositol is also used as an active supplement in functional food and cosmetics industries.
The production of inositol can be achieved by phytate hydrolysis, in vitro enzymatic synthesis, and in vivo microbial biosynthesis [13, 14]. At present, inositol is mainly produced by phytate hydrolysis under harsh conditions (low pH, high temperature and pressure) which is unfriendly to the environment. Recently, in vitro enzymatic synthesis routes were proposed for inositol production from glucose or starch [15, 16]. However, the enzymatic production of inositol has suffered from a number of draw-backs, such as complex product separation process and instability of the catalytic enzymes, making this route economically uncompetitive. In vivo microbial biosynthesis of inositol is considered as the most promising alternative to the conventional phytate hydrolysis method owing to its low production cost and environmental-friendly production process. So far, Escherichia coli is the main host studied for microbial production of inositol though metabolic engineering [14, 17, 18]. By blocking or reducing the carbon fluxes toward glycolysis and pentose phosphate pathways, the inositol production was significantly increased [14, 18]. However, one of the main disadvantages of using E. coli strain for commercial inositol production is the safety concerns associated with the lack of the GRAS (Generally Recognized as Safe) status of this bacteria.
Pichia pastoris (recently classified as Komagataella phaffii), is a methylotrophic budding yeast and could grow rapidly on defined chemical medium with glucose or glycerol as substrate [19]. P. pastoris has been extensively used as an efficient platform for heterologous recombinant proteins production due to its GRAS status, rapid growth rate and ability for high-cell-density fermentation [20]. Recently, P. pastoris was also exploited as a robust host for value-added biochemicals production, including S-adenosyl-L-methionine (SAM) [21], malic acid [22], lycopene [23], etc. indicating this yeast strain is indeed a versatile microbial cell factory.
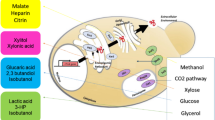
In this study, we aimed to engineer P. pastoris for efficient inositol production by dynamic regulation of carbon flux distribution (Fig. 1). First, an inositol-producing strain was constructed by enhancing the expression of endogenous IPS (encoding inositol-3-phosphate synthase, PAS_chr2-2_0113) gene which is the rate-limiting step for inositol biosynthesis and knocking out the PpITR1 (PAS_chr2-1_0489)/PpITR2 (PAS_chr4_0828) and pfk2 (PAS_chr1-4_0047) genes which are inositol transporters and β-subunit of 6-phosphofructokinase gene, respectively. Second, two heterologous enzymes IPS and IMP (inositol monophosphatase, PAS_chr4_0730) were overexpressed to further increase the inositol production. Third, the promoters of zwf (encoding glucose-6-phosphate dehydrogenase), pgi (encoding glucose-6-phosphate isomerase), and pfk1 (encoding α-subunit of 6-phosphofructokinase, PAS_chr2-1_0402) genes were replaced with the promoter of gut1 (encoding glycerol kinase, PAS_chr4_0783) gene. Consequently, the carbon flux distribution could be regulated in the presence of glucose via carbon catabolite repression (CCR). Last, the effect of oxygen availability on inositol production was investigated. The obtained mutant produced 30.71 g/L inositol in a high-cell-density fermentation in the 10 L bioreactor. Results from this study provide valuable guidance and promising strategies for engineering other yeast strains for inositol production.

Strategies for high-titer production of inositol in P. pastoris. Green arrows indicate the inositol biosynthesis pathway; orange arrows indicate glycerol catabolic pathway; genes in green indicate overexpression; genes in red indicate deletion or attenuation. G-3-P, glycerol-3-phosphate; DHAP, dihidroxyacetone-phosphate; G-6-P, glucose-6-phosphate; F-6-P, fructose-6-phosphate; F-1,6-BP, fructose-1,6-bisphosphate; I-3-P, inositol-3-phosphate; gut1, glycerol kinase; gut2, FAD-dependent glycerol-3-phosphate dehydrogenase; pgi, glucose-6-phosphate isomerase; pfk1, α-subunit of heterooctameric phosphofructokinase; pfk2, β-subunit of heterooctameric phosphofructokinase; IPS, inositol-3-phosphate synthase; IMP, inositol monophosphatase; zwf, glucose-6-phosphate dehydrogenase; PpITR1/PpITR2, inositol transporters
Results
Metabolic engineering of the native inositol biosynthesis pathway and transport system in P. pastoris
A markerless genome editing tool was initially developed based on mazF-zeoR counter-selection approach with some modifications [24]. Previous study demonstrated that the mazF-zeoR counter-selection approach is a highly efficient genome editing tool for P. pastoris. However, an unwanted scar (CYC1TT) was left in the genome after each round of gene editing (Additional file 1: Fig. S1) [24], which would limit the possible precise genomic modifications and increase the risk of homologous recombination between the scar sequences after multiple rounds of editing. In this study, the two CYC1TT sequences were removed, and a short-arm sequence (~ 100–300 bp) homologous to the 3’-end sequence of up-arm was inserted. Consequently, the up-arm and down-arm (~ 1000–1200 bp/each) which were much longer than short-arm would dominate the first round of recombination, while the short-arm would control the second round of recombination (Fig. 2A). Using the modified mazF-zeoR counter-selection approach, markerless genome editing could be achieved after two rounds of homologous recombination.

Markerless genome editing in P. pastoris using mazF-zeoR counter-selection approach. A Schematic illustrating the work flow of markerless genome editing system using mazF-zeoR counter-selection approach. Three homology arms with different lengths (up-arm and down-arm, ~ 1000–1200 bp; short-arm, ~ 100–300 bp) were used for gene deletion or integration. Two screening steps including two rounds of homologous recombination are involved. In step 1, the digested plasmid was transformed into P. pastoris under the selection of antibiotic zeocin, allowing the integration of “mazF-zeocin-short-arm” fragment onto the chromosome and the elimination of the target gene. In step 2, the obtained transformants were cultivated in BMMY medium containing methanol as the sole carbon source to induce the expression of mazF gene. The MazF selection stress promotes the removal of “mazF-zeocin” cassette through the second round of homologous recombination. B PCR analysis of JQ01 mutant after each selection step. C, D PCR analysis of JQ02 mutant after each selection step. M, 1-kb DNA ladder; WT, genomic DNA of P. pastoris GS115 as the control; Mut1, mutant strain after first round of recombination; Mut2, mutant strain after second round of recombination
The biosynthesis of inositol from the precursor glucose-6-phosphate (G-6-P) involves two crucial enzymes, IPS which can convert G-6-P to inositol-3-phosphate (I-3-P) and IMP which catalyzes the formation of inositol from I-3-P. Different from E. coli, P. pastoris has a native biosynthetic pathway for inositol production. However, the inositol biosynthesis in yeast is strictly regulated mainly by controlling the transcription of IPS gene [25]. Therefore, the promoter of IPS gene was replaced with the strong constitutive promoter pGAP (the promoter of glyceraldehyde-3-phosphate dehydrogenase gene) using the newly established mazF-zeoR counter-selection approach (Fig. 2B), creating mutant strain JQ01. It is reported that two transporters (ITR1 and ITR2) are responsible for extracellular inositol uptake in Saccharomyces cerevisiae [26]. By analyzing the genomic sequence of P. pastoris, two possible inositol transporters, PpITR1 and PpITR2, were identified to have strong similarity to ITR1 and ITR2 of S. cerevisiae, respectively. In order to decrease the potential inositol reassimilation and increase the inositol production, the two inositol transporter genes were deleted in strain JQ01 (Fig. 2C, D), generating JQ02. Fermentation results showed that ethanol was the major product for JQ01 and JQ02 (Fig. 3C). However, no detectable inositol production was observed in the fermentation broth of JQ01 and JQ02 (Fig. 3D), suggesting that some other factors might affect the production of inositol in P. pastoris.

Fed-batch fermentation profiles of P. pastoris GS115, JQ01, JQ02, JQ03, and JQ04. Strains were fermented in inorganic salt medium containing 20 g/L glycerol, thereafter 50 g/L glucose was added into the medium at the 24 h of fermentation for inositol production. A Glycerol consumption. B Glucose consumption. C Ethanol production. D Inositol production
Inositol production by slowing down the carbon flux of glycolytic pathway
The precursor for inositol biosynthesis is G-6-P, which is also an important intermediate for glycolysis pathway. Although the expression of IPS gene was enhanced, G-6-P might be still catabolized mainly in glycolysis pathway in P. pastoris, leading to negligible inositol production. In order to increase the G-6-P availability for inositol biosynthesis, the key gene—pgi—involved in catalyzing the conversion of G-6-P to fructose-6-phosphate (F-6-P) in glycolysis was initially selected to be deleted (Fig. 1). However, the attempt was unsuccessful using our mazF-zeoR counter-selection approach (data not shown), suggesting that the pgi gene plays a vital role in the central carbon metabolism of P. pastoris. Then, the pfk2 gene which is the β-subunit of heterooctameric phosphofructokinase in P. pastoris was chosen as another target gene to be deleted (Fig. 1). Fortunately, pfk2 gene was successfully deleted in strain JQ02, yielding mutant JQ03. Shake-flask fermentations were then carried out with wild-type strain and JQ03 to test their inositol production, using glycerol (initial carbon source for cell growth) and glucose (supplemented at 24 h of fermentation) as substrates. Results showed that the ethanol productions of wild-type strain and JQ03 mutant reached 10.61 and 11.54 g/L, respectively, indicating that ethanol was still the major product of these two strains (Fig. 3C). As expected, there was no detectable inositol production for wild-type strain. However, 0.71 g/L inositol was produced by mutant JQ03 at the end of fermentation (Fig. 3D). The ethanol production of mutant JQ03 was detected throughout the fermentation process, whereas the inositol accumulation was only observed after the addition of glucose (Fig. 3A–D). These results demonstrated that slowing down the carbon flux of the glycolytic pathway was an effective strategy to increase the G-6-P supply for inositol production.
Enhancement of inositol production by introduction of heterologous ScIPS and EcIMP
High inositol production was achieved in E. coli when plasmid was used for IPS and IMP overexpression [14, 18], which would usually give higher expression than single chromosome integration. Therefore, iterative integration of multiple-copy of IPS and IMP genes in P. pastoris was conducted in order to further enhance the inositol production. The ScIPS from S. cerevisiae and the EcIMP from E. coli which have been found to be efficient for inositol biosynthesis were selected and integrated into the PpITR1 locus of JQ03, generating mutant JQ04. Bidirectional promoter pHHX1 was employed for ScIPS and EcIMP overexpression [27]. Shake-flask fermentation showed that mutant JQ04 produced 0.80 g/L inositol, which was just slightly increased (12.7%) compared to that of strain JQ03 (Fig. 3D). In addition, the ethanol production of JQ04 reached 11.62 g/L, which was kept at the similar level to that of JQ03 (Fig. 3C). These results demonstrated that overexpression of heterologous ScIPS and EcIMP only had limited effect on inositol biosynthesis, implying that the supply of precursor G-6-P might be still not sufficient for inositol production in JQ04 and need to be further improved.
Further improvement of inositol production by dynamic regulation of glycolysis and pentose phosphate pathways
As demonstrated above, when glucose was used as carbon source for inositol production in P. pastoris, the carbon flux was mainly driven towards cell growth, primarily glycolysis and pentose phosphate pathways. In order to enhance the inositol production, these two pathways should be blocked or weakened. However, totally block the glycolysis and pentose phosphate pathways could lead to severe inhibition of cell growth [28]. Dynamic regulation is considered to be an effective strategy to modulate the balance between cell growth and product formation [29, 30]. Here, this strategy was applied to enhance the inositol production from glucose in P. pastoris. It is known that glycerol is an efficient carbon source for P. pastoris, but its utilization is repressed in presence of glucose. This repression is usually achieved by regulating the expression of gut1 gene which encodes glycerol kinase and mediates the first step of glycerol utilization in P. pastoris. In this study, we attempted to use the promoter of gut1 gene (pGUT1) to control the carbon flux of glycolysis and pentose phosphate pathways through carbon catabolite repression (CCR). Therefore, the promoters of zwf, pgi and pfk1 genes were replaced with the glycerol-inducible promoter pGUT1 in JQ04 (Fig. 1). The resultant strains were named as JQ05 (JQ04::pGUT1-zwf), JQ06 (JQ04::pGUT1-pgi), JQ07 (JQ04::pGUT1-zwf::pGUT1-pgi) and JQ08 (JQ07::pGUT1-pfk1). Consequently, the cell growth would not be disturbed when glycerol was used as carbon source, while the expressions of zwf, pgi and pfk1 genes could be inhibited through the addition of glucose, which could be beneficial for inositol production.
Mutants JQ05, JQ06, JQ07 and JQ08 were then cultured in inorganic salt medium with 20 g/L glucose or glycerol as sole carbon source to characterize their growth profiles. P. pastoris GS115 wild-type strain was used as the control. Results showed that all tested strains grew well in the glycerol medium, whereas prolonged lag-phase was observed for mutants JQ05, JQ06, JQ07 and JQ08 cultivated in glucose medium compared to wild-type strain (Fig. 4A, B), which were largely consistent with our expectations. A 12 h lag-phase was observed for strain JQ05, JQ06 and JQ07 (Fig. 4B). However, the lag-phase of mutant JQ08 was extended to 36 h, suggesting that the glycolysis and pentose phosphate pathways of JQ08 were strictly controlled.

Growth profiles and zwf, pgi, and pfk1 expressions of P. pastoris GS115, JQ05, JQ06, JQ07, and JQ08 grown in inorganic salt medium with glucose or glycerol as sole carbon source. A Growth profiles of various strains cultivated in glycerol medium. B Growth profiles of various strains cultivated in glucose medium. C qPCR results showing the mRNA expressions of zwf gene of various strains. D qPCR results showing the mRNA expressions of pgi gene of various strains. E qPCR results showing the mRNA expressions of pfk1 gene of various strains
In order to further verify the expressions of zwf, pgi and pfk1 genes in mutants JQ05, JQ06, JQ07 and JQ08, RT-qPCR was performed. The qPCR results showed that the expression of zwf gene in strains JQ05, JQ07 and JQ08 cultured in glucose medium were significantly decreased compared to those of JQ05, JQ07 and JQ08 cultured in glycerol medium (Fig. 4C), due to the glucose-mediated repression on pGUT1. In contrast, the expression of zwf gene in strain JQ06 possessing its native zwf promoter was kept at the similar level whether the carbon source in the medium was glucose or glycerol (Fig. 4C). As for pgi and pfk1 expressions, the native promoters of pgi (JQ05) and pfk1 (JQ05, JQ06 and JQ07) gave a higher transcription activity in glucose medium than those in glycerol medium (Fig. 4D, E). However, after the replacement of the native promoters of pgi and pfk1 with glycerol-inducible promoter pGUT1, the expressions of pgi (JQ06, JQ07 and JQ08) and pfk1 (JQ08) were greatly reduced in glucose medium compared to those in glycerol medium (Fig. 4D, E). These results are coincident with the results that the growth of mutants JQ05, JQ06, JQ07 and JQ08 were inhibited in the presence of glucose (Fig. 4B), demonstrating that the expressions of zwf, pgi and pfk1 genes in these mutants could be regulated by using glucose or glycerol as carbon source.
Shake-flask fermentations with mutants JQ05, JQ06, JQ07 and JQ08 were then performed using glycerol (for cell growth) and glucose (for inositol production) as substrates. Fermentation results showed that 1.00, 1.33 and 1.80 g/L inositol were produced by mutants JQ05, JQ06 and JQ07, respectively, which were increased by 25.0%, 66.3% and 125%, respectively, compared to that of mutant JQ04 (Fig. 5D). The ethanol productions of mutants JQ05, JQ06 and JQ07 were 13.35, 12.09 and 12.11 g/L, respectively, which were still much higher than inositol productions in these strains (Fig. 5C). The glycerol and glucose consumption profiles of JQ05, JQ06 and JQ07 were similar with those of JQ04 (Fig. 5A, B). In contrast, the carbon source consumption of mutant JQ08, especially the glucose consumption, was severely restrained (Fig. 5B), due to the block of glycolysis and pentose phosphate pathways. The ethanol production of JQ08 was reduced to 1.54 g/L (Fig. 5C). However, the inositol production was significantly enhanced in JQ08 (Fig. 5D). As high as 4.71 g/L inositol was generated in JQ08 at the end of fermentation, representing an increase of about 4.9-fold compared to that of mutant JQ04. Total 9.35 g/L glucose was utilized in the fermentation process of JQ08, indicating that the inositol yield of JQ08 reached 0.50 mol/mol glucose (Fig. 5E). However, the inositol yields of JQ05, JQ06 and JQ07 were just 0.02, 0.02 and 0.03 mol/mol glucose, respectively. These results indicated that dynamic regulation of glycolysis and pentose phosphate pathways was an effective strategy to enhance the inositol production in P. pastoris.

Fed-batch fermentation profiles of mutants JQ05, JQ06, JQ07, and JQ08. All mutants were fermented in inorganic salt medium containing 20 g/L glycerol. At the 24 h of fermentation, 50 g/L glucose was added into the medium for inositol production. A Glycerol consumption. B Glucose consumption. C Ethanol production. D Inositol production. E Stoichiometric yield of inositol
High-cell-density fermentation with strain JQ08
High inositol production and yield were obtained for strain JQ08 using glycerol and glucose as carbon sources. In order to comprehensively assess the fermentation capacity of JQ08, high-cell-density fermentation was carried out in a 10 L bioreactor. Glycerol was used as the initial carbon source for cell growth. When the wet cell weight (WCW) reached about 0.2 g/mL, glucose was then supplemented into the fermentation medium for inositol production. Initially, the level of dissolved oxygen (DO) was maintained at about 30% throughout the fermentation. The concentration of inositol reached 12.56 g/L at the end of fermentation (Fig. 6A). It is reported that oxygen limitation could strongly influence the core metabolism of P. pastoris by causing energy deprivation [31], and hypoxic condition has beneficial effect on recombinant protein secretion [32]. Therefore, lower DO levels were utilized to testify the effect of oxygen availability on inositol production in strain JQ08. Results showed that when DO levels were reduced to 15% and 5% after the addition of glucose, the inositol production of JQ08 reached 30.71 and 19.63 g/L (Fig. 6B, C), respectively, which were enhanced by 145% and 56.3%, respectively, compared to that of JQ08 fermented under the 30% DO condition. The results suggested that moderate control of oxygen availability has a positive effect on inositol production for mutant JQ08.

High-cell-density fermentation with mutant JQ08. The fermentations were performed in a 10 L bioreactor. Strain JQ08 was initial cultivated in the inorganic salt medium containing glycerol as the sole carbon source. When WCW reached about 0.2 g/mL, glucose was added into the medium for inositol production. Glucose was supplemented in batches. The DO was maintained at about 30% when glycerol was used as the carbon source for efficient cell growth. Then the level of DO was changed to 30% A, 15% B or 5% C after the addition of glucose for inositol biosynthesis
Discussion
In recent years, inositol and its derivatives have received considerable attention for their commercial applications in the fields of medicine, functional food, cosmetics and feed [33, 34]. Traditionally, the inositol was primarily produced by hydrolyzing plant-derived phytate, which would result in severe environmental pollution. Microbial biosynthesis of inositol has been regarded as one of the most promising alternatives to the phytate hydrolysis method. Numerous studies have been reported for metabolic engineering of different strains for inositol production [35]. To date, E. coli is the most efficient host for inositol biosynthesis, and high inositol production and yield were achieved in this strain [14, 18]. However, biochemicals produced by E. coli is restricted for medicine and food use purpose. In addition, to enhance the expression of genes involved in inositol biosynthesis, multi-plasmid expression approach was used in the most current reports [14, 18]. This on one hand brings extra burden to cellular metabolism; on the other, losing plasmid during the fermentation would lead to undulation in production. In this work, the methylotrophic yeast P. pastoris, an effective heterologous protein expression system, was exploited for efficient inositol production by dynamic regulation of carbon flux distribution without using plasmid expression system.
The main challenge for high-level inositol biosynthesis using microbial cell factory is the sufficient supply of G-6-P, which is the precursor for inositol biosynthesis and the essential intermediate for glycolysis and pentose phosphate pathways. Disruption or attenuation of glycolysis and pentose phosphate pathways could increase the yield of bioproducts derived directly from G-6-P. This strategy has been employed to glucaric acid and inositol production [35, 36]. In this study, we initially wanted to construct an inositol producing strain by manipulating the native inositol biosynthesis pathway and transport system in P. pastoris. However, the inositol accumulation was not observed in mutant JQ02 (Fig. 3D), suggesting that G-6-P was mainly consumed in glycolysis and pentose phosphate pathways. Totally block the glycolysis and pentose phosphate pathways would be an optimal option to enhance the supply of G-6-P, whereas the attempts consistently failed in P. pastoris. Mutant JQ03 with attenuated glycolysis pathway was successfully constructed by deletion of pfk2 gene, and inositol production was detected in the fermentation broth of JQ03 (Fig. 3D), which substantiated that slowing down the carbon flux of glycolytic pathway was beneficial for the production of inositol.
Appropriate modulation of the balance between cell growth and product formation is another challenge for inositol production from glucose. By using different carbon sources with direct access to growth and production modules, biochemical synthesis could be significantly improved [37, 38]. In this study, glycerol and glucose, which are both preferable carbon sources for P. pastoris, were selected and applied to support the cell growth and inositol production, respectively. In order to conserve more glucose for inositol formation, the promoters of key metabolic genes involved in glycolysis and pentose phosphate pathways, including zwf, pgi and pfk1, were replaced with the glycerol-inducible promoter pGUT1. Therefore, dynamic regulation of cell growth and inositol production was achieved via supplementing glycerol and glucose sequentially. The growth profile and RT-qPCR result of JQ08 substantiated that the expressions of zwf, pgi and pfk1 were successfully regulated by the addition of glucose (Fig. 4). In the previous study, gene knock-out or knock-down was employed to slow down the carbon flux of glycolysis and pentose phosphate pathways to enhance the supply of precursor G-6-P [18]. However, multiple knock-out/knock-down of key genes in glycolysis and pentose phosphate pathways severely inhibited the cell growth even when glycerol was used as the carbon source. In contrast, using the dynamic regulation strategy established in this study, the cell growth was totally not affected when glycerol was used as the carbon source (Fig. 4A). Using this strategy, high inositol production and yield were obtained in JQ08, and the accumulation of by-product ethanol was significantly reduced.
Environmental stresses, such as oxygen condition, have great influence on fermentation performance of microbial cell factory. It is reported that in fully aerobic conditions more glucose could be channeled to pentose phosphate pathway in P. pastoris, while increased glycolytic flux could be observed under the hypoxic conditions due to the upregulated expression of glycolytic genes like gap [31]. Therefore, it is reasonable to speculate that other genes controlled by the pGAP promoter would be upregulated in the hypoxic conditions as well. In our study, the promoter of native IPS gene which catalyzed the rate-limiting step for inositol biosynthesis was replaced with pGAP. Thus, the expression of native IPS gene could be enhanced under the low oxygen conditions. RT-qPCR was then performed to verify the expression of IPS in JQ08 under aerobic (DO: 30%) and hypoxic (DO: 15%) conditions. Results showed that the expression of IPS in JQ08 under hypoxic condition was significantly upregulated (about 3.1-fold increase) compared to that under the aerobic condition, which confirmed our speculation (Additional file 1: Fig. S3). This might be the reason that the inositol production of JQ08 was greatly increased under the low DO conditions in high-cell-density fermentation.
Conclusions
In conclusion, the GRAS yeast P. pastoris was successfully engineered as a high-inositol producer in this study by combining overexpression of native and heterologous genes. An effective dynamic regulation strategy was established and applied to enhance the inositol production from glucose. Using the dynamic regulation strategy, 30.71 g/L inositol was produced in an optimized fed-batch high-cell-density fermentation, which is the highest concentration achieved in yeast cell factory to date. The strategy developed herein could be readily adapted for the production of other high-value biochemicals.
Methods
Strains and culture media
All strains used in this study are listed in Table 1. E. coli strain Trans10 (Transgen Biotech, Beijing, China) was used for DNA cloning and plasmid propagation. P. pastoris GS115 was employed as the parental strain for genetic engineering and inositol production. E. coli Trans10 was routinely grown in Luria-Bertani (LB) medium supplemented with 100 µg/mL ampicillin as required. The recipes of media used for P. pastoris strains cultivation and selection were as follows: YPD medium: 20 g/L tryptone, 10 g/L yeast extract, 20 g/L glucose. BMGY medium: 20 g/L tryptone, 10 g/L yeast extract, 100 mM KH2PO4/K2HPO4 buffer (pH 6.0), 13.4 g/L YNB, 0.4 ppm biotin, 2% (v/v) glycerol. BMMY medium: 20 g/L tryptone, 10 g/L yeast extract, 100 mM KH2PO4/K2HPO4 buffer (pH 6.0), 13.4 g/L YNB, 0.4 ppm biotin, 1% (v/v) methanol. 100 µg/mL zeocin was added into the medium when needed.
Plasmid construction
All primers and plasmids used in this study can be found in Additional file 1: Table S1 and Tables 1, respectively. To construct the markerless editing plasmid pJQ (Additional file 1: Fig. S2A), the toxic mazF gene and antibiotic zeocin resistance gene were used as the selection markers. Firstly, the mazF gene from E. coli was amplified and cloned into the BamHI and AgeI sites of pPIC9K. Then, the pAOX-mazF expression cassette was amplified from the obtained plasmid, fused with ZeoR expression cassette (from pGAPZ A plasmid) by overlapping PCR and cloned into pEASY-T3 vector (Transgen Biotech, Beijing, China), creating plasmid pJQ. All the gene editing plasmids used in this study were constructed based on the mother vector pJQ. For the construction of markerless gene deletion plasmid (Additional file 1: Fig. S2B), three homology arms, up-arm (~ 1000–1200 bp), short-arm (~ 100–300 bp), down-arm (~ 1000–1200 bp), were amplified from the genomic DNA of P. pastoris GS115 and inserted into the SacII (up-arm) and SpeI (short-arm and down-arm) sites of pJQ vector, respectively. According to the genome editing demand, the sequence of short-arm was homologous to the 3’-end sequence of up-arm. For the construction of gene replacement or knock‑in plasmid, the cargo gene was inserted between the short-arm and down-arm of markerless gene deletion plasmid (Additional file 1: Fig. S2C). pGAP, pGUT1 and pHHX1 were amplified from the genomic DNA of P. pastoris GS115. ScIPS and EcIMP genes were obtained by PCR using the genomic DNA of S. cerevisiae and E. coli, respectively.
Mutant screening
The pJQ derivative plasmids for gene deletion, replacement or knock‑in were first linearized by NotI and then transformed into P. pastoris by electroporation according to the published method [39]. The transformants were selected on YPD or BMGY plates containing 100 mg/L zeocin. Colonies appeared on the plates were then screened by colony PCR to detect the deletion of target genes and the insertion of mazF-zeoR counter-selection marker on the chromosome of P. pastoris. The positive colonies were inoculated into BMMY liquid medium to induce the expression of mazF gene. The obtained cultures were diluted and plated on the YPD or BMGY plates. Colony PCRs were subsequently performed to confirm the eviction of mazF-zeoR counter-selection marker and the insertion of cargo genes or promoters.
Fed-batch fermentation
Shake-flask fermentations with various engineered strains were performed in 1 L shake flask with 200 mL reaction volume at 30 °C and 220 rpm. A modified inorganic salt medium (1.2 g/L KH2PO4, 0.5 g/L CaSO4, 6.5 g/L MgSO4, 5 g/L K2SO4, 18 g/L NH4H2PO4) containing 20 g/L glycerol was used as the fermentation medium. At 24 h of fermentation, 50 g/L glucose was supplemented into the medium for the inositol production. For the bioreactor fermentations, 10 L bioreactor (Shanghai Wanmuchun Biological Engineering Co., Shanghai, China) with a 7 L working volume was used. The initial glycerol concentration used in the inorganic salt medium was 50 g/L. After the glycerol was consumed, 50 g/L glucose was supplemented into the medium. Whenever the residual glucose concentration fell below 5 g/L, glucose solution (700 g/L) was pumped into the bioreactor to restore the glucose concentration to about 50 g/L. The temperature and pH were maintained at 30 °C and 5.0, respectively. The DO level was controlled at above 30% during 0–24 h (glycerol catabolism for cell growth) and then changed to 30%, 15% or 5% during the rest of fermentation period (glucose catabolism for inositol production) by automatic control of aeration rate and agitation speed. All P. pastoris strains were initially cultured in BMGY medium to prepare the seed culture (OD600 reached 10–12). Then, a 10% (v/v) inoculum of seed culture was used for all fed-batch fermentations. Shake-flask fermentations were performed in triplicate, while bioreactor fermentations were carried out in duplicate.
Analytical methods
Fermentation samples were collected every 6 or 12 h for the analysis. Glucose, glycerol, inositol, and ethanol in the fermentation broth were determined using high-performance liquid chromatography system (LC-20 A, Shimadzu, Kyoto, Japan) equipped with Agilent Hi-Plex Ca column (Agilent Technologies, Santa Clara, CA, USA) or Waters Sugar-Pak I column (Waters, Milford, MA, USA) and refractive index detector (RID). The mobile phase was ddH2O at a flow rate of 0.6 mL/min at 80 °C. Specifically, all the samples were filtered through the 0.22 μm filter before analysis. Cell density was monitored using a microplate reader at 600 nm (OD600). WCW was measured by centrifuging 1 mL of fermentation broth in a previously weighed 1.5 mL centrifuge tube at 12,000g for 2 min. The supernatant was discarded, and the difference in weight was defined as the WCW.
RT-qPCR analysis
The primers used in RT-qPCR are shown in Additional file 1: Table S1. P. pastoris strains were cultivated in inorganic salt medium with 20 g/L glucose or glycerol as carbon source. When OD600 reached about 10 (exponential growth phase), cells were harvested by centrifugation at 12,000g for 2 min. Total RNA extraction (RNA-easy Isolation Reagent, Vazyme, Beijing, China) and reverse transcription of cDNA (HiScript III RT SuperMix for qPCR, Vazyme, Beijing, China) were performed following the manufacturer’s instructions. All qPCRs were conducted according to the published procedure by using the ChamQ Universal SYBR qPCR Master Mix (Vazyme, Beijing, China) and the QuantStudio 6 Flex system (Applied Biosystems, Life Technologies, CA, USA) [40]. 2−∆∆Ct method was used to evaluate the transcription levels of the target genes [41], where the arg4 gene was used as the internal standard. All samples were run in triplicate.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its additional information file.
References
Michell RH. Inositol derivatives: evolution and functions. Nat Rev Mol Cell Biol. 2008;9:151–61.
Thomas MP, Mills SJ, Potter BVL. The “Other” inositols and their phosphates: synthesis, biology, and medicine (with recent advances in myo-inositol chemistry). Angew Chem Int Ed. 2016;55:1614–50.
Chen IW, Charalampous CF. Biochemical studies on inositol. IX. D-inositol 1-phosphate as intermediate in the biosynthesis of inositol from glucose 6-phosphate, and characteristics of two reactions in this biosynthesis. J Biol Chem. 1966;241:2194–9.
Croze ML, Soulage CO. Potential role and therapeutic interests of myo-inositol in metabolic diseases. Biochimie. 2013;95:1811–27.
Eden Evins A, Demopulos C, Yovel I, Culhane M, Ogutha J, Grandin LD, Nierenberg AA, Sachs GS. Inositol augmentation of lithium or valproate for bipolar depression. Bipolar Disord. 2006;8:168–74.
López-Gambero AJ, Sanjuan C, Serrano-Castro PJ, Suárez J, Rodríguez de Fonseca F. The biomedical uses of inositols: a nutraceutical approach to metabolic dysfunction in aging and neurodegenerative diseases. Biomedicines. 2020;8:295.
Shi D, Xia X, Cui A, Xiong Z, Yan Y, Luo J, Chen G, Zeng Y, Cai D, Hou L, et al. The precursor of PI(3,4,5)P3 alleviates aging by activating daf-18(Pten) and independent of daf-16. Nat Commun. 2020;11:4496.
Shiau S-Y, Su S-L. Juvenile tilapia (Oreochromis niloticus×Oreochromis aureus) requires dietary myo-inositol for maximal growth. Aquaculture. 2005;243:273–7.
Zehra S, Khan MA. Quantification of dietary inositol requirement for fingerling Channa punctatus (Bloch) based on growth, antioxidant status, hematological tools and liver inositol concentration. Aquaculture. 2019;512:734280.
Shiau S-Y, Su S-L. Dietary inositol requirement for juvenile grass shrimp, Penaeus monodon. Aquaculture. 2004;241:1–8.
Dai Z, Chung SK, Miao D, Lau KS, Chan AW, Kung AW. Sodium/myo-inositol cotransporter 1 and myo-inositol are essential for osteogenesis and bone formation. J Bone Miner Res. 2011;26:582–90.
Hayashi E, Maeda T, Tomita T. The effect of myo-inositol deficiency on lipid metabolism in rats: I. The alteration of lipid metabolism in myo-inositol deficient rats. Biochim Biophys Acta Lipids Lipid Metab. 1974;360:134–45.
Fujisawa T, Fujinaga S, Atomi H. An in vitro enzyme system for the production of myo-inositol from starch. Appl Environ Microbiol. 2017;83:e00550-17.
Tang E, Shen X, Wang J, Sun X, Yuan Q. Synergetic utilization of glucose and glycerol for efficient myo-inositol biosynthesis. Biotechnol Bioeng. 2020;117:1247–52.
You C, Shi T, Li Y, Han P, Zhou X, Zhang Y-HP. An in vitro synthetic biology platform for the industrial biomanufacturing of myo-inositol from starch. Biotechnol Bioeng. 2017;114:1855–64.
Lu Y, Wang L, Teng F, Zhang J, Hu M, Tao Y. Production of myo-inositol from glucose by a novel trienzymatic cascade of polyphosphate glucokinase, inositol 1-phosphate synthase and inositol monophosphatase. Enzyme Microb Technol. 2018;112:1–5.
Yi M, Yang L, Ma J, Liu H, He M, Hu C, Yu P. Biosynthesis of myo-inositol in Escherichia coli by engineering myo-inositol-1-phosphate pathway. Biochem Eng J. 2020;24:107792.
You R, Wang L, Shi C, Chen H, Zhang S, Hu M, Tao Y. Efficient production of myo-inositol in Escherichia coli through metabolic engineering. Microb Cell Fact. 2020;19:109.
Peña DA, Gasser B, Zanghellini J, Steiger MG, Mattanovich D. Metabolic engineering of Pichia pastoris. Metab Eng. 2018;50:2–15.
Karbalaei M, Rezaee SA, Farsiani H. Pichia pastoris: A highly successful expression system for optimal synthesis of heterologous proteins. J Cell Physiol. 2020;235:5867–81.
Qin X, Lu J, Zhang Y, Wu X, Qiao X, Wang Z, Chu J, Qian J. Engineering Pichia pastoris to improve S-adenosyl- l-methionine production using systems metabolic strategies. Biotechnol Bioeng. 2020;117:1436–45.
Guo F, Dai Z, Peng W, Zhang S, Zhou J, Ma J, Dong W, Xin F, Zhang W, Jiang M. Metabolic engineering of Pichia pastoris for malic acid production from methanol. Biotechnol Bioeng. 2021;118:357–71.
Zhang X, Wang D, Duan Y, Zheng X, Lin Y, Liang S. Production of lycopene by metabolically engineered Pichia pastoris. Biosci Biotechnol Biochem. 2020;84:463–70.
Yang J, Jiang W, Yang S. mazF as a counter-selectable marker for unmarked genetic modification of Pichia pastoris. FEMS Yeast Res. 2009;9:600–9.
Henry SA, Gaspar ML, Jesch SA. The response to inositol: Regulation of glycerolipid metabolism and stress response signaling in yeast. Chem Phys Lipids. 2014;180:23–43.
Nikawa J, Tsukagoshi Y, Yamashita S. Isolation and characterization of two distinct myo-inositol transporter genes of Saccharomyces cerevisiae. J Biol Chem. 1991;266:11184–91.
Vogl T, Kickenweiz T, Pitzer J, Sturmberger L, Weninger A, Biggs BW, Köhler E-M, Baumschlager A, Fischer JE, Hyden P, et al. Engineered bidirectional promoters enable rapid multi-gene co-expression optimization. Nat Commun. 2018;9:3589.
Guitart Font E, Sprenger GA. Opening a novel biosynthetic pathway to dihydroxyacetone and glycerol in Escherichia coli mutants through expression of a gene variant (fsaA(A129S)) for fructose 6-phosphate aldolase. Int J Mol Sci. 2020;21:9625.
Gupta A, Reizman IMB, Reisch CR, Prather KLJ. Dynamic regulation of metabolic flux in engineered bacteria using a pathway-independent quorum-sensing circuit. Nat Biotechnol. 2017;35:273–9.
Yang Y, Lin Y, Wang J, Wu Y, Zhang R, Cheng M, Shen X, Wang J, Chen Z, Li C, et al. Sensor-regulator and RNAi based bifunctional dynamic control network for engineered microbial synthesis. Nat Commun. 2018;9:3043.
Baumann K, Carnicer M, Dragosits M, Graf AB, Stadlmann J, Jouhten P, Maaheimo H, Gasser B, Albiol J, Mattanovich D, Ferrer P. A multi-level study of recombinant Pichia pastoris in different oxygen conditions. BMC Syst Biol. 2010;4:141.
Baumann K, Maurer M, Dragosits M, Cos O, Ferrer P, Mattanovich D. Hypoxic fed-batch cultivation of Pichia pastoris increases specific and volumetric productivity of recombinant proteins. Biotechnol Bioeng. 2008;100:177–83.
Vucenik I, Druzijanic A, Druzijanic N. Inositol hexaphosphate (IP6) and colon cancer: from concepts and first experiments to clinical application. Molecules. 2020;25:5931.
Özturan A, Arslan S, Kocaadam B, Elibol E, İmamoğlu İ, Karadağ MG. Effect of inositol and its derivatives on diabetes: a systematic review. Crit Rev Food Sci Nutr. 2019;59:1124–36.
Li Y, Han P, Wang J, Shi T, You C. Production of myo-inositol: Recent advance and prospective. Biotechnol Appl Biochem. 2021; https://doi.org/10.1002/bab.2181.
Shiue E, Brockman IM, Prather KLJ. Improving product yields on D-glucose in Escherichia coli via knockout of pgi and zwf and feeding of supplemental carbon sources. Biotechnol Bioeng. 2015;112:579–87.
Kim Y-S, Lee J-H, Kim N-H, Yeom S-J, Kim S-W, Oh D-K. Increase of lycopene production by supplementing auxiliary carbon sources in metabolically engineered Escherichia coli. Appl Microbiol Biotechnol. 2011;90:489–97.
Park JO, Liu N, Holinski KM, Emerson DF, Qiao K, Woolston BM, Xu J, Lazar Z, Islam MA, Vidoudez C, et al. Synergistic substrate cofeeding stimulates reductive metabolism. Nat Metab. 2019;1:643–51.
Pan R, Zhang J, Shen W-L, Tao Z-Q, Li S-P, Yan X. Sequential deletion of Pichia pastoris genes by a self-excisable cassette. FEMS Yeast Res. 2011;11:292–8.
Lu J, Struewing I, Wymer L, Tettenhorst DR, Shoemaker J, Allen J. Use of qPCR and RT-qPCR for monitoring variations of microcystin producers and as an early warning system to predict toxin production in an Ohio inland lake. Water Res. 2020;170:115262.
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2–∆∆CT method. Methods. 2001;25:402–8.
Funding
This research was supported by the National Key Research and Development Program of China (2021YFC2100203), the Agricultural Science and Technology Innovation Program (cxgc-ias-16), and the State Key Laboratory of Animal Nutrition Project (2004DA125184G2101).
Author information
Authors and Affiliations
Contributions
QZ and XW conducted the investigation and wrote the manuscript. HL, YW, YW, TT, XQ, XS, HH and BY conducted investigation. YB and JZ conceived and designed the research. All authors contributed to data analyses and results interpretation.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file
1: Table S1. Primers used in this study. Fig. S1. Comparation of genomeediting tool developed by Yang et al. [1] or used in ourstudy. A CYC1TT scar would leave in the genome after each round of gene editing in theprevious method. However, markerless genome editing could be achieved using ourmethod. Fig. S2. Maps of plasmids usedfor markerless genome editing in P. pastoris. A Plasmid pJQ was usedas the mother vector for the construction of markerless gene deletion orknock-in plasmids. B Map of plasmid used for markerless gene deletion. CMap of plasmid used for markerless gene knock-in. Fig.S3. qPCR results showing the mRNA expressions of IPSgene of JQ08 under aerobic (DO: 30%) and hypoxic (DO: 15%) conditions.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, Q., Wang, X., Luo, H. et al. Metabolic engineering of Pichia pastoris for myo-inositol production by dynamic regulation of central metabolism. Microb Cell Fact 21, 112 (2022). https://doi.org/10.1186/s12934-022-01837-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12934-022-01837-x