
Abstract
Background
Phenylalanine ammonia-lyase (PAL), as a key enzyme in the phenylalanine metabolism pathway in plants, plays an important role in the response to environmental stress. However, the PAL family responding to abiotic stress has not been fully characterized in rapeseed.
Results
In this study, we conducted a genome-wide study of PAL family, and analyzed their gene structure, gene duplication, conserved motifs, cis-acting elements and response to stress treatment. A total of 17 PALs were identified in the rapeseed genome. Based on phylogenetic analysis, the BnPALs were divided into four clades (I, II, IV, and V). The prediction of protein structure domain presented that all BnPAL members contained a conservative PAL domain. Promoter sequence analysis showed that the BnPALs contain many cis-acting elements related to hormone and stress responses, indicating that BnPALs are widely involved in various biological regulatory processes. The expression profile showed that the BnPALs were significantly induced under different stress treatments (NaCl, Na2CO3, AlCl3, and PEG), suggesting that BnPAL family played an important role in response to abiotic stress.
Conclusions
Taken together, our research results comprehensively characterized the BnPAL family, and provided a valuable reference for revealing the role of BnPALs in the regulation of abiotic stress responses in rapeseed.
Similar content being viewed by others

Introduction
Phenylpropane metabolic pathway is one of the three important secondary metabolic pathways of plants [1], through which plants directly or indirectly synthesize lignin, flavonoids, coumarins, alkaloids, and other substances containing phenylpropane skeleton [2]. As the key rate-limiting enzyme in the phenylpropane metabolism pathway, Phenylalanine ammonia-lyase (PAL; EC4.3.1.5) catalyzes the deamination of L-phenylalanine to produce trans-cinnamic acid [2, 3]. PAL is the first-rate limiting enzyme for phenylpropanoid metabolism, which regulates the anabolism of lignin, flavonoids, isoflavones and alkaloids [4]. In plants, there are many secondary metabolites synthesized through phenylpropane synthesis, such as anthocyanins, lignin, hormones and flavonoids, which play an important role in plant growth, development and adaptation to environmental stress [5]. For example, it was found that AtPAL1 and AtPAL2 are highly expressed during nitrogen stress and temperature fluctuations, leading to an accumulation of flavonoids in Arabidopsis [6]. Therefore, PAL plays an important role in plant resistance to environmental stresses.
PAL is widely found in various plants, and has been identified in various plants, such as Arabidopsis [7], Populus trichocarpa [8], rice (Oryza sativa) [9], walnut (Juglans regia) [10], Coleus (Solenostemon scutellarioides (L.) Codd) [4] and wheat (Triticum aestivum L.) [11]. In most higher plants, there are more than one PAL in their genomes. According to previous studies, the number of PALs is 4, 7, 9, 4, 12 and 14 in Arabidopsis [6], cucumber [12], rice (Oryza sativa) [9, 13], tobacco [14], walnut (Juglans regia) [10], and potato (Solanum tuberosum) [15], respectively. PAL protein is relatively conserved in plants. Although the quantity of PAL protein varies widely in different plants, their molecular weight is relatively stable, mainly between 275 and 330 kDa [16, 17]. PAL family contains multiple PALs, and each individual in the family exhibits a distinct expression pattern and a differential response to biotic and abiotic stresses [4].
Rapeseed (Brassica napus L.) is the second largest oilseed crop around the world possesses a complex genome. The growth and productivity of rapeseed are significantly influenced by the physiological, biochemical, and molecular levels under various abiotic stress conditions, such as soil salinization [18,19,20], soil acidification [21] and drought stress [22, 23]. So far, the PAL family and key PALs responding to abiotic stress have not been fully characterized in rapeseed. Here, we present a genome-wide study to identify the BnPALs in the rapeseed genome. To explore the structural diversity and evolution of BnPALs, we assessed their phylogenetic relationships, gene structure, conserved motifs and cis-acting elements in the promoter region. Additionally, we also demonstrated the expression profile of BnPALs treated with NaCl, Na2CO3, AlCl3 and PEG. These findings will not only deepen our understanding of the response of BnPALs to abiotic stress, but also facilitate further research on biological functions of the gene family and provide potential gene targets for high-yield breeding under various biotic and abiotic stresses in rapeseed.
Results
Identification and characterization of PALs in rapeseed
To identify PALs in rapeseed, four AtPAL protein sequences were used as queries for BlastP and HMM searches against the rapeseed genome “ZS11”. Consequently, 17 BnPALs possessing the PAL domain were identified (Table 1). Then, these genes are named as BnPAL1–BnPAL17. The BnPAL proteins above ranged from 210 to 754 amino acids in length (Table 1 and Supplementary file 1). Their relative molecular weights (MW) span from 22.43 kDa to 82.66 kDa. The theoretical isoelectric points (PI) of these proteins lie between 5.01 and 9.21, while their Aliphatic indices range from 73.71 to 97.18. The grand average of hydropathy (GRAVY) for all BnPALs is predicted to be in the range of -0.415 to -0.100, indicating that BnPAL proteins exhibit strong hydrophilicity. Subcellular localization predictions indicate that all BnPAL proteins are cytoplasmic.
Protein secondary structure predictions reveal that all BnPAL proteins comprise four structures: Alpha helix (Hh), Extended strand (Ee), Beta turn (Tt), and Random coil (Cc) (Supplementary file 2). Among the four structures, the proportion of Hh is the highest, ranging from 45.80 to 60.99%, followed by Cc and Ee in order, and the proportion of Tt is the lowest, ranging from 4.02 to 10.31%. Additionally, the 3D protein structures of BnPAL proteins were predicted using SWISS-MODEL. The results highlight that BnPAL6, BnPAL8 and BnPAL14 share high similarity, as do BnPAL7, BnPAL9, BnPAL13, BnPAL15, BnPAL16 and BnPAL17 (Fig. 1).

3D structure diagram of BnPAL proteins
Phylogenetic analysis of the PAL family
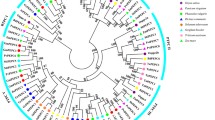
To elucidate the evolutionary relationships among BnPALs and other PAL proteins, we constructed a phylogenetic tree using multiple sequence alignments of PAL proteins from rapeseed (Brassica napus L.), Arabidopsis, and rice (Oryza sativa L.) (Fig. 2). The results showed that the PAL proteins were classified into five clades (I, II, III, IV, and V). Phylogenetic analysis results showed that 3, 4, 9, 5, and 9 PAL proteins were clustered into Clade I, Clade II, Clade III, Clade IV, and Clade V, respectively. Notably, all clades incorporated PAL proteins from both rapeseed and Arabidopsis, with the exception of Clade III, which exclusive featured PAL proteins from rice. This distribution underscores the close genetic relationship between PAL proteins from B. napus and Arabidopsis.

Phylogenetic tree of the PAL family in A. thaliana, O. sativa and B. napus. The neighbor-joining tree was generated through the MEGA11 program using the amino acid sequences of the PAL proteins by the neighbor-joining (NJ) method, with 1000 bootstrap replicates. The five major phylogenetic clades are labelled by different colored backgrounds
To further delve into the phylogenetic relationships of PALs within Brassica species, we constructed another phylogenetic tree based on PAL proteins from Brassica napus, Brassica rapa, and Brassica oleracea. This analysis revealed that these PAL proteins were grouped into four distinct clades (Fig. S1), aligning with prior classifications [10, 11]. Each of these clades encompassed PAL proteins from all three species, suggesting that BnPALs share a conserved evolutionary trajectory with PAL proteins from both Brassica rapa and Brassica oleracea.
Chromosomal locations and synteny evaluation of PALs
Chromosomal mapping showed that all the BnPALs are unevenly distributed across 9 identified chromosomes (Fig. 3). Notably, BnPAL16 and BnPAL17 were situated on unidentified chromosomes. BnPALs were mainly located on chromosomes A02, A04, A05, C04 and C07, which contained 2, 2, 2, 2 and 3 genes, respectively. There was only one BnPAL on A01, A07, C06 and C08 chromosomes, respectively, while no BnPAL was found on the rest chromosomes.

Distribution of BnPALs on chromosomes of B. napus.. The name of each chromosome is presented at the top of the corresponding green bar, and the gene names are given on the right side of them. The rules on the left indicate the physical position in megabases (Mb)
To reveal homologous gene functions and phylogenetic relationships between species, a collinearity analysis between PALs in B. napus, B. rapa and B. oleracea was performed. The results showed that the PALs of B. napus had 14 and 15 homologous gene pairs with the PALs of B. rapa and B. oleracea, respectively (Fig. 4 and Supplementary file 3). Among these homologous gene pairs, BnPAL15 had a collinear relationship with three PALs of B. rapa (BraA04g006280.3 C, BraA07g02160.3 C, and BraA09g044270.3 C) and two PALs of B. oleracea (BolC6t37294H and BolC8t50456H). Some BrPALs (BraA04g006280.3 C, BraA05g036420.3 C, BraA05g008320.3 C, BraA07g021160.3 C and BraA09g044270.3 C) and BoPALs (BolC4t22683H, BolC5t34439H, BolC6t37294H and BolC8t50456H) were found to be associated with two to four homologous gene pairs in B. napus, suggesting that PALs experience polyploidization in the process of evolution. In addition, we found that BnPAL8 and two PALs of B. oleracea showed a good collinear relationship. Meanwhile, no collinear genes of BnPAL5, BnPAL11 and BnPAL14 were detected in B. oleracea or B. rapa.

Synteny analysis of PALs among B. napus, B. rapa and B. oleracea.. The species names with the prefixes “B. napus”, “B. rapa” and “B. oleracea” indicate Brassica napus, Brassica rapa and Brassica oleracea, respectively. Grey lines in the background are the duplication events among B. napus, B. rapa and B. oleracea genomes, while the red lines indicate the syntenic PAL gene pairs. Green, yellow and blue bars represent the chromosomes of B. rapa, B. napus and B. oleracea, respectively. The chromosome number is labelled at the top of each chromosome
Gene duplication of BnPALs in B. napus
To better understand the evolutionary relationship of BnPALs in B. napus, gene duplication events were analyzed. The results showed that nine segmental duplication events with seven BnPALs in the genome of B. napus were identified, which were located on duplicated segments on chromosomes A01, A04, A05, A07, C06, C07, and C08 (Fig. 5). Among them, BnPAL15 had the most collinearity relationship with other BnPALs (BnPAL1, BnPAL6, BnPAL8, and BnPAL11), while BnPAL2, BnPAL3, BnPAL4, BnPAL9, BnPAL10, and BnPAL14 had no collinearity relationship with other BnPALs (Fig. 5 and Supplementary file 4).

Collinearity analysis between BnPALs. The gray lines indicate all synteny blocks between each chromosome and the thick red lines indicate duplicated PAL pairs. The circles from inside to outside represent gene density, unknown base ratio, GC ratio, GC skew and the length of chromosome (Mb), respectively. The chromosome name is shown at the bottom of each chromosome. The name and location of BnPALs are marked on the respective chromosome
Analysis of motifs and conserved domain of BnPAL proteins
To delve into the conservation and evolutionary relationships of PALs in B. napus, we analyzed conserved motifs using the MEME program. According to the results, a total of ten conserved motifs, designated as motif 1 through 10, were detected in the B. napus PAL protein family (Fig. 6A and B). Most PAL proteins contained motifs 4, 6, 8, and 9, suggesting that the BnPALs may encode proteins with similar functions. In clade II, the BnPAL proteins contained all the motifs, and these motifs were arranged in the same order (Fig. 6A and B), indicating that these PAL proteins may have similar biological functions. It is worth noting that different motifs had also been observed in the same clade, indicating that functional differentiation exists in the same clade. In particular, BnPAL3, BnPAL5 and BnPAL11 contained a considerable small number of motifs, suggesting a potential loss of function or functional differentiation in these genes. To perform the protein conserved domain analysis, the BnPAL protein sequences were subjected to NCBI for cdd-search, the results showed that all BnPAL proteins possess a Lyase ̲ aromatic domain (Fig. 6C). For most BnPAL proteins, this conserved domain is centrally situated.

Distribution of motifs and conserved domain in BnPAL proteins. (A) Groups of PALs in B. napus. (B) Schematic diagrams of all motifs in BnPAL proteins. Grey lines represent amino acid sequences. Different motifs are annotated by boxes of different colors and numbered 1–10. (C) Analysis of conserved domain of BnPAL proteins. The blue box represents conserved domain
Analysis of BnPAL gene structure and cis -acting elements in promoters
To explore the function of PALs in plant defense and abiotic stress responses in B. napus, a cis-acting element analysis was performed in the 2.0 kb promoter region of BnPALs. The cis-acting elements in the promoter region of BnPALs were identified using PlantCARE, and the kind with position of all cis-acting elements were marked with different color boxes (Fig. 7A and B). According to the results, the cis-acting elements could be mainly divided into 14 categories: abscisic acid responsiveness, anaerobic induction, auxin responsiveness, cell cycle regulation, defense & stress responsiveness, drought inducibility, endosperm expression, gibberellin responsiveness, light responsiveness, low-temperature responsiveness, MeJA responsiveness, meristem expression, salicylic acid responsiveness and zein metabolism regulation. The results showed that all the BnPALs, except BnPAL13, contained at least one of these cis-acting elements. Some BnPALs in the same clade had identical distribution of cis-acting elements in their promoter region. For example, BnPAL7 and BnPAL16 belonging to clade II, as well as BnPAL8 and BnPAL17 belonging to clade V, had the same cis-acting elements in their promoter regions (Fig. 7A and B). Additionally, we also found that most of the BnPALs contained cis-acting elements involved in anaerobic induction, abscisic acid, MeJA and defense & stress responsiveness, indicating that BnPALs may be induced or suppressed by various biotic and abiotic stresses.

Gene structure and cis -acting elements in the promoters of B. napus PALs. (A) Phylogenetic tree of PALs in B. napus. (B) Distribution of cis-acting elements in the promoter regions of BnPALs. Different cis-acting elements are annotated by boxes of different colors. (C) Gene structure of BnPALs. The purple box, blue green box and horizontal line represent UTRs, exons and introns, respectively
To gain insights into the structural diversity and evolutionary trajectory of BnPALs, the gene structure of BnPALs was analyzed. According to the results, the number of introns in BnPALs varied, ranging from one to four (Fig. 7C). BnPALs within the same clade exhibited similar gene structures. For instance, all BnPALs in clade II contained three exons and two introns, with the lengths of introns and exons being relatively consistent. Seven BnPALs (BnPAL6, BnPAL8, BnPAL9, BnPAL13, BnPAL14, BnPAL15 and BnPAL17) from clade IV and V displayed analogous gene structure, having the same number of exons and introns. Conversely, BnPALs in clade I varied in their number of introns and exons, suggesting diverse gene structures within this group.
Expression profile of BnPAL under abiotic stress conditions
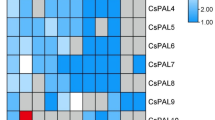
To further investigate the expression patterns of BnPALs under various stress conditions, qRT-PCR was performed to analysis on these genes under NaCl (1.2%), Na2CO3 (0.2%), AlCl3 (0.5mM) and PEG (PEG6000, 20%) stress conditions for 6 and 24 h (Fig. 8). Except for BnPAL10, which could not be detected by expression analysis even with five pairs of specific primers, qRT-PCR analysis revealed that the majority of the BnPALs were up-regulated (> 2-fold change) under at least one stress condition (Fig. 8). These results suggested that most of the BnPALs detected in this study exhibited significant responses to exogenous stressors. Most of the BnPALs up-regulated by Na2CO3 stress reached their peak expression at 6 h after treatment, indicating a rapid response to Na2CO3 stress. Four genes (BnPAL1, BnPAL11, BnPAL14 and BnPAL15) were highly up-regulated (≥ 5-fold change) under AlCl3 stress, suggesting that they have roles in AlCl3 stress response. We also found that the expression level of two (BnPAL1 and BnPAL11) and one (BnPAL14) gene under AlCl3 and Na2CO3 stress, respectively, was more than 10 times higher than that of control. Additionally, BnPAL14 was highly induced in response to all the stress treatments (> 5-fold change) (Fig. 8). According to the results, BnPAL9 was up-regulated under Na2CO3 stress but down-regulated under AlCl3 and PEG stresses, suggesting that BnPAL9 might play different roles in response to multiple stresses.

Expression patterns of BnPALs under various abiotic stresses. The y-axis represents relative expression, calculated using the 2−ΔΔCt formula. The x-axis represents different stress treatments. Expression profiles of BnPALs were obtained under NaCl (1.2%), Na2CO3 (0.2%), AlCl3 (0.5mM) and PEG (PEG6000, 20%) stress conditions, respectively. Samples were collected at 6 and 24 h after stress treatments. Data represent the mean ± standard error for three biological experiments. Student’s t-test was used to determine differences. *, P < 0.05, **P < 0.01
Discussion
Phenylalanine ammonia lyase (PAL) is an important enzyme involved in the phenylpropane pathway [7, 24]. Recently, there have been numerous reports on the analysis of PAL family in various species [9, 10, 13,14,15, 25,26,27,28,29,30,31]. However, the knowledge of PALs in rapeseed (B. napus), an important oil crop around the world, is still limited. In this study, a genome-wide analysis of PALs in B. napus was performed, and a total of 17 BnPALs were identified. Phylogenetic tree showed that BnPALs can be divided into four clades, which was consistent with the results of PAL family analysis in wheat [11]. Previous studies have shown that most PAL proteins are located in the cytoplasm [10, 11, 15, 32]. Similarly, the PAL proteins in B. napus were also predicted to be located in the cytoplasm (Table 1), suggesting that the subcellular localization of PAL protein in different species is conservative. The secondary structure prediction revealed that BnPAL predominantly consisted of alpha helices and random coils (Supplementary file 2), which was consistent with the results of PAL in other species [4, 33]. Taken together, the PAL family is relatively conservative in protein structure and gene evolution.
Compared to Arabidopsis thaliana, Brassica species including B. rapa and B. oleracea, have experienced a whole genome triplication (WGT) event during their evolutionary process [34]. Brassica napus (B. napus), an allotetraploid plant, was derived from natural hybridization between B. rapa and B. oleracea [35]. Therefore, there is a close genetic relationship between these species. Phylogenetic analysis showed that PALs in Arabidopsis and B. napus could be divided into four clades, each of which contained both AtPALs and BnPALs (Fig. 2). Meanwhile, the PALs in rice were divided into a single clade, exhibiting a relatively distant genetic relationship with PALs in Arabidopsis and B. napus. In addition, phylogenetic analysis among Brassica species showed that there is a relatively conservative evolutionary relationship among PALs from B. napus, B. rapa and B. oleracea. These results were consistent with the evolutionary relationship among these species.
Since B. napus originated from B. rapa and B. oleracea which have undergone WGT, theoretically, the number of genes in B. rapa and B. oleracea should be more than twice that of Arabidopsis, and the number of genes in B. napus should be more than five times that of Arabidopsis. However, the number of PALs in B. rapa, B. oleracea and B. napus is lower than expected (Fig. 4 and Supplementary file 3). These results suggest that part of PALs in B. rapa and B. oleracea were lost after WGT event. Notably, the number of PALs in B. napus is very close to the sum of PALs in B. rapa and B. oleracea. Additionally, collinearity analysis showed that almost every BnPAL can correspond to both BrPAL and BoPAL, indicating that most BnPALs were inherited from their ancestors (B. rapa and B. oleracea) during hybridization event.
In our study, BnPALs are widely dispersed across the genome, similar to PAL family studies of most other plants [7, 10, 11, 25]. According to the chromosome mapping results, nearly half of BnPALs are located in the A genome, while the other half are located in the C genome (Fig. 3; Table 1). The results of gene duplication analysis showed that most of the duplicated genes corresponding to BnPALs in B. napus genome A were located in genome C (Fig. 5), indicating that the BnPAL family expanded in B. napus genome mainly through dispersive duplication.
Recently, many studies have shown that PALs are involved in responding to various biotic and abiotic stresses [4, 6, 9, 15]. The cis-acting elements distributed in the gene promoter region have been shown to affect plant growth and development, environmental adaptation, and stress resistance [36,37,38,39]. Analysis of promoter cis-acting elements of the BnPALs is conducive to further exploring potential functions. In this study, we analyzed the promoter components of 17 BnPALs and identified various cis-acting elements. Among these cis-acting elements, most genes contained multiple core components that are involved in abscisic acid responsiveness, anaerobic induction, defense and stress responsiveness, drought inducibility and MeJA responsiveness (Fig. 7), indicating that BnPALs may play a crucial role in the response to a variety of biotic and abiotic stresses. The promoters of BnPALs classified in the same clade seem to contain similar types and numbers of cis-acting elements, suggesting that the BnPALs in the same clade may have functional similarities. Furthermore, some of the BnPALs in the same clade exhibited significant differences in the type and number of promoter components compared to other genes (Fig. 7). Therefore, BnPALs in the same clade may also have functional differentiation.
Since PAL plays an important role in plant resistance to environmental stresses, the expression of PALs is often induced when plants encounter biotic and abiotic stresses. According to previous studies, PALs are induced under cold stress in walnut [40], drought stress in cucumber [12] and lotus [33], high temperature stress in potato [15] and fungus stress in wheat [11]. In our study, the expression level of multiple BnPALs increased during various stress stimulations (Fig. 8), indicating that PAL gene family in B. napus has extensive responses to different stresses. In addition, the expression profiles of nearly all BnPALs fluctuated under AlCl3 exposure, suggesting a pronounced sensitivity of BnPALs to Al3+ stress. Intriguingly, while BnPAL1, BnPAL11, and BnPAL15 displayed markedly elevated expression compared to the control, BnPAL9’s expression dipped significantly below control levels at 24 h under Al3+ stress. This hints at potential functional redundancy and differentiation among these homologous genes. The expression level of BnPAL14 significantly increased under all stress treatments (Fig. 8), implying its multifaceted role in abiotic stress responses. Delving deeper into the biological functions of these genes will undoubtedly shed light on strategies for enhancing yield in rapeseed under various biotic stress conditions.
Conclusions
In this study, we conducted a comprehensive analysis of the PAL family in rapeseed. A total of 17 BnPALs were identified and classified into four clades. Further analysis of BnPALs including the phylogeny, gene structure, conserved motifs, chromosome localization, gene duplication and cis-acting elements provided insight into the molecular evolution of PAL family in rapeseed. The expression of most BnPALs increased significantly under stress treatment, indicating that the expression of PAL family in rapeseed was induced by abiotic stress. These results will contribute to a better understanding of the response of BnPALs to abiotic stress and lay the foundation for functional research of PALs in rapeseed.
Methods
Identification and characterization of PALs in rapeseed
As described in previous studies, we used BlastP and the Hidden Markov Model (HMM) [11] to identify PALs in the B. napus Zhongshuang 11 (hereafter referred to as ZS11) genome [41]. The genome database of B. napus ZS11 was downloaded from the BnPIR database (http://cbi.hzau.edu.cn/bnapus/index.php). Amino acid sequences of 4 AtPALs (AtPAL1, AtPAL2, AtPAL3 and AtPAL4) were obtained from The Arabidopsis Information Resource (TAIR) database (http://www.arabidopsis.org/). Then the full length of amino acid sequences of these AtPALs were subjected to BlastP against the B. napus genome with an e-value of 1 e− 10. Meanwhile, the HMM profile of the PAL domain (PF03634) was downloaded from the Pfam protein domain database (http://pfam.xfam.org/). Subsequently, the HMMER 3.1 software (http://www.hmmer.org/) was used to search the PALs with default parameters. Finally, combining the above two methods, we identified a total of 17 PALs in the rapeseed genome. The genome sequences of Brassica rapa (B. rapa) and Brassica oleracea (B. oleracea) were downloaded from BRAD database (https://brassicadb.cn/). Likewise, the PALs were also identified in B. rapa and B. oleracea genomes using the same method.
Structure, conserved motifs and physio-chemical properties of BnPAL proteins
The physico-chemical properties including molecular weight, theoretical PI, instability index, aliphatic index, and grand average of hydropathy (GRAVY) of BnPAL proteins were evaluated through ExPASy’s ProtParam tool (http://web.expasy.org/protparam/). The subcellular localization of BnPAL proteins was predicted by the WoLF PSORT (https://wolfpsort.hgc.jp/). The protein secondary structure analysis was performed by online tool SOPMA (http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html). The tertiary structure model of BnPAL protein was developed by SWISS-MODEL website (https://swissmodel.expasy.org/interactive). Gene structures including UTRs, introns and exons were shown by TBtools software (V 1.068; https://github.com/CJ-Chen/TBtools). The conserved motifs of BnPAL protein sequences were identified using the MEME program (https://meme-suite.org/meme/db/motifs) with default parameters [42]. The conserved motif structures were displayed by TBtools.
Phylogenetic analysis
To explore the evolutionary relationship of the PALs, a phylogenetic tree among B. napus, O. sativa, and A. thaliana were constructed. The sequence alignment was executed using Clustal W [43]. The phylogenetic tree was constructed using MEGA 11 software [44] with the neighbor-joining method and 1000 replicate iterations. The Interactive Tree Of Life (iTOL, https://itol.embl.de) was used to visualize the evolutionary tree [45].
Interspecies synteny analysis and gene duplication
To analyze the genetic relationships of PALs in different Cruciferae species, multiple sequence alignments were performed to detect the protein sequences of B. napus, B. rapa and B. oleracea with a similarity of more than 70%. Multiple Collinearity Scan Toolkit (MCScanX; https://github.com/wyp1125/MCScanX) was used to analyze the collinear region with default parameters [46]. Synteny analysis map of PALs among B. napus, B. oleracea and B. rapa was illustrated using the python-package JCVI (https://github.com/tanghaibao/jcvi). Gene duplication analysis was performed using the MCScanX program with default parameters, and the location and relationship of duplicated genes were displayed through Circos software [47].
Analysis of cis-acting elements in the BnPAL promoter
To investigate the putative cis-elements in the BnPAL promoter, a sequence of 2000 bp upstream of the initiation codon of the PAL in B. napus was selected as the regulatory promoter region. Then, these sequences were submitted to PlantCARE website (http://bioinformatics.psb.ugent.be/webtools/plantcare/html/) for cis-acting elements prediction [48], and the results were sorted and displayed by TBtools software [49].
Plant materials and growth conditions
Brassica napus ZS11 seeds were germinated on wet gauze (soaked with water) in a plant growth chamber at 20 to 22 °C and 65% humidity under a long-day condition (16-h-light/ 8-h-dark cycle). The one-week-old seedlings were then transferred into a previously described hydroponic system [19, 50] under the same culture conditions for nearly 20 days until the fourth leaves had extended. For stress treatment research, leaf samples from 4-week-old plants of ZS11 were collected after 6 and 24 h of 1.2% (w/v) NaCl, 0.2% (w/v) Na2CO3, 0.5 mM AlCl3 and 20% (w/v) PEG 6000 treatment. Seedlings without any stress treatment were used as the control. Each treatment includes three biological replications. Leaves were harvested immediately frozen in liquid nitrogen and stored at -80 °C for RNA extraction.
RNA extraction and quantitative real-time PCR
Total RNA was extracted using the RNA simple Total RNA kit (Tiangen Biotech, Beijing, China) according to the manufacturer’s protocol. cDNA was synthesized with 1 µg RNA from each sample with HiScript® II Q Select RT SuperMix with gDNA wiper (Vazyme, Nanjing, China). Gene-specific primers used for quantitative real-time PCR (qRT-PCR) listed in Supplementary file 5. qRT-PCR was run on the AriaMx real-time PCR system (Agilent Technologies). The following cycling parameters were used: initial denaturation at 95 °C for 5 min; 40 amplification cycles consisting of denaturation at 95 °C for 10s, annealing and extension at 60 °C for 30 s; The melting curve was then tested at 65–95 °C. The internal standard was the B. napus actin gene (BnaA01g27090D). Three biotic replicates were performed for each sample, and each replicate contained three technical replicates. Relative expression levels were calculated according to the 2−ΔΔCt method [51].
Data Availability
The PAL protein sequences of Arabidopsis were collected from the Arabidopsis information source (TAIR) database (http://www.arabidopsis.org). The genome sequences of B. napus PAL genes were downloaded from the BnPIR database (http://cbi.hzau.edu.cn/bnapus/index.php). All the datasets used and analyzed during the study are include in the article and its additional files.
References
Huang Y, Liang D, Xia H, Lin LJ, Wang J, Lv XL. Lignin and quercetin synthesis underlies Berry Russeting in ‘Sunshine Muscat’ grape. Biomolecules 2020, 10(5).
Raes J, Rohde A, Christensen JH, Van de Peer Y, Boerjan W. Genome-wide characterization of the lignification toolbox in Arabidopsis. Plant Physiol. 2003;133(3):1051–71.
MacDonald MJ, D’Cunha GB. A modern view of phenylalanine ammonia lyase. Biochem Cell Biol. 2007;85(3):273–82.
Zhu Q, Xie X, Lin H, Sui S, Shen R, Yang Z, Lu K, Li M, Liu YG. Isolation and functional characterization of a phenylalanine Ammonia-lyase gene (SsPAL1) from Coleus (Solenostemon scutellarioides (L.) Codd). Molecules. 2015;20(9):16833–51.
Blake VC, Kling JG, Hayes PM, Jannink J-L, Jillella SR, Lee J, Matthews DE, Chao S, Close TJ, Muehlbauer GJ, et al. The Hordeum Toolbox: the Barley Coordinated Agricultural Project genotype and phenotype resource. The Plant Genome. 2012;5(2):81–91.
Olsen KM, Lea US, Slimestad R, Verheul M, Lillo C. Differential expression of four Arabidopsis PAL genes; PAL1 and PAL2 have functional specialization in abiotic environmental-triggered flavonoid synthesis. J Plant Physiol. 2008;165(14):1491–9.
Huang J, Gu M, Lai Z, Fan B, Shi K, Zhou YH, Yu JQ, Chen Z. Functional analysis of the Arabidopsis PAL gene family in plant growth, development, and response to environmental stress. Plant Physiol. 2010;153(4):1526–38.
Shi R, Shuford CM, Wang JP, Sun YH, Yang Z, Chen HC, Tunlaya-Anukit S, Li Q, Liu J, Muddiman DC, et al. Regulation of phenylalanine ammonia-lyase (PAL) gene family in wood forming tissue of Populus trichocarpa. Planta. 2013;238(3):487–97.
Zeng JL, Ouyang LJ, Liu JL, He HH, Zhu CL, Peng XS, He XP, Fu JR, Chen XR, Bian JM, et al. Whole genome analysis and stress expression research of PAL gene in Rice. Genomics Appl Biol. 2018;37(9):3881–8.
Yan F, Li H, Zhao P. Genome-wide identification and transcriptional expression of the PAL Gene Family in Common Walnut (Juglans Regia L.). Genes (Basel) 2019, 10(1).
Zhan C, Li Y, Li H, Wang M, Gong S, Ma D, Li Y. Phylogenomic analysis of phenylalanine ammonia-lyase (PAL) multigene family and their differential expression analysis in wheat (Triticum aestivum L.) suggested their roles during different stress responses. Front Plant Sci. 2022;13:982457.
Shang QM, Li L, Dong CJ. Multiple tandem duplication of the phenylalanine ammonia-lyase genes in Cucumis sativus L. Planta. 2012;236:1093–105.
Hamberger B, Ellis M, Friedmann M, de Azevedo Souza C, Barbazuk B, Douglas CJ. Genome-wide analyses of phenylpropanoid-related genes in Populus trichocarpa, Arabidopsis thaliana, and Oryza sativa: the Populus lignin toolbox and conservation and diversification of angiosperm gene familiesThis article is one of a selection of papers published in the special issue on Poplar Research in Canada. Can J Bot. 2007;85(12):1182–201.
Reichert AI, He XZ, Dixon RA. Phenylalanine ammonia-lyase (PAL) from tobacco (Nicotiana tabacum): characterization of the four tobacco PAL genes and active heterotetrameric enzymes. Biochem J. 2009;424(2):233–42.
Mo F, Li L, Zhang C, Yang C, Chen G, Niu Y, Si J, Liu T, Sun X, Wang S et al. Genome-wide analysis and expression profiling of the phenylalanine Ammonia-lyase Gene Family in Solanum tuberosum. Int J Mol Sci 2022, 23(12).
Rawal HC, Singh NK, Sharma TR. Conservation, divergence, and genome-wide distribution of PAL and POX A gene families in plants. Int J Genomics. 2013;2013:678969.
Zhang X, Liu CJ. Multifaceted regulations of gateway enzyme phenylalanine ammonia-lyase in the biosynthesis of phenylpropanoids. Mol Plant. 2015;8(1):17–27.
Chen J, Zhang H, Tong J, Liu C, Ran J, Tang J, Liu J, Wen J, Zeng C, Wan H, et al. Genome-wide association analysis of root length traits in Brassica napus at germination stage under sodium carbonate stress. Euphytica. 2021;217(10):197.
Wan H, Chen L, Guo J, Li Q, Wen J, Yi B, Ma C, Tu J, Fu T, Shen J. Genome-wide Association Study reveals the Genetic Architecture Underlying Salt Tolerance-Related traits in rapeseed (Brassica napus L). Front Plant Sci. 2017;8:593.
Wan H, Qian J, Zhang H, Lu H, Li O, Li R, Yu Y, Wen J, Zhao L, Yi B et al. Combined Transcriptomics and Metabolomics Analysis reveals the molecular mechanism of Salt Tolerance of Huayouza 62, an Elite Cultivar in Rapeseed (Brassica napus L.). Int J Mol Sci 2022, 23(3).
Zhou H, Xiao X, Asjad A, Han D, Zheng W, Xiao G, Huang Y, Zhou Q. Integration of GWAS and transcriptome analyses to identify SNPs and candidate genes for aluminum tolerance in rapeseed (Brassica napus L). BMC Plant Biol. 2022;22(1):130.
Jamshidi N, Shirani R, Takht CF, Nazeri P, Ghafari M. Evaluation of rapeseed genotypes under drought stress condition. J Crop Ecophysiology. 2012;23(3):323–38.
Li J, Lin K, Zhang S, Wu J, Fang Y, Wang Y. Genome-wide analysis of myeloblastosis-related genes in Brassica napus L. and positive modulation of osmotic tolerance by BnMRD107. Front Plant Sci. 2021;12:678202.
Han H, Woeste KE, Hu Y, Dang M, Zhang T, Gao X-X, Zhou H, Feng X, Zhao G, Zhao P. Genetic diversity and population structure of common walnut (Juglans regia) in China based on EST-SSRs and the nuclear gene phenylalanine ammonia-lyase (PAL). Tree Genet Genomes 2016, 12(6).
Chang A, Lim MH, Lee SW, Robb EJ, Nazar RN. Tomato phenylalanine ammonia-lyase gene family, highly redundant but strongly underutilized. J Biol Chem. 2008;283(48):33591–601.
de Jong F, Hanley SJ, Beale MH, Karp A. Characterisation of the willow phenylalanine ammonia-lyase (PAL) gene family reveals expression differences compared with poplar. Phytochemistry. 2015;117:90–7.
Dong CJ, Shang QM. Genome-wide characterization of phenylalanine ammonia-lyase gene family in watermelon (Citrullus lanatus). Planta. 2013;238(1):35–49.
Fukasawa-Akada T, Kung SD, JC W. Phenylalanine ammonia-lyase gene structure, expression, and evolution in Nicotiana. Plant Mol Biol. 1996;30:711–22.
He Y, Zhong X, Jiang X, Cong H, Sun H, Qiao F. Characterisation, expression and functional analysis of PAL gene family in Cephalotaxus hainanensis. Plant Physiol Biochem. 2020;156:461–70.
Hou X, Shao F, Ma Y, Lu S. The phenylalanine ammonia-lyase gene family in Salvia miltiorrhiza: genome-wide characterization, molecular cloning and expression analysis. Mol Biol Rep. 2013;40(7):4301–10.
Kumar A, Ellis B. The phenylalanine Ammonia-lyase Gene Family in Raspberry. Structure, expression, and evolution. Plant Physiol 2001, 127(1).
Vishwakarma SK, Singh N, Kumaria S. Genome-wide identification and analysis of the PAL genes from the orchids Apostasia shenzhenica, Dendrobium catenatum and Phalaenopsis equestris. J Biomol Struct Dyn. 2023;41(4):1295–308.
Wu H, Gui S, Wang S, Ding Y. Molecular evolution and functional characterisation of an ancient phenylalanine ammonia-lyase gene (NnPAL1) from Nelumbo nucifera: novel insight into the evolution of the PAL family in angiosperms. BMC Evol Biol. 2014;14:100.
Cheng F, Wu J, Wang X. Genome triplication drove the diversification of Brassica plants. Hortic Res. 2014;1:14024.
Chalhoub B, Denoeud F, Liu S, Parkin IAP, Tang H, Wang X, Chiquet J, Belcram H, Tong C, Samans B, et al. Early allopolyploid evolution in the post-neolithic Brassica napus oilseed genome. Science. 2014;345(6199):950–3.
Himmelbach A, Liu L, Zierold U, Altschmied L, Maucher H, Beier F, Muller D, Hensel G, Heise A, Schutzendubel A, et al. Promoters of the barley germin-like GER4 gene cluster enable strong transgene expression in response to pathogen attack. Plant Cell. 2010;22(3):937–52.
Yamaguchi-Shinozaki K, Shinozaki K. A Nove1 cis-acting element in an Arabidopsis Gene 1s lnvolved in responsiveness to Drought, lowqemperature, or high-salt stress. Plant Cell. 1994;6:251–64.
Yamaguchi-Shinozaki K, Shinozaki K, TRANSCRIPTIONAL REGULATORY NETWORKS IN CELLULAR RESPONSES AND TOLERANCE TO DEHYDRATION AND COLD STRESSES. 2006, 57(1):781–803.
Ulmasov T, Murfett J, Hagen G, Guilfoyle TJ. Aux/lAA proteins repress expression of reporter genes containing natural and highly active synthetic auxin response elements. Plant Cell. 1997;9:1963–71.
Xu F, Deng G, Cheng S, Zhang W, Huang X, Li L, Cheng H, Rong X, Li J. Molecular cloning, characterization and expression of the phenylalanine ammonia-lyase gene from Juglans regia. Molecules. 2012;17(7):7810–23.
Song J, Guan Z, Hu J, Guo C, Yang Z, Wang S, Liu D, Wang B, Lu S, Zhou R, et al. Eight high-quality genomes reveal pan-genome architecture and ecotype differentiation of Brassica napus. Nat Plants. 2020;6(1):1–12.
Li X, Guo C, Ahmad S, Wang Q, Yu J, Liu C, Guo Y. Systematic analysis of MYB Family genes in Potato and their multiple roles in development and stress responses. Biomolecules 2019, 9(8).
Larkin MA, Blackshields G, Brown NP, Chenna R, McGettigan PA, McWilliam H, Valentin F, Wallace IM, Wilm A, Lopez R, et al. Clustal W and Clustal X version 2.0. Bioinformatics. 2007;23(21):2947–8.
Tamura K, Stecher G, Kumar S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol Biol Evol. 2021;38(7):3022–7.
Letunic I, Bork P. Interactive tree of life (iTOL) v4: recent updates and new developments. Nucleic Acids Res. 2019;47(W1):W256–9.
Wang Y, Tang H, Debarry JD, Tan X, Li J, Wang X, Lee TH, Jin H, Marler B, Guo H, et al. MCScanX: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012;40(7):e49.
Krzywinski M, Schein J, Birol I, Connors J, Gascoyne R, Horsman D, Jones SJ, Marra MA. Circos: an information aesthetic for comparative genomics. Genome Res. 2009;19(9):1639–45.
Lescot M, Déhais P, Thijs G, Marchal K, Moreau Y, Van de Peer Y, Rouzé P, Rombauts S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002;30(1):325–7.
Chen C, Chen H, Zhang Y, Thomas HR, Frank MH, He Y, Xia R. TBtools: an integrative Toolkit developed for interactive analyses of big Biological Data. Mol Plant. 2020;13(8):1194–202.
Tocquin P, Corbesier L, Havelange A, Pieltain A, Kurtem E, Bernier G, Périlleux C. A novel high efficiency, low maintenance, hydroponic system for synchronous growth and flowering of Arabidopsis thaliana. BMC Plant Biol 2003, 3(2).
Livak KJ, Schmittgen TJM. Analysis of relative gene expression data using real-time quantitative PCR and the 2-DDCt method. Methods. 2001;25(4):402–8.
Funding
This research was supported by the Natural Science Foundation of China (U22A20469), Hainan Provincial Natural Science Foundation of China (323QN192) and Research Initiation Fund of Hainan University (KYQD(ZR)-22103).
Author information
Authors and Affiliations
Contributions
X.Zhou. and H.W. conceived and designed the experiments; H.Zhang., J.H., Z.W., and X.Zhang. performed the main experiments; H.Zhao. performed the data collection and bioinformatics analysis; H.Zhang. and X.Zhang. wrote the manuscript; X.Zhou., G.Y. and H.W. revised the manuscript. All authors have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Zhang, H., Zhang, X., Zhao, H. et al. Genome-wide identification and expression analysis of phenylalanine ammonia-lyase (PAL) family in rapeseed (Brassica napus L.). BMC Plant Biol 23, 481 (2023). https://doi.org/10.1186/s12870-023-04472-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-023-04472-9