
Abstract
Background
hyperlipidemia acute pancreatitis (HTG-AP) is a major hidden danger affecting human health, however, whether there is a protective effect of resveratrol on HTG-AP is unclear. Therefore our study was aimed to investigate the preventive effect and the underlying mechanism of resveratrol in the HTG-AP mice model.
Methods
This research was divided into two parts. In the first part, mice were adaptively fed with normal chow or HFD for 6 weeks. From the second week, resveratrol-treated mice were in intragastric administration with resveratrol (45 mg/kg/d) for 4 weeks. In the second part, the procedures were the same as the first part. After the last intragastric administration with resveratrol, all mice were intraperitoneal injections of cerulean.
Results
We found resveratrol effectively inhibited pancreatic pathological injury in the HFD, AP, and HTG-AP mice. Resveratrol reduced the LPS, IL-6, TNF-α, and MCP-1 expressions in the HFD mice. Resveratrol also reduced TNF-α, MDA, and MCP-1 expressions and increased SOD and T-AOC expressions in the AP and HTG-AP mice. Furthermore, resveratrol suppressed the NF-κB pro-inflammatory signaling pathway in pancreatic tissues in the AP and HTG-AP mice. Moreover, resveratrol improved the gut microbiota in the HFD mice.
Conclusion
The resveratrol pre-treatment could attenuate pancreas injury, inflammation, and oxidative stress in the HTG-AP mice, via restraining the NF-κB signaling pathway and regulating gut microbiota. Therefore, Our study proved that the resveratrol pre-treatment had a preventive effect on HTG-AP.
Similar content being viewed by others
Background
The annual incidence of acute pancreatitis (AP) is between 13 and 45 cases per 100,000 populations worldwide [1]. Hypertriglyceridemia (HTG) is the third most common cause of AP accounting for up to 7% of the cases [2] with gallstones accounting for 60% [3] and alcohol for 30% [4]. Whereas, HTG is the most common established cause in some specific physiological states, such as pregnancy [5]. The clinical course of HTG-AP is highly similar to other causes of AP, but HTG is the only significant clinical feature. Furthermore, HTG-AP is often accompanied by higher severity and an increased complication rate [6]. Clinical studies have reported that HTG aggravated the course of AP and aggravated the inflammatory response [7, 8]. However, the underlying mechanism is still unclear. Therefore, a novel effective and risk-free HTG-AP therapeutic methods are in urgent demand. In this regard, natural compounds with potent antioxidative and anti-inflammatory activities represent invaluable resources for development as RA therapeutics.
Resveratrol (3,5,4′-trihydroxy-trans-stilbene, Rev) is a natural polyphenol, which widely exists in various plants, such as Polygonum cuspidatum, in fruits, including grapes and berries, in peanuts, and in red wine [9]. Resveratrol plays a regulatory role through a series of mechanisms, including scavenging ROS [10], antioxidative [11], and anti-inflammatory activities [12]. Recent studies have established that resveratrol owns the potential in the prevention or treatment of chronic inflammation-related disorders such as cardiovascular diseases, diabetes, obesity, and cancer [13,14,15,16]. Besides, it has an association with some metabolic syndrome diseases including glucose intolerance [17], altered cholesterolemia [18], and hypertriglyceridemia [19]. In view of this, we intended to address the question regarding the effect of resveratrol in HTG-AP and the underlying mechanisms in vivo using a caerulein-induced mice model with a high-fat diet (HFD), for cerulein-induced pancreatitis is the most-well-characterized and widely used experimental model for acute pancreatitis.
Herein, we presented evidence supporting the preventive effect of resveratrol on HTG-AP through alleviating hypertriglyceridemia, oxidative stress, and inflammation in HTG-AP mice and regulating HFD-induced gut microbiota disorders in vivo.
Methods
Animals
Forty-eight male C57BL/6 J mice, 18.0–20.0 g, aged 6 weeks, were purchased from Animal Center of West China Medical College, Sichuan University (Chengdu, China). All animals were housed in individual cages under a 12-hour light/dark-cycle environment, provided free water and food, and approved by the ethics committee of Affiliated Hospital of North Sichuan Medical College (No. 2022.03). All efforts were aimed to alleviate animal suffering.
Materials
Resveratrol (≥99%, Sigma, USA), sodium carboxymethyl cellulose (UNIVAL, Germany), cerulein (Sigma, USA), total cholesterol (TC) and total triglyceride (TG) content assay kits (Solarbio, China), LPS (cusabio, China), MCP-1 (Abcam, USA), TNF-α (Beyotime, China), IL-6 (Abcam, USA) malondialdehyde (MDA) (Solarbio, China), superoxide dismutase (SOD) (Solarbio, China), and total antioxidant capacity (T-AOC) (Solarbio, China) ELISA kits.
Model preparation and animal grouping
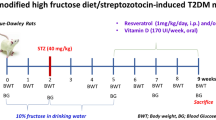
This research was divided into two parts. In the first part of experiments, mice were randomly assigned to 4 groups (n = 6) as follows: chow, chow. Rev., HFD, HFD. Rev. Mice were adaptively fed with normal chow or HFD for 2 weeks, and then chow. Rev. and HFD. Rev. groups were intragastric administration with resveratrol (45 mg/kg/d) for 4 weeks. Resveratrol (≥99%, Sigma, USA) was dissolved in 0.5% sodium carboxymethyl cellulose (UNIVAL, Germany). Then all mice were given an intraperitoneal injection of 50 mg/kg sodium pentobarbital and sacrificed by cervical dislocation. Blood and colon tissue were collected and 3 mice from each group were randomly selected to collect colon contents for 16 s rRNA sequencing of gut microbiota.
The second part of experiments was designed to determine the effect of pre-treatment with resveratrol in the HTG-AP mice and the HTG-AP mice model was established according to the reference [20]. In brief, mice were randomly assigned to 4 groups (n = 6) as follows: chow + cerulein, chow. Rev. + cerulein, HFD + cerulein, HFD. Rev. + cerulein. The procedures were the same as the first part of experiments. But 24 h after the last intragastric administration with resveratrol, all the groups were intraperitoneal injections of cerulein (Sigma, USA) by two times, each time at a dose of 40 μg/kg body weight. After 12 h, all mice were given an intraperitoneal injection of 50 mg/kg sodium pentobarbital and sacrificed by cervical dislocation. Blood and colon tissue were collected.
Measurement of TC and TG
Blood biochemical indicators of the lipid profile were assessed. The concentrations of total cholesterol (TC) were measured by Micro total cholesterol (TC) content assay kit (Solarbio, Beijing, China) and triglyceride (TG) was assessed by triglyceride content assay kit (Solarbio, Beijing, China) according to the manufacturer’s instructions.
Enzyme-linked immunosorbent assay (ELISA)
Blood was collected from mice in each group and centrifuged at 15000 rpm for 10 min to obtain the serum. In the first part of experiments, the levels of proinflammatory cytokines including LPS (cusabio, China), MCP-1 (Abcam, USA), TNF-α (Beyotime, China), and IL-6 (Abcam) were determined by ELISA according to the manufacturer’s instructions. In the second part of experiments, the levels of malondialdehyde (MDA) (Solarbio, Beijing, China), superoxide dismutase (SOD) (Solarbio, Beijing, China), and total antioxidant capacity (T-AOC) (Solarbio, Beijing, China) were measured by ELISA according to the manufacturer’s instructions. The OD value of each well was immediately read at 450 nm.
Histopathological assessment
Pancreatic tissues from each group were fixed in 4% paraformaldehyde at room temperature, embedded in paraffin, and sectioned at a thickness of 5 μm. In the immumohistochemical staining, TNF-α and MCP-1 antibody (1:400) and related conjugated secondary antibody were used. In the H&E staining, the tissues were stained with hematoxylin and eosin (H&E). The histopathological change was observed under the light microscope (Olympus, Tokyo, Japan) at 400× magnification.
Western blot assay
After the pancreatic tissues were obtained, proteins were extracted using RIPA lysis buffer (Beyotime, Beijing, China), and protein concentrations were measured using a BCA protein assay kit (Vazyme, Nanjing, China) following the manufacturer’s protocols. The protein samples were then separated by 10% SDS-PAGE and transferred onto polyvinylidene fluoride (PVDF) membranes. Next, the PVDF membranes were blocked by 5% skim milk. After being cultured for 2 h at room temperature, they were then incubated overnight at 4 °C with specific primary antibodies including p65, p-p65, TNF-α, and IL-6. After washing three times, the blots were subsequently incubated with a goat horseradish peroxidase-conjugated secondary antibody for 2 h at room temperature. After four times washing for 10 min in Tris-buffered saline with Tween-20 (TBST), the membranes were detected using a chemiluminescence detection system. The intensity of the bands was quantified by ImageJ software. β-actin served as a loading control.
DNA extraction
The colon content DNA of mice in each group was extracted with ZR Fecal DNA Extraction Kit (Zymo Research, CA, USA). The buffer solution was added to 200 mg feces from each group to prepare fecal homogenate, and the sediments were centrifuged using a vortex mixer after incubation at 70 °C. Then the supernatant was extracted, and inhibitors were added. The sediments were centrifuged, aspirated the supernatant again. The buffer solution was added and incubated at 70 °C for 10 min, and 200 μl absolute ethanol was added ultimately. After the sample was purified, the DNA sample is obtained, which is quantified by an ultraviolet spectrophotometer and tested for purity. After this, the DNA quality is analyzed by agarose gel electrophoresis.
16sRNA sequencing of gut microbiota
Bacterial RNA was amplified by RT-PCR targeting the V3-V4 hypervariable regions of the 16 s RNA gene and using specific primers (319F: 5′ ACTCCTACGGGAGGCAGCAG 3′; 806R: 3′ ACTCCTACGGGAGGCAGCAG 5′). Amplicons were pooled and paired-end sequenced on an Illumina MiSeq (Illumina) in the Shanghai Personal Biotechnology Co., Ltd. (Shanghai, China). The Quantitative Insights Into Microbial Ecology (QIIME, v1.8.0) pipeline was employed to process the sequencing data, as previously described. Sequence processing and microbial composition analysis were performed with the Quantitative Insights into Microbial Ecology (QIIME) software package, version 1.9.1. After quality filters, the remaining high-quality sequences were clustered into operational taxonomic units (OTUs) at 97% sequence using the reference-based USEARCH (version 5.2) pipeline in QIIME, using the May 2013 release of the GreenGenes 99% OTU database as a closed reference. The raw data and sequencing sample information have been submitted to the SILVA database to classify.
Statistical analysis
All data are expressed as mean ± SD. Statistical analysis was performed using GraphPad Prism 7 (GraphPad software, USA). Statistical differences among the groups were determined using one-way ANOVA or two-way ANOVA to compare differences between experimental and control groups. Results with p < 0.05 were considered statistically significant. All experiments were performed at least in triplicate.
Results
Resveratrol decreased serum levels of TC and TG in HFD and HTG-AP mice
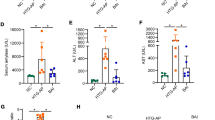
The levels of serum TC and TG were conducted to evaluate the alterations in lipid profiles in different groups. As shown in Fig. 1, the serum concentration of TC and TG varied among groups. In the HFD model group, the TC and TG levels were markedly higher than those in the chow group, which indicated that high lipid levels were associated with HFD (Fig. 1A). Similarly, the levels of TC and TG in the HFD + cerulein model group (HTG-AP) were significantly higher than those in the chow + cerulein group (Fig. 1B). However, administration of resveratrol markedly decreased the TC and TG levels compared with the HFD model group and HFD + cerulein model group in the same way.

Serum lipid profiles after treatment with resveratrol in different models. The concentrations of TC and TG in HFD model (A), TC and TG in HFD + cerulein model (B) were analyzed among different groups. TC, total cholesterol; TG, triglyceride. Bars represent the mean ± S.D. from three independent experiments. *p < 0.05, **p < 0.01, ****p < 0.0001
Resveratrol alleviated histopathological damage of pancreatic tissue in HFD and HTG-AP mice
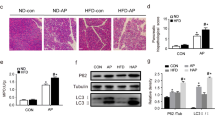
The pancreatic tissues from all the groups were collected and stained with H&E to observe the histopathological change. In the HFD model group, the pancreatic tissue of chow group displayed a clear tissue structure with no obvious abnormality in the pancreatic ducts, islet, and acini. No obvious inflammatory cell infiltration was observed. And the histopathological characteristics in chow. Rev. group was similar to the chow group. However, pancreatic tissue in the HFD group displayed interstitial edema, and a small number of inflammatory cells infiltrated into the perivascular and stroma. These cells were mainly lymphocytes with round nuclei and deep staining. Nevertheless, an improvement in pathological changes was observed following resveratrol treatment compared with those of the HFD group (Fig. 2A). Likewise, in the HTG-AP model group, pancreatic tissue of chow + cerulein group displayed local acinar epithelial cell degeneration and necrosis, nucleus contraction, a small number of inflammatory cells infiltrated in the stroma and around the blood vessels, and these cells were mainly round hyperchromatic lymphocytes. And the pathological changes in HFD + cerulein group had similarities to those of chow + cerulein group following the hyperplasia of acinar stromal and slight separation of some acinus. Encouragingly, all these pathological changes can be reversed by treatment with resveratrol (Fig. 2B).

Pathological changes in pancreatic tissues following treatment with resveratrol. Pancreatic tissues stained with H&E were visualized with a light microscope at a magnification of × 200 and × 400. Scale bar = 100 μm. A Pathological changes in HFD mice; B Pathological changes in HTG-AP mice. Green arrow: lymphocytic infiltration, blue arrow: acinar separation, and black arrow: acinar epithelial cells are denaturated and necrotic
Resveratrol down-regulated the expressions of MCP-1 and TNF-α in HFD and HTG-AP mice
Researchers have demonstrated that MCP-1 and TNF-α were overexpressed when the pancreatic tissue was damaged [21, 22]. Therefore, to evaluate the alterations in different groups, the expression of MCP-1 was measured in the HFD model group. The expression of MCP-1 and TNF-α were conducted in the HTG-AP model group. As for the expression of MCP-1, it was over-expressed in the HFD-treated, cerulein-treated, and HFD + cerulein-treated group (Fig. 3A and B) and this can be reversed by adding resveratrol. Similarly, as for the expression of TNF-α, it was over-expressed in the cerulein-treated and HFD + cerulein-treated group (Fig. 3C) compared with the resveratrol-treated group. Furthermore, the integral optical density (IOD) values of MCP-1 and TNF-α were shown. The expressions of MCP-1 and TNF-α were both up-regulated in HFD or HFD + cerulein group, but the addition of resveratrol can revert this (Fig. 3D).

Immunohistochemical staining of HFD and HTG-AP model mice for MCP-1 and TNF-α. A The expression of MCP-1 in HFD model group. Resveratrol-treated mice showed lower level of MCP-1 than the model group. B The expression of MCP-1 in HTG-AP model. C The expression of TNF-α in HTG-AP model group. D Bar graphs of the integral optical density (IOD) of tissue MCP-1 and TNF-α levels. Blue box: The target protein is deposited in the pancreas tissue *p < 0.05, **p < 0.01
Resveratrol reduced serum levels of inflammatory cytokines in HFD mice
The expression levels of inflammatory cytokines, including LPS, MCP-1, TNF-α, and IL-6 in the serum of HFD mice were measured by ELISA. The higher levels of LPS, MCP-1, TNF-α, and IL-6 in the HFD group compared with those in the chow group suggested that HFD may be related to the overexpression of inflammatory cytokines and the inflammatory response (Fig. 4). The HFD group displayed the highest levels of these cytokines compared with the chow group. However, the levels of LPS (Fig. 4A), MCP-1 (Fig. 4B), TNF-α (Fig. 4C), and IL-6 (Fig. 4D) were all significantly reduced following the administration of resveratrol. These results suggested that resveratrol might have a therapeutic effect against inflammation in HFD.

Serum proinflammatory cytokines levels, including LPS, MCP-1, TNF-α and IL-6. The concentrations of (A) LPS, (B) MCP-1, (C) TNF-α and (D) IL-6 in the different groups were determined by ELISA. Bars represent the mean ± S.D. from three independent experiments. *p < 0.05, **p < 0.01, ***p < 0.001
Resveratrol decreased oxidative stress level in HTG-AP mice
The oxidative stress-related markers in pancreatic tissue were measured to determine the antioxidant effects of resveratrol. The MDA activity, standing for lipid peroxidation, was markedly higher in HFD + cerulein group compared with the chow + cerulein, and it was reduced following resveratrol treatment (Fig. 5A). Additionally, the SOD activity, represent for free radical level, was significantly lower in HFD + cerulein group. The resveratrol-treated group suffered an increase in the activity of SOD (Fig. 5B). Ultimately, T-AOC activity, standing for total antioxidant level, in the resveratrol-treated group was higher than HFD + cerulein group (Fig. 5C). These results manifested that resveratrol can reverse the oxidative stress caused by cerulein.

The levels of oxidative stress-related markers in pancreatic tissue. The activities of (A) MDA, (B) SOD, and (C) T-AOC in the different groups were determined by ELISA. Bars represent the mean ± S.D. from three independent experiments. *p < 0.05, **p < 0.01
Resveratrol restrained NF-κB signaling pathway in HTG-AP mice pancreatic tissues
It is reported that the activation of nuclear factor-κB (NF-κB) signaling pathway can induce the expression and release of its downstream inflammatory cytokine IL-6 [23]. And TNF-α can serve as an activator of the NF-κB pathway [24]. The results of WB analysis for IL-6, TNF-α, and NF-κB pathway-related proteins in pancreatic tissue of HTG-AP mice were shown in Fig. 6. Bands of IL-6, TNF-α, NF-κB p65, phosphorylation-p65 (p-p65), IκBα and p-IκBα were displayed (Fig. 6A&H). The expressions of IL-6 (Fig. 6B), TNF-α (Fig. 6C), NF-κB p65 (Fig. 6D), p-p65 (Fig. 6E), and p-IκBα (Fig. 6H) were all significantly decreased in both chow. Rev. + cerulein and HFD. Rev. + cerulein groups, and similar results were shown in chow groups. These results suggested that resveratrol inhibited the activation of the NF-κB signaling pathway in pancreatic tissues, thus suppressing the activation of pro-inflammatory signaling, concerning the expressions of IL-6 and TNF-α.

Effect of resveratrol on the NF-κB signaling pathway. A The protein levels of IL-6, TNF-α, NF-κB p65 and p-p65 were measured by western blotting. B Relative protein level of IL-6; C Relative protein level of TNF-α; D Relative protein level of NF-κB p65; E Relative protein level of p-p65. F The protein levels of NF-κB IκBα and p-IκBα and p-p65 were measured by western blotting. G Relative protein level of NF-κB IκBα; H Relative protein level of NF-κB p-IκBα. β-actin served as a loading control. Bars represent the mean ± S.D. from three independent experiments. *p < 0.05, **p < 0.01
Resveratrol influenced the microbial diversity and the structure of community of feces in HFD mice
To explore the effect of resveratrol on gut microbiota in HFD mice, fecal samples were collected to analyze its diversity and richness. Multiple alpha diversity metrics of richness and diversity revealed that no significant difference was observed between groups, as shown in Chao1 and the observed species method (Fig. 7A). Bray-Curtis distance-based PCoA analysis was employed to determine the similarities and differences in the composition of gut microbiota among groups. HFD group showed a difference in gut microbiota compared with the chow group. HFD + Rev. group showed a movement in the first principal component (PC1) towards the direction of the chow group, thus HFD + Rev. shortened the distance with the chow group (Fig. 7B). Hierarchical clustering analysis revealed that the microbial communities in the resveratrol-treated group showed more similarities to those in the chow group (Fig. 7C). In short, resveratrol treatment can reverse the HFD-induced variations.

Effect of resveratrol on microbial diversity and structure of community of feces in HFD mice. A Richness and diversity of fecal microbiota. No significant difference was observed among groups. B Principal coordinate (PCoA) analysis of Bray-Curtis distance. C Cluster analysis of unweighted pair group method with arithmetic mean (UPGMA) based on Bray-Curtis distance
Resveratrol affected the composition of gut microbiota in HFD mice
The composition of gut microbiota was shown in Fig. 8A. In general, HFD-relative changes in fecal gut microbiota were characterized with a significantly higher relative abundance of Firmicutes, Actinobacteria, and Proteobacteria but a markedly lower relative abundance of Bacteroidetes. After treatment with resveratrol, it decreased the relative abundance of Firmicutes, Proteobacteria, and Actinobacteria, and increased the relative abundance of Bacteroidetes. Furthermore, the ratio of Firmicutes/Bacteroidetes was increased highly in the HFD group compared with the chow group and resveratrol treatment can reverse this (Fig. 8B). Consistent with beta diversity, clustering analysis of the top 50 genera highlighted differences in their distributions after treatment with resveratrol (Fig. 8C). Then, we analyzed the difference in the genus of each group and the statistically significant genera were shown. Compared with the chow group, the HFD mice showed a higher level in the relative abundance of Allobaculum and Streptococcus but a lower level in the relative abundance of Lactobacillus (Fig. 9). Specifically, compared with the HFD group, the HFD. Rev. group promoted the recovery of the relative abundance of Allobaculum and Lactobacillus and decrease the relative abundance of Streptococcus. In addition, in the chow and chow. Rev. groups, we found that the relative abundance of these three genera was consistent with the HFD and HFD. Rev. groups (Fig. 9).

Effect of resveratrol on composition of gut microbiota in HFD mice. A Difference in the relative abundance of Firmicutes, Bacteroidetes, Actinobacteria and Proteobacteria phylum among groups. B The ratio of Firmicutes/Bacteroidetes in phylum level. C The heat map of top 50 abundant genus. Double hierarchical dendrogram shows the bacterial distribution. *p < 0.05, **p < 0.01

Resveratrol-related alterations at genus levels of Allobaculum, Lactobacillus and Streptococcus. Bars represent the mean ± S.D. from three independent experiments. *p < 0.05, **p < 0.01
Discussion
Acute pancreatitis (AP) caused by hypertriglyceridemia (HTG-AP) is usually associated with a repeated attack of AP [25]. HTG can be divided into primary and secondary types. Primary HTG is caused by a high-fat, high-carbohydrate diet and other genetic and environmental factors, as well as lack of physical activity which can lead to disorders of TG synthesis and metabolism [26]. Secondary HTG is usually induced by unrecognized diseases, including obesity, diabetes, pregnancy, metabolic syndrome, and drugs such as estrogen and tamoxifen can also lead to the occurrence of HTG [6]. Studies have shown that the pathogenesis of HTG-AP is related to the inflammatory response, [27] microcirculatory disorder, [28] Ca2+ overload and endoplasmic reticulum stress, [29, 30] oxidative stress [31] and accumulation of free fatty acid [32]. A retrospective analysis showed that HTG-AP patients are generally younger than AP, and are more likely to suffer from cardiopulmonary and renal insufficiency and systemic inflammatory response syndrome (SIRS) [7]. There are some effective treatments on HTG-AP, such as insulin, heparin, plasmapheresis, and anti-HTG drugs. The chemopreventive effects of resveratrol in a rat model of cerulein-induced AP have been reported [33], however, whether resveratrol has a protective effect on HTG-AP is unknown. In our study, we successfully established the HTG-AP model according to the reference [20], finding that HTG-AP was more serious than AP, and proved that the resveratrol pre-treatment had a preventive effect on HTG-AP.
NF-κB transcription factor plays an important role in inflammation, immune response, survival and apoptosis [34]. This pathway regulates the production of pro-inflammatory cytokines, the aggregation of inflammatory cells, and the promotion of inflammatory response. Many studies have shown that the NF-κB pathway is involved in the inflammatory process and cancer development. For example, Th17 type cytokines, IL-6 and TNF-α can synergistically activate STAT3 and NF-κB pathways to promote the growth of colorectal cancer cells [35]. In addition, STAT3 and NF-κB pathways are also active in pancreatic cancer [36]. Clinical evidence shows that NF-κB pathway components play an important role in tumorigenesis and development, regulating gene expression related to cell survival and proliferation, drug resistance, metastasis, and angiogenesis [37]. Therefore, NF-κB can be used as a molecular target for some cancers.
Nowadays, several studies have been identified that some phytochemicals have inhibitory effects on the NF-κB pathway [38]. Among the polyphenols, resveratrol, curcumin, epigallocatechin gallate, genistein and cardamom have been the most well-studied. They have the ability to block NF-κB nuclear transport or restrain NF-κB activation to inhibit the proliferation ability of cancer cells. For example, resveratrol can treat glioblastoma multiforme by inhibiting PI3K/Akt/NF-κB signal transduction and inhibiting MMP-2 expression [39]; curcumin has a therapeutic effect on oxaliplatin-resistant colon cancer cell lines by inhibiting CXC chemokine/NF-κB signaling pathway [40]; Epigallocatechin gallate (EGCG) can treat nasopharyngeal carcinoma via regulating the cellular localization of NF-κB p65 and reducing the transcriptional regulation effect of NF-κB p65 on Twist1 expression [41].
Gut microbiota is closely associated with lipid metabolism disorder and systemic inflammation of obese mice [42]. Currently, Firmicutes, Bacteroides, Actinobacteria and Proteobacteria accounted for more than 90% of the gut microbiota [43]. Studies have shown that long-term HFD changed the gut microbiota, leading to increased intestinal permeability, mucosal immune response, obesity and chronic inflammation [44]. The prevalence of Firmicutes, Actinobacteria and Proteobacteria is positively associated with HFD, whereas Bacteroides show the opposite effect. Obesity and obesity-related pathologies are related to the occurrence of chronic low-grade inflammation [45]. Most patients with obesity exhibit increased circulating levels of inflammatory markers such as IL-6, IL-1, TNF and MCP1 [46]. As we know, some gut microbiota plays a role in the pro-inflammatory effect and others have an anti-inflammatory effect. Lactobacillus is a well-known probiotic that is proven to be related to reducing colitis in several models of inflammatory bowel diseases [47]. But the prevalence of Allobaculum level has been shown to be associated with neuronal and intestinal inflammation [48]. Streptococcus pneumonia is the most common type of streptococcus, and it can cause diseases such as pneumonia, meningitis and otitis media [49]. It has been reported in the HFD mice that the improvement of resveratrol on gut microbiota [50]. Indeed, the resveratrol-treated HFD group can up-regulate the relative abundance of anti-inflammatory Lactobacillus but down-regulate the relative abundance of pro-inflammatory Allobaculum and streptococcus compared with the HFD group. Similar results were observed in the chow and chow. Rev. group. These results demonstrated that resveratrol pre-treatment improved the gut microbiota in the HFD mice, thereby reducing the pancreatic damage following induced by caerulein.
In this study, the HFD mice model and HFD + cerulein (HTG-AP) mice model were established to measure the effect of resveratrol. In the HFD mice model, the expression levels of inflammatory and chemotactic cytokines TNF-α, LPS, IL-6, MCP-1 and the damage of pancreatic tissue were decreased, and the composition of gut microbiota was different. Also, in the HTG-AP mice model, resveratrol pre-treatment decreased the expressions of TNF-α, MCP-1, MDA the injury of pancreatic tissue, increased the levels of SOD and T-AOC. The results indicated that resveratrol pre-treatment can inhibit the secretion of pro-inflammatory cytokines and promote the antioxidant stress capacity. Furthermore, the NF-κB signaling pathway was inhibited by resveratrol, and the expressions of TNF-α and IL-6 related to the NF-κB signaling pathway activation were also decreased. All these results suggested that resveratrol pre-treatment can reduce oxidative stress, attenuate pancreatic tissue injury and restrain inflammatory response by inhibiting the NF-κB signaling pathway.
Conclusion
Using in vivo experiments, we provided evidence that HTG could promote AP injury, aggravating inflammation and oxidative stress. Moreover, the resveratrol pre-treatment could attenuate pancreas injury, inflammation, and oxidative stress in the HTG-AP mice, via restraining the NF-κB signaling pathway and regulating gut microbiota. Our study proved that the short-term resveratrol pre-treatment had a preventive effect on HTG-AP. However, the effects of long-term resveratrol pre-treatment and treatment on HTG-AP need more research.
Availability of data and materials
The initial data used to support the findings of this study are available from the corresponding author upon request. The sequencing data used in this study are stored in NCBI SRA database (SUB10626305). https://www.ncbi.nlm.nih.gov/sra/PRJNA778434.
References
Yadav D, Lowenfels AB. The epidemiology of pancreatitis and pancreatic Cancer. Gastroenterology. 2013;144:1252-1261. 5D5. https://doi.org/10.1053/j.gastro.2013.01.068.
Kota SK, Kota SK, Jammula S, Krishna SV, Modi KD. Hypertriglyceridemia-induced recurrent acute pancreatitis: A case-based review. Indian J Endocrinol Metab. 2012;16:141–3. https://doi.org/10.4103/2230-8210.91211.
Forsmark CE, Baillie J. AGA Institute technical review on acute pancreatitis. Gastroenterology. 2007;132:2022-2044. 5D5. https://doi.org/10.1053/j.gastro.2007.03.065.
UK guidelines for the management of acute pancreatitis. Gut 2005; 54 Suppl 3: iii1–9. 5D5: https://doi.org/10.1136/gut.2004.057026.
Valdivielso P, Ramírez-Bueno A, Ewald N. Current knowledge of hypertriglyceridemic pancreatitis. Eur J Intern Med. 2014;25:689–94. https://doi.org/10.1016/j.ejim.2014.08.008.
Adiamah A, Psaltis E, Crook M, Lobo DN. A systematic review of the epidemiology, pathophysiology and current management of hyperlipidaemic pancreatitis. Clin Nutr. 2018;37:1810–22. https://doi.org/10.1016/j.clnu.2017.09.028.
Bosques-Padilla FJ, Vázquez-Elizondo G, González-Santiago O, Del Follo-Martínez L, González OP, González-González JA, et al. Hypertriglyceridemia-induced pancreatitis and risk of persistent systemic inflammatory response syndrome. Am J Med Sci. 2015;349:206–11. https://doi.org/10.1097/maj.0000000000000392.
Zeng Y, Zhang W, Lu Y, Huang C, Wang X. Impact of hypertriglyceridemia on the outcome of acute biliary pancreatitis. Am J Med Sci. 2014;348:399–402. https://doi.org/10.1097/maj.0000000000000333.
Gambini J, Inglés M, Olaso G, Lopez-Grueso R, Bonet-Costa V, Gimeno-Mallench L, et al. Properties of Resveratrol: In Vitro and In Vivo Studies about Metabolism, Bioavailability, and Biological Effects in Animal Models and Humans. Oxidative Med Cell Longev. 2015;2015:837042. https://doi.org/10.1155/2015/837042.
Liang Q, Wang XP, Chen TS. Resveratrol protects rabbit articular chondrocyte against sodium nitroprusside-induced apoptosis via scavenging ROS. Apoptosis. 2014;19:1354–63. https://doi.org/10.1007/s10495-014-1012-1.
Li YR, Li S, Lin CC. Effect of resveratrol and pterostilbene on aging and longevity. Biofactors. 2018;44:69–82. https://doi.org/10.1002/biof.1400.
Byun EB, Sung NY, Park JN, Yang MS, Park SH, Byun EH. Gamma-irradiated resveratrol negatively regulates LPS-induced MAPK and NF-κB signaling through TLR4 in macrophages. Int Immunopharmacol. 2015;25:249–59. https://doi.org/10.1016/j.intimp.2015.02.015.
Zordoky BN, Robertson IM, Dyck JR. Preclinical and clinical evidence for the role of resveratrol in the treatment of cardiovascular diseases. Biochim Biophys Acta. 2015;1852:1155–77. https://doi.org/10.1016/j.bbadis.2014.10.016.
Szkudelski T, Szkudelska K. Resveratrol and diabetes: from animal to human studies. Biochim Biophys Acta. 2015;1852:1145–54. https://doi.org/10.1016/j.bbadis.2014.10.013.
de Ligt M, Timmers S, Schrauwen P. Resveratrol and obesity: Can resveratrol relieve metabolic disturbances? Biochim Biophys Acta. 2015;1852:1137–44. https://doi.org/10.1016/j.bbadis.2014.11.012.
Singh CK, Ndiaye MA, Ahmad N. Resveratrol and cancer: Challenges for clinical translation. Biochim Biophys Acta. 2015;1852:1178–85. https://doi.org/10.1016/j.bbadis.2014.11.004.
Dao TM, Waget A, Klopp P, Serino M, Vachoux C, Pechere L, et al. Resveratrol increases glucose induced GLP-1 secretion in mice: a mechanism which contributes to the glycemic control. PLoS One. 2011;6:e20700. https://doi.org/10.1371/journal.pone.0020700.
Shao D, Wang Y, Huang Q, Shi J, Yang H, Pan Z, et al. Cholesterol-Lowering Effects and Mechanisms in View of Bile Acid Pathway of Resveratrol and Resveratrol Glucuronides. J Food Sci. 2016;81:H2841–h2848. https://doi.org/10.1111/1750-3841.13528.
Andrade JM, Paraíso AF, de Oliveira MV, Martins AM, Neto JF, Guimarães AL, et al. Resveratrol attenuates hepatic steatosis in high-fat fed mice by decreasing lipogenesis and inflammation. Nutrition. 2014;30:915–9. https://doi.org/10.1016/j.nut.2013.11.016.
Zheng J, Wu J, Chen J, Liu J, Lu Y, Huang C, et al. Therapeutic effects of quercetin on early inflammation in hypertriglyceridemia-related acute pancreatitis and its mechanism. Pancreatology. 2016;16:200–10. https://doi.org/10.1016/j.pan.2016.01.005.
Bhatia M, Brady M, Kang YK, Costello E, Newton DJ, Christmas SE, et al. MCP-1 but not CINC synthesis is increased in rat pancreatic acini in response to cerulein hyperstimulation. Am J Physiol Gastrointest Liver Physiol. 2002;282:G77–85. https://doi.org/10.1152/ajpgi.00031x.2002.
Ye B, Rong P, Liu L, Wang W, Zhang S. Protective effect of necrostatin-1 on the damage of pancreas islet cells induced by TNF-α. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 2020;45:752–8. https://doi.org/10.11817/j.issn.1672-7347.2020.190534.
Zhang M, Wang C, Wu J, Ha X, Deng Y, Zhang X, et al. The effect and mechanism of KLF7 in the TLR4/NF-κB/IL-6 inflammatory signal pathway of adipocytes. Mediat Inflamm. 2018;2018:1756494. https://doi.org/10.1155/2018/1756494.
Nakajima S, Chi Y, Gao K, Kono K, Yao J. eIF2α-Independent Inhibition of TNF-α-Triggered NF-κB Activation by Salubrinal. Biol Pharm Bull. 2015;38:1368–74. https://doi.org/10.1248/bpb.b15-00312.
Kilinc F, Senates E, Demircan F, Pekkolay Z, Gozel N, Guven M, et al. Are There Differences in the Management of Acute Pancreatitis Cases Due to Severe Hypertriglyceridemia in Pregnant Women? Med Sci Monit. 2018;24:5619–23. https://doi.org/10.12659/msm.910343.
Shah AS, Wilson DP. Primary hypertriglyceridemia in children and adolescents. J Clin Lipidol. 2015;9:S20–8. https://doi.org/10.1016/j.jacl.2015.04.004.
Khatua B, El-Kurdi B, Singh VP. Obesity and pancreatitis. Curr Opin Gastroenterol. 2017;33:374–82. https://doi.org/10.1097/mog.0000000000000386.
Yan MX, Li YQ, Meng M, Ren HB, Kou Y. Long-term high-fat diet induces pancreatic injuries via pancreatic microcirculatory disturbances and oxidative stress in rats with hyperlipidemia. Biochem Biophys Res Commun. 2006;347:192–9. https://doi.org/10.1016/j.bbrc.2006.06.063.
Zhang W, Zhao Y, Zeng Y, Yu X, Yao J, Zhao S, et al. Hyperlipidemic versus normal-lipid acute necrotic pancreatitis: proteomic analysis using an animal model. Pancreas. 2012;41:317–22. https://doi.org/10.1097/MPA.0b013e31822981fb.
Kanak MA, Shahbazov R, Yoshimatsu G, Levy MF, Lawrence MC, Naziruddin B. A small molecule inhibitor of NFκB blocks ER stress and the NLRP3 inflammasome and prevents progression of pancreatitis. J Gastroenterol. 2017;52:352–65. https://doi.org/10.1007/s00535-016-1238-5.
Pérez S, Pereda J, Sabater L, Sastre J. Redox signaling in acute pancreatitis. Redox Biol. 2015;5:1–14. https://doi.org/10.1016/j.redox.2015.01.014.
Hahn SJ, Park JH, Lee JH, Lee JK, Kim KA. Severe hypertriglyceridemia in diabetic ketoacidosis accompanied by acute pancreatitis: case report. J Korean Med Sci. 2010;25:1375–8. https://doi.org/10.3346/jkms.2010.25.9.1375.
Carrasco C, Holguín-Arévalo MS, Martín-Partido G, Rodríguez AB, Pariente JA. Chemopreventive effects of resveratrol in a rat model of cerulein-induced acute pancreatitis. Mol Cell Biochem. 2014;387:217–25. https://doi.org/10.1007/s11010-013-1887-0.
Girard S, Kadhim H, Roy M, Lavoie K, Brochu ME, Larouche A, et al. Role of perinatal inflammation in cerebral palsy. Pediatr Neurol. 2009;40:168–74. https://doi.org/10.1016/j.pediatrneurol.2008.09.016.
De Simone V, Franzè E, Ronchetti G, Colantoni A, Fantini MC, Di Fusco D, et al. Th17-type cytokines, IL-6 and TNF-α synergistically activate STAT3 and NF-kB to promote colorectal cancer cell growth. Oncogene. 2015;34:3493–503. https://doi.org/10.1038/onc.2014.286.
Zhang X, Ren D, Wu X, Lin X, Ye L, Lin C, et al. miR-1266 Contributes to Pancreatic Cancer Progression and Chemoresistance by the STAT3 and NF-κB Signaling Pathways. Mol Ther Nucleic Acids. 2018;11:142–58. https://doi.org/10.1016/j.omtn.2018.01.004.
Zhang Q, Lenardo MJ, Baltimore D. 30 Years of NF-κB: A Blossoming of Relevance to Human Pathobiology. Cell. 2017;168:37–57. https://doi.org/10.1016/j.cell.2016.12.012.
Shanmugam MK, Lee JH, Chai EZ, Kanchi MM, Kar S, Arfuso F, et al. Cancer prevention and therapy through the modulation of transcription factors by bioactive natural compounds. Semin Cancer Biol. 2016:40–1. https://doi.org/10.1016/j.semcancer.2016.03.005.
Jiao Y, Li H, Liu Y, Guo A, Xu X, Qu X, et al. Resveratrol Inhibits the Invasion of Glioblastoma-Initiating Cells via Down-Regulation of the PI3K/Akt/NF-κB Signaling Pathway. Nutrients. 2015;7:4383–402. https://doi.org/10.3390/nu7064383.
Ruiz de Porras V, Bystrup S, Martínez-Cardús A, Pluvinet R, Sumoy L, Howells L, et al. Curcumin mediates oxaliplatin-acquired resistance reversion in colorectal cancer cell lines through modulation of CXC-Chemokine/NF-κB signalling pathway. Sci Rep. 2016;6:24675. https://doi.org/10.1038/srep24675.
Li YJ, Wu SL, Lu SM, Chen F, Guo Y, Gan SM, et al. (−)-Epigallocatechin-3-gallate inhibits nasopharyngeal cancer stem cell self-renewal and migration and reverses the epithelial-mesenchymal transition via NF-κB p65 inactivation. Tumour Biol. 2015;36:2747–61. https://doi.org/10.1007/s13277-014-2899-4.
Sen T, Cawthon CR, Ihde BT, Hajnal A, DiLorenzo PM, de La Serre CB, et al. Diet-driven microbiota dysbiosis is associated with vagal remodeling and obesity. Physiol Behav. 2017;173:305–17. https://doi.org/10.1016/j.physbeh.2017.02.027.
Bibbò S, Ianiro G, Giorgio V, Scaldaferri F, Masucci L, Gasbarrini A, et al. The role of diet on gut microbiota composition. Eur Rev Med Pharmacol Sci. 2016;20:4742–9.
Luo Q, Cheng D, Huang C, Li Y, Lao C, Xia Y, et al. Improvement of colonic immune function with soy Isoflavones in high-fat diet-induced obese rats. Molecules. 2019;24:5D5. https://doi.org/10.3390/molecules24061139.
Gregor MF, Hotamisligil GS. Inflammatory mechanisms in obesity. Annu Rev Immunol. 2011;29:415–45. https://doi.org/10.1146/annurev-immunol-031210-101322.
Weisberg SP, McCann D, Desai M, Rosenbaum M, Leibel RL, Ferrante AW Jr. Obesity is associated with macrophage accumulation in adipose tissue. J Clin Invest. 2003;112:1796–808. https://doi.org/10.1172/jci19246.
Zhu Q, Gao R, Wu W, Qin H. The role of gut microbiota in the pathogenesis of colorectal cancer. Tumour Biol. 2013;34:1285–300. https://doi.org/10.1007/s13277-013-0684-4.
Alhasson F, Das S, Seth R, Dattaroy D, Chandrashekaran V, Ryan CN, et al. Altered gut microbiome in a mouse model of gulf war illness causes neuroinflammation and intestinal injury via leaky gut and TLR4 activation. PLoS One. 2017;12:e0172914. https://doi.org/10.1371/journal.pone.0172914.
Mitchell TJ. Streptococcus pneumoniae: infection, inflammation and disease. Adv Exp Med Biol. 2006;582:111–24. https://doi.org/10.1007/0-387-33026-7_10.
Qiao Y, Sun J, Xia S, Tang X, Shi Y, Le G. Effects of resveratrol on gut microbiota and fat storage in a mouse model with high-fat-induced obesity. Food Funct. 2014;5:1241–9. https://doi.org/10.1039/c3fo60630a.
Zhang X, Yang G. Preventive effect of resveratrol on Caerulein-induced acute pancreatitis in high-fat diet-feeding mice; 2021.
Acknowledgements
Funding
This work was supported by Key projects of Sichuan Education Department (14ZA0194) and Special project of science and technology strategic cooperation between city and University (18SXHZ0371).
Author information
Authors and Affiliations
Contributions
XYZ and GDY conceived and designed the experiments. XYZ, YLC, ZM, HYZ, and LYZ performed the experiments and analyzed the data. XYZ and GDY wrote the manuscript. The author(s) read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Approved by The Ethics Committee of North Sichuan Medical College (No 2022.03), all experiments were performed in accordance with relevant guidelines and regulations. All methods are reported in accordance with ARRIVE guidelines for the reporting of animal experiments.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, X., Yang, G., Chen, Y. et al. Resveratrol pre-treatment alleviated caerulein-induced acute pancreatitis in high-fat diet-feeding mice via suppressing the NF-κB proinflammatory signaling and improving the gut microbiota. BMC Complement Med Ther 22, 189 (2022). https://doi.org/10.1186/s12906-022-03664-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12906-022-03664-4