
Abstract
Background
Soybean is one of the four major crops in China. The occurrence of viruses in soybean causes significant economic losses.
Results
In this study, the soybean leaves from stay-green plants showing crinkle were collected for metatranscriptomic sequencing. A novel geminivirus, tentatively named soybean geminivirus A (SGVA), was identified in soybean stay-green plants. Sequence analysis of the full-length SGVA genome revealed a genome of 2762 nucleotides that contain six open reading frames. Phylogenetic analyses revealed that SGVA was located adjacent to the clade of begomoviruses in both the full genome-based and C1-based phylogenetic tree, while in the CP-based phylogenetic tree, SGVA was located adjacent to the clade of becurtoviruses. SGVA was proposed as a new recombinant geminivirus. Agroinfectious clone of SGVA was constructed. Typical systemic symptoms of curly leaves were observed at 11 dpi in Nicotiana benthamiana plants and severe dwarfism was observed after 3 weeks post inoculation. Expression of the SGVA encoded V2 and C1 proteins through a potato virus X (PVX) vector caused severe symptoms in N. benthamiana. The V2 protein inhibited local RNA silencing in co-infiltration assays in GFP transgenic 16C N. benthamiana plants. Further study revealed mild symptoms in N. benthamiana plants inoculated with SGVA-ZZ V2-STOP and SGVA-ZZ V2-3738AA mutants. Both the relative viral DNA and CP protein accumulation levels significantly decreased when compared with SGVA-inoculated plants.
Conclusions
This work identified a new geminivirus in soybean stay-green plants and determined V2 as a pathogenicity factor and silencing suppressor.
Similar content being viewed by others


Background
Soybean is one of the four major crops in China. Soybean seeds are rich in oil and protein, and therefore served as an important resource for food and industrial products. Soybean is vulnerable to infection by many viruses [1, 2]. More than 67 viruses have been reported to infect soybean crops worldwide [3]. Among them viruses in the family Geminiviridae have been found in soybeans such as common bean curly stunt virus (CBCSV) [4], soybean chlorotic spot virus (SoCSV) [5], mungbean yellow mosaic India virus (MYMIV) [6], african cassava mosaic virus (ACMV) [7], cowpea golden mosaic virus (CPGMV), dolichos yellow mosaic virus (DoYMV) and soybean mild mottle virus (SbMMoV) [8].
Geminiviruses are plant pathogens causing significant economic losses to many crops worldwide [9]. Currently, according to new 2020 ICTV taxonomy release the family is divided into fourteen genera [10], namely Becurtovirus, Begomovirus, Capulavirus, Citlodavirus, Curtovirus, Eragrovirus, Grablovirus, Maldovirus, Mastrevirus, Mulcrilevirus, Opunvirus, Topilevirus, Topocuvirus, and Turncurtovirus. Begomovirus, the largest genus within the family Geminiviridae, contains circular single-strand DNA, which can be either bipartite composed of both DNA-A and DNA-B components or monopartite containing a single DNA-A like component. The open reading frames (ORFs) of the DNA-A component encode six genes which are coat protein (ORF AV1/V1), movement protein (ORF AV2/V2), replication associated protein (ORF AC1/C1), transcriptional activator (ORF AC2/C2), a replication enhancer (ORF AC3/C3) and a AC4/C4 protein [11].
In the present study, we identified a new monopartite geminivirus from soybean plants in China, designated as soybean geminivirus A (SGVA). Infectious DNA clone of SGVA was constructed and inoculated into Nicotiana benthamiana plants via agrobacterium-mediated infiltration to show that SGVA causes disease symptoms. We tested the role of protein encoded by SGVA using PVX expression system and found that V2 and C1 are crucial for PVX symptom development and virus accumulation. We further identified V2 as the viral suppressor of RNA silencing (VSR) and as a pathogenicity factor.
Results
Identification and sequencing of an unknown geminivirus
Soybean samples from Zhengzhou, China that showed crinkle and stay-green symptoms were collected (Fig. 1A) and mRNA library was constructed and sequenced using Illumina HiSeq X ten platform. In total 69,986,830 clean reads were obtained after removing low-quality reads and adaptor sequences. After assembly using the CLC Genomics Workbench (version:6.0.4), 62,243 primary unigenes were generated (Table S1). These unigenes were then assembled for a second time using CAP3 EST software to acquire the final unigene sequences. After final assembly in total 54,208 contigs were obtained (Table S2). One contig (contig7689) of 2020 nucleotides in length with the most reads of 2367 was acquired through sequencing. As shown in Fig. S1 both tomato leaf curl Cebu virus and ageratum yellow vein China virus showing the most abundant expression were mapped to the contig 7689. Using BLASTn analysis the contig showed 98.37% nucleotide identity on 97% coverage with an unpublished soybean geminivirus sequence (GenBank No. MH428829) and 83.62% nucleotide identity with tomato leaf curl Java virus (GenBank accession No. AB162141) on 55% coverage. The name “soybean geminivirus A” isolate Zhengzhou (SGVA-ZZ) is proposed for this virus. The occurrence of SGVA was detected in two symptomatic field samples (Fig. 1B). According to the sequence of contig 7689, the full-length sequence of SGVA was assembled using primer pair AF/AR and BF/BR to amplify full length of SGVA and a fragment of 2377 nt in length (Fig. 1C) and deposited in GenBank as accession No. MZ505080. The whole genome was 2762 nt in length. SGVA was found to contain six putative open reading frames (ORFs), including the V1 (786 nt) and V2 (312 nt) genes on the viral sense strand, and the C1 (1086 nt), C2 (417 nt), C3 (456 nt) and C4 (291 nt) genes on the complementary strand (Fig. 1D). A geminiviral conserved 9-base nucleotide sequence (TAATATTAC) [12] was also present in the genome of SGVA.

Symptoms, mRNA sequencing and detection of SGVA. A Symptoms of SGVA infection in the soybean stay-green plant. B PCR detection of the full length genome (2762 bp) of SGVA in the two field samples using primer pair AF/AR. Lane 1 and 3 are symptomatic soybean plants. Lane 2 and 4 are healthy soybean plants. C PCR amplification of the full length genome (2762 bp) and 0.86 copy of the genome (2377 bp). M, DNA marker. Lane 1 and 3 are healthy soybean leaves, Lane 2 and 4 are symptomatic soybean leaves. D Schematic diagram of the SGVA-ZZ genome
Phylogenetic analysis of the viral genome
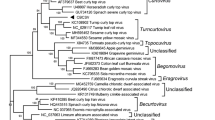
The neighbour-joining phylogenetic analyses based on the nucleotide sequence of the full-length genome and the amino acid sequence of coat protein (CP) (V1) and C1 were performed. SGVA-ZZ was located adjacent to the clade of begomoviruses in both the full genome-based and C1-based phylogenetic tree (Fig. 2A, B), while in the CP-based phylogenetic tree, SGVA-ZZ was located adjacent to the clade of becurtoviruses (Fig. 2C). Hence, SGVA-ZZ was proposed as a possible new recombinant geminivirus.

Phylogenetic analysis of SGVA. A Phylogenetic relationships of full genome sequences of SGVA with selected geminiviruses. The tree was constructed by the Neighbor-joining method with 1,000 bootstrap replicates in the MEGA X software. B, C Phylogenetic relationships of SGVA with selected geminiviruses based on C1 (B) and CP (C) amino acid sequences. The tree was constructed by the Neighbor-joining method with 1,000 bootstrap replicates in the MEGA X software
Infectivity of SGVA in Nicotiana benthamiana
The full-length cDNA clone of SGVA was constructed into vector pGD to acquire the recombinant vector, pGDSGVA (Fig. 3A). The resultant clone was agro-infiltrated into 4-week-old leaves of N. benthamiana plants. Typical systemic symptoms of curly leaves were observed at 11 days post inoculation (dpi), and severe dwarfism was observed after 3 weeks post inoculation (Fig. 3B). At 33 dpi the flowering of SGVA infected N. benthamiana plants was inhibited compared to control plants (Fig. S2). To verify the presence of viral DNA in systemic leaves at 13 dpi, total DNA was extracted and detected using the primer pair CP-F/R (Fig. 3C). The results indicated that the infection rate was 100% for SGVA during three biological repeats of 10, 8 and 8 plants, respectively. Furthermore, SGVA accumulation in systemic leaves at 13 dpi was indicated by performing western blotting with SGVA CP polyclonal antibodies (Fig. 3D).

Systemic infection of SGVA-ZZ in Nicotiana benthamiana. A Construction of infectious cDNA clone. Whole viral genome was cloned using fragments F1 (2762 bp) and F2 (2377) digested with BglII/HindIII and HindIII/SalI, respectively and sequentially ligated into pGD vector to obtain the recombinant clone PGDSGVA-ZZ. B Systemic symptoms induced by SGVA-ZZ in N. benthamiana plants were observed at 13 dpi and 23 dpi, respectively. C The presence of SGVA-ZZ in N. benthamiana systemic leaves was confirmed (13 dpi) by PCR using primers specially targeting SGVA-ZZ CP. D SGVA accumulation in N. benthamiana systemic leaves was detected by conducting western blotting analysis with anti-SGVA CP antibodies. Coomassie Brilliant Blue (CBB) staining of the large subunit of RuBisCo served as a loading control
Screening for potential virulence factors encoded by SGVA-ZZ
To determine the potential virulence factors encoded by SGVA, six ORFs were transiently expressed in N. benthamiana plants via a PVX-based heterologous expression system. By 12 dpi, the N. benthamiana plants inoculated with PVX-V2 and PVX-C1 showed apical necrosis that ultimately led to the death of plants, and the plants inoculated with PVX-V1, PVX-C2, PVX-C3, and PVX-C4 produced PVX-like symptoms (Fig. 4A). To determine whether the severe symptoms were associated with higher virus accumulation, the expression of the CP in the systemic leaves was detected using western blotting with antibodies against PVX CP. The infection of PVX-V2, PVX-C1 and PVX-C2 promoted viral accumulation of PVX, while the accumulation of PVX-V1, PVX-C3 showed lower CP levels and the CP accumulation of PVX-C4 was equivalent to PVX (Fig. 4B). Furthermore, the viral RNA accumulation was determined using qRT-PCR (Fig. 4C), and the result was consistent with western blotting analysis.

SGVA V2 and C1 enhance pathogenicity of PVX. A Symptoms of N. benthamiana plants inoculated with PVX, PVX-V1, PVX-V2, PVX-C1, PVX-C2, PVX-C3 and PVX-C4. Symptoms were photographed at 12 dpi. The accumulation of PVX was detected by western blotting (B) and quantitative reverse transcription PCR (qRT-PCR) (C) respectively. Coomassie brilliant blue-stained RuBisCo large subunit protein (CBB) was used to show sample loadings. The expression of NbUBC was used as an internal control in qRT-PCR. The results are presented as means ± SD from three biological replicates per treatment. Bars represents the mean ± standard deviation (SD). The statistical significance between treatments was determined using Duncan's multiple range test (p* < 0 .05, p** < 0.01)
Identification of SGVA V2 as an RNA silencing suppressor
V2 was previously reported as a VSR in some geminiviruses [13]. To further investigate the RNA silencing suppressor encoded by SGVA V2, ORF of V2 were introduced into the binary vector pGD. Leaves of 16C transgenic N. benthamiana plants carrying GFP were infiltrated with a mixture of agrobacterium harboring 35S-GFP and either a test or a control vector (Fig. 5A). The pGD + GFP agrobacterium culture harboring empty vector was infiltrated into 16C plants as negative control and P19 + GFP agrobacterium culture expressing the P19 silencing suppressor of tomato bushy stunt virus was infiltrated as positive control. At 3 d post infiltration, the leaf expressing V2 + GFP showed obvious and stronger green fluorescence under UV light similar to that produced by P19 + GFP (Fig. 5A), which correlates with the enhanced accumulation of GFP proteins by western blotting analysis and GFP mRNA by qRT-PCR analysis (Fig. 5B, C). These results suggest that SGVA V2 is an RNA silencing suppressor.

Suppression of RNA silencing by SGVA V2. A The leaves of GFP-transgenic 16C line were co-infiltrated with agrobacterium suspension harboring 35S-GFP expressing GFP and the recombinant vectors expressing SGVA V2 protein as indicated. The leaves expressing pGD + GFP were used as negative control and leaves expressing P19 + GFP were used as positive control. The photographs were taken under UV light at 3 d post infiltration. B Western blotting analysis of GFP accumulation in the co-infiltrated leaf patches at 3 d post infiltration. Coomassie brilliant blue-stained RuBisCo large subunit protein (CBB) was used to show sample loadings. C qRT-PCR analysis of GFP accumulation in the co-infiltrated leaf patches at 3 d post infiltration. The expression of NbUBC was used as an internal control in qRT-PCR. The results are presented as means ± SD from three biological replicates per treatment. Bars represents the mean ± standard deviation (SD). The statistical significance between treatments was determined using Duncan's multiple range test (p* < 0 .05, p** < 0.01)
Identification of V2 as a pathogenicity determinant
To further study the function of V2, we constructed two V2 mutants of SGVA-ZZ. SGVA-ZZV2-STOP mutant in which the start codons in the V2 gene (ATG) was modified to ATC. Besides we performed an alignment using the amino acid sequence of SGVA-ZZ V2 and that of other geminivirus V2 with the DNAMAN software (Fig. S3). Two conserved amino acids (L37, I38) in the V2s were found and were substituted with alanine to generate SGVA-ZZV2-3738AA mutant (Fig. 6A). Then, N. benthamiana were inoculated separately with SGVA-ZZ, SGVA-ZZV2-STOP and SGVA-ZZV2-3738AA. At 14 dpi, the SGVA-ZZ inoculated N. benthamiana plants showed obvious symptoms of curly leaves, while the SGVA-ZZV2-STOP and SGVA-ZZV2-3738AA inoculated plants did not show obvious virus symptoms (Fig. 6B). Western blot and qRT-PCR analysis results showed that SGVA-ZZ viral DNA and CP accumulations were significantly reduced in the systemic leaves of the SGVA-ZZV2-STOP and SGVA-ZZV2-3738AA inoculated plants compared to that in the systemic leaves of the SGVA-ZZ inoculated plants, indicating that V2 expression was essential for SGVA-ZZ systemic infection and conserved amino acids L37I38 mutation inhibited the systemic infection of SGVA-ZZ (Fig. 6C, D). Furthermore, by 21 dpi, the SGVA-ZZ inoculated N. benthamiana plants showed severe symptoms, while the SGVA-ZZV2-STOP and SGVA-ZZV2-3738AA inoculated plants showed mild symptoms of curly leaves (Fig. 6E). Western blot and qRT-PCR analysis further confirmed the symptoms observed (Fig. 6F, G). Furthermore, the VSR activity of the two V2 mutants were examined in N. benthamiana plants. The pGD + GFP agrobacterium culture harboring empty vector was infiltrated into N. benthamiana plants as negative control and P19 + GFP agrobacterium culture was infiltrated as positive control. At 3 d post infiltration, the leaf expressing V2 + GFP showed obvious and stronger green fluorescence under UV light similar to that produced by P19 + GFP while V2-STOP + GFP and V2-3738AA + GFP showed weak fluorescence as pGD + GFP control (Fig. 7A). Further detection of GFP proteins and mRNA by western blotting and qRT-PCR analysis confirmed the observation (Fig. 7B, C).

SGVA V2 functions as a pathogenicity determinant. A SGVA-ZZV2-STOP and SGVA-ZZV2-3738AA was constructed as shown in the schematic diagram. Symptoms of N. benthamiana plants inoculated with SGVA-ZZ, SGVA-ZZV2-STOP and SGVA-ZZV2-3738AA were photographed at 14 dpi and 21 dpi, respectively (B, E). The accumulation of SGVA-ZZ was detected by western blotting (C, F) and quantitative reverse transcription PCR (qRT-PCR) (D, G) respectively. Coomassie brilliant blue-stained RuBisCo large subunit protein (CBB) was used to show sample loadings. The expression of NbUBC was used as an internal control in qRT-PCR. The results are presented as means ± SD from three biological replicates per treatment. Bars represents the mean ± standard deviation (SD). The statistical significance between treatments was determined using Duncan's multiple range test (p* < 0 .05, p** < 0.01)

V2 mutants abolishes RNA silencing suppressor activity of V2. A The leaves of Nicotiana benthamiana were co-infiltrated with agrobacterium suspension harboring 35S-GFP expressing GFP and the recombinant vectors expressing SGVA V2, V2-STOP and V2-3738AA proteins as indicated. The leaves expressing pGD + GFP were used as negative control and leaves expressing P19 + GFP were used as positive control. The photographs were taken under UV light at 3 d post infiltration. B Western blotting analysis of GFP accumulation in the co-infiltrated leaf patches at 3 d post infiltration. Coomassie brilliant blue-stained RuBisCo large subunit protein (CBB) was used to show sample loadings. C qRT-PCR analysis of GFP accumulation in the co-infiltrated leaf patches at 3 d post infiltration. The expression of NbUBC was used as an internal control in qRT-PCR. The results are presented as means ± SD from three biological replicates per treatment. Bars represents the mean ± standard deviation (SD). The statistical significance between treatments was determined using Duncan's multiple range test (p* < 0 .05, p** < 0.01)
Discussion
The family Geminiviridae consists of fourteen genera that mainly affect a wide range of plants. Of all the fourteen genera, Begomovirus is the largest member of the group, which has about 445 species. Several begomoviruses have been reported to infect soybean [4,5,6,7,8]. In this study we characterized a new geminivirus named SGVA from diseased soybean stay-green plants showing leaf curling symptoms in Zhengzhou, China. Sequence analysis reveals that SGVA shares 98.37% identity with the unpublished Genbank sequences (GenBank No. MH428829). According to the phylogenetic analysis, SGVA was adjacent to the clade of begomoviruses based on full-length nucleotide sequence and C1 amino acid sequences, while SGVA was adjacent to the clade of becurtoviruses based on CP amino acid sequences. SGVA was proposed as a new recombinant geminivirus. The high genetic variability of geminivirus populations is predominantly driven by their high mutational dynamics combined with recombination [14,15,16].
In this study we found that the over-expression of V2 protein induced systemic necrosis in N. benthamiana plants, and indicating that it functions as a key virulence factor. Besides, many V2 proteins encoded by different geminiviruses have been demonstrated to be important virulence determinants [13, 17, 18]. PVX-C1-inoculated plants started to show systemic necrosis at 5 dpi. Previous research showed that PVX expressing C1 of apple geminivirus (AGV) developed visible necrotic lesions at 13 dpi [19]. To further confirm the function of V2 in viral pathogenicity, two SGVA mutants, SGVA-ZZV2-STOP and SGVA-ZZV2-3738AA, were constructed. Both mutants showed milder symptoms and lower viral accumulation compared with SGVA inoculated Nicotiana benthamiana plants indicating V2 functions as a pathogenicity factor during SGVA infection. The over-expression of V1 and C3 showed lower viral accumulation than PVX possibly due to the activation of host defense related pathways by V1 and C3 over-expression and thereby inhibited viral accumulation. C2 over-expression promoted PVX CP accumulation at higher level than the mRNA accumulation. C2 plays role in the suppression of host immune responses and regulation of the ubiquitin/proteasome system (UPS) [20]. It is possible that C2 interferes with the UPS pathway and promote the CP accumulation. Multiple virus encoded RNA silencing suppressors are involved in viral pathogenicity, such as pepper vein yellows virus P0, tomato leaf curl Java virus V2 protein, rice stripe mosaic virus P4 protein, watermelon silver mottle virus nonstructural protein S [18, 21,22,23]. In our study SGVA encoded V2 protein acts as a RNA silencing suppressor and pathogenicity determinant which is consistent with the previous studies.
Here in our study we identified a new geminivirus in soybean stay-green plants and established the infectious clone system in Nicotiana benthamiana. We’ve tried different inoculation methods including agro-inoculation, shot-gun and sap-inoculation to inoculate two varieties of soybean, whereas the infection was not established in soybeans plants. Considering SGVA may be a threat to soybean production in China, further efforts to elucidate the impact of SGVA on soybean plants are underway.
Conclusions
In this work we identified a new monopartite geminivirus from soybean stay-green plants in China, designated as soybean geminivirus A (SGVA). Infectious DNA clone of SGVA was constructed and inoculated into Nicotiana benthamiana plants via agrobacterium-mediated infiltration to show that SGVA causes disease symptoms. We further identified V2 as the viral suppressor of RNA silencing (VSR) and as a pathogenicity factor. Conserved amino acids L37I38 of V2 are essential for the viral pathogenicity.
Materials and methods
Plant materials and virus inoculation
During field survey in August 2020, soybean samples exhibiting crinkle and stay-green symptoms were collected from Zhengzhou, Henan province of China. N. benthamiana plants were grown in pots in a growth room under a 16 h light/8 h dark photoperiod at 25 °C with 60% humidity. For agroinfiltration, Agrobacteria strain GV3101 carrying infectious viral clones were suspended in infiltration buffer (10 mM MgCl2, 10 mM MES, and 200 μM acetosyringone, pH 5.6) at an OD600 of 1, kept at room temperature for 2 to 4 h and infiltrated into N. benthamiana leaves using a 1-mL needleless syringe.
Plasmid construction
Primers used for plasmid construction are listed in Table S3. All the available constructs were sequenced.
PVX recombinant vectors was constructed by introducing coding sequences of SGVA encoded proteins into potato virus X (PVX) vector pGR106 via ClaI and SalI digestion, followed by ligation with T4 DNA ligase (NEB). The fragments used were amplified using primer pairs PVX-V2/V1/C1/C2/C2/C4-F/R.
To construct vectors for RNA silencing analysis, the coding sequence of corresponding genes were amplified and inserted into PstI/BamHI digested pGD vectors via homologous recombination using Clonexpress II one step cloning kit (Vazyme, China) [24], the homologous arm length was 20 bp. The fragments used were amplified using primer pairs PGD-V2 -F/R.
To construct full-length cDNA clone of SGVA, a recombinant plasmid containing 1.8 copies of the full-length fragment of the viral genome was constructed. First, a 2762 bp full length was amplified from total DNA extracted in symptomatic leaves using Q5 high-fidelity polymerase (NEB, Beijing,China) with primer pair SGVAF1F/ SGVAF1R and ligated into binary vector pGD vector between the cauliflower mosaic virus (CaMV) 35S promoter and the nopaline synthase terminator (t-Nos) via the restriction sites BglII/ HindIII to produce construct pGDSGVAF1. After sequencing, a 2377 bp partial fragment was amplified using primer pair SGVAF2F/SGVAF2R and ligated into pGDSGVAF1 vector via the restriction sites HindIII/SalI to produce construct pGDSGVA.
To construct V2 mutant vectors, a 2762 bp full length was amplified from pGDSGVA-ZZ using primer pairs SGVAV2STOPF/ SGVAF1R, SGVAF1F/ SGVAV2STOPR and ligated into pGD vector via the restriction sites BglII/ HindIII to produce construct pGDSGVAv2stopF1, then a 2377 bp partial fragment was amplified using primer pair SGVAV2STOPF/ SGVAF2R, SCVF2F/ SGVAV2STOPR and ligated into pGDSGVAv2stopF1 vector via the restriction sites HindIII/SalI to produce construct pGDSGVAv2stop. SGVAV2-3738AA mutant was constructed using primer pairs SGVA3738AAF and SGVA3738AAR in the same way.
Nucleic Acid Extraction, Metatranscriptomic Sequencing, and Data analysis
The total RNA of samples was subjected to an rRNA removal procedure using a Ribo-zero Magnetic kit according to the manufacturer’s instructions (Epicentre, an Illumina® company). Next, cDNA libraries were constructed using a TruSeq™ RNA sample prep kit (Illumina). Barcoded libraries were paired-end sequenced on an Illumina HiSeq X ten platform at Shanghai biotechnology Co., Ltd. (Shanghai, China) according to the manufacturer’s instructions (www.illumina.com).
To obtain clean reads, the Fastax online software (version: 0.0.13, http://hannonlab.cshl.edu/fastx_toolkit/index.html) was used to screen out unqualified reads from the raw reads; this step removed joint sequences, reads with low overall quality (mainly reads with lengths less than 20 bp), reads with base N (base with uncertain identity), reads with more than 20% of the bases possessing Q-values ≤ 10, and or low end quality. Then, the reads were trimmed to remove bases with a quality score of soybean. Clean reads were de novo assembled using CLC Genomics Workbench (version:6.0.4) according to the scaffolding contig algorithm (word-size = 45, minimum contig length = 300) [25,26,27]. These various steps produced the primary unigenes. These were then assembled for a second time using CAP3 EST software [28] to acquire the final unigene sequence set. This unigene set was used for further exploration of the transcriptome. The final unigene set was compared against the NCBI non-redundant (Nr) database using BLASTX [29], with an E-value < 1e−5. Then the final unigenes annotated with virus were used for the virus analysis.The assembled contigs were queried by homology search tools (BLASTn and BLASTx) against public database (GenBank) in the NCBI (the National Center for Biotechnology Information).
Phylogenetic analysis
Sequences alignments were conducted using the ClustalW method, and phylogenetic trees were constructed by the neighbor joining method using MEGA version X [30]. The tree was evaluated with 1000 bootstrap replicates. Sequences of geminiviruses were retrieved from GenBank and used for comparison and phylogenetic analysis in this study.
Western blotting analysis
Agro-infiltrated leaves were harvested for western blotting assay. Total protein was extracted from 0.2 g leaves using the extraction buffer containing 20% glycerol, 20 mM Tris–HCl (pH 7.5), 1 mM EDTA, 150 mM NaCl, 1 mM PMSF, 1 × Protease inhibitor cocktail (Sigma, China). Total protein was separated in SDS–polyacrylamide gel electrophoresis, followed by transfer to nitrocellulose membranes. The membranes were probed using anti-PVX CP polyclonal antibodies or SGVA CP polyclonal antibodies followed by an HRP-conjugated secondary antibody. Antigens of PVX CP and SGVA CP was acquired via prokaryotic expression. The detection signals were developed using an ECL reagent as instructed. PVX CP and SGAV CP accumulation were photographed under a chemiluminescence apparatus (Amersham imager 680). CBB staining of the large subunit of RuBisCo served as a loading control. Due to the figure size the original size image of the blots was supplied in the supplementary information.
Quantitative RT-PCR
Total RNA was extracted from harvested N. benthamiana leaves using Trizol reagent (invitrogen) and treated with RNase-free DNase I. First strand cDNA was synthesized using 500 ng total RNA, an oligo d(T) primer, random primer, and M-MLV reverse transcriptase as instructed. Ten-fold diluted cDNA product was used for qPCR on an Eppendorf Real-Time PCR system using a SYBR Green master mix (Takara). The NbUBC genes (GenBank accession number: AB026056.1) was used as internal controls. All the primers used for qRT-PCR are listed in Table S3. The relative gene expression levels were calculated using the 2−△△CT method.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its supplementary information files.
References
Hartman GL, West ED, Herman TK. Crops that feed the World 2. Soybean—worldwide production, use, and constraints caused by pathogens and pests. Food Secur. 2011;3:5–17.
Hill JH, Whitham SA. Control of Virus Diseases in Soybeans - ScienceDirect. Adv Virus Res. 2014;90:355–90.
Ali A. Rapid detection of fifteen known soybean viruses by dot-immunobinding assay. J Virol Methods. 2017;249:126–9.
Zhang R, Wu X, Jiang X, Wu X, Cheng X. Molecular characterization of common bean curly stunt virus: a novel recombinant geminivirus in China. Adv Virol. 2020;165:257–60.
Coco D, Calil IP, Brustolini OJB, Santos AA, Inoue-Nagata AK, Fontes EPB. Soybean chlorotic spot virus, a novel begomovirus infecting soybean in Brazil. Adv Virol. 2013;158:457–62.
Ramesh SV, Shivakumar M, Ramteke R, Bhatia VS, Chouhan BS, Goyal S, et al. Quantification of a legume begomovirus to evaluate soybean genotypes for resistance to yellow mosaic disease. J Virol Methods. 2019;268:24–31.
Mgbechi-Ezeri JU, Alabi OJ, Naidu RA, Kumar PL. First Report of the Occurrence of African cassava mosaic virus in a Mosaic Disease of Soybean in Nigeria. Plant Dis. 2008;92:1709.
Mollel HG, Sseruwagi P, Ndunguru J, Alicai T. Desmodium mottle virus, the first legumovirus (genus Begomovirus) from East Africa. Adv Virol. 2017;162:1799–803.
Mansoor S, Zafar Y, Briddon RW. Geminivirus disease complexes: the threat is spreading. Trends Plant Sci. 2006;11:209–12.
Walker PJ, Siddell SG, Lefkowitz EJ, Mushegian AR, Adriaenssens EM, Alfenas-Zerbini P, et al. Changes to virus taxonomy and to the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2021). Arch Virol. 2021;166:2633–48.
Zerbini FM, Briddon RW, Idris A, Martin DP, Moriones E, Navas-Castillo J, et al. ICTV Virus Taxonomy Profile: Geminiviridae. J Gen Virol. 2017;98:131–3.
Stenger DC, Revington GN, Stevenson MC, Bisaro DM. Replicational release of geminivirus genomes from tandemly repeated copies: evidence for rolling-circle replication of a plant viral DNA. Proc Natl Acad Sci U S A. 1991;88:8029–33.
Luna AP, Romero-Rodríguez B, Rosas-Díaz T, Cerero L, Rodríguez-Negrete EA, Castillo AG, Bejarano ER. Characterization of Curtovirus V2 Protein, a Functional Homolog of Begomovirus V2. Front Plant Sci. 2020;11:835.
Duffy S, Holmes EC. Phylogenetic evidence for rapid rates of molecular evolution in the single-stranded DNA begomovirus tomato yellow leaf curl virus. J Virol. 2008;82:957–65.
Duffy S, Holmes EC. Validation of high rates of nucleotide substitution in geminiviruses: phylogenetic evidence from East African cassava mosaic viruses. J Gen Virol. 2009;90:1539–47.
Lima ATM, Silva JCF, Silva FN, Castillo-Urquiza GP, Silva FF, Seah YM, Mizubuti ESG, Duffy S, Zerbini FM. The diversification of begomovirus populations is predominantly driven by mutational dynamics. Virus Evol. 2017;3:vex005.
Li P, Jing C, Ren H, Jia Z, Ghanem H, Wu G, Li M, Qing L. Analysis of Pathogenicity and Virulence Factors of Ageratum leaf curl Sichuan virus. Front Plant Sci. 2020;11:527787.
Sharma P, Ikegami M. Tomato leaf curl Java virus V2 protein is a determinant of virulence, hypersensitive response and suppression of posttranscriptional gene silencing. Virology. 2010;396:85–93.
Zhan B, Zhao W, Li S, Yang X, Zhou X. Functional Scanning of Apple Geminivirus Proteins as Symptom Determinants and Suppressors of Posttranscriptional Gene Silencing. Viruses. 2018;10:488.
Guerrero J, Regedanz E, Lu L, Ruan J, Bisaro DM, Sunter G. Manipulation of the Plant Host by the Geminivirus AC2/C2 Protein, a Central Player in the Infection Cycle. Front Plant Sci. 2020;11:591.
Huang CH, Foo MH, Raja JAJ, Tan YR, Lin TT, Lin SS, Yeh SD. A Conserved Helix in the C-Terminal Region of Watermelon Silver Mottle Virus Nonstructural Protein S Is Imperative For Protein Stability Affecting Self-Interaction, RNA Silencing Suppression, and Pathogenicity. Mol Plant Microbe Interact. 2020;33:637–52.
Wang L, Tian P, Yang X, Zhou X, Zhang S, Li C, Yang X, Liu Y. Key Amino Acids for Pepper Vein Yellows Virus P0 Protein Pathogenicity, Gene Silencing, and Subcellular Localization. Front Microbiol. 2021;12:680658.
Zhang C, Chen D, Yang G, Yu X, Wu J. Rice Stripe Mosaic Virus-Encoded P4 Is a Weak Suppressor of Viral RNA Silencing and Is Required for Disease Symptom Development. Mol Plant Microbe Interact. 2020;33:412–22.
Goodin MM, Dietzgen RG, Schichnes D, Ruzin S, Jackson AO. pGD vectors: versatile tools for the expression of green and red fluorescent protein fusions in agroinfiltrated plant leaves. Plant J. 2002;31:375–83.
Bräutigam A, Mullick T, Schliesky S, Weber AP. Critical assessment of assembly strategies for non-model species mRNA-Seq data and application of next-generation sequencing to the comparison of C(3) and C(4) species. J Exp Bot. 2011;62:3093–102.
Garg R, Patel RK, Tyagi AK, Jain M. De novo assembly of chickpea transcriptome using short reads for gene discovery and marker identification. DNA Res. 2011;18:53–63.
Su CL, Chao YT, Alex Chang YC, Chen WC, Chen CY, Lee AY, Hwa KT, Shih MC. De novo assembly of expressed transcripts and global analysis of the Phalaenopsis aphrodite transcriptome. Plant Cell Physiol. 2011;52:1501–14.
Huang X, Madan A. CAP3: A DNA sequence assembly program. Genome Res. 1999;9:868–77.
Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–10.
Kumar S, Stecher G, Li M, Knyaz C, Tamura K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol Biol Evol. 2018;35:1547–9.
Acknowledgements
We thank all the colleagues in our laboratory for providing useful discussions and technical assistance.
Funding
This work was funded by the Henan Science Fund for Excellent Young Scholars (202300410194), and Key Specialized Research and Development Breakthrough Project in Henan province (212102110451).
Author information
Authors and Affiliations
Contributions
Y. S., and Y. X. designed the experiments and wrote this manuscript. Q.-L.L., Y.-Y.Z. performed the experiments. W.-G.L., L.-L.Y., and X.-Y.H. assisted in the preparation of samples and analyses of data. Y.-J.S. and Y.-Q.L. critically reviewed the manuscript. L.-L.C., and H.-L.L. contributed to the data discussion, and corrected the manuscript. The author(s) read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The permission from Henan Agricultural University to collect the Nicotiana benthamiana plants documented in this work was obtained. The use of Nicotiana benthamiana plants during the experiment complies with national and international guidelines, and complies with local laws and regulations. The use of all plant materials does not bring any risk to other species in nature. Dr. Xue Yang undertook the formal identification of the Nicotiana benthamiana plants and the voucher specimen of this material has been deposited in the herbarium of Henan Agricultural University (Deposit no. Nb-061915).
Consent for publication
Not applicable
Competing interests
The authors declare no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Supplementary Table S1.
The statistics of primary assembly. Supplementary Table S2. The statistics of final assembly. Supplementary Table S3. List of PCR primers used in this work. Supplementary Fig. S1. The species expression abundance results based on the reads expression level of each species. Supplementary Fig. S2. Systemic symptoms induced by SGVA-ZZ in N. benthamiana plants were observed at 33 dpi. Supplementary Fig. S3. The multiple alignment of V2 amino acids of geminiviruses. Supplementary Fig. S4. Full scan date of the immunoblots in this work.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Li, Q., Zhang, Y., Lu, W. et al. Identification and characterization of a new geminivirus from soybean plants and determination of V2 as a pathogenicity factor and silencing suppressor. BMC Plant Biol 22, 362 (2022). https://doi.org/10.1186/s12870-022-03745-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-022-03745-z