
Abstract
Background
Drought is one of the main environmental factors limiting plant growth and development. Pennisetum purpureum Schum. was used to explore the mitigation effects of exogenous strigolactone (SL) on drought stress during the seedling stage. The effects of different concentrations (1, 3, 5, and 7 μmol·L− 1) of SL on the photosynthesis characteristics, growth performance, and endogenous abscisic acid (ABA) of P. purpureum under drought stress were studied.
Results
Exogenous SL could effectively alleviate the inhibitory effect of drought stress on P. purpureum growth. Compared with drought stress, the net photosynthesis rate, stomatal conductance, transpiration rate, and water-use efficiency of the leaves of P. purpureum after SL treatment significantly increased, thereby exerting a significant mitigation effect on the decrease in photosystem II maximum photochemical efficiency and the performance index based on light absorption caused by drought. Moreover, the exogenous application of SL can effectively increase the fresh and dry weight of the leaves and roots and the main-root length. After applying SL for 120 h, the ABA content of P. purpureum decreased significantly. The activity of key enzymes of photosynthesis significantly increased after 48 h of external application of SL to P. purpureum.
Conclusions
SL treatment can improve the photosynthesis performance of P. purpureum leaves under drought conditions and increase the antioxidant capacity of the leaves, thereby reducing the adverse effects of drought, promoting the growth of P. purpureum, and effectively improving the drought resistance of P. purpureum.
Similar content being viewed by others

Background
Drought is one of the most important environmental factors affecting plant growth and development [1]. With the looming global climate change, the frequency and intensity of regional droughts are also increasing yearly [2]. On one hand, drought can reduce the photosynthesis rate of leaves and destroy plant photosynthesis organs, in turn leading to the inability of plants to effectively utilize absorbed light energy [3, 4]. On the other hand, drought can affect nutrient acquisition, transport, distribution, and storage, leading to decreased plant-biomass accumulation and root vigor [5, 6]. Plant hormones play a crucial role in regulating plant responses to abiotic stresses [7]. Many phytohormones including abscisic acid (ABA), ethylene (ET), salicylic acid (SA), and jasmonic acid (JA) are involved in the regulation of plant stomata and inducing the expression of genes related to stomatal aperture [7]. Hormones such as ABA, SA, and JA also play important roles in improving chlorophyll (Chl) content, relative water content, and proline content, as well as in increasing the fresh and dry weight of roots and above-ground parts [8]. These hormones are also involved in the scavenging of reactive oxygen species (ROS), maintaining the redox status of plants under stress conditions, and enhancing antioxidant defense responses [9]. Thus, phytohormones play an important role in regulating plant response to abiotic stresses.
Strigolactones (SLs), a class of hormones produced with carotenoids as precursors, affect plant growth and development by regulating various hormone-response pathways of plants and their growth to improve their adaptation to adverse stresses [10, 11]. SLs further reportedly inhibit plant branching, control shape root morphology, enhance primary root cell growth to inhibit adventitious root development and growth, and regulate plant secondary growth [12, 13]. They are also associated with drought resistance, cold tolerance, salt tolerance and other stress tolerance [14,15,16,17], and photomorphogenesis [18] and serve as positive regulators of plant responses to certain abiotic stresses [19]. Sattar et al. [20] applied SL to maize (Zea mays L.) under drought stress and found that SL improves the water relationship of maize seedlings, increases photosynthesis pigments and gas-exchange parameters, and improves antioxidant enzyme activity. Thus, the tolerance of maize seedlings to drought stress is enhanced. Ha et al. [21] found that Arabidopsis thaliana plants deficient in SL biosynthesis and signal transduction exhibit hypersensitivity to drought and salt stress. Treatment with exogenous SL restores the drought-sensitive phenotype of SL-deficient mutants, but not SL-responsive mutants, and enhances drought tolerance in wild-type (WT) plants. The role of SLs as a positive regulator in stress response has been confirmed. Min et al. [22] studied the foliar application of SL to alleviate drought stress in grapes (Vitis vinifera L.). They found that GR24 treatment increases the Chl content and Pn of plants and reduces the electrolyte leakage, ROS content, indole acetic acid and zeatin riboside contents in the roots and leaves under drought stress. Consequently, the adverse effects of drought are ameliorated.
Photosynthesis is one of the most important physiological processes that are inhibited in plants under drought stress. Studies have shown that SL can inhibit the activity of Chl-degrading enzymes, regulate the binding of Chl to membrane proteins, maintain the stability of the chloroplast thylakoid membrane, and continuously enhance the photosynthesis capacity [23]. SL can also increase the stomatal conductance (Gs), transpiration rate (Tr), and intercellular CO2 concentration (Ci) of plants under stress conditions, thereby increasing the Pn [16]. SL can ease the photosynthesis process as well by maintaining the stability of the photosystem II (PSII) supercomplex, increasing D1 protein turnover, the photosynthetic electron transport and the demand for ATP and NADPH in the Calvin cycle, and the efficiency of the photosystem, ultimately promoting plant growth and development [24,25,26]. Photosynthesis in higher plants can be classified as C3, C4, and crassulacean acid metabolism according to the way they fix carbon during the production of different initial photosynthesis products [27]. Among the many enzymes involved in the C4 photosynthesis pathway, phosphoenolpyruvate carboxylase (PEPC), pyruvate phosphate dikinase (PPDK), and NADP-malic enzyme (NADP-ME) are considered to be the most important [28,29,30]. The average Pn of rice (Oryza sativa L.) transduced with C4 model cereal (Setaria italica L.) PPDK and NADP-ME genes increases by 18 and 12%, respectively, and is positively correlated with increased photosynthesis pigment content [31]. Transgenic rice plants transfer more absorbed light energy to photochemical reactions than WT plants, and transgenic plants exhibit increased yield as evidenced by increased plant height, spike length, spike weight, and thousand grain weight [31]. This finding indicates that the activity of key photosynthesis enzymes is important for photosynthesis and plant yield.
Pennisetum purpureum Schum. is a perennial C4 plant with tall plants, well-developed root systems, strong tillering capacity, and huge biomass [32]. P. purpureum has few pests and diseases, fast growth, and high yield. It contains a large amount of crude protein and soluble sugar, which is a kind of high-yielding and high-quality forage grass with economic and ecological benefits for sustainable development. At present, studies on P. purpureum have focused on application and nutritional value [32,33,34], whereas the effects of plant growth regulators such as SL on drought stress in P. purpureum have not been reported. Accordingly, the present study was conducted to determine the effects of exogenous SL on the growth, photosynthesis characteristics, fresh weight, dry weight, main root length, endogenous abscisic acid (ABA), and key photosynthesis enzyme activities of P. purpureum under drought stress. We aimed to reveal the regulatory mechanism of exogenous SL application on P. purpureum in response to drought stress and provide a theoretical basis for the application and research of exogenous plant growth regulators such as SLs and thus improve the drought resistance of P. purpureum.
Results
Effect of SL on the photosynthesis parameters of leaves of P. purpureum under drought stress
Table 1 shows the net Pn and Tr of P. purpureum leaves, which continuously decreased with the severity of drought. Gs, Ci, and WUE showed a decreasing, increasing, and decreasing trend. These results all significantly differed from those of the normally watered control (P < 0.05). Under drought stress, the foliar application of SL to P. purpureum effectively alleviated the inhibitory effect caused by drought stress. After 24 h of hormone spraying, the Pn and Gs of the T1, T2, and T3 treatment groups were significantly elevated, with significant differences (P < 0.05) from the drought-treatment group (D group). The most significant relief effect was observed in T2, which was close to that of the normal watering group. The Tr of all hormone-treatment groups was significantly higher than that of the D group, with the most significant effect on T2 and T3. The Ci of all hormone-treatment groups was significantly higher than that of the D group. WUE significantly improved only in T1, which significantly differed from that of the D group (P < 0.05).
After 48 h of hormone spraying, the Pn, Gs, and Tr in T1, T2, and T3 were higher than those in the D group, whereas the Pn, Gs, and Tr in T4 were lower than those in the D group. Ci most significantly increased in T1, and WUE showed an inhibitory trend compared with the D group in all three groups except T3. This finding indicated that the relief effect of hormone on Tr was higher than that on Pn. After 120 h of hormone spraying, the Pn, Gs, Tr, and WUE were significantly higher in T1, T2, and T3 than those in the D group. Overall, T1 showed the most significant relief effect, and Ci showed a decreasing trend in all treatment groups.
Effect of SL on the physiological properties of Chl fluorescence in leaves of P. purpureum under drought stress
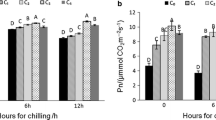
Figure 1 shows that the maximum photochemical efficiency of PSII (Fv/Fm) and the performance index based on the absorption of light energy (PIabs) of the leaves of P. purpureum under drought stress were significantly lower than those of the normal control. This finding indicated that the photosynthesis performance of P. purpureum was severely affected by photoinhibition under drought stress, and the photoinhibition became more serious with the severity of drought. The photoinhibition that occurred in PSII of P. purpureum was significantly alleviated by spraying SL on its leaves, and Fv/Fm and PIabs were significantly enhanced. After 24 h of hormone spraying, T1, T2, T3, and T4 showed significant alleviating effects compared with the D group, and the Fv/Fm values of T1, T2, and T4 were significantly higher than that in the D group. After 48 h of hormone spraying, T1, T2, T3, and T4 differed significantly (P < 0.05) compared with the D group. The Fv/Fm values of T2, T3, and T4 were significantly higher than those of the D group. After 24 and 48 h of hormone spraying, the PIabs of T2 differed significantly (P < 0.05) from that of the D group and reached the level of the normal control group. Therefore, the relief effect of T2 was the most significant. After 120 h of hormone spraying, the relief effect of T1, T2, T3, and T4 was significantly higher than that of the D group. The Fv/Fm values of each treatment group were significantly higher than that of the D group, and the relief effect of PIabs was the most obvious in T2.

Effect of spraying SL on the physiological characteristics of Chl fluorescence of P. purpureum leaves under drought stress. Effects of SL on Fv/Fm (a, b, and c) and PIabs (d, e, and f) of P. purpureum leaves. Different lowercase letters at the same time indicate significant differences (P < 0.05). N: normal watering treatment; D: drought stress treatment only; T1, T2, T3, and T4 represent the different treatment concentrations of SL under drought stress (1, 3, 5, and 7 μmol·L− 1 respectively)
Effect of SL on P. purpureum biomass under drought stress
As shown in Table 2, drought stress had a significant effect on the biomass of P. purpureum, and its leaf fresh weight and dry weight, root fresh weight and dry weight, and leaf and root water content were significantly lower than those of the normal watering-treatment group. The application of exogenous SL to P. purpureum under drought stress alleviated the accumulation of biomass in P. purpureum, but it was related to the SL concentration. After 24 h of hormone spraying, the relief effect was most significant for leaf fresh weight and dry weight in T2 and T3, for root fresh weight and dry weight in T1, and for leaf and root water content in T2. After 48 h of hormone spraying, the leaf fresh weight, dry weight, and leaf water content were most significantly relieved by T1, followed by T2. The root fresh weight and dry weight were most significantly relieved by T1 and T2. No significant differences were observed in root water content at this time. After 120 h of hormone spraying, the leaf fresh weight, dry weight, and leaf water content were most significantly relieved by T1, the root fresh weight and dry weight were most significantly relieved by T3, and the root water content was most significantly relieved by T2. Therefore, T1 and T2 had the most significant effect on the biomass of P. purpureum under drought stress.
Effect of SL on the root length of P. purpureum under drought stress
As shown in Fig. 2, the root length of the D group was significantly lower than that of the normal treatment group. After spraying SL, their root lengths significantly improved and differed significantly from those of the D group (P < 0.05). The most significant effects on the root system were observed in T2 at 24, 48, and 120 h after hormone spraying. The effect of T1 on the root system was better than that of T3 and T4 in the first 48 h. The effect of T3 and T4 on the root system was better than that of T1 after 120 h.

Effect of spraying SL on the root length of P. purpureum under drought stress. Effects of SL on root length (a, b, and c) of P. purpureum. Different lowercase letters at the same time indicate significant differences (P < 0.05). N: normal watering treatment; D: drought stress treatment only; T1, T2, T3, and T4 represent the different treatment concentrations of SL under drought stress (1, 3, 5, and 7 μmol·L− 1 respectively)
Effect of SL on the ABA content of P. purpureum under drought stress
Figure 3 shows that with prolonged drought time, the ABA content in P. purpureum leaves showed a continuous increasing trend, and that in P. purpureum roots showed an increasing, decreasing, and increasing trend. The ABA content of P. purpureum leaves under drought stress was initially lower and then became higher than that of the normal control group. Meanwhile, the ABA content of P. purpureum roots under drought stress was always higher than that of the normal control group. After spraying SL on P. purpureum under drought stress, the ABA content of leaves showed a continuous increase in all treatment groups, and the ABA content of the roots showed a continuous decrease. However, the ABA content in roots was always higher than that in leaves. After 24 h of hormone spraying, the ABA content in the leaves of all treatment groups was higher than that of the D group. The ABA content in the roots of T3 was higher than that of the D group, and the ABA content in the roots of the remaining three groups was lower than that of the D group. After 48 h of hormone spraying, the ABA content in the leaves of T1, T2, and T3 was higher than that of the D group. The ABA content in the leaves of T4 was lower than that of the D group. The ABA content in the roots of T3 was higher than that of the D group. The ABA content in the roots of T1, T2, and T4 was lower than that of the D group. After 120 h of hormone spraying, the ABA content in the leaves and roots of the treatment groups (all concentrations) was lower than that of the D group.

Effect of spraying SL on the ABA content of P. purpureum under drought stress. Effects of SL on leaf ABA content (a, b, and c) and root ABA content (d, e, and f) of P. purpureum. Different lowercase letters at the same time indicate significant differences (P < 0.05). N: normal watering treatment; D: drought stress treatment only; T1, T2, T3, and T4 represent the different treatment concentrations of SL under drought stress (1, 3, 5, and 7 μmol·L− 1 respectively)
Effect of SL on key photosynthesis enzyme activities in P. purpureum under drought stress
As shown in Fig. 4, the NADP-ME activity in the D group was significantly lower than that in the normal-watering group, the PEPC and PPDK activities were higher than those in the normal-watering group, and the PPDK activity differed significantly (P < 0.05) from that of the normal-watering group. The NADP-ME, PEPC, and PPDK activities were significantly higher in T2 and T3 after spraying SL onto P. purpureum under drought stress. The NADP-ME activity in T1 was higher than that in the D group, and the PEPC and PPDK activities were lower than those in the D group. The NADP-ME and PEPC activities were significantly higher in T4 than in the D group, and the PPDK activity did not change significantly compared with the D group.

Effect of spraying SL on the activities of key enzymes in the photosynthesis of P. purpureum under drought stress. Effects of SL on NADP-ME (a), PEPC (b), and PPDK (c) activities of P. purpureum. Different lowercase letters at the same time indicate significant differences (P < 0.05). N: normal watering treatment; D: drought stress treatment only; T1, T2, T3, and T4 represent the different treatment concentrations of SL under drought stress (1, 3, 5, and 7 μmol·L− 1 respectively)
Discussion
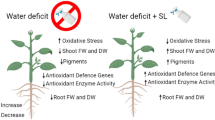
Drought is one of the major abiotic stress factors affecting plant growth. To cope with drought stress, plants have developed various self-protection and self-defense mechanisms during their long-term evolution [35, 36]. Plants respond to drought by regulating morphological and physiological characteristics such as root growth, leaf structure, stomatal movement, and photosynthesis. However, the ability of plants to adapt to drought by altering their own physiological and biochemical metabolism is limited and has significant effects only in the early stages of drought; with the severity of drought, the plant’s own metabolism becomes disturbed, eventually affecting plant growth and morphogenesis, resulting in reduced biomass accumulation [37, 38]. The results of this experimental study showed that under drought stress, the fresh weight, dry weight, and water content of the leaves and roots of P. purpureum were significantly lower than those of the normal-watering group. After foliar spraying of SL onto P. purpureum under drought stress, its dry and fresh weight and water content increased, and the growth inhibition caused by drought stress was alleviated, with the best alleviation effect observed in T1 and T2. This finding indicated that the application of SL promoted the growth of P. purpureum under drought conditions, which may be related to phytohormone interactions. More studies have shown that SL can interact with growth hormones and cytokinins to regulate plant biomass accumulation and promote plant growth and development [39, 40]. Sattar et al. [20] also showed that exogenous application of SL increases the dry weight and relative water content of maize in above- and below-ground under drought stress.
Drought stress causes the stomatal closure of plant leaves, causing a decrease in leaf Gs, which in turn reduces Tr to reduce water loss. However, stomatal closure reduces water loss while also reducing CO2 entry, decreasing the CO2 concentration in the leaves and leading to a continuous decrease in Pn, thereby inflicting more serious damage to plants with increased drought severity [41, 42]. In this experiment, drought stress caused a significant decrease in net Pn, Gs, Tr, and WUE in P. purpureum leaves, and Ci initially decreased and then increased. The inhibition of photosynthesis and reduction of Pn caused by drought stress can be attributed to stomatal and nonstomatal factors. If Gs and Ci decreased, the net Pn was primarily affected by stomatal factors. If Gs decreased but Ci increased, the net Pn was affected mostly by nonstomatal factors, indicating that photosynthesis was limited by stomatal and nonstomatal factors in this experiment [43,44,45]. The net Pn, Gs, Tr, and WUE of P. purpureum leaves were enhanced by the exogenous application of SL. Particularly, T1 and T2 had the most significant enhancement effect, whereas T4 had an insignificant alleviation effect, indicating that the low concentration of SL could alleviate the inhibitory effect of drought stress on P. purpureum, and enhance its photosynthesis capacity. This finding may be due to the fact that SL can regulate the binding of Chl to membrane proteins and thus maintain the stability of chloroplast cyst-like membranes, thereby enhancing the plant’s ability to use light energy to overcome photosystem damage and the photosynthesis performance of the plant [17]. Sedaghat et al. [46] obtained similar findings in their study on the effect of SL on wheat (Triticum aestivum L.) under drought stress, in which SL application enhances the net Pn, Gs, and Tr of wheat. Thula et al. [47] showed that the effect on photosynthesis is primarily attributed to SL compared with other plant hormones. In the present study, SL had a more pronounced effect on stomatal conductance. Stomatal conductance and transpiration rates were significantly higher in TI and T2 than in the D group at 24, 48, and 120 h. The effect of SL on stomatal conductance and transpiration rate was higher in T2 than in the D group. The opening of stomata led to increased transpiration rate and also promoted the entry of atmospheric CO2, which in turn enhanced the photosynthesis rate [48]. This phenomenon could be a strategy of SL to alleviate drought stress by opening stomata and thus maintain the CO2 supply and enhance photosynthesis performance. Stomatal conductance may have also been influenced by osmoregulation and soluble sugar and proline contents may have been elevated by SL treatment. Consequently, the water-retention capacity of the plant and the water-uptake capacity of the plant roots were enhanced, thereby maintaining the water balance in the plant and thus opening the stomata [46, 49].
Drought stress inflicts substantial damage to the PSII reaction center in plants, and the damage to PSII accelerates Chl degradation and exacerbates the denaturation and inactivation of complex proteins in the PSII structure. These phenomena increase the photoinhibition and significantly reduce the light-energy-conversion efficiency of plants [50,51,52]. Chl fluorescence parameters can reflect the absorption and conversion of light energy in plant leaves under drought stress in terms of light energy conversion and electron-transfer efficiency of leaf PSII, which is an important basis for determining the degree of photosystem damage under drought stress conditions [53, 54]. Among them, Fv/Fm is the maximum photochemical efficiency of PSII, which can reflect the maximum capacity of photosystem reaction centers to utilize light energy, whereas PIabs reflects the overall performance of PSII [55,56,57]. In the present study, drought stress led to a significant decrease in Fv/Fm and PIabs in P. purpureum leaves, which continuously decreased with prolonged stress time. This finding indicated that the PSII reaction center of P. purpureum leaves was damaged by stress, and the damage became more serious with drought severity. Fv/Fm and PIabs were significantly elevated after the exogenous application of SL, indicating that SL effectively improved the function of PSII and alleviated photoinhibition. This finding may be due to the fact that SL alleviates the photosynthesis process by maintaining the stability of PSII or increasing the turnover of D1 protein and by improving the photosynthetic electron transport and the demand for NADPH in the Calvin cycle. Min et al. [22] obtained similar results in a study on grapes, where spraying of SL under drought stress elevated Fv/Fm levels. However, in the present study, the alleviating effect of SL on drought stress showed concentration dependence. Fv/Fm and PIabs had the best alleviating effect in T2 at all three sampling time points, and the difference was significant (P < 0.05) compared with the D group. Conversely, T4 produced a mitigating effect only on Fv/Fm, with almost no effect on PIabs, and PIabs was lower than those of the D group at 120 h. The high concentration may have slightly inhibited some of the PSII activity. Therefore, selecting the appropriate concentration in the application of exogenously administered plant hormones is important to relieve environmental stress.
The root system is the main organ for water and nutrient absorption in plants, and drought usually leads to self-thinning of the root system, resulting in decreased root-absorption capacity and the death of some roots by abscission, seriously affecting normal plant growth [58, 59]. In the present experiment, drought stress inhibited the primary root growth, and with the increasing degree of drought stress, the primary root length was significantly lower than that of the normal-watering group. The spraying of SL significantly promoted the growth of primary roots, and the difference was significant compared with the D group (P < 0.05). In particular, T2 had the most significant relief effect. Similar results have been found for SL in A. thaliana. Ruyter-Spira et al. [60] showed that SL positively regulates the primary root length in A. thaliana, and SL controls root elongation by regulating the content of A. thaliana growth hormone. The exogenous application of SL restores the root phenotype, increases the length of the original roots, and promotes the growth and development of plants under stress conditions.
Plants must constantly adjust their ABA levels to adapt to changes under physiological and environmental conditions. ABA accumulates rapidly under saline and drought conditions and is considered to be a “stress hormone” [61, 62]. The interactions among SL, growth hormone, ABA, and other plant hormones jointly regulate abiotic stresses. In particular, the interaction between SL and ABA is the key to abiotic stress tolerance [63, 64]. The interaction between SL and ABA plays an important role in integrating stress signals and regulating stomatal development and function. The results of this experimental study indicate that different concentration treatments and different time periods after hormone treatment exert different effects on the ABA content of P. purpureum. More ABA was produced P. purpureum leaves in the first 48 h after spraying SL. ABA may not yet be produced in large quantities during mild drought, and SL application may have stimulated ABA production and maintained a dynamic balance between the hormones to enhance plant tolerance. With increased drought severity, the ABA content of the D group increased continuously, whereas the application of SL alleviated the adverse effects of drought, such that the ABA content did not increase further and decreased significantly compared with the D group. The genes related to the SL signaling pathway may have also co-regulated ABA levels [65, 66]. Toh et al. [67] showed that the exogenous application of SL reduced the ABA content in A. thaliana seeds and reduced dormancy induced by heat stress. A recent study in soybean seeds also showed that SL application reduced ABA levels in soybean seeds under alkali stress and promoted seed germination [68].
PEPC, NADP-ME, and PPDK are key enzymes of C4 photosynthesis and their main role is to concentrate CO2 for the Calvin cycle. Under adverse conditions, the photosynthesis efficiency of plants can be enhanced, stress tolerance improved, and crop yield ultimately increased by regulating the activities of key photosynthesis enzymes in plants [69,70,71,72,73]. In the present experiment, PEPC and PPDK activities increased under drought stress, suggesting that P. purpureum can cope with drought by elevating the activities of key enzymes of leaf photosynthesis, consistent with the findings of Doubnerová et al. [74]. However, NADP-ME activity decreased under drought stress, and enzyme activity may have been reduced during the NADP-ME-catalyzed production of pyruvate, NADPH, and CO2. PEPC, PPDK, and NADP-ME activities increased after spraying with SL, and the difference was significant (P < 0.05) compared with the D group. This finding indicated that the ability of P. purpureum to fix CO2 was significantly enhanced and photosynthesis performance was significantly improved after SL application. The NADP-ME activity in D, T1, T2, and T3 and the PEPC and PPDK activities in D, T2, and T3 were positively correlated with the net Pn of the leaves, indicating that the key enzymes of the photosynthesis photosynthesis pathway played important roles in their photosynthesis.
Conclusions
In conclusion, the exogenous application of SL under drought stress significantly increased photosynthetic performance of P. purpureum leaves, reducing leaf photochemical damage, improving light-energy-utilization efficiency, and thus improving its adaptability in drought environment. Furthermore, SL application also increased the accumulation of biomass and the length of primary roots of P. purpureum to promote the growth of P. purpureum under drought conditions for better resistance to drought.
Methods
Test materials
The test material was P. purpureum, which was collected from the nursery of the National Juncao Engineering Technology Research Center of Fujian Agriculture and Forestry University. SL was purchased from Beijing Solabao Technology Co.
Experimental design
The experiments were performed in a greenhouse, and P. purpureum was cultivated using the soil-cultivation method. When P. purpureum grew to seven leaves, P. purpureum was selected and divided into six treatment groups: control group (N), with normal-watering treatment and SL concentration of 0; drought-stress group (D), with watering stopped for 7 days to induce natural drought, soil moisture content maintained at 40–45%, and SL concentration of 0; and T1, T2, T3, and T4, sprayed with 1, 3, 5, and 7 μmol·L− 1 SL after drought stress, respectively. Five replicates were set up for each treatment. In the SL-treatment group, each SL concentration was sprayed onto P. purpureum leaves at 8:00 every morning continuously for 2 days on the basis that the leaves and the back of the leaves were covered with water droplets. Photosynthesis indices and Chl fluorescence were measured at 24, 48, and 120 h after SL treatment. The root length and fresh dry weight of leaves and roots were measured. Leaf and root samples were collected, and the samples were rapidly transferred to a − 80 °C refrigerator for storage after quick freezing with liquid nitrogen.
Determination of photosynthesis parameters
The photosynthesis parameters of P. purpureum were measured using a CIRAS-3 portable photosynthesizer (PP-Systems Company Amesbury, MA01913, USA). The light intensity was set at 1200 μmol·(m2·s)− 1, the CO2 concentration was set at 380 μmol·mol− 1, and the air relative humidity was set at 75%. The photosynthesis parameters were measured under these conditions. The net Pn [μmol·(m2·s)− 1], Gs [μmol·(m2·s)− 1], Tr [mmol·(m2·s)− 1], Ci (μmol·mol− 1), and water-use efficiency (WUE; mmol·mol− 1) were measured in each treatment group. Five plants were measured for each treatment.
Chl fluorescence determination
The maximum photochemical efficiency (Fv/Fm) and the performance index based on absorbed light energy (PIabs) were determined using a plant-efficiency analyzer. The top third unfolded leaf of the plant was selected, and the leaf was dark-adapted for 20 min before Chl fluorescence-parameter determination. Five plants were measured for each treatment.
Biomass determination
P. purpureum was rinsed with distilled water, and the leaves and roots were dried and cut. The fresh weight of P. purpureum was immediately weighed with an analytical balance. Then, the plant was dried to constant weight with an electric blast dryer at 105 °C, and its dry weight was obtained. Three plants were measured per treatment. The formula for calculating the water content was as follows: water content (%) = (fresh weight − dry weight) / (fresh weight) × 100.
Root-length determination
Three plants were taken from each treatment at 24, 48, and 120 h after SL treatment, and the length of the main root of P. purpureum was measured in centimeters using a meter ruler (minimum scale of 1 mm).
Determination of ABA content
Leaf and root samples were taken at 24, 48, and 120 h after SL treatment. Then, they were snap frozen in liquid nitrogen and transferred into a − 80 °C refrigerator. The leaf and root samples were ground into powder with liquid nitrogen, and 0.1 g of sample was loaded into a centrifuge tube. Then, 0.9 mL of PBS buffer was added, shaken well, centrifuged at 3000 g and 4 °C for 10 min, and prepared for use. Ten microliters of supernatant was collected for ABA content determination according to the instructions of a Phytohormone Abscisic Acid (ABA) ELISA Kit (YX-010201P, purchased from Shanghai Preferred Biotechnology Co.). Three plants were measured for each treatment.
Determination of key enzyme activities of photosynthesis pathways
Leaf sampling was performed at 48 h after SL treatment. The samples were snap frozen in liquid nitrogen and transferred into a − 80 °C refrigerator. The leaf samples were ground into powder with liquid nitrogen, and 0.1 g of the sample was loaded into a centrifuge tube. After adding 1 mL of extraction solution, it was homogenized in an ice bath, centrifuged at 8000 g and 4 °C for 10 min, and placed on ice. Ten microliters of supernatant was collected to determine the key photosynthesis enzyme activities according to the instructions of a NADP-ME kit (YX-W-A104, purchased from Shanghai Preferred Biotechnology Co.), a PEPC kit (YX-W-B203, purchased from Shanghai Preferred Biotechnology Co.), and a PPDK kit (YX-W-PPDK, purchased from Shanghai Preferred Biotechnology Co.). Three plants were measured for each treatment.
Statistical analysis
Data were statistically analyzed using SPSS 20.0 (SPSS Inc., Chicago, IL, USA). One-way ANOVA and Duncan’s multiple comparison analysis were used to test the significance of differences between treatments (P < 0.05). The analysis results were expressed as the mean and standard error.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Abbreviations
- SL:
-
Strigolactone
- ABA:
-
Abscisic acid
- WT:
-
Wild-type
- Chl:
-
Chlorophyll
- ROS:
-
Reactive oxygen species
- Pn:
-
Net photosynthetic rate
- Gs:
-
Stomatal conductance
- Tr:
-
Transpiration rate
- Ci:
-
Intercellular CO2 concentration
- WUE:
-
Water use efficiency
- Fv/Fm:
-
Maximum photochemical efficiency
- PIabs:
-
Performance index based on absorbed light energy
- CAM:
-
Crassulacean acid metabolism
- PEPC:
-
Phosphoenolpyruvate carboxylase
- PPDK:
-
Pyruvate phosphate dikinase
- NADP-ME:
-
NADP-malic enzyme
References
Luo L, Xia H, Lu BR. Editorial: crop breeding for drought resistance. Front Plant Sci. 2019;10:314.
Banks JM, Percival GC, Rose G. Variations in seasonal drought tolerance rankings. Trees. 2019;33:1063–72.
Hein JA, Sherrard ME, Manfredi KP, Abebe T. The fifth leaf and spike organs of barley (Hordeum vulgare L.) display different physiological and metabolic responses to drought stress. BMC Plant Biol. 2016;16(1):248.
Hejnák V, Tatar Ö, Atasoy GD, Martinková J, Çelen AE, Hnilička F, et al. Growth and photosynthesis of upland and pima cotton: response to drought and heat stress. Plant Soil Environ. 2015;61(11):507–14.
Hosseini SA, Réthoré E, Pluchon S, Ali N, Billiot B, Yvin JC. Calcium application enhances drought stress tolerance in sugar beet and promotes plant biomass and beetroot sucrose concentration. Int J Mol Sci. 2019;20(15):3777.
Williams A, de Vries FT. Plant root exudation under drought: implications for ecosystem functioning. New Phytol. 2020;225(5):1899–905.
Hasan MM, Rahman MA, Skalicky M, Alabdallah NM, Waseem M, Jahan MS, et al. Ozone induced stomatal regulations, MAPK and phytohormone signaling in plants. Int J Mol Sci. 2021;22(12):6304.
Mubarik MS, Khan SH, Sajjad M, Raza A, Hafeez MB, Yasmeen T, et al. A manipulative interplay between positive and negative regulators of phytohormones: a way forward for improving drought tolerance in plants. Physiol Plant. 2021;172(2):1269–90.
Billah M, Aktar S, Brestic M, Zivcak M, Khaldun ABM, Uddin MS, et al. Progressive genomic approaches to explore drought- and salt-induced oxidative stress responses in plants under changing climate. Plants (Basel). 2021;10(9):1910.
López-Ráez JA, Shirasu K, Foo E. Strigolactones in plant interactions with beneficial and detrimental organisms: the yin and yang. Trends Plant Sci. 2017;22(6):527–37.
Alvi AF, Sehar Z, Fatma M, Masood A, Khan NA. Strigolactone: an emerging growth regulator for developing resilience in plants. Plants (Basel). 2022;11(19):2604.
Bhoi A, Yadu B, Chandra J, Keshavkant S. Contribution of strigolactone in plant physiology, hormonal interaction and abiotic stresses. Planta. 2021;254(2):28.
Bürger M, Chory J. The many models of strigolactone signaling. Trends Plant Sci. 2020;25(4):395–405.
Wang WN, Min Z, Wu JR, Liu BC, Xu XL, Fang YL, et al. Physiological and transcriptomic analysis of cabernet sauvginon (Vitis vinifera L.) reveals the alleviating effect of exogenous strigolactones on the response of grapevine to drought stress. Plant Physiol Biochem. 2021;167:400–9.
Zhang X, Zhang L, Sun Y, Zheng S, Wang J, Zhang T. Hydrogen peroxide is involved in strigolactone induced low temperature stress tolerance in rape seedlings (Brassica rapa L.). Plant Physiol Biochem. 2020;157:402–15.
Ling F, Su Q, Jiang H, Cui J, He X, Wu Z, et al. Effects of strigolactone on photosynthetic and physiological characteristics in salt-stressed rice seedlings. Sci Rep. 2020;10(1):6183.
Zhou X, Tan Z, Zhou Y, Guo S, Sang T, Wang Y, et al. Physiological mechanism of strigolactone enhancing tolerance to low light stress in cucumber seedlings. BMC Plant Biol. 2022;22(1):30.
Wei CQ, Chien CW, Ai LF, Zhao J, Zhang Z, Li KH, et al. The Arabidopsis B-box protein BZS1/BBX20 interacts with HY5 and mediates strigolactone regulation of photomorphogenesis. J Genet Genomics. 2016;43(9):555–63.
Lu T, Yu H, Li Q, Chai L, Jiang W. Improving plant growth and alleviating photosynthetic inhibition and oxidative stress from low-light stress with exogenous GR24 in tomato (Solanum lycopersicum L.) seedlings. Front Plant Sci. 2019;10:490.
Sattar A, Ul-Allah S, Ijaz M, Sher A, Butt M, Abbas T, et al. Exogenous application of strigolactone alleviates drought stress in maize seedlings by regulating the physiological and antioxidants defense mechanisms. Cereal Res Commun. 2022;50:263–72.
Ha CV, Leyva-González MA, Osakabe Y, Tran UT, Nishiyama R, Watanabe Y, et al. Positive regulatory role of strigolactone in plant responses to drought and salt stress. Proc Natl Acad Sci U S A. 2014;111(2):851–6.
Min Z, Li R, Chen L, Zhang Y, Li Z, Liu M, et al. Alleviation of drought stress in grapevine by foliar-applied strigolactones. Plant Physiol Biochem. 2019;135:99–110.
Naeem MS, Warusawitharana H, Liu H, Liu D, Ahmad R, Waraich EA, et al. 5-aminolevulinic acid alleviates the salinity-induced changes in brassica napus as revealed by the ultrastructural study of chloroplast. Plant Physiol Biochem. 2012;57:84–92.
Turan S, Kumar S, Cornish K. Photosynthetic response of in vitro guayule plants in low and high lights and the role of non-photochemical quenching in plant acclimation. Ind Crop Prod. 2014;54:266–71.
Lu T, Meng Z, Zhang G, Qi M, Sun Z, Liu Y, et al. Sub-high temperature and high light intensity induced irreversible inhibition on photosynthesis system of tomato plant (Solanum lycopersicum L.). Front Plant Sci. 2017;8:365–81.
Zhou Y, Schideman LC, Park DS, Stirbet A, Rupassara SI, Krehbiel JD, et al. Characterization of a chlamydomonas reinhardtii mutant strain with improved biomass production under low light and mixotrophic conditions. Algal Res. 2015;11:134–74.
Aldous SH, Weise SE, Sharkey TD, Waldera-Lupa DM, Stühler K, Mallmann J, et al. Evolution of the phosphoenolpyruvate carboxylase protein kinase family in C3 and C4 Flaveria spp. Plant Physiol. 2014;165(3):1076–91.
Covshoff S, Szecowka M, Hughes TE, Smith-Unna R, Kelly S, Bailey KJ, et al. C4 photosynthesis in the rice paddy: insights from the noxious weed echinochloa glabrescens. Plant Physiol. 2016;170(1):57–73.
Chen YB, Lu TC, Wang HX, Shen J, Bu TT, Chao Q, et al. Posttranslational modification of maize chloroplast pyruvate orthophosphate dikinase reveals the precise regulatory mechanism of its enzymatic activity. Plant Physiol. 2014;165(2):534–49.
Wang J, Gao H, Guo Z, Meng Y, Yang M, Li X, et al. Adaptation responses in C4 photosynthesis of sweet maize (Zea mays L.) exposed to nicosulfuron. Ecotoxicol Environ Saf. 2021;214:112096.
Swain A, Behera D, Karmakar S, Dash M, Dash BP, Swain P, et al. Morphophysiological alterations in transgenic rice lines expressing PPDK and ME genes from the C4 model Setaria italica. J Plant Physiol. 2021;264:153482.
Monção FP, Costa MAMS, Rigueira JPS, de Sales ECJ, Leal DB, da Silva MFP, et al. Productivity and nutritional value of BRS capiaçu grass (Pennisetum purpureum) managed at four regrowth ages in a semiarid region. Trop Anim Health Prod. 2020;52(1):235–41.
Huang PF, Mou Q, Yang Y, Li JM, Xu ML, Huang J, et al. Effects of supplementing sow diets during late gestation with Pennisetum purpureum on antioxidant indices, immune parameters and faecal microbiota. Vet Med Sci. 2021;7(4):1347–58.
Ujang FA, Roslan AM, Osman NA, Norman A, Idris J, Farid MAA, et al. Removal behaviour of residual pollutants from biologically treated palm oil mill effluent by Pennisetum purpureum in constructed wetland. Sci Rep. 2021;11(1):18257.
Zhou J, Chen S, Shi W, David-Schwartz R, Li S, Yang F, et al. Transcriptome profiling reveals the effects of drought tolerance in Giant Juncao. BMC Plant Biol. 2021;21(1):2.
Gupta A, Rico-Medina A, Caño-Delgado AI. The physiology of plant responses to drought. Science. 2020;368(6488):266–9.
Miller G, Suzuki N, Ciftci-Yilmaz S, Mittler R. Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ. 2010;33(4):453–67.
Zia R, Nawaz MS, Siddique MJ, Hakim S, Imran A. Plant survival under drought stress: implications, adaptive responses, and integrated rhizosphere management strategy for stress mitigation. Microbiol Res. 2021;242:126626.
Cheng X, Ruyter-Spira C, Bouwmeester H. The interaction between strigolactones and other plant hormones in the regulation of plant development. Front Plant Sci. 2013;4:199.
Banerjee A, Roychoudhury A. Strigolactones: multi-level regulation of biosynthesis and diverse responses in plant abiotic stresses. Acta Physiol Plant. 2018;40(5):1–10.
Wang Z, Li G, Sun H, Ma L, Guo Y, Zhao Z, et al. Effects of drought stress on photosynthesis and photosynthetic electron transport chain in young apple tree leaves. Biol Open. 2018;7(11):bio035279.
Mukarram M, Choudhary S, Kurjak D, Petek A, Khan MMA. Drought: sensing, signalling, effects and tolerance in higher plants. Physiol Plant. 2021;172(2):1291–300.
Drake JE, Power SA, Duursma RA, Medlyn BE, Aspinwall MJ, Choat B, et al. Stomatal and non-stomatal limitations of photosynthesis for four tree species under drought: a comparison of model formulations. Agric For Meteorol. 2017;247:454–66.
Bellasio C, Quirk J, Beerling DJ. Stomatal and non-stomatal limitations in savanna trees and C4 grasses grown at low, ambient and high atmospheric CO2. Plant Sci. 2018;274:181–92.
Dusenge ME, Duarte AG, Way DA. Plant carbon metabolism and climate change: elevated CO2 and temperature impacts on photosynthesis, photorespiration and respiration. New Phytol. 2019;221:32–49.
Sedaghat M, Emam Y, Mokhtassi-Bidgoli A, Hazrati S, Lovisolo C, Visentin I, et al. The potential of the synthetic strigolactone analogue GR24 for the maintenance of photosynthesis and yield in winter wheat under drought: investigations on the mechanisms of action and delivery modes. Plants (Basel). 2021;10(6):1223.
Thula S, Moturu TR, Salava H, Balakhonova V, Berka M, Kerchev P, et al. Strigolactones stimulate high light stress adaptation by modulating photosynthesis rate in Arabidopsis. J Plant Growth Regul. 2022:1–16. https://doi.org/10.1007/s00344-022-10764-5.
Lawson T, Vialet-Chabrand S. Speedy stomata, photosynthesis and plant water use efficiency. New Phytol. 2019;221(1):93–8.
Yang Y, Gu M, Chen J, Zhang R, Liu Z, Shi Y, et al. Comparative transcriptomes reveal the mitigation effect of GR24 in alfalfa under drought stress. J Plant Growth Regul. 2022:1–12. https://doi.org/10.1007/s00344-022-10779-y.
Ma H, Li P, Liu X, Li C, Zhang S, Wang X, et al. Poly-γ-glutamic acid enhanced the drought resistance of maize by improving photosynthesis and affecting the rhizosphere microbial community. BMC Plant Biol. 2022;22(1):11.
Zhuang J, Wang Y, Chi Y, Zhou L, Chen J, Zhou W, et al. Drought stress strengthens the link between chlorophyll fluorescence parameters and photosynthetic traits. PeerJ. 2020;8:e10046.
Killi D, Raschi A, Bussotti F. Lipid peroxidation and chlorophyll fluorescence of photosystem II performance during drought and heat stress is associated with the antioxidant capacities of C3 sunflower and C4 maize varieties. Int J Mol Sci. 2020;21(14):4846.
Helm LT, Shi H, Lerdau MT, Yang X. Solar-induced chlorophyll fluorescence and short-term photosynthetic response to drought. Ecol Appl. 2020;30(5):e02101.
Zhu L, Wen W, Thorpe MR, Hocart CH, Song X. Combining heat stress with pre-existing drought exacerbated the effects on chlorophyll fluorescence rise kinetics in four contrasting plant species. Int J Mol Sci. 2021;22(19):10682.
Wu HY, Tang HK, Liu LA, Shi L, Zhang WF, Jiang CD. Local weak light induces the improvement of photosynthesis in adjacent illuminated leaves in maize seedlings. Physiol Plant. 2021;171(1):125–36.
Shin YK, Bhandari SR, Lee JG. Monitoring of salinity, temperature, and drought stress in grafted watermelon seedlings using chlorophyll fluorescence. Front Plant Sci. 2021;12:786309.
Bano H, Athar HU, Zafar ZU, Kalaji HM, Ashraf M. Linking changes in chlorophyll a fluorescence with drought stress susceptibility in mung bean [Vigna radiata (L.) Wilczek]. Physiol Plant. 2021;172(2):1244–54.
Kim Y, Chung YS, Lee E, Tripathi P, Heo S, Kim KH. Root response to drought stress in rice (Oryza sativa L.). Int J Mol Sci. 2020;21(4):1513.
Li C, Li L, Reynolds MP, Wang J, Chang X, Mao X, et al. Recognizing the hidden half in wheat: root system attributes associated with drought tolerance. J Exp Bot. 2021;72(14):5117–33.
Ruyter-Spira C, Kohlen W, Charnikhova T, van Zeijl A, van Bezouwen L, de Ruijter N, et al. Physiological effects of the synthetic strigolactone analog GR24 on root system architecture in Arabidopsis: another belowground role for strigolactones? Plant Physiol. 2011;155(2):721–34.
Hong JH, Seah SW, Xu J. The root of ABA action in environmental stress response. Plant Cell Rep. 2013;32(7):971–83.
Lim CW, Baek W, Jung J, Kim JH, Lee SC. Function of ABA in stomatal defense against biotic and drought stresses. Int J Mol Sci. 2015;16(7):15251–70.
Marzec M, Daszkowska-Golec A, Collin A, Melzer M, Eggert K, Szarejko I. Barley strigolactone signalling mutant hvd14.D reveals the role of strigolactones in abscisic acid-dependent response to drought. Plant Cell Environ. 2020;43(9):2239–53.
Liu J, He H, Vitali M, Visentin I, Charnikhova T, Haider I, et al. Osmotic stress represses strigolactone biosynthesis in Lotus japonicus roots: exploring the interaction between strigolactones and ABA under abiotic stress. Planta. 2015;241(6):1435–51.
Feng Z, Liang X, Tian H, Watanabe Y, Nguyen KH, Tran CD, et al. SUPPRESSOR of MAX2 1 (SMAX1) and SMAX1-LIKE2 (SMXL2) negatively regulate drought resistance in Arabidopsis thaliana. Plant Cell Physiol. 2022:pcac080. https://doi.org/10.1093/pcp/pcac080.
Bu Q, Lv T, Shen H, Luong P, Wang J, Wang Z, et al. Regulation of drought tolerance by the F-box protein MAX2 in Arabidopsis. Plant Physiol. 2014;164(1):424–39.
Toh S, Kamiya Y, Kawakami N, Nambara E, McCourt P, Tsuchiya Y. Thermoinhibition uncovers a role for strigolactones in Arabidopsis seed germination. Plant Cell Physiol. 2012;53(1):107–17.
Nisa Z u, Mi X, Anwar S, Chen C, Jin X, Yu L, et al. Strigolactone enhances alkaline tolerance in soybean seeds germination by altering expression profiles of ABA biosynthetic and signaling genes. J Plant Biol. 2022:1–9. https://doi.org/10.1007/s12374-022-09357-2.
Liu X, Li X, Zhang C, Dai C, Zhou J, Ren C, et al. Phosphoenolpyruvate carboxylase regulation in C4-PEPC-expressing transgenic rice during early responses to drought stress. Physiol Plant. 2017;159(2):178–200.
Zhang H, Xu W, Wang H, Hu L, Li Y, Qi X, et al. Pyramiding expression of maize genes encoding phosphoenolpyruvate carboxylase (PEPC) and pyruvate orthophosphate dikinase (PPDK) synergistically improve the photosynthetic characteristics of transgenic wheat. Protoplasma. 2014;251(5):1163–73.
Chen Q, Wang B, Ding H, Zhang J, Li S. Review: the role of NADP-malic enzyme in plants under stress. Plant Sci. 2019;281:206–12.
Tang Y, Li X, Lu W, Wei X, Zhang Q, Lv C, et al. Enhanced photorespiration in transgenic rice over-expressing maize C4 phosphoenolpyruvate carboxylase gene contributes to alleviating low nitrogen stress. Plant Physiol Biochem. 2018;130:577–88.
He Y, Xie Y, Li X, Yang J. Drought tolerance of transgenic rice overexpressing maize C4-PEPC gene related to increased anthocyanin synthesis regulated by sucrose and calcium. Biol Plantarum. 2020;64(13):6–149.
Doubnerová Hýsková V, Miedzińska L, Dobrá J, Vankova R, Ryšlavá H. Phosphoenolpyruvate carboxylase, NADP-malic enzyme, and pyruvate, phosphate dikinase are involved in the acclimation of Nicotiana tabacum L. to drought stress. J Plant Physiol. 2014;171(5):19–25.
Acknowledgements
The authors are thankful to all the anonymous reviewers, research team, and funding agencies.
Funding
This work was supported by the National Natural Science Foundation of China (32101418, 42075116) and Major Project of Fujian Province (2021NZ029009).
Author information
Authors and Affiliations
Contributions
YL performed the experiments and wrote the manuscript. STL, QXF, JZ1, XLH and LZ performed parts of the experiments. FLY designed the experiments, provided experimental materials and financial assistance. JZ2 conceived the experiments, revised and edited the manuscript and provided financial support. All authors have read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, Y., Li, S., Feng, Q. et al. Effects of exogenous Strigolactone on the physiological and ecological characteristics of Pennisetum purpureum Schum. Seedlings under drought stress. BMC Plant Biol 22, 578 (2022). https://doi.org/10.1186/s12870-022-03978-y
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-022-03978-y