
Abstract
Background
Canopy architecture is critical in determining the light environment and subsequently the photosynthetic productivity of fruit crops. Numerous CCT domain-containing genes are crucial for plant adaptive responses to diverse environmental cues. Two CCT genes, the orthologues of AtPRR5 in pear, have been reported to be strongly correlated with photosynthetic performance under distinct canopy microclimates. However, knowledge concerning the specific expression patterns and roles of pear CCT family genes (PbCCTs) remains very limited. The key roles played by PbCCTs in the light response led us to examine this large gene family in more detail.
Results
Genome-wide sequence analysis identified 42 putative PbCCTs in the genome of pear (Pyrus bretschneideri Rehd.). Phylogenetic analysis indicated that these genes were divided into five subfamilies, namely, COL (14 members), PRR (8 members), ZIM (6 members), TCR1 (6 members) and ASML2 (8 members). Analysis of exon–intron structures and conserved domains provided support for the classification. Genome duplication analysis indicated that whole-genome duplication/segmental duplication events played a crucial role in the expansion of the CCT family in pear and that the CCT family evolved under the effect of purifying selection. Expression profiles exhibited diverse expression patterns of PbCCTs in various tissues and in response to varying light signals. Additionally, transient overexpression of PbPRR2 in tobacco leaves resulted in inhibition of photosynthetic performance, suggesting its possible involvement in the repression of photosynthesis.
Conclusions
This study provides a comprehensive analysis of the CCT gene family in pear and will facilitate further functional investigations of PbCCTs to uncover their biological roles in the light response.
Similar content being viewed by others


Background
CCT [CONSTANS (CO), CONSTANS-LIKE (COL) and TIMING OF CAB1 (TOC1)] transcription factors, which encode proteins with a conserved motif (CCT domain) of ~ 43 amino acid residues towards their carboxy-terminus, constitute a plant-specific family [1, 2]. The CCT domain has important functions in nuclear localization, protein–protein interactions, transcriptional regulation and nuclear protein transport [3,4,5]. CCT family genes can be divided into five categories based on the sequencing information of Arabidopsis: COL, PRR (Pseudo-Response Regulator), ZIM (Zinc-finger Protein Expressed in Inflorescence Meristem), TCR1 (Tunicamycin-induced COL-Related 1) and ASML2 (Activator of Spomin::LUC2) subfamilies [6, 7]. The CCT family has been comprehensively analysed in some plants, including Arabidopsis, rice, maize, brachypodium, sorghum, foxtail millet, barley, Aegilops tauschii and Medicago truncatula [7,8,9,10,11], but not yet in woody perennial fruit crops.
It is well known that CCT family members play diverse and important roles in flowering, circadian rhythms, development and abiotic stress tolerance [12]. The COL subfamily has been extensively studied and can be further subdivided into three smaller groups according to the degree of conservation and number of B-box domains [13,14,15]. In addition to the CCT domain, COL proteins contain one or two zinc-finger B-box domains towards the amino terminus, which are thought to be involved in protein–protein interactions [16, 17]. The first cloned CCT family gene in Arabidopsis was AtCO, which is required to promote photoperiodic flowering, at least in part by activating the expression of the AtFT (flower time) and AtSOC1 (suppressor of overexpression of CO1) genes [18,19,20]. Other CO homlogues (COLs) have also been associated with abiotic stress tolerance, plant growth, development and metabolic processes, apart from playing a key role in photoperiodic flowering induction [21,22,23,24,25,26]. PRR subfamily genes contain a pseudoreceiver domain towards the amino terminus, as well as a CCT domain at the carboxy terminus [26]. PRRs are key genetic components of interconnected transcriptional-translational feedback loops that regulate circadian clock-output pathways [26,27,28,29]. In Arabidopsis, all PRR genes (AtPRR1, AtPRR3, AtPRR5, AtPRR7, and AtPRR9) that have redundant functions directly regulate the expression of genes implicated in abiotic responses, cell elongation, and photoperiodic flowering responses [26,27,28, 30]. In addition to the CCT domain, the ZIM subfamily contains both a C2C2-GATA zinc-finger domain and a TIFY domain [31, 32]. ZIM subfamily genes, including ZIM and ZIM-like (ZML) genes, are involved in hypocotyls and petiole elongation, cryptochrome 1-dependent responses to excess light and wound-induced lignification [33,34,35]. TCR1 and ASML2 subfamilies represent two distinct classes of the CCT family, and both encode proteins possessing just a single CCT domain [6, 7]. The AtTCR1 gene has been shown to be transcriptionally induced by Arabidopsis endoplasmic reticulum stress [7]. AtCIA2, AtCIL, and their barley homologues (HvCMF3 and HvCMF7) belong to the TCR1 subfamily and play critical roles in chloroplast development, thylakoid morphology, photosynthetic activity and various abiotic stress responses [36,37,38,39]. The functions of ASML2 genes are generally not well understood. Overexpression of AtASML2 results in enhanced expression of a subset of sugar-inducible genes in Arabidopsis [6].
As sessile organisms, plants need to depend on their ability to adapt to complex changes in the surrounding environment [40, 41]. As a consequence, they are equipped with sophisticated mechanisms that integrate environmental cues, such as light signalling (light quality, light intensity and photoperiod), and their endogenous regulators to optimize their growth and productivity [30, 42, 43]. There is increasing evidence that CCT genes are associated with light responses and/or photosynthetic capacity. Ghd7, the homologue of the CCT gene in rice, is a major locus that is responsible for natural variation in chlorophyll content at the heading stage [44]. Another CCT gene in rice, CRCT (CO2 responsive CCT protein), controls the capacity for starch synthesis, which can indirectly affect the photosynthetic rate under elevated CO2 conditions [45, 46]. In Arabidopsis, AtBBX4 is a CCT domain protein with an abundance that is positively modulated by phyB under red light, thereby promoting photomorphogenic development [47].
Pear is one of the most economically important fruit crops in the world. In the field, the canopy architecture of pear is critical in determining the light environment and thereby indirectly affects the source–sink relationship [48]. To understand the potential mechanism of pear photosynthetic variability responses to heterogeneous light environments within canopies, we conducted physiological and transcriptomic surveys to capture progressive stages of photosynthetic differentiation between distinct canopy structures [49]. We found that the two orthologues (LOC103943360 and LOC103951583) of AtPRR5 in pear were hub genes of the module positively correlated with pear photosynthetic rate and might play key roles in photosynthetic performance under distinct canopy microclimates [49]. Here, we address the important question of whether other members (PbCCTs) of the CCT family in pear could also participate in the regulation of photosynthesis and light signal response processes. However, little information is available on the identification and functional characterization of PbCCTs in pear, an important fruit crop. With the complete pear genome sequences of Pyrus bretschneideri publicly available [50], it is now possible to perform a genome-wide comprehensive analysis of PbCCTs. In this study, we identified PbCCTs and analysed the chromosomal locations, phylogenetic relationships and gene structure, as well as the expression patterns of some members in various tissues and in response to different light signals. In addition, transient expression analysis was used to investigate the possible roles of the PbPRR2 gene, a key CCT member, in response to light signals. This comprehensive study of the CCT family may provide valuable information for further research and utilization of PbCCTs, helping to enhance our understanding of the possible roles of PbCCTs in the adaptation of pear to changing ambient light signalling.
Methods
Genome-wide identification of CCT family genes
The hidden Markov model profile of the CCT domain (PF06203) downloaded from the Pfam database (http://pfam.xfam.org/) was used for identification of the PbCCTs from the downloaded database of the Pear Genome Project (http://peargenome.njau.edu.cn/) using the HMMER programme (version 3.1b2) with a threshold e-value < 10e−10 [50]. Using the same criterion, CCT family sequences from Arabidopsis thaliana (ftp://ftp.ensemblgenomes.org/pub/plants/release-38/fasta/arabidopsis_thaliana/), Oryza sativa (ftp://ftp.ensemblgenomes.org/pub/plants/release-38/fasta/oryza_sativa/), and two other Rosaceae species (GDR; https://www.rosaceae.org/), including apple and woodland strawberry (Fragaria vesca, diploid wild species), were obtained. To confirm the reliability of the search results, the obtained sequences were further examined based on the presence of conserved domains of CCT proteins using the InterProScan software package (version 5.25–64.0). Finally, a self-blast of protein sequences was performed to remove redundancy. Alternative splice variants were not considered. Any two protein sequences that showed a perfect match were deemed to be redundant gene pairs, and the shorter sequence was removed from the potential CCT gene list.
Analysis of synteny relationships, chromosomal locations, protein properties, gene structure and conserved motifs
The Multiple Collinearity Scan toolkit (MCScan X) was used to identify whole-genome duplication (WGD)/segmental and tandem duplications in the pear genome [51]. The KaKs_Calculator 2.0 was used to determine nonsynonymous (Ka) and synonymous (Ks) substitutions [52]. To exhibit the syntenic relationship of the orthologous CCT genes between pear and other selected species (apple/strawberry/Arabidopsis/rice), syntenic analysis was performed using MCScan X software. The physical location information for each CCT gene was retrieved from the pear/apple/strawberry genome database. The synteny relationships and location data among the three Rosaceae species (pear, apple and strawberry) were then plotted using TBtools software [53].
The molecular weights and theoretical isoelectric points of the PbCCT proteins were predicted by the compute pI/Mw tool in the ExPASY server (https://web.expasy.org/compute_pi/). The exon–intron structures of the PbCCTs were identified with the Gene Structure Display Server (GSDS, http://gsds.cbi.pku.edu.cn/) programme by the alignment of cDNA sequences with the corresponding genomic sequences. The InterProScan programme (http://www.ebi.ac.uk/interpro/) was used to characterize the domains and motifs of the pear CCT family.
Phylogenetic analysis
To investigate the phylogenetic relationship between pear and Arabidopsis, unrooted neighbour-joining (NJ) trees were constructed using MEGA7 software with 1000 bootstrap replicates [54]. The numbers generated for each clade represent the bootstrap support values expressed as percentages. The same method was adopted to construct the NJ phylogenetic tree for the five subfamilies of the pear CCT family.
Plant materials, growth conditions and treatments
To investigate the effects of light quality on PbCCT gene expression, one-year-old grafted seedlings of ‘Wonhwang’ (P. pyrifolia Nakai cv. ‘Wonhwang’) pear cultivar were obtained from the experimental orchard (30.292°N, 114.143°E) of the Research Institute of Fruit and Tea. Uniform and healthy plants were transplanted into plastic pots and cultured in an intelligent growth chamber (RLD-1000E-4, Le Electronic Instrument Co., Ltd., Ningbo, China) maintained at 25 °C and 70% relative humidity (RH). The commercially available light source used in this study was cool-red (R)/blue (B)/green (G) light-emitting diode (LED) panels. The distance between the lamps and pear leaves was ~ 20 cm. For the R light gradient treatments, fixed B light (1800 lx) and G light (5000 lx) intensities were also provided. The gradual increase in R light included six light intensity treatments, i.e., 500 lx (R500), 1000 lx (R1000), 1500 lx (R1500), 2000 lx (R2000), 2500 lx (R2500) and 3000 lx (R3000). For the B light gradient treatments, the seedlings were sequentially incubated in six B light intensity gradients, i.e., 1000 lx (B1000), 1500 lx (B1500), 2000 lx (B2000), 2500 lx (B2500), 3000 lx (B3000) and 3500 lx (B3500), while R light (2000 lx) and G light (5000 lx) were fixed light intensities. The fully expanded leaves from three individual plants were defined as three biological replicates. After light irradiation, leaf samples were harvested by rapid freezing in liquid nitrogen and stored at -80 °C until further use.
To test whether PbPRR2 could regulate photosynthetic performance in response to varying R light signals, PbPRR2 was transiently overexpressed in N. benthamiana leaves. One day after infiltration, N. benthamiana plants started to receive LED light treatment. The distance between the lamps and samples was ~ 30 cm. During the experiment, the temperature was maintained at 25 °C, while the RH was maintained at 70%. A gradient of R light was established, while B light (5000 lx) and G light (1800 lx) were relatively uniform. To determine the R light irradiance to which each leaf was exposed, light intensities (45–105 µmol m−2 s−1) were measured on the upper surface of each individual leaf using an LI-180 spectrometer (LI-COR Inc., USA).
Measurements of the light environment among different pear tree canopy positions
To investigate the light environment among different pear tree canopy positions, sunlight spectra were measured with an LI-180 spectrometer at 5 cm above the surface of the leaves in a specific location. Adult ‘Wonhwang’ pear trees (ten years old) were grown in the experimental orchard of the Research Institute of Fruit and Tea. The experiment was carried out in a randomized complete block design with three replications. Trees from each block were randomly selected, which represented biological replicates. Each tested tree was divided into four canopy positions, i.e., sunny side-interior part of the canopy, sunny side-exterior part of the canopy, shady side-interior part of the canopy, and shady side-exterior part of the canopy. The interior and exterior parts of the canopy were approximately 0–1.0 m and more than 1.0 m away from the trunk, respectively. For each biological replicate of each canopy position, light quality parameters were measured at three independent positions (three technical replicates). All sunlight spectra measurements were performed every 2 h from 08:00 to 16:00 on sunny and clear days (105 days after flowering). All data were analysed by one-way ANOVA (IBM SPSS Statistics 19 software) followed by Duncan’s multiple range tests with a significance level of P < 0.01.
Expression analysis of PbCCTs
To investigate the expression patterns of PbCCTs in different pear tissues, the normalized RPKM (reads per kilobase per million mapped reads) values of pear CCT genes were extracted from the previously published RNA-Seq data of leaf, ovary, petal, shoot, stigma, and fruit (15 days after full bloom) [55]. The results were visualized using a heatmap with transformed log2 (RPKM + 1) values using the ‘pheatmap’ R package (https://cran.r-project.org/web/packages/pheatmap/index.html).
To investigate the possible functions of the PbCCTs in varying light quality environments, PbCCTs with higher expression levels (RPKM values > 10) in leaves were selected for further analysis using qRT–PCR. Total RNA was extracted from the frozen leaves using the RNAprep Pure Plant Kit (Polysaccharides & Polyphenolics-rich) (Tiangen, China) according to the manufacturer’s protocol, followed by RNA integrity examination on 1.0% agarose gels stained with ethidium bromide. First-strand cDNA synthesis and qRT–PCR were performed as described previously [48]. Primer sequences for qRT–PCR analysis were designed using Primer Premier 5 (Additional file 1). Two reference genes, i.e., PbSKD1 and PbYLS8, which were shown to be stably expressed in pear leaves [48], were used as internal controls to normalize the qRT–PCR data. Relative quantification was calculated according to the Ct method (2−△△CT). For each sample, three independent biological replicates were performed to acquire reliable results. Statistical analyses were performed as described above.
Gene cloning, vector construction and transient transformation
The cDNA sequence of PbPRR2 was amplified from ‘Wonhwang’ pear leaf cDNA using gene-specific primers (Additional file 1) and then cloned into the pDONR221 vector by the BP reaction (Gateway, Invitrogen). Several independent clones were sequenced to confirm the correct sequence of PbPRR2 and then transferred by Gateway LR reaction into the destination vector pHEX2. All Gateway reactions were performed as recommended by the manufacturer (Thermo Fisher Scientific, https://www.thermofisher.com). Sequence alignment was performed using the Clustal X2 programme (http://www.clustal.org/) and GENEDOC. pHEX-GUS, pHEX-PbPRR2 and pBIN61-p19 (suppressor of gene silencing p19, which could dramatically enhance transient expression of a broad range of proteins) constructs were separately introduced into Agrobacterium tumefaciens strain GV3101 by the electroporation method [56]. A. tumefaciens cultures carrying each expression vector were mixed with an equal volume of A. tumefaciens strain containing pBIN61-p19 and coinfiltrated into N. benthamiana leaves as described by Hellens et al. (2005) [57]. Three days after infiltration, photosynthetic measurements were taken for 20 leaves per construct (n = 20) under different R light levels using the portable CIRAS-3 photosynthesis system (PP. Systems Inc., USA). Data are expressed as the mean ± SEM and were analysed using IBM SPSS Statistics 19 software. The differences between means were determined by the Student’s t test with a significance level of P < 0.001.
Results
Identification and genomic distribution of the CCT family in pear
In total, 42 PbCCTs were identified in the pear genome (Additional file 2). CCT family members were classified into COL, PRR, ZIM, TCR1 and ASML2 subfamilies and then systematically named according to their family name and sequence similarity. Our analysis revealed that the COL subfamily consisted of the highest number of CCT genes in pear, with 33.3% (14 PbCOLs) of the total PbCCTs (Additional file 2). Both PRRs and ASMLs constituted the second largest subfamily, with 19.0% (8 PbPRRs and 8 PbASML2s) of the PbCCTs. The ZIM and TCR1 subfamilies were the smallest, with 14.3% (6 PbZIMs and 6 PbTCR1s) of the PbCCTs. The molecular weight of these PbCCT proteins ranged from 22.8 kD to 93.6 kD, and their isoelectric point values were between 4.27 and 9.45.
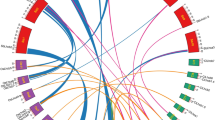
PbCCTs were unevenly distributed over 15 of the 17 pear chromosomes, with no PbCCT gene found on chromosomes 2 and 4 (Fig. 1). Among these, chromosomes 1, 3, 8 and 12 contained the fewest PbCCTs, with only one member (2.4%) on each chromosome, while chromosome 17 possessed the highest number of PbCCTs, with five (11.9%) of the 42 members. However, it should be noted that eight PbCCTs remained on unmapped scaffolds. Similar to that in PbCCTs, the putative CCT genes in the other two Rosaceae species (apple and strawberry) also exhibited random chromosomal distribution (Fig. 1 and Additional file 3).

Localization and synteny of CCT genes in pear and two other Rosacease species. Chromosomes of pear, apple and strawberry are shown in sky-blue, dark sea-green and goldenrod colours. The approximate positions of each CCT gene are marked with short grey lines on the circles. Gene pairs with syntenic relationships are connected by coloured lines
To clarify the potential mechanism of evolution of the PbCCT gene family, both WGD/segmental duplication and tandem duplication events were investigated in this study. Among all identified PbCCTs, a total of seven gene pairs were localized to WGD/segmentally duplicated regions, while there was no gene in tandem repeats (Additional file 4), indicating that WGD/segmental duplication events were the major contributors to the expansion of the pear CCT family. All members of the pear genome have undergone two genome duplication events, ancient WGD (Ks ~ 1.50–1.80) and recent WGD (Ks ~ 0.15–0.30) [50, 58]. The Ks values of two duplicated gene pairs were 0.162 and 0.207, implying that they might be derived from the relatively recent WGD event (approximately 30–45 MYA; MYA: million years ago); five duplicated gene pairs had smaller Ks values (0.004–0.041), suggesting that they might come from more recent segmental duplication events (Additional file 4). Moreover, with one exception (PbTCR5-PbTCR6), the Ka/Ks ratios of the other duplicated pairs were less than 0.26, implying that the pear CCT gene family had mainly undergone strong purifying selection (Additional file 4).
To further explore the synteny relationships of CCT family genes between pear (Rosaceae/Maloideae) and the other four representative species, Arabidopsis (dicot model plant), rice (monocot model plant), apple (Rosaceae/Maloideae) and strawberry (Rosaceae/Rosoideae), we performed interspecies comparative synteny analysis in a pairwise manner (Additional file 5). A total of 41, 33, 35 and 12 PbCCT genes were found to exhibit synteny relationships with CCT genes from apple, strawberry, Arabidopsis and rice, respectively, for example, PbASML1 (pear)—MD13G1220700/MD16G1225800 (apple)—FvH4_4g04500.1 (strawberry)—At2g33350 (Arabidopsis)—Os10t0466500-01 (rice). These results provide insights that will assist in the prediction of the possible roles of PbCCTs.
Phylogenetic analyses of CCT genes
To explore the phylogenetic relationship of the CCT family, an unrooted neighbour-joining phylogenetic tree was established based on the alignment of the full-length CCT protein sequences from pear and Arabidopsis (Fig. 2). In most clades, internal nodes were supported by confidence values of at least 80%, indicative of good consistency in the topology, which further corroborated the reliability of the tree. To test the reliability of the tree topology, protein domain architecture was used to provide additional support for the proposed phylogeny. The majority of members belonging to the same phylogenetic group exhibited common motif compositions (Fig. 3D). For example, TIFY and C2C2-GATA zinc-finger domains are specifically shared by the ZIM subfamily. The presence of the pseudoreceiver domain is also clade dependent in the PRR subfamily. The conserved intron/exon structural characteristics also supported the fine structure of the phylogenetic trees. For example, all the coding sequences of the PbPRRs were interrupted by 5 or 7 introns, while the TCR1 subfamily contained no more than two introns (Fig. 3C).

Phylogenetic relationships and subfamily designations in CCT proteins from pear and Arabidopsis based on the neighbour-joining method. The reliability of the predicted tree was tested by bootstrapping with 1000 replicates. The percentage of neighbour-joining bootstrap replications (> 40%) is shown above each node. These proteins were divided into seven clades and are represented by different colours

Phylogenetic relationships, expression profiles, gene structure and protein structure of pear CCT genes. A Neighbour-joining trees constructed for CCT genes from the ASML2, ZIM, PRR, COL and TCR1 families. B Heatmap showing the expression profiles of CCT genes in different tissues, including leaf, ovary, petal, shoot, stigma and fruit. The colour gradient from red to green indicates that expression values change from high to low. C Structure of CCT genes with exon(s) in green, UTR regions in blue, and solid lines between the coloured boxes indicating introns. The number indicates the phases of the corresponding introns. D Structures of CCT proteins with the CCT DNA binding domains represented by orange boxes, the zinc-finger B-box domain in royal blue, pseudoreceiver domain in aerugo, TIFY domain in green, C2C2-GATA zinc-finger domain in purple and leucine-rich repeat N-terminal domain represented by sky-blue boxes
According to the classification criteria of the CCT family in Arabidopsis, PbCCTs were classified into seven major clades, Clades I-VII (Fig. 2). To clarify the phylogenetic relationships, Clades IV, V, VI, and VII were further divided into three, two, two, and four subgroups, respectively. Remarkably, we found that all seven clades included genes from both pear and Arabidopsis, indicating that ancestral genes of the seven clades diverged before the differentiation of pear and Arabidopsis. Interestingly, one subgroup, i.e., Clade VIb, included only PbCCTs and not AtCCTs, implying that these genes might have specialized roles that were either lost in Arabidopsis or were acquired after divergence from their common ancestor.
All PbCOLs were categorized into three clades, with well-supported bootstrap values: five PbCOLs in Clade I, four PbCOLs in Clade II, and five PbCOLs in Clade III (Figs. 2 and 3A). Clade I comprised five PbCOLs (PbCOL1/4/5/8/9), featuring a conserved CCT domain with two upstream zinc-finger B-box domains (Fig. 3D). PbCOL members (PbCOL2/3/6/7) in Clade II exhibited one B-box domain and a CCT domain. The gene structures of Clades I and II were highly conserved, containing two exons and one intron (Fig. 3C). The PbCOL homologues (PbCOL10-PbCOL14) were clustered with Clade III of the Arabidopsis COL subfamily, which possesses a normal B-box domain, a second divergent B-box domain and a CCT domain (Fig. 2). We found that this phylogenetic classification of PbCOLs in Clades I and II was the same as the classification of the Arabidopsis COL subfamily based on the difference in the B-box domain (Figs. 2 and 3D). However, PbCOL10, PbCOL11 and PbCOL12 in Clade III contained only one or no B-box domain (Fig. 3D). These patterns suggested that the corresponding genes might have lost the B-box-type zinc finger domain.
The PRR subfamily was further divided into three main subgroups based on their phylogenetic relationship, which were designated Clades IVa, IVb and IVc (Fig. 2). Four members (PbPRR1/4/5/6) of Clade IVa and two members (PbPRR2/3) in Clade IVb were highly conserved, containing eight exons and seven introns (Fig. 3C). Clade IVc included two pear PRR genes (PbPRR7/8) that clustered with the AtPRR1 (AtTOC1) gene from the same branch (Fig. 2).
The TCR subfamily could be divided into two subgroups, i.e., Clade Va and Clade Vb (Fig. 2). For Clade Va, PbTCR1 and PbTCR4 were clustered with two AtTCR genes (At5g57180 and At4g25990). Clade Vb contained four pear members (PbTCR2/3/5/6), which were clustered with two AtTCR genes (AtTCR1 and At5g14370).
All pear ZIM genes were also divided into two subgroups (Fig. 2). In detail, Clade VIa comprised three pear PbZIM members (PbZML4/5/6), which clustered with Arabidopsis ZIM subfamily genes (AtZIM, AtZML1 and AtZML2), while three pear ZIM homologues (PbZML1/2/3) were identified as a distinct subgroup (Clade VIb) that had no counterpart in Arabidopsis.
Pear ASML2 members were classified into four subgroups and were characterized by only conserved CCT domains (Figs. 2 and 3D). Clades VIIa and VIIb were divided from the same branch and contained PbASML3/4 and PbASML7/8, respectively. PbASML1/2 were clustered to Clade VIIc, and PbASML5/6 belonged to Clade VIId.
Expression profiles of CCT genes in different tissues and under varying light signal environments
To investigate the tissue expression profiles of the PbCCTs in pear, we analysed their transcript levels based on publicly available RNA-seq data of different tissues, including leaf, ovary, petal, shoot, stigma and fruit (Fig. 3B). In general, the candidate PbCCTs showed variation in tissue expression patterns. Many PbPRRs and PbCOLs exhibited high transcript abundance levels in all six tissues, whereas most PbASMLs were expressed at relatively lower levels in multiple tissues. On the other hand, several PbCCTs exhibited tissue-specific expression. For example, PbCOL6 and PbTCR3 were mainly expressed in leaves, whereas PbCOL9 showed the highest transcript abundance in petals. Some duplicated gene pairs also showed divergent transcript levels. For instance, PbZML3 showed very low expression in six different tissues, whereas its duplicated gene, PbZML2, was highly expressed in all tested tissues.
We investigated the environmental light spectrum changes among different pear tree canopy positions. Compared with the exterior part of the canopy, the levels of R and B light decreased significantly in the interior part of the canopy, indicating that intensity changes in light quality were important signatures in fruit orchards (Additional file 6). Some genes from the CCT family have been shown to regulate growth and development by responding to R and B light signals [47, 59, 60]; therefore, the response of PbCCT induction to light quality treatments was further characterized by qRT–PCR. Overall, most of the selected PbCCTs showed highly diverse expression patterns under the enhancement of R/B light radiation (Fig. 4). These results suggested that they were sensitive to light quality signals, thus inducing different responses according to the external light conditions. Under the R light treatments, the expression of three genes (PbCOL6, PbTCR1, PbTCR2) reached a peak with low R light intensities (500 lx and 1000 lx) and was then downregulated during subsequent increased exposure to R light (Fig. 4). Additionally, PbCOL11 and PbTCR3 presented an increasing trend with increasing R light exposure. One homologous pair (PbPPR2 and PbPPR3) displayed strong rhythmic expression patterns, suggesting that these genes could respond to R light changes during their regulation of pear growth and development. Notably, compared with R light at 500 lx (R500), the abundance of PbPRR2 and PbTCR3 transcripts dramatically increased more than 13.5-fold under a light intensity of 2500 lx (R2500) and 107.6-fold under a light intensity of 3000 lx (R3000), respectively. We also analysed the expression pattern of the PbCCTs in the B light treatment (Fig. 4). Among them, the transcript levels of four genes (PbPPR2, PbPRR3, PbTCR1 and PbTCR3) were induced by enhanced B light, some of which decreased markedly at the highest abundance (B3500). Notably, the relative expression level of PbPPR2 increased approximately 208.9-fold at an intensity of 3000 lx (B3000) relative to the control and that of PbTCR3 increased approximately 172.0-fold at an intensity of 3500 lx (B3500). In addition, two PbCCTs (PbCOL5 and PbTCR4) showed decreased expression levels in response to progressively increasing B light signals.

Expression profiles of 16 selected PbCCT genes under varying light signal environments. The Y-axes show the relative gene expression level under blue light treatment (blue bars, left) and the relative gene expression level under red light treatment (red line, right). B1000/1500/2000/2500/3000/3500 (x-axis) indicate six blue light intensity gradients, i.e., 1000 lx, 1500 lx, 2000 lx, 2500 lx, 3000 lx and 3500 lx. R500/1000/1500/2000/2500/3000 (x-axis) indicate six red light intensity gradients, i.e., 500 lx, 1000 lx, 1500 lx, 2000 lx, 2500 lx and 3000 lx. Error bars indicate the standard deviation from three biological replicates by qRT–PCR analysis. Different capital letters indicate significant differences at P < 0.01
PbPRR2 might be involved in negatively regulating photosynthetic performance
Combining the previous transcriptomic study [49] and the present bioinformatics analysis and expression analysis, PbPRR2 (LOC103943360), a close homologue of the Arabidopsis circadian clock gene AtPRR5, was chosen as a strong candidate for functional verification. Because AtPRR5 is implicated in photomorphogenesis in R light, which is considered the most efficient wavelength for driving photosynthesis [61,62,63,64], these findings prompted us to investigate the potential role of PbPRR2 under a broad range of R light intensities. PbPRR2 has a 2013-bp open reading frame and encodes a protein of 670 amino acids (GenBank accession number: MZ826141). The amino acid sequences encoded by PbPRR2 and AtPRR5 (AT5G24470), the orthologue of PbPRR2 from Arabidopsis, were 40.12% identical (Additional file 7). The PbPRR2 protein featured a PR domain at the N-terminus and a CCT motif at the C-terminus.
To further test the possible role of PbPRR2 in the regulation of photosynthetic properties under changing R light signals, PbPRR2 was transiently overexpressed in N. benthamiana leaves and compared with the control leaves. Fluctuating profiles of net photosynthetic rate, stomatal conductance and internal CO2 were observed in both PbPRR2-infiltrated and control leaves with increasing R light intensity (Fig. 5). However, significantly reduced levels of these photosynthetic parameters were observed in leaves inoculated with the pHEX2-PbPRR2 construct compared with the pHEX2-GUS control. These observations indicated that PbPRR2 might suppress the red light-dependent enhancement of photosynthetic performance.

Photosynthetic performance of PbPRR2 transiently expressed in tobacco with increasing R light intensity. Leaves were infiltrated with PbPRR2 (red line) or the control pHEX2-GUS (blue line). Data for three key photosynthetic parameters, i.e., net photosynthetic rate (A), stomatal conductance (B) and internal CO2 (C), are presented as the mean ± SEM (n = 20). Asterisks indicate significant difference at P < 0.001
Discussion
Photosynthetic performance is an important agricultural trait vital to the adaptation of horticultural crops to ambient light conditions [65]. Although CCT genes are well-established regulators of photoperiodic flowering pathways and circadian rhythms, their regulatory roles in other light-dependent agricultural traits, including photosynthesis, merit further exploration [10, 12, 61, 66]. Understanding the structural characteristics of pear CCT family genes and their specific responses to different light environments will help to identify some important candidates that may be involved in canopy light signal regulation. Here, we performed a comprehensive analysis of the PbCCT family genes. The number of CCT genes in pear (42) and apple (49) was nearly twice that in strawberry (24) (Fig. 1). In the process of genome evolution, gene duplication is always involved in gene family expansion, either through WGD, segmental duplication or tandem duplication [67, 68]. None of the PbCCTs were from tandem duplication, and all PbCCT gene duplications in pear were caused by WGD/segmental duplication events (Additional file 4); similar results were also observed for other gene families in pear [69,70,71,72]. The more recent WGD event presumably occurred in the ancestor of Maloideae, but not in Rosoideae [73, 74], which likely resulted in the higher gene numbers of the CCT gene family in pear and apple. After gene duplication, some duplicates may undergo functional divergence. The Ka/Ks ratios of the PbCCT duplication pair indicated that purifying selection was a major force driving the evolution of new functions for PbCCTs.
Phylogenetic analysis showed that most of the CCT family clades contained both pear and Arabidopsis proteins, suggesting that the two species displayed relatively conserved evolution (Fig. 2). Gene expression patterns of PbCCTs in response to varying light quality could provide important clues regarding gene functions during the light response process. For instance, two PbCCTs (PbCOL5 and PbCOL7) showed progressively decreasing expression levels with increasing R light intensity (Fig. 4). AtCO1, an Arabidopsis orthologue of PbCOL5 (Fig. 2), is a key player in the induction of flowering [18]. AtCO protein was degraded under R light by Phytochrome B [75]. Our results showed that shaded light had a lower intensity of R/B light (Additional file 6). These findings suggested that increased PbCOL5 expression induced early flowering in response to the lower R light level to which pear shoots were exposed when shaded by neighbouring vegetation. Additionally, the expression of AtCOL7, which is a homologue gene of PbCOL7 in Arabidopsis (Fig. 2), was rapidly downregulated in response to high R:FR (red:far-red light) [60]. Overexpression of AtCOL7 was shown to enhance branching number under high R:FR conditions. It would be interesting to determine whether PbCOL7 could increase branching by perceiving decreasing R light signalling.
In Arabidopsis, AtCOL13 was shown to act as a positive regulator of R light-mediated inhibition of hypocotyl elongation [76]. The transcription level of PbCOL11, a homologue gene of AtCOL13, was upregulated robustly under the enhanced R light environment (Figs. 2 and 4), implying that PbCOL14 might act in response to R light signalling to regulate photomorphogenesis in pear.
Our results provide evidence indicating that PbPRR2 might have a novel role in the regulation of photosynthetic performance under varying light signal environments. Our phylogenetic analysis showed that the homologous pair (PbPRR2/PbPRR3) clustered together with Arabidopsis AtPRR5 (Fig. 2). The functions of AtPRR5 were implicated in the mechanisms underlying the control of flowering time and photomorphogenesis, as well as the circadian rhythm [61, 77]. Moreover, AtPRR5-ox plants were shown to be highly sensitive to continuous R light and seemed to be slightly hypersensitive to B light [77]. The “red or far-red light signalling pathway” was found to be an enriched category in AtPRR5 direct targets [26]. Here, we noticed that the expression of PbPRR2 and PbPRR3 showed robust free-running rhythms under continuously increasing R light (Fig. 4). Our previous study indicated that these two genes, which are involved in the ‘circadian rhythm-plant’ pathway, were strongly correlated with photosynthetic performance [49]. A key piece of evidence supporting the role of PbPRR2 in the light response was that it resulted in inhibition of photosynthetic performance compared with that of the control plants (Fig. 5). Therefore, PbPRR2 (together with PbPRR3) should be taken into consideration for a better understanding of the molecular links between circadian rhythms and light signalling-controlled photosynthetic performance.
Conclusions
In short, a total of 42 PbCCTs were identified in pear and divided into five subfamilies, as supported by phylogenetic relationships, intron–exon structures and conserved motifs. Expression analysis indicated that the PbCCT family might have diverse functions, and some members were sensitive to light induction, indicating that PbCCT genes were involved in light environmental adaptation. One CCT gene named PbPRR2 was indicated to be associated with negatively regulating photosynthetic performance under the enhanced R light environment. All the results presented in this study provide comprehensive information on the CCT family in pear and lay a foundation for further research on the function of the PbCCT gene.
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its supplementary information files.
Abbreviations
- Ka:
-
Nonsynonymous substitution
- Ks:
-
Synonymous substitution
- NJ:
-
Neighbour-joining
- RH:
-
Relative humidity
- R:
-
Red
- B:
-
Blue
- G:
-
Green
- LED:
-
Light-emitting diode
- RPKM:
-
Reads per kilobase per million mapped reads
- FR:
-
Far-red
- R500/1000/1500/2000/2500/3000:
-
Six red light intensity gradients, i.e., 500 lx, 1000 lx, 1500 lx, 2000 lx, 2500 lx and 3000 lx
- B1000/1500/2000/2500/3000/3500:
-
Six blue light intensity gradients, i.e., 1000 lx, 1500 lx, 2000 lx, 2500 lx, 3000 lx and 3500 lx
- WGD:
-
Whole-genome duplication
- MYA:
-
Million years ago
References
Li Y, Xu M. CCT family genes in cereal crops: a current overview. Crop J. 2017;5:449–58.
Mengarelli DA, Zanor MI. Genome-wide characterization and analysis of the CCT motif family genes in soybean (Glycine max). Planta. 2021;253:15.
Hotta CT, Xu X, Xie Q, Dodd AN, Johnson CH, Webb AAR. Are there multiple circadian clocks in plants? Plant Signal Behav. 2008;3:342–4.
Gangappa SN, Botto JF. The BBX family of plant transcription factors. Trends Plant Sci. 2014;19:460–70.
Kaewphalug W, Huehne PS, Sriboonlert A. Characterization of a CONSTANS-like gene from pigeon orchid (Dendrobium crumenatum Swartz) and its expression under different photoperiod conditions. Hortic J. 2017;86:252–62.
Masaki T, Tsukagoshi H, Mitsui N, Nishii T, Hattori T, Morikami A, et al. Activation tagging of a gene for a protein with novel class of CCT-domain activates expression of a subset of sugar-inducible genes in Arabidopsis thaliana. Plant J. 2005;43:142–52.
Iwata Y, Yamada T, Koizumi N. Transcriptional regulation of an Arabidopsis gene encoding a CCT domain-containing protein during endoplasmic reticulum stress. Plant Biotechnol. 2008;25:397–402.
Cockram J, Thiel T, Steuernagel B, Stein N, Taudien S, Bailey PC, et al. Genome dynamics explain the evolution of flowering time CCT domain gene families in the poaceae. PLoS One. 2012;7:e45307.
Zheng X, Li X, Ge C, Zhang J, Shi M, Chen J, et al. Characterization of the CCT family and analysis of gene expression in Aegilops tauschii. PLoS One. 2017;12:e0189333.
Jin M, Liu X, Jia W, Liu H, Li W, Peng Y, et al. ZmCOL3, a CCT gene represses flowering in maize by interfering circadian clock and activating expression of ZmCCT. J Integer Plant Biol. 2018;60:465–80.
Ma L, Yi D, Yang J, Liu X, Pang Y. Genome-wide identification, expression analysis and functional study of CCT gene family in Medicago truncatula. Plants. 2020;9:513.
Liu HY, Zhou XC, Li QP, Wang L, Xing YZ. CCT domain-containing genes in cereal crops: flowering time and beyond. Theor Appl Genet. 2020;133:1385–96.
Kurokura T, Samad S, Koskela E, Mouhu K, Hytönen T. Fragaria vesca CONSTANS controls photoperiodic flowering and vegetative development. J Exp Bot. 2017;68:4839–50.
Li YF, Zhao YQ, Zhang M, Jia GX, Zaccai M. Functional and evolutionary characterization of the CONSTANS-like family in Lilium×formolongi. Plant Cell Physiol. 2018;59:1874–88.
Song N, Xu Z, Wang J, Qin Q, Jiang H, Si W, et al. Genome-wide analysis of maize CONSTANS-LIKE gene family and expression profiling under light/dark and abscisic acid treatment. Gene. 2018;673:1–11.
Xu D, Jiang Y, Li J, Holm M, Deng XW. The B-box domain protein BBX21 promotes photomorphogenesis. Plant Physiol. 2018;176:2365–75.
Li J, Gao K, Yang X, Khan WU, Guo B, Guo T, et al. Identification and characterization of the CONSTANS-like gene family and its expression profiling under light treatment in Populus. Int J Bio Macromolecules. 2020;161:999–1010.
Suárez-López P, Wheatley K, Robson F, Onouchi H, Valverde F, Coupland G. CONSTANS mediates between the circadian clock and the control of flowering in Arabidopsis. Nature. 2001;410:1116–20.
An H, Roussot C, Suárez-López P, Corbesier L, Vincent C, Piñeiro M, et al. CONSTANS acts in the phloem to regulate a systemic signal that induces photoperiodic flowering of Arabidopsis. Development. 2004;131:3615–26.
Yoo SK, Chung KS, Kim J, Lee JH, Hong SM, Yoo SJ, et al. CONSTANS activates SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 through flowering locus T to promote flowering in Arabidopsis. Plant Physiol. 2005;139:770–8.
Min JH, Chung JS, Lee KH, Kim CS. The CONSTANS-like 4 transcription factor, AtCOL4, positively regulates abiotic stress tolerance through an abscisic acid-dependent manner in Arabidopsis. J Integr Plant Biol. 2015;57:313–24.
Liu J, Shen J, Xu Y, Li X, Xiao J, Xiong L. Ghd2, a CONSTANS-like gene, confers drought sensitivity through regulation of senescence in rice. J Exp Bot. 2016;67:5785–98.
Muntha ST, Zhang L, Zhou Y, Zhao X, Hu Z, Yang J, et al. Phytochrome A signal transduction 1 and CONSTANS-LIKE 13 coordinately orchestrate shoot branching and flowering in leafy Brassica juncea. Plant Biotechnol J. 2018;17:1333–43.
Ohmiya A, Oda-Yamamizo C, Kishimoto S. Overexpression of CONSTANS-like 16 enhances chlorophyll accumulation in petunia corollas. Plant Sci. 2019;280:90–6.
Cao D, Lin Z, Huang L, Damaris RN, Li M, Yang P. A CONSTANS-LIKE gene of Nelumbo nucifera could promote potato tuberization. Planta. 2021;253:65.
Nakamichi N, Kiba T, Kamioka M, Suzuki T, Yamashino T, Higashiyama T, et al. Transcriptional repressor PRR5 directly regulates clock-output pathways. Proc Natl Acad Sci USA. 2012;109:17123–8.
Farré EM, Liu T. The PRR family of transcriptional regulators reflects the complexity and evolution of plant circadian clocks. Curr Opin Plant Biol. 2013;16:621–9.
Liu TL, Newton L, Liu MJ, Shiu SH, Farré EM. A G-box-like motif is necessary for transcriptional regulation by circadian pseudo-response regulators in Arabidopsis. Plant Physiol. 2016;170:528–39.
Majeed N, Panigrahi KCS, Sukla LB, John R, Panigrapy M. Regulatory mechanisms across networks of the circadian clock and senescence pathways. J Plant Biochem Biotechnol. 2020;29:665–74.
Liu T, Carlsson J, Takeuchi T, Newton L, Farré EM. Direct regulation of abiotic responses by the Arabidopsis circadian clock component PRR7. Plant J. 2013;76:101–14.
Zhang L, You J, Chan Z. Identification and characterization of TIFY family genes in brachypodium distachyon. J Plant Res. 2015;128:995–1005.
Saha G, Park JI, Kayum MA, Nou IS. A genome-wide analysis reveals stress and hormone responsive patterns of TIFY family genes in Brassica rapa. Front Plant Sci. 2016;7:936.
Shikata M, Takemura M, Yokota A, Kohchi T. Arabidopsis ZIM, a plant-specific GATA factor, can function as a transcriptional activator. Biosci Biotechnol Biochem. 2003;67:2495–7.
Shaikhali J, Barajas-Lopéz JD, Ötvös K, Kremnev D, Garcia AS, Srivastava V, et al. The CRYPTOCHROME1-dependent response to excess light is mediated through the transcriptional activators ZINC FINGER PROTEIN EXPRESSED IN INFLORESCENCE MERISTEM LIKE1 and ZML2 in Arabidopsis. Plant Cell. 2012;24:3009–25.
Vélez-Bermúdez IC, Salazar-Henao JE, Fornalé S, López-Vidriero I, Franco-Zorrilla JM, Grotewold E, et al. A MYB/ZML complex regulates wound-induced lignin genes in maize. Plant Cell. 2015;27:3245–59.
Li M, Hensel G, Mascher M, Melzer M, Budhagatapalli N, Rutten T, et al. Leaf variegation and impaired chloroplast development caused by a truncated CCT domain gene in albostrians barley. Plant Cell. 2019;31:1430–45.
Li M, Hensel G, Melzer M, Junker A, Tschiersch H, Arend D, et al. Mutation of the ALBOSTRIANS ohnologous gene HvCMF3 impairs chloroplast development and thylakoid architecture in barley due to reduced plastid translation. bioRixv. 2019; https://doi.org/10.1101/756833.
Gawroński P, Burdiak P, Scharff LB, Mielecki J, Górecka M, Zaborowska M, et al. CIA2 and CIA2-LIKE are required for optimal photosynthesis and stress responses in Arabidopsis thaliana. Plant J. 2021;105:619–38.
Li M, Ruwe H, Melzer M, Junker A, Hensel G, Tschiersch H, et al. The Arabidopsis AAC proteins CIL and CIA2 are sub-functionalized paralogs involved in chloroplast development. Front Plant Sci. 2021;12:681375.
Valim HF, McGale E, Yon F, Halitschke R, Fragoso V, Schuman MC, et al. The clock gene TOC1 in shoots, not roots, determines fitness of nicotiana attenuata under drought. Plant Physiol. 2019;181:305–18.
Lyu G, Li D, Li S. Bioinformatics analysis of BBX family genes and its response to UV-B in Arabidopsis thaliana. Plant Signal Behav. 2020;15:9.
Wang S, Zhang C, Zhao J, Li R, Lv J. Expression analysis of four pseudo-response regulator (PRR) genes in chrysanthemum morifolium under different photoperiods. Peer J. 2019;7:e6420.
Huq E. Direct convergence of light and auxin signaling pathways in Arabidopsis. Mol Plant. 2018;11:515–7.
Wang Q, Xie W, Xing H, Yan J, Meng X, Li X, et al. Genetic architecture of natural variation in rice chlorophyll content revealed by a genome-wide association study. Mol Plant. 2015;8:946–57.
Morita R, Sugino M, Hatanaka T, Misoo S, Fukayama H. CO2-responsive CONSTANS, CONSTANS-like, and time of Chlorophyll a/b binding protein expression1 protein is a positive regulator of starch synthesis in vegetative organs of rice. Plant Physiol. 2015;167:1321–31.
Morita R, Inoue K, Ikeda KI, Hatanaka T, Misoo S, Fukayama H. Starch content in leaf sheath controlled by CO2-responsive CCT protein is a potential determinant of photosynthetic capacity in rice. Plant Cell Physiol. 2016;57:2334–41.
Heng Y, Jiang Y, Zhao X, Zhou H, Wang X, Deng XW, et al. BBX4, a phyB-interacting and modulated regulator, directly interacts with PIF3 to fine tune red light-mediated photomorphogenesis. Proc Natl Acad Sci USA. 2019;116:26049–56.
Liu Z, Cheng K, Qin Z, Wu T, Li X, Tu J, et al. Selection and validation of suitable reference genes for qRT-PCR analysis in pear leaf tissues under distinct training systems. Plos One. 2018;13:e0202472.
Liu Z, An L, Lin S, Wu T, Li X, Tu J, et al. Comparative physiological and anscriptomic analysis of pear leaves under distinct training systems. Sci Rep. 2020;10:18892.
Wu J, Wang Z, Shi Z, Zhang S, Ming R, Zhu S, et al. The genome of the pear (Pyrus bretschneideri Rehd.). Genome Res. 2013;23:396–408.
Wang Y, Tang H, DeBarry JD, Tan X, Li J, Wang X, et al. MCScan X: a toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012;40:e49.
Wang D, Zhang Y, Zhang Z, Zhu J, Yu J. KaKs_Calculator 2.0: a toolkit incorporating gamma-series methods and sliding window strategies. Genomics Proteomics Bioinformatics. 2010;8:77–80.
Chen C, Chen H, Zhang Y, Thomas HR, Frank MH, He Y, et al. TBtools: an integrative toolkit developed for interactive analyses of big biological data. Mol Plant. 2020;13:1194–202.
Kumar S, Stecher G, Tamura K. MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol Biol Evol. 2016;33:1870–4.
Zhang S, Ma M, Zhang H, Zhang S, Qian M, Zhang Z, et al. Genome-wide analysis of polygalacturonase gene family from pear genome and identification of the member involved in pear softening. BMC Plant Biol. 2019;19:587.
Voinnet O, Rivas S, Mestre P, Baulcombe D. An enhanced transient expression system in plants based on suppression of gene silencing by the p19 protein of tomato bushy stunt virus. Plant J. 2003;33:949–56.
Hellens RP, Allan AC, Friel EN, Bolitho K, Grafton K, Templeton MD, et al. Transient expression vectors for functional genomics, quantification of promoter activity and RNA silencing in plants. Plant Methods. 2005;1:13.
Fawcett JA, Maere S, Van de Peer Y. Plants with double genomes might have had a better chance to survive the cretaceous-tertiary extinction event. Proc Natl Acad Sci USA. 2009;106:5737–42.
Sawa M, Nusinow DA, Kay SA, Imaizumi T. FKF1 and GIGANTEA complex formation is required for day-length measurement in Arabidopsis. Science. 2007;318:261–5.
Wang H, Zhang Z, Li H, Zhao X, Liu X, Ortiz M, et al. CONSTANS-LIKE 7 regulates branching and shade avoidance response in Arabidopsis. J Exp Bot. 2013;64:1017–24.
Yamamoto Y, Sato E, Shimizu T, Nakamich N, Sato S, Kato T, et al. Comparative genetic studies on the APRR5 and APRR7 genes belonging to the APRR1/TOC1 quintet implicated in circadian rhythm, control of flowering time, and early photomorphogenesis. Plant Cell Physiol. 2003;44:1119–30.
Kato T, Murakami M, Nakamura Y, Ito S, Nakamichi N, Yamashino T, et al. Mutants of circadian-associated PRR genes display a novel and visible phenotype as to light responses during de-etiolation of Arabidopsis thaliana seedlings. Biosci Biotechnol Biochem. 2007;71:834–9.
Izzo LG, Mele BH, Vitale L, Vitale E, Arena C. The role of monochromatic red and blue light in tomato early photomorphogenesis and photosynthetic traits. Environ Exp Bot. 2020;179:104195.
Liu J, van Iersel MW. Photosynthetic physiology of blue, green, and red light: light intensity effects and underlying mechanisms. Front Plant Sci. 2021;12:619987.
Fu QS, Zhao B, Wang YJ, Ren S, Guo YD. Stomatal development and associated photosynthetic performance of capsicum in response to differential light availabilities. Photosynthetica. 2010;48:189–98.
Lee YS, Jeong DH, Lee DY, Yi J, Ryu CH, Kim SL, et al. OsCOL4 is a constitutive flowering repressor upstream of Ehd1 and downstream of OsphyB. Plant J. 2010;63:18–30.
Su W, Ren Y, Wang D, Su Y, Feng J, Zhang C, et al. The alcohol dehydrogenase gene family in sugarcane and its involvement in cold stress regulation. BMC Genomics. 2020;21:521.
Wang J, Wang Z, Jia C, Miao H, Zhang J, Liu J, et al. Genome-wide identification and transcript analysis of TCP gene family in Banana (Musa acuminata L.). Biochem Genet. 2021;60(1):204. https://doi.org/10.1007/s10528-021-10100-8.
Cao Y, Han Y, Li D, Lin Y, Cai Y. MYB transcription factors in Chinese pear (Pyrus bretschneideri Rehd.): genome-wide identification, classification, and expression profiling during fruit development. Front Plant Sci. 2016;7:577.
Abdullah M, Cao Y, Cheng X, Meng D, Chen Y, Shakoor A, et al. The sucrose synthase gene family in Chinese pear (Pyrus bretschneideri Rehd.): structure, expression, and evolution. Molecules. 2018;23:1144.
Wang L, Wang L, Zhang Z, Ma M, Wang R, Qian M, et al. Genome-wide identification and comparative analysis of the superoxide dismutase gene family in pear and their functions during fruit ripening. Postharvest Biol Technol. 2018;143:68–77.
Wang Y, Lü J, Chen D, Zhang J, Qi KJ, Cheng R, et al. Genome-wide identification, evolution, and expression analysis of the KT/HAK/KUP family in pear. Genome. 2018;61:755–65.
Qiao X, Li Meng, Li L, Yin H, Wu J, Zhang S. Genome-wide identification and comparative analysis of the heat shock transcription factor family in Chinese white pear (Pyrus bretschneideri) and five other Rosaceae species. BMC Plant Biol. 2015;15:12.
Tang C, Qiao X, Zhu X, Khan W, Wu J, Zhang S. Expression and evolutionary analysis of soluble inorganic pyrophosphatase gene family in pear and four other Rosaceae species. Plant Syst Evol. 2020;306:46.
Valverde F, Mouradov A, Soppe W, Ravenscroft D, Samach A, Coupland G. Photoreceptor regulation of CONSTANS protein in photoperiodic flowering. Science. 2004;303:1003–6.
Liu B, Long H, Yan J, Ye L, Zhang Q, Chen H, et al. A HY5-COL3-COL13 regulatory chain for controlling hypocotyl elongation in Arabidopsis. Plant Cell Environ. 2021;44:130–42.
Sato E, Nakamichi N, Yamashino T, Mizuno T. Aberrant expression of the Arabidopsis circadian-regulated APRR5 gene belonging to the APRR1/TOC1 quintet results in early flowering and hypersensitiveness to light in early photomorphogenesis. Plant Cell Physiol. 2002;43:1374–85.
Acknowledgements
Not applicable
Funding
This work was financially supported by the National Key R&D Program of China (no. 2018YFD1000200, 2020YFD1000202), the China Agriculture Research System of MOF and MARA (no. CARS-28–11), the Major Program of Hubei Agricultural Science and Technology Innovation Center (no. 2020–620-000–002-05), and the National Natural Science Foundation of China (nos. 31801819). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
ZL and TW conceived and designed the research. ZL, JLL and LA performed the experiments. ZL, LY, YSC and XSN analyzed the data. ZL wrote the paper. ZQQ revised the paper. All authors have read and approved the manuscript for publication.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
The experiments did not involve endangered or protected species. The data collection of plants was carried out with permission of related institution, and complied with national or international guidelines and legislation.
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
List of primers used inthis study.
Additional file 2.
Information on PbCCT genes identified in the genome of pear.
Additional file 3.
Lists of CCT genes identified in apple,strawberry, Arabidopsis and rice.
Additional file 4.
Ka/Ks calculation of theduplicated PbCCT gene pairs in pear.
Additional file 5.
One-to-one orthologousrelationships between pear and apple/strawberry/Arabidopsis/rice.
Additional file 6.
Comparison of diurnalcourses of red light and blue light measured from different canopy positions. All measurements weretaken every 2 h between 08:00 and 16:00. Eachvalue represents the mean ± SEM (n = 9). The capital letters above the bars indicatesignificant differences (P < 0.01).
Additional file 7.
Alignment of the amino acid sequences of PbPRR2 and AtPRR5. The redframe indicates the relatively conserved pseudoreceiver domain and CCT domain.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Liu, Z., Liu, JL., An, L. et al. Genome-wide analysis of the CCT gene family in Chinese white pear (Pyrus bretschneideri Rehd.) and characterization of PbPRR2 in response to varying light signals. BMC Plant Biol 22, 81 (2022). https://doi.org/10.1186/s12870-022-03476-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-022-03476-1