
Abstract
Background and aims
Caveolin-1 (CAV-1) in adipocyte tissue and other body parts possesses numerous biological functions. In the present study, we sought to investigate the interaction between CAV-1 polymorphism and dietary fat quality indexes on visceral adiposity index (VAI) and body adiposity index (BAI) among overweight and obese women.
Methods
This study was conducted on 386 women aged 18–48 years old. Biochemical measurements were assessed by standard protocols. We used a food frequency questionnaire (FFQ) to calculate the dietary intake and the indexes of dietary fat quality intake. Anthropometric values and body composition were measured by standard methods. Finally, the CAV-1 genotype was measured using the PCR–RFLP method.
Results
We found marginally significant differences between AA and GG genotypes of waist-to-hip ratio (WHR) (P = 0.06) and BAI (P = 0.06) of participants after adjusting for potential confounders. For dietary intakes, after adjusting with the energy intake, mean differences in biotin (P = 0.04) and total fiber (P = 0.06) were significant and marginally significant, respectively. The interaction between two risk alleles (AA) with omega-6 to omega-3 ratio (W6/W3) on BAI, after adjustment for potential confounders (age, physical activity, energy intake, education), was marginally positive (β = 14.08, 95% CI = − 18.65, 46.81, P = 0.07). In comparison to the reference group (GG), there was a positive interaction between the two risk alleles (AA) with W6/W3 ratio on VAI (β = 2.81, 95% CI = 1.20, 8.84, P = 0.06) in the adjusted model.
Conclusions
We found that there might be an interaction between CAV-1 genotypes with dietary quality fat indexes on VAI and BAI among overweight and obese women.
Similar content being viewed by others


Introduction
Overweight and obesity represent notable risk factors for many non-communicable diseases, and contribute significantly to global mortality rates [1,2,3]. Indeed, it has been estimated that by 2030, obesity and overweight will dramatically increase [4]. Obesity and overweight are associated with many diseases and clinical conditions, such as diabetes, cardiovascular disease (CVD), disability, depression, and early mortality [5].
Various anthropometric indexes can be used to get an overview of the body composition and health status of the body [6]. For example, body mass index (BMI), hip circumference (HC), waist circumference (WC), and WHR can be used to measure and monitor obesity [7]. In recent years, novel anthropometric indexes such as VAI and BAI have been introduced to measure body fat distribution accurately [8]. BAI appears to represent a good assessment of fat percentage using HC, height, and gender [9]. Whilst VAI is used to evaluate adipose tissue and its function based on gender by using anthropometric and biochemical parameters, such as waist circumference, body mass index, blood triglyceride concentration, and blood cholesterol [10].
Throughout the body, including adipose tissue, the plasma membrane contains a small (50–100 nm) complex, invaginations, and flask-shaped specialized structures, called caveolae, that have cholesterol and sphingolipids in their structure [11]. Caveolae are involved in lipid metabolism, signal transduction, and endocytosis [12,13,14,15]. Caveolae contain proteins called caveolin [16], which are integral membrane proteins that have scaffolding functions, and are regulators of many signaling pathways [17, 18]. There are three forms of caveolin: caveolin 1, caveolin 2, and caveolin 3, which are expressed differently and can all affect obesity [19]. Caveolin-1 is expressed in adipose tissue, caveolin-2 is expressed with caveolin-1 to form hetero-oligomers, and caveolin-3 is the muscle-specific isoform of caveolin [12, 15]. As an interactive gene, caveolin interacts with cholesterol and plays an essential role in its metabolism and regulation [20]. Due to the notable interaction of caveolin with cholesterol, animal studies have shown the significant effect of a high-cholesterol diet on the expression of the caveolin gene [21].
The quality of dietary fat intake is related to obesity and body fat percentage, and to examine this relationship, all aspects of fatty acids in the diet should be considered [22]. Indeed, most studies have been performed on the role of diet in the expression of the caveolin gene in animals. According to such studies, caveolin can be influenced and regulated by obesity and diet [23]. Fat consumption is known to impact adipose tissue [24]. Moreover, to evaluate the quality of dietary fat, several indicators can be used, including the cholesterol-saturated fat index (CSI) [25] and W6/W3 ratio [26].
This study sought to investigate the interaction between the quality of dietary fat intake and the CAV-1 gene and their relationship with visceral adiposity index and body adiposity index.
Methods and materials
Participants
Three hundred eighty-six women aged 18–48 years were randomly selected from among participants in a cross-sectional study performed in 2016–2017. The inclusion criteria were: obese or overweight (BMI > 25 kg/m2), no alcohol consumption, and no smoking. Women with CVD, kidney failure, stroke, thyroid disease, liver disease, cancer, inflammatory illnesses, and those taking any therapeutic medications and weight loss supplements, or any supplements that affect weight were excluded from the study. Pregnant and menopausal women and those who reported a total daily energy intake outside 800–4200 kcal (3344–17,556 kJ) were also excluded from the study. The study protocol was approved by the ethics committee of Tehran University of Medical Sciences (TUMS) with the following identification: IR.TUMS.VCR.REC.1397.920. All methods were carried out by relevant guidelines. All participants of the study completed a written informed consent.
Measurement of biochemical parameters
All blood samples were collected at the Nutrition and Biochemistry laboratory of the School of Nutritional Sciences and Dietetics, TUMS. Serum triglycerides (TG) concentrations were assayed with triacylglycerol kits (Pars Azmoon Inc, Tehran, Iran) by using enzymatic colorimetric tests with Glycerol-3-phosphate oxidase Phenol 4-Aminoantipyrine Peroxidase (GPO-PAP). Total cholesterol (total-chol) levels were measured by the cholesterol oxidase Phenol 4-Aminoantipyrine Peroxidase (CHOD-PAP), and low-density lipoprotein (LDL) and high-density lipoprotein (HDL) were measured by the direct method and immune inhibition.
Assessment of anthropometric measures
Weight was measured with digital scales and recorded to the nearest 100 g while the subjects were minimally clothed and unshod. Height was measured using tape while the subjects were standing, unshod, and had shoulders in a normal position. BMI was calculated by dividing the weight by the square of the height, waist circumference (WC) was measured at the narrowest part of the abdomen, HC was measured as the maximum circumference over the buttocks, and the minimal circumference was recorded to the nearest 0.1 cm. To reduce error, all measurements were taken by the same technician. For measuring the fat distribution, we used BAI and VAI (for women) with the following formulas:
-
BAI was calculated using the following formula [9]:
$${\text{BAI }} = \frac{{{\text{Hip }}\;{\text{Circumference}}\;\left( {{\text{cm}}} \right)}}{{height \left( m \right)^{1.5} }} - 18$$ -
VAI was calculated using the formula [10]:
$${\text{Women:}}\;{\text{VAI }} = \left( {\frac{{{\text{Waist}}\;{\text{Circumference}}\;{ }\left( {{\text{cm}}} \right)}}{{36.58 + \left( {1.89 \times BMI} \right)}}} \right) \times \left( {\frac{TG}{{0.81}}} \right) \times \left( {\frac{1.52}{{HDL}}} \right)$$
Assessment of dietary intake
To evaluate participants’ dietary intake, we used a 147-item semi-quantitative FFQ, with high validity and reliability [27], during face-to-face interviews. The extracted FFQ values were then changed to grams/day. For the evaluation of macro-and micronutrient content, N4 software was used.
Measurement of fat quality indexes
Fat quality indexes include CSI and W6/W3 ratio which are calculated through their respective formulas.
-
CSI: Indicates the state of cholesterol and saturated fats, which helps a person in self-care against the state of cholesterol [25].
$${\text{CSI}} = \frac{Cholesterol}{{Saturated\;fat}}$$ -
w–6/w–3 ratio: Omega 6/Omega 3 are two essential fats that are categorized as PUFAs. The total amount of omega-6 s and omega-3 s are divided by each other to get a ratio [26].
$$W{-}6/W{-}3\; Ratio = \frac{{\sum {\upomega }{-}6{ }}}{{\sum {\upomega }{-}3}}$$
Genotyping
For genotyping the Cav-1 polymorphisms, DNA was extracted from whole blood via a Mini Columns kit (Type G; Genall; Exgene). The polymerase chain reaction-restriction fragment length polymorphism (PCRRFLP) technique was employed to investigate Cav-1 polymorphisms (rs3807992) in gene fragments (major allele G and minor allele A). PCR was carried out using the following primers:
F:3′AGTATTGACCTGATTTGCCATG5′R:5′GTCTTCTGGAAAAAGCACATGA-3′, according to pervious study [28]. We gave risk alleles GG, AG, and AA the values 2, 1, and 0.
Assessment of other variables
We assessed the participants’ physical activity with a validated International Physical Activity Questionnaire (IPAQ) [29]. We also designed and used a standard socio-economic demographic questionnaire to collect general background information, including education and marital status.
Statistical analysis
Statistical analysis was performed using SPSS v.25 software (SPSS Inc., IL, USA) and the significance level was, a priori, considered P ≤ 0.05, while P = 0.06 was considered as marginally significant. There were no missing values in the analysis. Our data followed a normal distribution, which was confirmed using the Kolmogorov–Smirnov test. All continuous data were expressed as means and standard deviation (SD), and categorical variables were expressed as numbers and percentages. The Pearson’s chi-square test was used for categorical variables and to determine the Hardy–Weinberg Equilibrium. One-way analysis of variance (ANOVA) was used to evaluate the association between dietary intake indexes, anthropometric parameters, and biochemical parameters, and analysis of covariance (ANCOVA) was used to evaluate and control for confounders. BMI consider as collinear variable for anthropometrics and body composition variables, these variables control with energy intake, age, and physical activity. To investigate the mean differences between groups, post-hoc multiple comparison analysis (Bonferroni corrected), was used. To estimate interactions between CAV-1 genotypes and VAI and BAI, we used a generalized linear model, conducting a crude and adjusted model, where we controlled several potential confounders (age, energy intake, physical activity, education). GG allele carriers were consider as the reference group.
Result
Study population characteristics
A total of 386 overweight and obese women were evaluated in this study. The participant characteristics were: age 36.67 (9.10) years, BMI 31.26 (4.29) kg/m2, VAI 2.48 (2.13), BAI 29.58 (7.62), CSI 9.74 (3.43), and W6/W3 ratio 12.65 (0.10). Among the participants, 286 (70.8%) women were married, and 110 (27.2%) had a good economic status. The overall prevalence of CAV-1 genotypes in the study was 50% (193), 23.3% (90), and 25.5% (103) for AA, AG, and AA, respectively.
Baseline characteristics of study participants among the CAV-1 genotypes
The baseline characteristics of study participants, categorized according to the CAV-1 genotypes, are presented in Table 1. As shown in this table, P values for all variables were reported before the adjustment in the crude model, and after adjustment with potential confounders, including age, physical activity, energy intake, and BMI. BMI was considered as collinear for anthropometrics and body composition variables. In the crude model, there was a significant mean difference among the study participants in terms of BMI (P = 0.01), diastolic blood pressure (DBP) (P = 0.02), HDL (P = 0.001), LDL (P = 0.03), and VAI (P = 0.006). After adjustment for potential confounders, the mean difference became marginally significant in the WHR and BAI of participants. The mean difference of HDL and VAI remained significant in the adjusted model. According to Bonferroni post-hoc testing, the mean difference in WHR, VAI, and HDL was between AA and GG genotype groups, such that the mean was higher in AG genotype for WHR and VAI and was higher in GG genotype for HDL, while, in BAI, there was a difference between AA and AG genotypes, such that in the AG genotype was higher. In categorical variables, a significant mean difference among the participants was seen regarding their education status after controlling for cofounders. There was no significant difference in terms of other variables in Table 1.
Dietary intakes of all subjects according to CAV-1 genotypes in obese and overweight women
Dietary intakes of all subjects according to CAV-1 genotypes in obese and overweight women are presented in Table 2. Regarding fat components of foods, no significant difference was found.in crude and adjusted models. The result was the same for macronutrients and energy, with no significant difference in the crude and adjusted models. According to micronutrients, there was no significant difference among participants in the crude model. But after adjustment for energy intake, mean differences in Biotin (P = 0.04) and total fiber (P = 0.06) were significant and marginally significant, respectively. There was no significant difference in terms of other variables in Table 2.
The association of genotype variant of CAV-1 with BAI and VAI in crude and adjusted linear regression models in overweight and obese women
The association of the genotype variant of CAV-1 with BAI and VAI in the crude and adjusted linear regression model is presented in Table 3. After controlling for the potential confounders (age, energy intake, physical activity), in comparison to the reference group (GG), there was a marginally significant positive association between the AA genotype and BAI (β = 0.94, 95% CI = 0.006, 4.33, P = 0.06). A positive association between AG genotype and VAI, compared to the reference group (GG), was observed in the crude model (β = 67.77, 95% CI = 26.06, 109.48, P = 0.001). On the other hand, after adjusting for confounders (age, energy intake, physical activity), a positive association was found in comparison to the reference group (GG) for the AA and AG genotypes with VAI (β = 42.12, 95% CI = −2.74, 86.98, P = 0.06), (β = 84.12, 95% CI = 32.16, 136.07, P = 0.002), respectively.
GG consider as reference group
Interactions between CAV-1 genotypes and CSI on BAI and VAI in overweight and obese women
The interaction of CAV-1 genotypes with CSI on BAI and VAI in the crude and adjusted model is presented in Table 4. In the crude model, in terms of BAI, there was a negative interaction between two risk alleles (AG) with CSI (β = − 0.30, 95% CI = − 0.99, 0.37, P = 0.38) in comparison to the reference group (GG), and after controlling for potential confounders (age, physical activity, energy intake, education), this interaction remained negative between AG and CSI (β = −0.58, 95% CI = − 1.67, 0.50, P = 0.29). For VAI, a positive interaction was observed between two risk alleles (AA) with CSI in the crude and adjusted model, in comparison to reference group (GG) (β = 0.11, 95% CI = − 0.09, 0.31, P = 0.28), (β = 0.11, 95% CI = 0.11, 0.34, P = 0.04), respectively (Fig. 1).

Interaction between CAV-1 genotypes (GG as the reference group) and CSI intake on VAI. VAI (The P value for AA genotype: 0.61; P value for CSI intake: 0.99; P value for interaction between AA genotype and CSI intake: 0.28). In this figure low and high intake of CSI is based on medians, low intake is considered as ≤ 9.405 and high intake is considered as ≥ 9.405
Interactions between CAV-1 genotypes and W6/W3 ratio on BAI and VAI in overweight and obese women
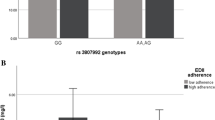
In the crude model, the interaction between two risk alleles (AG) with W6/W3 ratio on BAI, in comparison to the reference group (GG), was negative but not significant (β = − 6.45, 95% CI = −37.63, 24.73, P = 0.68), and the interaction between two risk alleles (AA) with W6/W3 ratio on BAI after adjustment for potential confounders (age, physical activity, energy intake, education) was marginally positive (β = 14.08, 95% CI = − 18.65, 46.81, P = 0.07) (Fig. 2). In terms of VAI, in comparison to the reference group (GG), there was a positive interaction between two risk alleles (AA) with W6/W3 ratio (β = 2.81, 95% CI = 1.20, 8.84, P = 0.06) in the adjusted model (Fig. 3).

Interaction between CAV-1 genotypes (GG as the reference group) and W6/W3 intake on BAI. VAI (The P value for AA genotype: 0.76; P value for W6/W3 intake: 0.52; P value for interaction between AA genotype and W6/W3 intake: 0.76). In this figure low and high intake of W6/W3 is based on medians, low intake is considered as ≤ 12.646 and high intake is considered as ≥ 12.646

Interaction between CAV-1 genotypes (GG as the reference group) and W6/W3 intake on VAI. VAI (The P value for AA genotype: 0.51; P value for W6/W3 intake: 0.80; P value for interaction between AA genotype and W6/W3 intake: 0.50). In this figure low and high intake of W6/W3 is based on medians, low intake is considered as ≤ 12.646 and high intake is considered as ≥ 12.646
Discussion
To our knowledge, this cross-sectional study is the first to investigate the interaction between CAV-1 genotypes and dietary fat quality indexes on VAI and BAI among overweight and obese women. Accordingly, our results showed that there might be an interaction between CAV-1 genotypes and dietary fat quality indexes (CSI, W6/W3 ratio) on BAI and VAI in overweight and obese women.
We found that the interaction of increased adherence to W6/W3 ratio consumption with AA carriers of CAV-1 was positive on VAI and BAI. We also noted a positive interaction between CSI with AA allele carriers on VAI. To our knowledge, no study has been conducted directly on this subject, but according to some related studies, CAV-1 may directly impact the regulation of HDL, TG, and cholesterol, and in general, CAV-1 affects fatty acid metabolism [20, 30, 31]. In another study, the CAV-1 expression was decreased in the adipocytes of obese subjects [32]. An animal study showed that consumption of a high-cholesterol diet could affect the expression of CAV-1 [21]. We found that AA allele carriers of CAV-1 have higher BAI than other groups, while in terms of VAI, the AG genotype group was higher than others. BAI and VAI represent novel indicators of obesity that have been scarcely studied [33, 34]. Females with AA allele carriers had a significantly higher BMI compared to the reference group (GG), which is consistent with previous studies [28]. According to Catalan et al., CAV-1 expression in visceral adipose tissue and subcutaneous adipose tissue is positively associated with BMI and body fat [30].
According to our results, females with the AG allele carriers have higher WHR and DBP values than the GG group, while HDL and LDL values were higher in the reference group (GG). As previously mentioned, CAV-1 has a critical role in lipid homeostasis, which can affect adipose tissue, blood vessels, and the liver to alter the regulation of TG, cholesterol, VLDL, and HDL [31]. However, our results did not show any significant difference in the macronutrients, energy, and fat components of the foods of subjects, with the only difference found in biotin and total fiber. The subjects' varied intake of foods high in biotin or containing biotin is likely the cause of this difference.
Caveolae are formed from lipid rafts and contain cholesterol, glycosphingolipids, and CAV-1 [35], and can elicit an uptake of lipid metabolites, including several fatty acid species, triacylglycerol, and cholesterol [36,37,38]. The roles of CAV-1 are variegated, with its most essential functions in cholesterol homeostasis, signal transduction, cellular and systemic lipid metabolism, and regulation of lipid and lipoprotein metabolism [30, 31, 39]. CAV-1 is expressed in different body parts, such as fibroblasts, epithelial and endothelial cells, and adipocytes [40]. Interestingly, the CAV-1 works as an element in lipid droplets and can affect lipid droplet accumulation and breakdown [41]. According to Chang et al., a high-fat diet causes CAV-1 to be secreted more in adipose tissue than a normal diet in mice; therefore, adipose tissue may be considered the main source of CAV-1 secretion [42]. Fat consumption can change the effect of Cav-1 polymorphism function on obesity and metabolic syndrome [28]. For example, high SFA intake can increase the adverse effects of Cav-1 in terms of metabolic syndrome and obesity [28]. One study revealed that A allele carriers with a high intake of PUFA, had a lower risk of metabolic syndrome. Consumption of SFA and PUFA can change caveolae contents and cell signaling [43,44,45].
CAV-1 is involved in lipogenesis and adipogenic processes as CAV-1 mRNA [46]., and CAV-1 can functionally suppress transforming growth factor-beta (TGF-β) by modifying the phosphorylation state of SMAD., Another potential mechanism is that the interaction between CAV-1 with transforming growth factor-beta type 1 receptor (TαR-1) may influence TGF-β inhibition [47]. Further, CAV-1 can impact cytokine signaling by interactions with the Janus kinase (JAK) family, cytokine receptors, and the proteasome pathway [15, 48].
Several limitations in the present study must be considered when interpreting our findings. This study was cross-sectional, thereby precluding causal inferences from being drawn. Next, the sample size used to conduct this study was small, and we used an FFQ to investigate the intakes of subjects, which, given its’ self-reported nature, may be influenced by recall bias. Furthermore, this study was conducted only on women, so the results may not generalize to all sexes. Nevertheless, despite the noted limitations, our study has numerous strengths, including; this is, to our knowledge, the first study to have investigated the interaction between CAV-1 genotypes with dietary fat quality indexes on VAI and BAI among overweight and obese women, moreover; we used detailed genetic factors in the study. Further, since we only conducted this study on women, greater specificity and insight may be gleaned.
Conclusion
Based on the present study’s finding, it appears that probably there may be an interaction between CAV-1 genotypes with dietary quality fat indexes and an association with VAI and BAI among overweight and obese women. Two risk alleles (AA) significantly interacted with CSI and W6/W3 ratio consumption and impacted BAI and VAI. In addition, we saw an association between the AG genotype and VAI. Nevertheless, we recommend that more studies be done in this area to confirm the integrity of our findings.
Availability of data and materials
The data that confirm the findings of this study are available from Khadijeh Mirzaei Data are available from the authors upon reasonable request and with permission of Khadijeh Mirzaei.
Abbreviations
- BAI:
-
Body adiposity index
- BMI:
-
Body mass index
- CAV-1:
-
Caveolin-1
- CI:
-
Confidence interval
- CSI:
-
Cholesterol-saturated fat index
- CVDs:
-
Cardiovascular diseases
- DBP:
-
Diastolic blood pressure
- DNA:
-
Deoxyribonucleic acid
- FFQ:
-
Food frequency questionnaire
- HC:
-
Hip circumference
- HDL:
-
High-density lipoprotein
- Il-1β:
-
Interleukin 1 beta
- IPAQ:
-
International physical activity questionnaire
- JAK:
-
Janus kinase
- LDL:
-
Low-density lipoprotein
- MET:
-
Metabolic equivalent of task
- mRNA:
-
Messenger ribonucleic
- PUFA:
-
Polyunsaturated fatty acid
- SD:
-
Standard deviation
- TG:
-
Triglyceride
- TGF-β:
-
Transforming growth factor beta
- TNF-α:
-
Tumor necrosis factor alpha
- TβR-1:
-
Transforming growth factor-beta type 1 receptor
- VAI:
-
Visceral adiposity index
- WC:
-
Waist circumference
- WHR:
-
Waist-to-hip ratio
- W6/W3:
-
Omega-6 to omega-3 ratio
References
Finucane MM, Stevens GA, Cowan M, Lin JK, Paciorek CJ, Singh GM, et al. HHS public access participants. Lancet. 2011;377(9765):557–67.
Ng M, Fleming T, Robinson M, Thomson B, Graetz N, Margono C, et al. Global, regional, and national prevalence of overweight and obesity in children and adults during 1980–2013: a systematic analysis for the Global Burden of Disease Study 2013. Lancet [Internet]. 2014;384(9945):766–81.
Kelishadi R. Childhood overweight, obesity, and the metabolic syndrome in developing countries. Epidemiol Rev. 2007;29(1):62–76.
Kelly T, Yang W, Chen CS, Reynolds K, He J. Global burden of obesity in 2005 and projections to 2030. Int J Obes. 2008;32(9):1431–7.
Hruby A, Hu FB. The epidemiology of obesity: a big picture. Pharmacoeconomics. 2015;33(7):673–89.
Al-Daghri NM, Al-Attas OS, Wani K, Alnaami AM, Sabico S, Al-Ajlan A, et al. Sensitivity of various adiposity indices in identifying cardiometabolic diseases in Arab adults. Cardiovasc Diabetol. 2015;14(1):1–8.
Yong L, GuangHui T, WeiWei T, LiPing L, XiaoSong Q. Can body mass index, waist circumference, waist-hip ratio and waist-height ratio predict the presence of multiple metabolic risk factors in Chinese subjects? BMC Public Health [Internet]. 2011;11(35):1–10. https://doi.org/10.1186/1471-2458-11-35.
Saqlain M, Akhtar Z, Karamat R, Iqbal M, Fiaz M, Zafar MM, et al. Body mass index versus other adiposity traits: best predictor of cardiometabolic risk. Iran J Public Health. 2019;48(12):2224–31.
Bergman RN, Stefanovski D, Buchanan TA, Sumner AE, Reynolds JC, Sebring NG, et al. A better index of body adiposity. NIH Public Access. 2012;19(5):1083–9. https://doi.org/10.1038/oby.2011.38.
Amato MC, Giordano C, Galia M, Criscimanna A, Vitabile S, Midiri M, et al. Visceral adiposity index: a reliable indicator of visceral fat function associated with cardiometabolic risk. Diabetes Care [Internet]. 2010;33(4):920–2. https://doi.org/10.2337/dc09-1825.
Bastiani M, Parton RG. Caveolae at a glance. J Cell Sci. 2010;123(22):3831–6.
Cohen AW, Hnasko R, Schubert W, Lisanti MP. Role of caveolae and caveolins in health and disease. Physiol Rev. 2004;84(4):1341–79.
Frühbeck G. The Sir David Cuthbertson Medal Lecture Hunting for new pieces to the complex puzzle of obesity. Proc Nutr Soc. 2006;65(04):329–47.
Parton RG, Simons K. The multiple faces of caveolae. Nat Rev Mol Cell Biol. 2007;8(3):185–94.
Frühbeck G, López M, Diéguez C. Role of caveolins in body weight and insulin resistance regulation. Trends Endocrinol Metab [Internet]. 2007;18(5):177–82.
Song KS, Tang Z, Li S, Lisanti MP. Mutational analysis of the properties of caveolin-1. A novel role for the C-terminal domain in mediating homo-typic caveolin–caveolin interactions. J Biol Chem [Internet]. 1997;272(7):4398–403. https://doi.org/10.1074/jbc.272.7.4398.
Inokuchi J. Insulin resistance as a membrane microdomain disorder. Biol Pharm Bull [Internet]. 2006;29(8):1532–7.
Parton RG, Hanzal-Bayer M, Hancock JF. Biogenesis of caveolae: a structural model for caveolin-induced domain formation. J Cell Sci. 2006;119(5):787–96.
Williams TM, Lisanti MP. The Caveolin genes: from cell biology to medicine. Ann Med. 2004;36(8):584–95.
Frank PG, Cheung MWC, Pavlides S, Llaverias G, Park DS, Lisanti MP. Caveolin-1 and regulation of cellular cholesterol homeostasis. Am J Physiol Hear Circ Physiol. 2006;291(2):H677–86. https://doi.org/10.1152/ajpheart.01092.2005.
Hahn-Obercyger M, Graeve L, Madar Z. A high-cholesterol diet increases the association between Caveolae and insulin receptors in rat liver. J Lipid Res [Internet]. 2009;50(1):98–107. https://doi.org/10.1194/jlr.M800441-JLR200.
Barbieiri P, Nunes JC, Torres AG, Nishimura RY, Zuccolotto DCC, Crivellenti LC, et al. Indices of dietary fat quality during midpregnancy is associated with gestational diabetes. Nutrition. 2016;32(6):656–61.
Lopez IP, Milagro FI, Marti A, Moreno-Aliaga MJ, Martinez JA, De Miguel C. High-fat feeding period affects gene expression in rat white adipose tissue. Mol Cell Biochem [Internet]. 2005;275(1–2):109–15. https://doi.org/10.1007/s11010-005-1082-z.
Gómez-Ruiz A, Milagro FI, Campión J, Martínez JA, De Miguel C. Caveolin expression and activation in retroperitoneal and subcutaneous adipocytes: Influence of a high-fat diet. J Cell Physiol. 2010;225(1):206–13.
Mitchell DT, Korslund MK, Brewer BK, Novascone MA. Development and validation of the cholesterol-saturated fat index (CSI) Scorecard. J Am Diet Assoc [Internet]. 1996;96(2):132–6.
Simopoulos AP. The importance of the ratio of omega-6/omega-3 essential fatty acids. Biomed Pharmacother. 2002;56(8):365–79.
Mirmiran P, Hosseini Esfahani F, Mehrabi Y, Hedayati M, Azizi F. Reliability and relative validity of an FFQ for nutrients in the Tehran Lipid and Glucose Study. Public Health Nutr. 2010;13(5):654–62.
Abaj F. Caveolin-1 genetic polymorphisms interact with fatty acid types to modulate metabolic syndrome. Br J Nutr. 2022;127(9):1281–8. https://doi.org/10.1017/S0007114521002221.
Aadahl M, Jørgensen T. Validation of a new self-report instrument for measuring physical activity. Med Sci Sports Exerc. 2003;35(7):1196–202.
Catalán V, Gómez-Ambrosi J, Rodríguez A, Silva C, Rotellar F, Gil MJ, et al. Expression of caveolin-1 in human adipose tissue is upregulated in obesity and obesity-associated type 2 diabetes mellitus and related to inflammation. Clin Endocrinol (Oxf). 2008;68(2):213–9.
Frank PG, Pavlides S, Cheung MWC, Daumer K, Lisanti MP. Role of caveolin-1 in the regulation of lipoprotein metabolism. Am J Physiol Cell Physiol. 2008;295(1):242–9.
Fernández-Real JM, Catalán V, Moreno-Navarrete JM, Gómez-Ambrosi J, Ortega FJ, Rodriguez-Hermosa JI, et al. Study of caveolin-1 gene expression in whole adipose tissue and its subfractions and during differentiation of human adipocytes. Nutr Metab. 2010;7:1–9.
Blus E, Wojciechowska-Kulik A, Majewska E, Baj Z. Usefulness of new indicators of obesity (BAI and VAI) in estimation of weight reduction. J Am Coll Nutr. 2020;39(2):171–7.
Jabłonowska-Lietz B, Wrzosek M, Włodarczyk M, Nowicka G. New indexes of body fat distribution, visceral adiposity index, body adiposity index, waist-to-height ratio, and metabolic disturbances in the obese. Kardiol Pol. 2017;75(11):1185–91.
Garg A, Agarwal AK. Caveolin-1: a new locus for human lipodystrophy. J Clin Endocrinol Metab. 2008;93(4):1183–5.
Su X, Abumrad NA. Cellular fatty acid uptake: a pathway under construction. Trends Endocrinol Metab [Internet]. 2009;20(2):72–7.
Pilch PF, Liu L. Fat caves: Caveolae, lipid trafficking and lipid metabolism in adipocytes. Trends Endocrinol Metab. 2011;22(8):318–24.
Grayson TH, Chadha PS, Bertrand PP, Chen H, Morris MJ, Senadheera S, et al. Increased caveolae density and caveolin-1 expression accompany impaired NO-mediated vasorelaxation in diet-induced obesity. Histochem Cell Biol. 2013;139(2):309–21.
Otis JP, Shen M-C, Quinlivan V, Anderson JL, Farber SA. Intestinal epithelial cell caveolin 1 regulates fatty acid and lipoprotein cholesterol plasma levels. Dis Model Mech [Internet]. 2017;10(3):283–95.
Gámez-Ruiz A, Milagro FI, Campián J, Martínez JA, De Miguel C. High-fat diet feeding alters metabolic response to fasting/non fasting conditions. Effect on caveolin expression and insulin signalling. Lipids Health Dis. 2011;10:1–10.
Cohen AW, Razani B, Schubert W, Williams TM, Wang XB, Iyengar P, et al. Role of caveolin-1 in the modulation of lipolysis and lipid droplet formation. Diabetes [Internet]. 2004;53(5):1261–70. https://doi.org/10.2337/diabetes.53.5.1261.
Chang C-C, Chen C-Y, Wen H-C, Huang C-Y, Hung M-S, Lu H-C, et al. Caveolin-1 secreted from adipose tissues and adipocytes functions as an adipogenesis enhancer. Obesity [Internet]. 2017;25(11):1932–40. https://doi.org/10.1002/oby.21970.
Perona JS. PT NU SC. BBA—biomembr [Internet]. 2017. https://doi.org/10.1016/j.bbamem.2017.04.015.
Chapkin RS, Mcmurray DN, Davidson LA, Patil BS, Fan Y, Lupton JR. Review article bioactive dietary long-chain fatty acids: emerging mechanisms of action. Br J Nutr. 2008;100(6)1152–7. https://doi.org/10.1017/S0007114508992576.
Lay S Le, Li Q, Proschogo N, Rodriguez M, Gunaratnam K, Cartland S, et al. Caveolin-1-dependent and -independent membrane domains. J Lipid Res. 2009;50(8):1609–20. https://doi.org/10.1194/jlr.M800601-JLR200.
Scherer PE, Lisanti MP, Baldini G, Sargiacomo M, Mastick CC, Lodish HF. Induction of caveolin during adipogenesis and association of GLUT4 with caveolin-rich vesicles. J Cell Biol. 1994;127(5):1233–43.
Razani B, Zhang XL, Bitzer M, Von Gersdorff G, Böttinger EP, Lisanti MP. Caveolin-1 regulates transforming growth factor (TGF)-β/SMAD signaling through an interaction with the TGF-β type I receptor. J Biol Chem. 2001;276(9):6727–38.
Felley-Bosco E, Bender FC, Courjault-Gautier F, Bron C, Quest AFG. Caveolin-1 down-regulates inducible nitric oxide synthase via the proteasome pathway in human colon carcinoma cells. Proc Natl Acad Sci USA. 2000;97(26):14334–9.
Acknowledgements
The authors thank the study participants for their assistance in physical examinations. They also thank those involved in nutritional assessment and database management. This manuscript has been granted by Tehran University of Medical Sciences (Grant No: 97-03-161-41017).
Funding
This manuscript has been granted by Tehran University of Medical Sciences (Grant No: 97-03-161-41017). The funder had no role in the design, analysis or writing of this article.
Author information
Authors and Affiliations
Contributions
The project was designed and wrote by RGE and KhM. Data were analyzed by FS. Data were collected by AM, and CC proofread the manuscript for native English writing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Ethics approval and consent to participate Ethics approval for the study protocol was confirmed by The Human Ethics Committee of Tehran University of Medical Sciences (TUMS) with the following identification: IR.TUMS.VCR.REC.1397.920. All participants signed a written informed consent that was approved by the Ethics committee. All methods were carried out in accordance with relevant guidelines and regulations.
Consent for publication
Not Applicable.
Competing interests
The authors express that they have no competing interests.
Sources of Support
Tehran University of Medical Sciences.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Ghaffarian-Ensaf, R., Shiraseb, F., Mirzababaei, A. et al. Interaction between caveolin-1 polymorphism and dietary fat quality indexes on visceral adiposity index (VAI) and body adiposity index (BAI) among overweight and obese women: a cross-sectional study. BMC Med Genomics 15, 258 (2022). https://doi.org/10.1186/s12920-022-01415-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12920-022-01415-5