
Abstract
Background
Since 2011, numerous highly virulent and antigenic variant viral strains have been reported in pigs that were vaccinated against the swine pseudorabies virus. These infections have led to substantial economic losses in the Chinese swine industry.
Results
This study, constructed a novel recombinant vaccine strain with gI/gE deletion (PRV-GD2013-ΔgI/gE) by overlapping PCR and homologous recombination technology. The growth curves and plaque morphology of the recombinant virus were similar to those of the parental strain. However, PRV-GD2013-ΔgI/gE infection was significantly attenuated in mice compared with that of PRV-GD2013. Two-week-old piglets had normal rectal temperatures and displayed no clinical symptoms after being inoculated with 105 TCID50 PRV-GD2013-ΔgI/gE, indicating that the recombinant virus was avirulent in piglets. Piglets were immunized with different doses of PRV-GD2013-ΔgI/gE, or a single dose of Bartha-K61 or DMEM, and infected with PRV-GD2013 at 14 days post-vaccination. Piglets given high doses of PRV-GD2013-ΔgI/gE showed no obvious clinical symptoms, and their antibody levels were higher than those of other groups, indicating that the piglets were completely protected from PRV-GD2013.
Conclusions
The PRV-GD2013-ΔgI/gE vaccine strain could be effective for immunizing Chinese swine herds against the pseudorabies virus (PRV) strain.
Similar content being viewed by others

Highlights
-
1.
Bartha-K61 did not provide full protection against the virulent PRV strains in China.
-
2.
The PRV mutant was generated by homologous recombination.
-
3.
The PRV mutant was non-toxic and immunogenic for immunized piglets.
-
4.
The PRV mutant provided effective protection against the PRV variant challenge.
Background
Pseudorabies (PR), widely known as Aujeszky′s disease, is an infectious disease for most livestock and wild animals caused by the pseudorabies virus (PRV). Pigs are a natural host while a variety of other vertebrates such as cattle, sheep, dogs, cats, goats, and rabbits, can be infected [1,2,3,4]. PRV infection in pigs produces different clinical symptoms, namely neurological symptoms and high mortality in piglets, growth retardation and respiratory disorders in growing pigs, and reproductive failure such as miscarriage and stillbirth in pregnant sows [5].
PRV is a double-stranded linear DNA virus of around 145 kb that belongs to the Alphaherpesvirinae subfamily of the Herpesviridae family [6]. The genome of PRV can be divided into a unique short region (US, 9 kb), a unique long region (UL, 110 kb), a terminal repeat sequence (TRS), and an internal repeat sequence (IRS) [7]. PRV encodes 10 glycoproteins, which are classified as essential and non-essential based on whether they are required for viral replication [6, 7]. The gE glycoprotein is encoded by the US8 gene. Its 125th valine and 126th cysteine have an important influence on the biological function of the virus. Deletion of these two amino acids reduces the neurotoxicity of the virus without affecting its immunogenicity [8]. The gI and gE glycoproteins together form a heterodimer complex, which is present in the infected cell membrane and the viral envelope membrane, and participates in the invasion of the nervous system by the pseudorabies virus and its transmission process [9,10,11]. The original vaccine strain, Bartha-K61, contains a genetic deletion of the gI, gE, US9, and US2 genes and is used worldwide, resulting in effective control of PR [12,13,14]. Between the 1990s and 2010, only a few swine PR cases were reported [15]. However, since 2011, PR outbreaks have occurred in pigs that were immunized with Bartha-K61, leading to severe economic losses in China [14, 16,17,18,19]. Consequently, several gene mutant vaccines of gE, gI&gE, gE&TK, gI&gE&TK deletion from PR variant have been generated [20,21,22,23]. Furthermore, studies have found that the effective reproduction rate (which represents the average number of secondary infections resulting from a typically infected swine at a given time within a population in which not all hosts are necessarily susceptible) during each year for the variant PRV was higher than one during 2012 to 2017 in China. This indicates a high risk of a variant PRV epidemic and that the safety or immunogenicity of these mutant vaccines is far from being satisfactory [24].
The overlapping PCR technique was developed by Horton et al. in 1989 [25]. This technology utilizes PCR to carry out effective gene recombination and site-directed mutagenesis in vitro. It can rapidly link two or more DNA fragments together without affecting the non-target DNA sequences. This technology facilitates the correct assembly of multiple fragments and accelerates complex genetic engineering, such as the production of polyclonal antibodies, somatic cell knockout of human cells, vaccine research, and assembly of viral genomes [26].
Our preliminary studies showed that the PRV variant isolated from infected swine that were immunized with Bartha-K61 had high pathogenicity and mortality. More than 200 pig farms were infected with this virus in China, which caused direct economic losses of more than 200 million RMB during outbreaks. Biological characterization and sequence analysis confirmed that the isolated strain, hereafter referred to as PRV-GD2013, had many specific amino acid mutations when compared with different variants found in our previous study in China. Bartha-K61 was unable to provide sufficient protection against PRV-GD2013. Unfortunately, there is no effective vaccine on the domestic market that can control this PRV mutant. Therefore, there is an urgent need to develop a new vaccine that can control the PRV epidemic in pig farms. In this study, a gI/gE-deleted mutant was constructed by overlapping PCR and homologous recombination technology from a PRV strain PRV-GD2013, and the safety and immunogenicity of the PRV mutant were evaluated in piglets.
Results
Characterization of the gI/gE-deleted PRV mutant
The pBLE (pBluescript SK (+)) and pBLE-EGFP (enhanced green fluorescent protein) vectors were used to construct pBLE-gI-gE and pBLE-gI-EGFP-gE vectors by overlapping PCR, and a recombinant PRV mutant was obtained in which gI/gE was deleted by homologous recombination (Fig. 1). After being subjected to five rounds of plaque purification under fluorescence microscopy, a gI/gE-deleted PRV mutant (PRV-GD2013-ΔgI/gE-EGFP) containing an EGFP gene was generated. Using the same method, a gI/gE-deleted PRV mutant (PRV-GD2013-ΔgI/gE) without an EGFP gene was produced. Lastly, the screening results were determined by fluorescence microscopy (Fig. 2). All the PRV-GD2013-ΔgI/gE infected cell without EGFP fluorescence compared to PRV-GD2013-ΔgI/gE-EGFP, which indicated that we successfully obtained the recombinant PRV-GD2013-ΔgI/gE (Fig. 2a, b). Recombinant viruses were analyzed by PCR (Fig. S1). The PCR results showed that the gene band size of line 1 was small when compared with lines 2 and 3, which indicated that we successfully deleted the gI/gE of PRV-GD2013, and, the DNA sequencing further verified this result (data not shown).

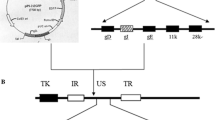
Construction strategy of the PRV-GD2013-ΔgI/gE recombinant strain. a, b Position of the left and right homologous recombination arms (L-arm and R-arm). c, d The genome of PRV-GD2013 and the relative site of US6 (gD), US7 (gI), US8 (gE), US9, and US2. e Construction of the transfer plasmids pBLE-gI-gE, including the target deletion region, L-arm, and R-arm. f Construction of the transfer plasmids pBLE-gI-EGFP-gE, including the target deletion region, L-arm, R-arm, and inserted EGFP expression cassette. g The genome of PRV-GD2013-ΔgI/gE-EGFP and the relative site of US6, US7s, the EGFP expression cassette, US8s, US9, and US2. h The genome of PRV-GD2013-ΔgI/gE and the relative site of US6, US7s, US8s, US9, and US2. UL, unique long region; US, unique short region; IR, internal repeat sequences; TR, terminal repeat sequences; US7s, section US7; US8s, section US8. EGFP expression cassette; EGFP (enhanced green fluorescent protein)

BHK-21 cells were infected with recombinant PRV. CPEs caused by PRV-GD2013-ΔgI/gE (a), PRV-GD2013-ΔgI/gE-EGFP (b), PRV-GD2013 (c), and control (d). CPEs were observed using a fluorescent microscope at two dpi
Growth characteristics of the gI/gE-deleted recombinant virus
A one-step growth curve showed that there was no significant difference in the replication of PRV-GD2013-ΔgI/gE and the parental PRV-GD2013 strain in BHK-21 cells (p < 0.05) (Fig. 3a). After 24 h of infection, the peak titer of the recombinant virus was 108.0–108.5 TCID50/ml, which was the same as the parental strain. Meanwhile, the plaque areas between PRV-GD2013-ΔgI/gE reconstituted viruses and PRV-GD2013 were not significantly different (Fig. 3b).

a One-step growth curve of PRV-GD2013-ΔgI/gE compared with its parental viruses. Monolayers of BHK-21 cells were inoculated with PRV-GD2013-ΔgI/gE and PRV-GD2013 at 1 MOI. The cell culture supernatants were collected at different time points (0, 4, 8, 12, 16, 20, 24, 28, 32, and 36 hpi), and were used to calculate the TCID50 of each virus. b Plaque morphology and plaque size measurement of PRV-GD2013-ΔgI/gE and PRV-GD2013 on BHK-21 cells at 60 h post-infection. The one-step growth curve and plaque size were measured by one-way repeated measurement analysis variance and least significance (LSD). Differences were considered statistically significant when p < 0.05
Virulence of the gI/gE-deleted recombinant virus in mice
Sixty mice were randomly divided into 12 groups of five and injected subcutaneously with different doses of PRV-GD2013 and PRV-GD2013-ΔgI/gE. The lethal doses (LD50) were 102.67 TCID50 (PRV-GD2013) and 104.33 TCID50 (PRV-GD2013-ΔgI/gE). The virulence of PRV-GD2013 in mice was higher than that of PRV-GD2013-ΔgI/gE, causing earlier death at the same dose (Table 1). The clinical symptoms of infected mice included pruritus at the injection site, biting of the skin, and hair loss.
Safety of the gI/gE-deleted recombinant virus in piglets
To evaluate the safety of gI/gE-deleted recombinant virus in piglets, fifteen 2-week-old piglets were randomly divided into 3 groups of five and inoculated with PRV-GD2013 (group A), 105 TCID50 PRV-GD2013-ΔgI/gE (group B) and DMEM (group C) intranasally (i.n.). During the experiment, rectal temperatures, virus shedding, clinical symptoms and the level of gB/gE-specific antibodies were monitored and recorded. Piglets in experimental group A displayed a fever above 41 °C on the 2nd day after the infection, accompanied by mental depression, anorexia, respiratory distress, and neurological symptoms. Following inoculation, virus shedding was detected in all piglets and the first piglet died after 3 days, three died after 4 days, and the remaining piglet died after 5 days (Table 2). All piglets in groups B and C exhibited normal rectal temperatures, no challenge virus shedding was detected (Table 2) and all survived without showing any clinical symptoms during the experiment, which was a stark contrast to group A (Fig. 4a). No gB-specific antibodies were detected in all groups (Fig. 4b). However, the gE-specific antibodies were detected in group B, not in groups A and C (Fig. 4c). At 15 day post-inoculation (dpi) all surviving piglets in groups B and C were euthanized and necropsied. Necropsies of all of the piglets in group A showed obvious pathological lesions in the brain, lymph nodes, lung, kidney, liver, and spleen. Severe cerebral hemorrhage was the most obvious feature. However, the tissues and organs of piglets in groups B and C were normal and exhibited no obvious pathological changes when compared with the reactions of group A (Fig. 5a). These data indicate that the gI/gE-deleted recombinant virus was attenuated in piglets.

Pathogenic testing in piglets using recombinant PRVs. a Rectal temperature of piglets after inoculation with PRV-GD2013, PRV-GD2013-ΔgI/gE, or DMEM. b gB-specific antibody levels. Sample with S/N ratios ≤0.60, were classified as positive for gB antibodies. c Detection of gE-specific antibody levels. Samples with S/N ratios ≤0.60 were classified as positive for gE antibodies. All data are presented as the mean ± SD

a Pathological examination of organ tissues. Groups of piglets (n = 3) were inoculated with 105 TCID50 PRV-GD2013, 105 TCID50 PRV-GD2013-ΔgI/gE, or DMEM. At 17 dpi, all surviving piglets were euthanized and necropsied. Tissue samples from the brain, lymph nodes, lung, kidney, liver, spleen were collected and used for pathological examination. b Pathological changes in various organ tissues of immunized piglets that were challenged with PRV-GD2013. Groups of piglets (n = 5) were inoculated with 105 TCID50 PRV-GD2013-ΔgI/gE, 104 TCID50 PRV-GD2013-ΔgI/gE, 103 TCID50 PRV-GD2013-ΔgI/gE, Bartha-K61, or DMEM. At 14 dpc, all surviving piglets were euthanized and necropsied. Tissue samples from the brain, lymph nodes, lung, kidney, liver, spleen were collected and used for pathological examination
Protective efficacy of gI/gE-deleted recombinant virus in piglets
Twenty-five piglets were randomly divided into five groups of five and injected with 105 PRV-GD2013-ΔgI/gE (group A), 104 PRV-GD2013-ΔgI/gE (group B), 103 PRV-GD2013-ΔgI/gE (group C), Bartha-K61 (group D), or DMEM (group E). Piglets in all groups exhibited normal rectal temperatures and no adverse clinical symptoms before the challenge. After 2 weeks of immunization, all of the piglets were infected with PRV-GD2013. All piglets in groups A and B survived with just a transient fever (40 ~ 41.5 °C), with no other clinical symptoms and virus shedding (Table 2). However, Piglets in groups C, D, and E were not completely protected when compared to groups A and B. In group C, piglets had only one occasion of elevated rectal temperatures (40.4 ~ 41.5 °C). One piglet showed virus shedding from the 4th to the 8th day after infection (Table 2) and displayed mild clinical symptoms including mental depression, anorexia, and respiratory distress. Piglets in group D had elevated rectal temperatures (40.4 ~ 41.5 °C) on two separate occasions. Four piglets exhibited virus shedding from the 3th to the 9th day after infection (Table 2), alongside displaying severe clinical symptoms including mental depression, anorexia, and respiratory distress, two of which showed obvious sequela (growth retardation and intermittent convulsion). All piglets in group E showed virus shedding (Table 2) and exhibited rectal temperatures over 41 °C twice post-challenge, which was accompanied by mental depression, anorexia, respiratory distress, and neurological symptoms. Three piglets in group E died, on the 4th, 5th, and 7th day respectively post-challenge (dpc).
Serum samples taken from all piglets were analyzed for gB and gE-specific antibodies at 0, 3, 7, 10, 14 (0), 17 (3), 21 (7), 24 (10), and 28 (14) days post-immunization. The numbers in parentheses represent the number of days after the infection was injected. gB-specific antibodies in all of the immunized piglets in groups A, B, C, and D, were detected at 7, 7, 10, and 10 dpi respectively. Levels of gB-specific antibodies showed a slight short-term decrease at three dpc, and then gradually increased. No gE-specific antibodies were detected in any piglets before the challenge. Production of gE-specific antibodies was detected in groups A, B, and C at 10 dpc, after which, gE-specific antibody levels gradually increased. At 14 dpc, gE-specific antibodies were detected in group D piglets. However, no gB and gE-specific antibodies were detected in group E piglets at any point during the experiment when compared to groups A-D (Fig. 6b, c).

Immunization and challenge experiments in piglets. a Rectal temperature of piglets after challenge with PRV-GD2013. b gB-specific antibody levels. Samples with S/N ratios ≤0.60 were classified as positive for gB antibodies. c gE-specific antibody levels. Samples with S/N ratios ≤0.60 were classified as positive for gE antibodies. All data are presented as the mean ± SD
At 15 dpc, all surviving piglets were euthanized and then necropsied. The tissues and organs of piglets in groups A and B were normal and no obvious pathological changes were observed. The brains and livers of piglets in groups C and D showed slight hemorrhage and lung consolidation compared to groups A and B. However, the pathological features of the piglets in group C were milder than those of group D. All piglets in group E exhibited pathological changes in the brain, lymph nodes, lung, kidney, liver, spleen. Severe cerebral hemorrhage was the most obvious feature when compared to other groups (Fig. 5b).
Piglets immunized with 105 TCID50 PRV-GD2013-ΔgI/gE and 104 TCID50 PRV-GD2013-ΔgI/gE showed almost no obvious pathological lesions after challenged by PRV-GD2013. However, piglets immunized with 103 TCID50 PRV-GD2013-ΔgI/gE, Bartha-K61, and DMEM showed different degrees of pathological lesions when compared to 105 TCID50 PRV-GD2013-ΔgI/gE and 104 TCID50 PRV-GD2013-ΔgI/gE group. Piglets immunized with 103 TCID50 PRV-GD2013-ΔgI/gE exhibited some inflammatory cell infiltration in the brain, as well as alveolar wall thickening, hepatic portal area inflammatory cell infiltration, loose lymphocyte arrangement, and necrotic and disintegrated partial lymphocytes. The piglets in Bartha-K61 group also showed inflammatory cell infiltration in the brain, lung consolidation, inflammatory cell infiltration, lung abscesses, hemorrhage, and alveolar effusion, liver occasional punctate necrosis and hepatocyte cavitation. The piglets in the DMEM group all had obvious inflammatory cell infiltration in the brain, inflammatory cell infiltration, lung abscesses, hemorrhage, alveolar effusion, liver occasional punctate necrosis and hepatic portal area inflammatory cell infiltration, spleen hemorrhage, renal tubular epithelial cell degeneration, necrosis, loose lymphocyte arrangement, necrotic and disintegrated partial lymphocytes. The pathological lesions in the DMEM group were significantly more severe than those in Bartha-K61 group (Fig. 7).

Histological examination of brain (A1–A5), lung (B2-B5), liver (C2–C5), spleen (D2–D5), kidney (E2–E5), and lymph nodes (F2–F5) of the piglets in different groups. A1–F1 Correspond to piglets in 105 TCID50 PRV-GD2013-ΔgI/gE vaccinated groups. A2–F2 Correspond to piglets in 104 TCID50 PRV-GD2013-ΔgI/gE vaccinated groups. A3–F3 Correspond to piglets in 103 TCID50 PRV-GD2013-ΔgI/gE vaccinated groups. A4–F4 Correspond to piglets in Bartha k61 vaccinated groups. A5–F5 Correspond to piglets in unvaccinated groups. Histopathologic examination and H&E staining. Magnification, 200 ×
Discussion
In the 1960s, PRV was found to be endemic in pig farms in China. Vaccination was considered to be an effective strategy to control and eradicate PRV. At present, Bartha-K61 is a commonly used vaccine in China with a good safety record, good immunogenicity, and effective protection. However, since 2011, Chinese pigs vaccinated with Bartha-K61 have been showing typical symptoms of PR, which is strongly infectious to piglets leading to a 50% mortality rate [14, 16]. Furthermore, the PRV-GD2013 strain was isolated from Bartha-K61-vaccinated pig herds, indicating that Bartha-K61 vaccine has limited protection against the PRV variants found in China. Since 2017, some variant PRVs have been detected in the eastern provinces of China [27]. Interestingly, there is a close relationship between PRV and geographical location [27]. Thus, there may be a need for the development of new vaccines in different regions of China. The GD2013 strain was compared with other PRV variants. Genetically, the GD2013 strain is closely related to the strains isolated in China in recent years, but it is relatively distant from isolates of western countries, including Becker and Kaplan. Additionally, when compared with the Classic SC strain, there are nucleotide insertions and mutations in the gE gene of the GD2013 strain, and, when compared with the JS-2012-China-2012, QXY-China-2013 and HeN1-China-2014, which has 8, 12 and 9 amino acid differences (Fig. S2). Thus, the newly emerging GD2013 strain is a PRV variant with unique molecular characteristics. However, the mechanism of how the variant evolved is unknown. Based on these findings, the development of a live attenuated vaccine based on a novel PRV variant would be crucial for PRV control. In this study, we constructed and evaluated PRV-GD2013-ΔgI/gE, derived from PRV variant PRV-GD2013 as a live vaccine candidate.
A live vaccine candidate requires safety, broad protection, and immunogenicity. The criteria of target genes for constructing a live vaccine candidate are inessential for replication but is essential for virulence [28]. Extensive studies have shown that gI and gE perfectly meet these standards [13, 29,30,31]. gI/gE forms a complex and is responsible for neurotropism and reactivation. Our studies confirmed that the growth curve and plaque size of PRV-GD2013-ΔgI/gE was not significantly reduced when compared to that of PRV-GD2013. Also, the LD50 of PRV-GD2013 was higher than PRV-GD2013-ΔgI/gE in mice, indicating that the gI/gE genes are related to the virulence of PRV. PRV can infect pigs of all ages and the mortality rate of suckling piglets can be as high as 100% [5]. The efficacy evaluation demonstrated that a single inoculation of 105 TCID50 PRV-GD2013-ΔgI/gE was capable of providing full protection for piglets challenged with 105 TCID50 PRV-GD2013. However, the DMEM and Bartha-K61 vaccinations did not provide full protection for piglets challenged with 105 TCID50 PRV-GD2013. These results demonstrate that immunization with the PRV-GD2013-ΔgI/gE strain with a lower TCID50 affords sufficient protection for piglets challenged with PRV-GD2013.
Furthermore, in the growing pigs, the gB-specific antibodies generated by the PRV-GD2013-ΔgI/gE vaccine stimulated significant levels of the gB antibody against PRV-GD2013, whereas the commercial Bartha-K61 vaccine demonstrated substantially lower protection against PRV-GD2013, with the gE-specific antibodies not being detected throughout the experiment. These data further suggest that Bartha-K61 is insufficient for the prevention of PRV infection, but that the PRV-GD2013-ΔgI/gE strain is non-toxic to two-week-old piglets, and is immunogenic for immunized piglets.
Conclusions
We generated a PRV-GD2013-ΔgI/gE PRV mutant with gI/gE deletion using overlapping PCR and homologous recombination techniques. The PRV-GD2013-ΔgI/gE strain is non-toxic to two-week-old piglets and has full immunogenicity against PRV-GD2013. Antibodies produced by PRV-GD2013-ΔgI/gE-immunized piglets can be distinguished from those produced by wild-type strain infection using commercial gE-ELISA and gB-ELISA kits. This technology can be used for differential diagnosis. We suggest that the recombinant virus PRV-GD2013-ΔgI/gE is an attractive vaccine candidate to control the current epidemic of swine PR in China.
Methods
Cells and viruses
The PRV strain GD2013 was isolated in 2013 from the brain tissue of a sick piglet that had been vaccinated with Bartha-K61 in Guangdong Province, China. All viruses were propagated in BHK-21 cells (ATCC), which were cultured in Dulbecco’s Modified Eagle’s medium (DMEM, Gibco) supplemented with 10% fetal bovine serum (FBS, Hyclone) at 37 °C in a humidified 5% CO2 incubator.
Construction of transfer vector by overlapping PCR and homologous recombination technology
Construction of a pBLE-gI-gE transfer vector
DNA from the GD2013 strain was used as a template for PCR amplification using the primers listed in Table 3. The areas of the PRV genome flanking of gI and gE genes were amplified using primers P1/P2 and P3/P4, respectively. The amplified gene fragments were used as the recombination homologous arms (L-arm and R-arm) of the pBLE-gI-gE transfer vector. P2 and P3 contained the overlapping complementary sequences, while P1 and P4 directed cloning into the vector. Both products were gel-purified after the first round of PCR. The second round of PCR used the two resulting PCR products and the P1 and P4 primers to obtain the fragment encoding gI-gE. The fragments were then assembled by homologous recombination and ligated into NotI- and ClaI- cleaved pBluescript KS (+) vectors to generate the pBLE-gI-gE transfer vector (Fig. 1). The recombinant plasmid was confirmed by DNA sequencing. All primers used to amplify the sequence are listed in Table 3.
Construction of the pBLE-gI-EGFP-gE transfer vector
The coding sequences flanking the gI and gE gene fragment were amplified using primers P1′/P2′ and P3′/P4′, respectively. The amplified gene fragments were used as the L-arm and R-arm of the pBLE-gI-gE transfer vector. A signal sequence fragment, CMV-EGFP-SV40 polyA, was determined from the pEGFP-C3 plasmid by PCR using primers P5′/P6′. The P2′/P5′ and P3′/P6′ primer pairs contained the overlapping complementary sequences, while P1′and P4′ contained NotI and ClaI recognition sites for directed cloning into the vector. Both products were gel-purified after the first round of PCR. The second round of PCR used the three resulting PCR products to obtain the fragment encoding gI–EGFP-gE. The fragments were then assembled by homologous recombination and ligated into NotI- and ClaI- cleaved pBluescript KS (+) vectors to construct the pBLE-gI-EGFP-gE transfer vector (Fig. 1). The recombinant plasmid was confirmed by DNA sequencing. All primers used to determine the sequence are listed in Table 3.
Generation of recombinant viruses
The genomic DNA of PRV-GD2013 was extracted and purified using commercially available kits (AXYGEN, LOT#: AP-MIN-BF-VNA-250, USA) according to the manufacturer’s instructions. The pBLE-gI-EGFP-gE transfer plasmid and the PRV-GD2013 genomic DNA were co-transfected into BHK-21 (2 × 105 cells/dish) cells using the Lipofectamine® 2000 transfection reagent (Invitrogen), according to the manufacturer’s instructions. After cytopathic effects (CPEs) were observed, the transfected culture was harvested. After two to three freeze-thaw cycles, monolayers of BHK-21 cells were inoculated with 100 μl of lysate per well and covered with 1% low-melting agarose. Recombinant viruses with green fluorescent plaques were screened under fluorescent microscopy. After several rounds of plaque purification, the recombinant virus stably expressing EGFP was screened and hereafter referred to as PRV-GD2013-ΔgI/gE-EGFP. Similarly, PRV-GD2013-ΔgI/gE-EGFP genomic DNA and pBLE-gI-gE were co-transfected into BHK-21 cells, in the same way, to generate a recombinant virus without EGFP expression, hereafter referred to as PRV-GD2013-△gI/gE (Fig. 1). Gene recombination was identified by PCR (using the primers were listed in Table 3, gI-F/gI-R, gE-F/gE-R, gIF/gER) and DNA sequencing.
Growth kinetics
Growth kinetics were determined by using a one-step growth curve and plaque size calculation. Monolayers of BHK-21 cells were inoculated with PRV-GD2013-ΔgI/gE and PRV-GD2013 at an MOI of 1 respectively. At 2 hours post-infection (hpi), the monolayers were washed twice with phosphate-buffered saline (PBS), and 2 ml of DMEM (containing 2% FBS) was added. The culture supernatants (200 μl) were collected at different time points (0, 4, 8, 12, 16, 20, 24, 28, and 32 hpi). The supernatants collected at these nine-time points were then used to calculate the TCID50 of the virus according to the Reed-Muench formula. One-step growth curves were drawn based on the result of these calculations. Plaque sizes were performed for PRV-GD2013-ΔgI/gE and PRV-GD2013 as described previously [32].
Animal experiments
Pathogenic testing in mice
Sixty-five two-week-old specific-pathogen-free (SPF) female BALB/c mice were obtained from the Laboratory Animal Center of Southern Medical University and were randomly divided into 13 groups of five. Groups 1–6 were subcutaneously injected in the inguinal region with 0.1 ml of different doses (10, 102, 103, 104, 105 or 106 TCID50) of PRV-GD2013. Groups 7–12 were subcutaneously injected in the inguinal region with different doses (10, 102, 103, 104, 105 or 106 TCID50) of PRV-GD2013-ΔgI/gE. The mice in group 13 were subcutaneously injected in the inguinal region with 0.1 ml of DMEM and served as negative controls. Clinical signs and mortality in the mice were observed and recorded daily after inoculation for 14 days. All surviving mice were euthanized by intravenous administration of an overdose of sodium pentobarbital after the experiment. The LD50 of the virus was calculated according to the Reed-Muench formula.
Pathogenic testing in piglets
Fifteen two-week-old piglets without PRV antibodies were obtained from a local farm and were randomly divided into three groups of five and housed separately. Group A was inoculated i.n. with 2 ml of 105 TCID50 PRV-GD2013. Group B was inoculated i.n. with 2 ml of 105 TCID50 PRV-GD2013-ΔgI/gE. Group C was inoculated i.n. with 2 ml of DMEM to serve as a negative control. Following inoculation, rectal temperatures and clinical symptoms were monitored and recorded daily. Virus shedding was determined by the daily collection of nasal and rectal swabs and blood samples were collected at 0, 3, 7, 10, 14, and 17 dpi. At 17 dpc, all surviving piglets were euthanized by intravenous administration of an overdose of sodium pentobarbital and were then necropsied within 2 h of death. Any piglets that died before the end of the experiments were immediately necropsied. Tissue samples were collected for pathological examination.
Immunization and challenge experiments
Twenty-five two-week-old piglets without PRV antibodies were obtained from a local farm and were randomly divided into five groups of five and housed separately. Groups A-C were vaccinated intramuscularly (i.m.) with different doses (105, 104, 103 TCID50) of PRV-GD2013-ΔgI/gE. Group D was vaccinated i.m. with 105 TCID50 Bartha-K61. Group E was inoculated i.m. with DMEM to serve as a negative control. After 2 weeks post-vaccination, all piglets were challenged i.n. with 105 TCID50 of PRV-GD2013. After vaccination and challenge, rectal temperatures and clinical symptoms were monitored and recorded daily. Virus shedding was determined by the daily collection of nasal and rectal swabs, and blood samples were collected at 0, 3, 7, 10, and 14 dpi. At the end of the experiment, all surviving piglets were euthanized by intravenous administration of an overdose of sodium pentobarbital and were then necropsied within 2 h of death. Any piglets that died before the end of the animal experiments were immediately necropsied. Tissue samples from the brain, lung, liver, kidney, spleen, and lymph nodes were collected for histopathologic examination.
Enzyme-linked immunosorbent assay (ELISA)
Levels of gB- and gE-specific antibodies were measured using commercial PRV antibody detection kits (IDEXX, USA) according to the manufacturer’s instructions.
Statistical analysis
All experimental data were analyzed using SPSS 13.0 software and Sigma Plot 12.0. All data are presented as mean ± SD.
Availability of data and materials
The datasets generated and/or used during the current study are not available to public as it is owned by the South China Agricultural University, China. However, these can be requested via email from the corresponding authors; Mrs. Mingqiu Zhao (zmingqiu@scau.edu.cn) and/or Prof. Dr. Jinding Chen (jdchen@scau.edu.cn).
Abbreviations
- BHK-21:
-
Baby Hamster Syrian Kidney
- CPEs:
-
Cytopathogenic effects
- DNA:
-
Deoxyribonucleic acid
- dpc:
-
Days post-challenge
- dpi:
-
Days post-inoculation
- ELISA:
-
Enzyme-linked immunosorbent assay
- gB:
-
Glycoprotein B
- gC:
-
Glycoprotein C
- gD:
-
Glycoprotein D
- gE:
-
Glycoprotein E
- gG:
-
Glycoprotein G
- gI:
-
Glycoprotein I
- hpi:
-
Hours post-infection
- i.m.:
-
Intramuscularly
- i.n.:
-
Intranasally
- LD50:
-
50% lethal dose
- MOI:
-
Multiplicity of infection
- PCR:
-
Polymerase chain reaction
- PR:
-
Pseudorabies
- PRV:
-
Pseudorabies virus
- TCID50:
-
50% tissue culture infectious dose
- TK:
-
Thymidine kinase
- TRS:
-
Terminal repeat sequence
- IRS:
-
Internal repeat sequence
- US:
-
Unique short region
- UL:
-
Unique long region
References
Jons A, Gerdts V, Lange E, Kaden V, Thomas C. Mettenleiter. Attenuation of dUTPase-deficient pseudorabies virus for the natural host. Vet Microbiol. 1997;56(1–2):47–54. https://doi.org/10.1016/S0378-1135(96)01353-3.
Dijkstra JM, Gerdts V, Klupp BG, Mettenleiter TC. Deletion of glycoprotein gM of pseudorabies virus results in attenuation for the natural host. J Gen Virol. 1997;34(9):2147–51. https://doi.org/10.1099/0022-1317-78-9-2147.
Klupp BG, Lomniczi B, Visser N, Fuchs W, Mettenleiter TC. Mutations affecting the UL21 gene contribute to avirulence of pseudorabies virus vaccine strain Bartha. Virology. 1995;212(2):466–73. https://doi.org/10.1006/viro.1995.1504.
Wittmann G, Rziha HJ. Aujeszky’s disease (Pseudorabies) in pigs. Dev Vet Virol. 1989;9:230–325. https://doi.org/10.1007/978-1-4613-1587-2_7.
Pomeranz LE, Reynolds AE, Hengartner CJ. Molecular biology of Pseudorabies virus: impact on Neurovirology and veterinary medicine. Microbiol Mol Biol Rev. 2005;69(3):462–500. https://doi.org/10.1128/MMBR.69.3.462-500.2005.
Klupp BG, Hengartner CJ, Mettenleiter TC, Enquist LW. Complete, annotated sequence of the pseudorabies virus genome. J Virol. 2004;78(1):424–40. https://doi.org/10.1128/JVI.78.1.424-440.2004.
Oirschot JTV, Kaashoek MJ, Rijsewijk FAM, Stegeman JA. The use of marker vaccines in eradication of herpesviruses. J Biotechnol. 1996;44(1–3):75–81. https://doi.org/10.1016/0168-1656(95)00129-8.
Mettenleiter TC, Klupp BG, Weiland F, Visser N. Characterization of a quadruple glycoprotein-deleted pseudorabies virus mutant for use as a biologically safe live virus vaccine. J Gen Virol. 1994;75(7):1723–33. https://doi.org/10.1099/0022-1317-75-7-1723.
Babic N, Makoschey BKA, Flamand A, Mettenleiter TC, Klupp BG. Glycoprotein gH of Pseudorabies virus is essential for penetration and propagation in cell culture and in the nervous system of mice. J Gen Virol. 1996;77(9):2277–85. https://doi.org/10.1099/0022-1317-77-9-2277.
Grimm KS, Klupp BG, Harald G, Müller FM, Walter F, Mettenleiter TC. Analysis of viral and cellular factors influencing herpesvirus-induced nuclear envelope breakdown. J Virol. 2012;86(12):6512–21. https://doi.org/10.1128/JVI.00068-12.
Kritas SK, Pensaert MB, Mettenleiter TC. Role of envelope glycoproteins gI, gp63 and gIII in the invasion and spread of Aujeszk's disease virus in the olfactory nervous pathway of the pig. J Gen Virol. 1994;75(9):2319–27. https://doi.org/10.1099/0022-1317-75-9-2319.
Lomniczi B, Watanabe S, Ben-Porat T, Kaplan AS. Genome location and identification of functions defective in the Bartha vaccine strain of pseudorabies virus. J Virol. 1987;61(4):796–801. https://doi.org/10.1016/0166-0934(87)90156-X.
Szpara ML, Tafuri YR, Lance PS, Rafi S, Verstrepen KJ, Matthieu L, et al. A wide extent of inter-strain diversity in virulent and vaccine strains of alphaherpesviruses. PLoS Pathog. 2011;7(10):e1002282. https://doi.org/10.1371/journal.ppat.1002282.
An TQ, Peng JM, Tian ZJ, Zhao HY, Li N, Liu YM, et al. Pseudorabies virus variant in bartha-k61-vaccinated pigs, China, 2012. Emerg Infect Dis. 2013;19(11):1749–55. https://doi.org/10.3201/eid1911.130177.
Mahy BWJ. Animal viruses: molecular biology. Emerg Infect Dis. 2008;14:867a–867. https://doi.org/10.3201/eid1405.080077.
Luo Y, Li N, Cong X, Wang CH, Du M, Li L, et al. Pathogenicity and genomic characterization of a pseudorabies virus variant isolated from Bartha-K61-vaccinated swine population in China. Vet Microbiol. 2014;174(1-2):107–15. https://doi.org/10.1016/j.vetmic.2014.09.003.
Wang CH, Yuan J, Qin HY, Luo Y, Cong X, Li Y, et al. A novel gE-deleted pseudorabies virus (PRV) provides rapid and complete protection from lethal challenge with the PRV variant emerging in Bartha-K61-vaccinated swine population in China. Vaccine. 2014;32(27):3379–85. https://doi.org/10.1016/j.vaccine.2014.04.035.
Wu R, Bai C, Sun J, Chang S, Zhang X. Emergence of virulent pseudorabies virus infection in northern China. J Vet Sci. 2013;14(3):363–5. https://doi.org/10.4142/jvs.2013.14.3.363.
Yu X, Zhou Z, Hu D, Zhang Q, Han T, Li X, et al. Pathogenic pseudorabies virus, China, 2012. Emerg Infect Dis. 2014;20(1):102–4. https://doi.org/10.3201/eid2001.130531.
Cong X, Lei JL, Xia SL, Wang YM, Li Y, Li S, et al. Pathogenicity and immunogenicity of a gE/gI/TK gene-deleted pseudorabies virus variant in susceptible animals. Vet Microbiol. 2016;182:170–7. https://doi.org/10.1016/j.vetmic.2015.11.022.
Gu Z, Jing D, Wang J, Hou C, Sun H, Yang W, et al. A novel inactivated gE/gI deleted pseudorabies virus (PRV) vaccine completely protects pigs from an emerged variant PRV challenge. Virus Res. 2015;195:57–63.
Wang T, Xiao Y, Yang Q, Wang Y, Sun Z, Zhang C, et al. Construction of a gE-deleted Pseudorabies virus and its efficacy to the new-emerging variant PRV challenge in the form of killed vaccine. Biomed Res Int. 2015;2015:1–10. https://doi.org/10.1155/2015/684945.
Wang J, Song Z, Ge A, Guo R, Qiao Y, Xu M, et al. Safety and immunogenicity of an attenuated Chinese pseudorabies variant by dual deletion of TK&gE genes. BMC Vet Res. 2018;14(1):287–98. https://doi.org/10.1186/s12917-018-1536-7.
He W, Auclert LZ, Zhai X, Wong G, Zhang C, Zhu H, et al. Interspecies transmission, genetic diversity, and evolutionary dynamics of Pseudorabies virus. J Infect Dis. 2019;219(11):1705–15. https://doi.org/10.1093/infdis/jiy731.
Ho SN, Hunt HD, Horton RM, Pullen JK, Pease LR. Site-directed mutagenesis by overlap extension using the polymerase chain reaction. Gene. 1989;77(1):51–9. https://doi.org/10.1016/0378-1119(89)90358-2.
Jones ML, Barnard RT. Chimerization of multiple antibody classes using splice overlap extension PCR. Biotechniques. 2005;38(2):181–2. https://doi.org/10.2144/05382BM01.
Zhai X, Zhao W, Li K, Zhang C, Wang C, Su S, et al. Genome characteristics and evolution of Pseudorabies virus strains in eastern China from 2017 to 2019. Virol Sin. 2019;34(1–2). https://doi.org/10.1007/s12250-019-00140-1.
Wu CY, Liao CM, Chi JN, Chien MS, Huang CJ. Growth properties and vaccine efficacy of recombinant pseudorabies virus defective in glycoprotein E and thymidine kinase genes. J Biotechnol. 2016;229:58–64. https://doi.org/10.1016/j.jbiotec.2016.05.009.
Kratchmarov R, Tal K, Greco TM, Taylor MP, Toh Hean CN, Cristea IM, et al. Glycoproteins gE and gI are required for efficient KIF1A-dependent anterograde axonal transport of alphaherpesvirus particles in neurons. J Virol. 2013;87(17):9431–40. https://doi.org/10.1128/JVI.01317-13.
Zhu L, Yi Y, Xu Z, Cheng L, Tang S, Guo W. Growth, physicochemical properties, and morphogenesis of Chinese wild-type PRV Fa and its gene-deleted mutant strain PRV SA215. Virol J. 2011;8(1):272–2. https://doi.org/10.1186/1743-422X-8-272.
Hu RM, Zhou Q, Song WB, Sun EC, Zhang MM, He QG, et al. Novel pseudorabies virus variant with defects in TK, gE and gI protects growing pigs against lethal challenge. Vaccine. 2015;33(43):5733–40. https://doi.org/10.1016/j.vaccine.2015.09.066.
Tong W, Li G, Liang C, Liu F, Tian Q, Cao Y, et al. A live, attenuated pseudorabies virus strain JS-2012 deleted for gE/gI protects against both classical and emerging strains. Antivir Res. 2016;130:110–7. https://doi.org/10.1016/j.antiviral.2016.03.002.
Acknowledgements
We would sincerely like to thank Dr. Zunyang Song for assistance in experiment.
Funding
This work was supported by the National Key Research Development Program of China (Nos. 2016YFD0500700, 2017YFD0500600), the Science and Technology Program of Guangzhou, China (Nos. 201803020005), and the National Natural Science Foundation of China (Nos. 31672590, U1405216, 31472200), the Science and Technology Planning Project of Guangdong Province of China (Nos. 2019B02021103) and the Key Research Projects of Universities of Guangdong Province. The funding body had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
WL, JC and MZ conceived and designed the experiment. WL, DZ, MZ, EZ and BX performed the experiments. WL and HL carried out the analysis. WL wrote the manuscript. WL revise the manuscript. All the authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
The use of piglets in this study was approved by Daguang farming Co., Ltd. and the use of mice in this study was approved by the Laboratory Animal Center of Southern Medical University. All procedures involving animals were conducted in compliance with the guidelines of the Animal Welfare Act and the Guide for the Care and Use of Laboratory Animals of South China Agricultural University (Guangzhou, China, Approval No. SYXK [Yue] 2014–0136).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
Recombinant viruses were analyzed by PCR. DNAMarker: DL5000; 1: PRV-GD2013-△gI/gE; 2: PRV-GD2013-△gI/gE-EGFP; 3: PRV-GD2013; 4: Negative control.
Additional file 2: Figure S2.
Multiple alignment of the gE protein in PRV-GD2013, JS-2012-China-2012, QXY-China-2013 and HeN1-China-2014.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, W., Zhuang, D., Li, H. et al. Recombinant pseudorabies virus with gI/gE deletion generated by overlapping polymerase chain reaction and homologous recombination technology induces protection against the PRV variant PRV-GD2013. BMC Vet Res 17, 164 (2021). https://doi.org/10.1186/s12917-021-02861-6
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12917-021-02861-6