
Abstract
Background
Suboptimal root zone temperature (RZT) causes a remarkable reduction in growth of horticultural crops during winter cultivation under greenhouse production. However, limited information is available on the effects of suboptimal RZT on nitrogen (N) metabolism in cucumber seedlings. The aim of this study is to investigate the effects of 24-Epibrassinolide (EBR) on nitrate and ammonium flux rate, N metabolism, and transcript levels of NRT1 family genes under suboptimal RZT in cucumber seedlings.
Results
Suboptimal RZT (LT) negatively affected on cucumber growth and proportionately decreased EBR contents, bleeding rate, root activity, enzyme activities of nitrate reductase (NR), nitrite reductase (NiR), glutamine synthetase (GS), and glutamate synthase (GOGAT), nitrate (NO3−) influx rate, ammonium (NH4+) efflux rate, and transcript levels of nitrate transporter (NRT1) encoding genes. However, exogenous EBR reduced the harmful effects of suboptimal RZT and increased endogenous EBR contents, bleeding rate, root activity, enzyme activities of NR, NiR, GS, and GOGAT, NH4+ and NO3− flux rates and contents, and N accumulation. EBR-treated seedlings also upregulated the transcript levels of nitrate transporters CsNRT1.1, CsNRT1.2A, CsNRT1.2B, CsNRT1.2C, CsNRT1.3, CsNRT1.4A, CsNRT1.5B, CsNRT1.5C, CsNRT1.9, and CsNRT1.10, and downregulated CsNRT1.5A and CsNRT1.8. LT treatment upregulated the expression level of CsNRT1.5A, while exogenous BZR application downregulated the expression level of NRT1 genes.
Conclusion
These results indicate that exogenous application of EBR alleviated the harmful effects of suboptimal RZT through changes in N metabolism, NH4+ and NO3− flux rates, and NRT1 gene expression, leading to improved cucumber seedlings growth. Our study provides the first evidence of the role of EBR in the response to suboptimal RZT in cucumber, and can be used to improve vegetable production.
Similar content being viewed by others


Background
Environmental factors influence plant growth and developments, and temperature is particularly important. Air temperature is unstable, while root zone temperature (RZT) is considered both stable and more important for study [1]. Temperature and light intensity are key factors affecting plant growth and development [2], which are also influenced by humidity and nutrient availability [3, 4]. Among these factors temperature is very important, that effects overall plant developmental process in short time [3, 7]. The previous studies reported that low temperature stress leads to over production of reactive oxygen species (ROS) and reduce antioxidant enzyme activates, reduction in chlorophylls and photosynthetic capacity, hormonal imbalance, ion uptake and accumulation, thus caused a significant reduction in plant growth and yield [2, 4, 6, 8]. Physiological and molecular studies have shown that plant growth is affected by RZT [3]. RZT influences physiological and biological processes, thereby affecting nutrient uptake and availability [5, 6]. Plant nutrient absorption rate is dependent on RZT [7], and can alter ion balance and nitrogen metabolism [8]; a small increase in RZT could induce large changes in plant growth and development [6].
During winter cultivation, air temperature is unstable, while soil temperature changes slowly and is maintained around 10–15 °C [9]. RZT plays a critical role in plant root physiology, morphology, growth, nutrient and water uptake, and translocation from root to leaf. Even horticultural crops exposed to suboptimal RZT may experience heavy losses of early productivity [3, 5,6,7]. Earlier studies reported that RZT severely affected nitrogen metabolism in cucumber and reduced growth and yield [2]. These studies demonstrated the importance of RZT on plant growth and development. The potential mechanisms of growth inhibition at ambient RZT may involve water and nutrient uptake rates but are largely unknown.
Nitrogen (N) is an essential macronutrient and its availability in soil affects plant growth and development, as well as all metabolic processes [10]. N is a major constituent of proteins and nucleotides, as well as of chlorophyll, numerous metabolites, and cellular components [11]. Nutrient availability and uptake affect plant growth and development [2]. Ammonium (NH4+) and nitrate (NO3−) are the principal soil N sources for plants [12]. Plant fine roots absorb NO3− and assimilate NH4+ into organic N via the GOGAT enzyme, once inside root cells, nitrate (NO3−) can be reduced to ammonium (NH4+) by nitrate and nitrite reductase and then assimilated into organic nitrogen through the glutamine synthase (GS)-GOGAT cycle [13,14,15,16]. Plants take up nitrate and transport it across the specialized plasma membrane made of root epidermal and cortical cells through a complex transport system [17, 18]. The mechanisms by which nitrate influx and efflux occur have been characterized at both the physiological and molecular levels [13, 19]. Plant cells consist of two nitrate uptake systems; one is a low-affinity system, either constitutive low-affinity system (cLATS) or inducible low affinity transport system (iLATS), which are encoded by NRT1 genes; the other is a high affinity transport system, either constitutive high affinity transport system (cHATS) or inducible high affinity transport system (iHATS), which are encoded by NRT2 genes [12, 20]. In Arabidopsis, eleven NRT1 and seven NRT2 gene homologues have been identified, but a limited number are considered responsible for nitrate uptake from soil [13, 21]. In iHATS, NRT2.1 and 2.2 and NRT1.1 in iLATS appear to play a major role in NO3− influx [11, 17]. LATS and HATS are involved in root xylem loading and unloading of nitrate (AtNRT1.5 and AtNRT1.8) and transport into the leaf [12, 17].
Brassinosteroids (EBRs) are growth-promoting steroid phytohormones in plants [22, 23]. EBRs play vital roles in a wide range of developmental processes in plants from germination to fruit development [24, 25]. Exogenous application of EBRs regulates a variety of physiological, biochemical, and molecular processes which enhance plant tolerance to a variety of abiotic stresses, such as low temperatures, heavy metals, and drought [25, 26]. 24-epibrassinolide (EBR) is the most active synthetic analog of the EBR family and can improve tolerance of low temperatures in pepper, tomato, eggplant, cucumber, and ryegrass [22, 27,28,29,30]. The mechanism of EBR activity in plant responses to abiotic stress has been reported [25, 31,32,33]. EBR promotes plant tolerance to heat, cold, drought, and salinity by correlating with higher expression of stress marker genes, including heat shock proteins (HSPs) and cold responsive genes (COR) [25, 29, 34]. EBR Plant exposed to low/cold stress caused negative effects on chlorophyll, photosynthesis, nutrients accumulation and antioxidant enzyme activity, thus leads to reduced plant growth and yield. The pervious study reported that chilling stress downregulate thousands of genes in involved in many developmental process, including chlorophyll and photosynthesis, antioxidant enzymes, hormones and transcriptional factors, while exogenous EBR application reduce the negative effect of chilling/cold stress on pepper seedling [29]. Cold stress caused a significant reduction in antioxidant enzyme activities and increase ROS (reactive oxygen species) accumulation, thus leads to reduce chlorophylls and photosynthetic capacity, while exogenous EBR application reduce the harmful effects and improve growth [37]. These findings are suggested that, EBR enhances biosynthesis of chlorophyll and photosynthetic machinery and activates stress tolerance enzymes, thus reduce the harmful effects of abiotic stresses [35,36,37]. A previous study reported that EBR regulated nitrogen uptake and metabolism in Arabidopsis via the EBR signaling pathway [23, 38]. Additionally, EBR receptor BRI1 (BRASSINOSTEROID INSENSITIVE 1) mutant bri1–5 induced expression of AMT1 (ammonium transporter 1) and GS and GOGAT encoded genes, showing that EBR signaling transcription factors BES1 (BRI1-EMS SUPPERSSOR 1) and BZR1 (BRASSINAZOLE RESISTANT 1) are involved in pathways of EBR-mediated nitrogen metabolism and uptake [23, 39,40,41]. A recent study reported that EBR enhanced low temperature and weak light stress tolerance in tomato, by improving nitrogen metabolism, stimulating nitrate and ammonium accumulation, and accelerating nitrogen conversion into free amino acids [28]. These amino acids are involved in biosynthesis of chlorophylls, proteins, primary and secondary metabolites, and enzyme biosynthesis [38]. These findings suggest an active role for EBR in stress and in nitrogen uptake and metabolism to reduce the harmful effect of stress. However, little is known about the role of EBR in nitrate and ammonium ion influx and in regulation of nitrogen metabolism under suboptimal RZT. This study will comprehensively determine the role of EBR in nitrogen uptake, metabolism, and accumulation under suboptimal RZT in cucumber seedlings.
Cucumber, which is widely grown in greenhouses in northern parts of China during summer and winter seasons, is intolerant to suboptimal RZT, leading to large yield losses [2, 9]. Therefore, suboptimal RZT is a major limiting factor for winter cultivation of cucumber in greenhouses [9, 42, 43]. In this study, we investigated the effect of EBR on cucumber seedling physiology and growth under suboptimal RZT. We hypothesized that exogenous EBR applied to leaves may enhance cucumber seedling growth by increasing enzyme activities and expression of genes involved in nitrogen metabolism as well as regulating nutrient uptake (ion influx rate). The key objectives of this study were to: (1) investigate the effect of RZT on plant physiology; (2) examine whether exogenous EBR application can effectively enhance nitrogen metabolism and uptake rate (ion influx rate); and (3) examine whether exogenous EBR regulates NRT1 expression in cucumber. The results could improve understanding of the role of EBR in nitrogen metabolism, uptake, and response to RZT, which is useful for greenhouse vegetable production.
Results
Effect of EBR on cucumber seedlings growth under suboptimal RZT
Cucumber seedlings growth and growth-related parameters were investigated seven days after exposure to various treatments of suboptimal RZT and EBR, as presented in Table 1. Suboptimal RZT significantly reduced the growth of cucumber seedlings (Table 1). Briefly, plant height, hypocotyl dimeter, leaf area, total fresh weight and seedling index were decreased by 26.59, 29.43, 28.71, 38.04 and 39.18% respectively, in the LT treatment compared to the NT treatment, but decreased by 31.40, 27.54, 32.75, 38.19, and 33.89% respectively, when compared to EBR treated seedlings. The differences between NT and EBR treatments were not significant. Moreover, the plant height, root fresh weight, shoot fresh weight, and total fresh weight of cucumber seedlings in the EBR treatment were significantly higher than in the LT and BZR treatments. These results suggest that EBR alleviated the harmful effects of suboptimal RZT temperature, thus leading to improved cucumber seedlings growth.
Effect of exogenous EBR application on endogenous accumulation of EBR
The endogenous EBR contents were measured in cucumber leaves seven days after exposure to suboptimal RZT. As shown in Fig. 1, the LT (control; suboptimal RZT) treatment significantly decreased endogenous EBR accumulation in the leaf by 28.24% compared with the control (NT, normal RZT). Moreover, exogenous EBR application significantly increased endogenous EBR accumulation in cucumber leaf by 43.34 and 58.09% compared with LT and BZR treatments, respectively, under suboptimal RZT, while it increased by 26.66% over that of the NT treatment (normal RZT).

Changes in endogenous EBR contents in leaves of cucumber seedlings after suboptimal RZT with EBR and BZR application. Leaf samples were harvested seven days after treatment. Data are the means of four replicates with standard deviation (SD) shown by vertical bars on top of columns. Columns with the same lowercase letter are not significantly different by the least significant difference (LSD) test at P = 0.05
Effect of EBR on root activity and bleeding rate
Root activity is an important parameter for suboptimal RZT because it reflects the strength of overall metabolic processes in root tissues including respiration, oxidation, and enzymatic activities involved in nutrient uptake. Suboptimal RZT caused a significant reduction in root activity of cucumber seedlings, but these increased significantly in the EBR treatments (Fig. 2 A. Briefly, the EBR treatment increased root activity by 17.05, 35.98, and 32.76%, when compared with the NT, LT, and BZR treatments, respectively. Additionally, the LT and BZR treatments negatively affected the root activity of cucumber seedlings, with reductions of 25.07 and 21.29%, respectively, compared to the NT treatment.

Changes in root activity (A) and bleeding rate (B) cucumber seedling after seven days under suboptimal RZT with EBR and BZR application. Treatments with the same lowercase letter are not significantly different by the least significant difference (LSD) test at P = 0.05
Suboptimal RZT also negatively affected bleeding rate (Fig. 2 B. In the LT treatment, bleeding rate was reduced by 46.73% compared to that of the NT treatment, while exogenous EBR application significantly increased bleeding rate by 47.31, and 55.65%, compared to that of the LT and BZR treatments (Fig. 2 B). The bleeding rate did not differ detectably between the NT and EBR treatments. These findings suggest that suboptimal RZT negatively affected root activity and bleeding rate and caused a significant reduction in growth.
Effect of EBR on NR, NiR, GS and GOGAT enzyme activities
Nitrate reductase (NR), nitrite reductase (NiR), glutamine synthetase (GS), and glutamate synthase (GOGAT) are key enzymes involved in nitrogen metabolism. We investigated these enzymes in leaves of cucumber seedlings after seven days of suboptimal RZT. The enzymes involved in nitrogen metabolism (NR, NiR, GS, and GOGAT) were adversely affected by suboptimal RZT (LT), when compared to the NT and EBR treatments, as presented in Fig. 3. Exogenous EBR significantly increased the activities of NR, NiR, GS, and GOGAT under suboptimal RZT, as compared to the LT treatments, but differences between the NT and EBR treatments were not significant (Fig. 3). Glutamine synthetase activity was significantly higher in the EBR treatment compared to the NT treatment (Fig. 3 B). These findings suggested that EBR regulates the activities of NR, NiR, GS, and GOGAT under suboptimal RZT in cucumber seedlings.

Effect of EBR on A; nitrate reductase (NR), B; glutamine synthetase (GS), C; glutamate synthase (GOGAT), and D; nitrite reductase (NiR) activities in leaves of cucumber seedlings under suboptimal RTZ. Treatments with the same lowercase letter are not significantly different by the least significant difference (LSD) test at P = 0.05
Effect of EBR on NH4 + and NO3 − fluxes rate
The NH4+ and NO3− flux rates were investigated in the roots of cucumber seedlings seven days after exposure to suboptimal RZT. Suboptimal RZT negatively affected NH4+ and NO3− flux rates, but exogenous EBR application significantly increased NO3− and NH4+ flux rates in root of cucumber seedlings (Figs. 4 & 5).

Effects of suboptimal RZT and EBR on NO3− (A; scatter NO3− flux rate, B; average NO3− flux rate) flux rate in roots of cucumber seedlings under suboptimal RZT treatment. Flux rate was recorded for 10 min in roots seven days after treatment. Each point is the mean of nine individual seedlings and bars indicate standard deviations. Treatments with the same letters are not significantly different by the least significant difference (LSD) test at P = 0.05

Effects of suboptimal RZT and EBR on NH4+ (A; scatter NH4+ flux rate, B; average NH4+ flux rate) flux rate in roots of cucumber seedlings under suboptimal RZT treatment. Flux rate was recorded for 10 min in roots seven days after treatment. Each point is the mean of nine individual seedlings and bars indicate standard deviations. Treatments with the same letters are not significantly different by the least significant difference (LSD) test at P = 0.05
The results indicated that, NO3− influx rate in cucumber roots were also adversely affected by suboptimal RZT, as shown in Fig. 4. The average influx rates indicate that exogenous EBR application significantly increased NO3− influx rate by 72.11 and 86.02%, when compared with LT and BZR treatments, respectively (Fig. 4). NO3− flux rate decreased significantly (by 54.85%) in the LT compared to the NT treatment. Additionally, EBR increased the nitrate influx rate by 15.75% compared to that of the NT treatment, but this difference was not significant (Fig. 4). The influx rate of nitrate did not differ significantly between LT and BZR (Fig. 4).
Similarly, the NH4+ efflux rate was significantly higher in the EBR treatment, but significantly lower in the BZR and LT treatments (Fig. 5). Exogenous EBR application increased average NH4+ efflux rate by 15.75, 71.01, and 76.44%, as compared to the NT, LT, and BZR treatments, respectively (Fig. 5). The NT treatment showed a 71.53% increase in average NH4+ flux rate compared to the LT treatment (Fig. 5). The difference between the NT and EBR treatments was not significant from 0 to 2.5 min (Fig. 5) but became significant over time (Fig. 5). The differences between the LT and BZR treatments were not significant (Fig. 5). These results suggest that suboptimal RZT caused negative effects on cucumber roots and led to a reduction in the flux rate of NO3− and NH4+ which decreased cucumber seedling growth rate. Additionally, exogenous EBR application reduce the detrimental effects of suboptimal RZT through increasing NH4+ and NO3− flux rates.
The effect of EBR on N accumulation under suboptimal RZT
Nitrogen (N) is a major component of proteins, amino acids, nucleic acids, chlorophyll, and enzymes, and thus its accumulation is important for plant growth and development. The results of present study indicated that the LT treatment caused a severe reduction in N accumulation in cucumber root, leaf, and shoot tissues when exposed to suboptimal RZT for seven days, with decreases of 19.30, 12.41 and 48.56%, respectively, when compared with those tissues under the NT treatment (Fig. 6). Further, exogenous EBR application significantly increased the root and shoot total N contents, and plant N accumulation, by 7.41, 29.27, and 50.68%, respectively, when compared with those values for the LT treatment. The maximum root/shoot N accumulation was reported in LT and BZR treated seedlings over that observed in the NT and EBR treatments (Fig. 6).

Nitrogen accumulation in cucumber (root and shoot) seven days after exposure of suboptimal RZT Treatments with the same letters are not significantly different by the least significant difference (LSD) test at P = 0.05
Effect of EBR on NO3 − and NH4 + contents
Suboptimal RZT (LT) significantly decreased the accumulation of nitrate and ammonium in leaves of cucumber seedlings (Fig. 7). Moreover, exogenous EBR application significantly increased nitrate and ammonium accumulation upon exposure to suboptimal RZT by 40.61 and 30.28%, respectively, compared to the LT treatment, and by 43.32 and 34.44%, compared with the BZR treatment. The NT treatment significantly increased nitrate and ammonium contents by 48.77 and 40.66%, respectively, compared to those of the LT treatment. Additionally, nitrate content in the EBR and NT treatments were not significantly different (Fig. 7 A), but ammonium contents were significantly higher (14.89%) in NT when compared to EBR (Fig. 7 B).

EBR regulates NO3−-N and NH4+-N contents in cucumber leaves under suboptimal RZT. Treatments with the same lowercase letter are not significantly different by the least significant difference (LSD) test at P = 0.05
Nitrate transporter (NRT1) gene expression
The effects of suboptimal RZT on nitrate transporter 1 (NRT1) genes (low affinity transporters) were investigated in cucumber leaves under suboptimal RZT (Fig. 8). LT significantly downregulated the transcription levels of NRT1 genes, but these genes were significantly upregulated by exogenous EBR application under suboptimal RZT.

EBR regulates transcript levels of NRT1 genes in leaves of cucumber seedlings under suboptimal RZT. Leaf samples were harvested seven days after treatment. Columns indicate the means of independent measurements of four replications per treatment and bars indicate the SD of the mean. Treatments with the same lowercase letter(s) are not significantly different by the least significant difference (LSD) test at P = 0.05
Transcription levels of CsNRT1.1, CsNRT1.2A, CsNRT1.2B, CsNRT1.2C, CsNRT1.3, CsNRT1.4A, CsNRT1.5B, CsNRT1.5C, CsNRT1.9, and CsNRT1.10 significantly increased in EBR treated seedlings under suboptimal RZT. Additionally, EBR downregulated the expression of CsNRT1.5A and CsNRT1.8, while CsNRT1.4B showed the same trend in the NT, EBR, and BZR treatments, but was downregulated by the LT treatment. Among the LATS gene family, CsNRT1.1, CsNRT1.2A, CsNRT1.2B, CsNRT1.2C, and CsNRT1.5B showed higher expression than did other members of this family when treated with exogenous EBR under suboptimal RZT. These findings indicated that EBR activated the expression of NRT1 genes and led to increased N metabolism, thus improving cucumber seedling growth under suboptimal RZT.
Discussion
EBR is a growth-promoting steroid hormone which plays an active role in a wide range of developmental processes, including abiotic stress tolerance [25, 34, 35]. EBR increases tolerance to abiotic stresses including chilling [29], heat [44], drought [10], and salinity [45]. The previous study shows that EBR regulate thousands of genes in pepper to reduce the harmful effects of chilling stress [46]. Nitrogen is an essential, that’s promotes plant growth and development, as well as alleviate the inhibitory effects of abiotic stresses, but their metabolism is sensitive suboptimal RZT [2]. The previous studies suggested that nitrogen metabolism plays a fundamental role in biosynthesis of chlorophyll and photosynthetic capacity [8]. It has been widely observed that abiotic stress induced reduction in chlorophyll and photosynthetic capacity accompanied by the decrease in the nitrogen metabolic enzyme activities, like NR, NiR, GS, GOGAT [2, 6, 12]. Exogenous EBR application alleviates the harmful effects of low temperature and weak light stress through enhancing the nitrogen metabolism and photochemical efficiency in tomato seedling [28]. Suboptimal RZT causes a significant reduction in plant growth and growth-related parameters [1, 2]. Previous studies reported that exogenous EBR application increased low temperature stress tolerance, regulated levels of endogenous plant hormones (including EBR contents), and regulated expression of an EBR biosynthesis gene (CsDWF) [34, 47, 48]. In the present study, endogenous EBR contents increased in EBR treated seedlings (Fig. 1) and resulted in significant increases in cucumber seedling growth under suboptimal RZT (Table 1). These findings are in line with those of a previous study, who reported that exogenous EBR increased cucumber and pepper seedlings growth under low temperature stress [29, 48].
Suboptimal RZT negatively affects plant growth by affecting chlorophyll, photosynthetic capacity and nitrogen metabolism [2, 6]. Earlier studies reported that suboptimal RZT severely affected plant and root physiology through changes to ion balance [6], nitrogen metabolism [49], chlorophyll [50], photosynthesis [4, 7, 51], and antioxidant enzyme activities and that negatively affected plant metabolic processes [2, 42, 50, 52], thus leading to reduced plant growth (Table 1). Regulation of RZT leads to instantaneous changes in root physiology and affects root metabolism and morphology [4, 6], thereby impacting critical root functions such as nutrient uptake and absorption rate [42]. Additionally, roots are the main source of water and minerals uptake and are responsible for translocation from the soil to plant tissues [49, 53]. A previous study suggested that low root zone temperature caused a significant reduction in root activity, thus leading to a significant reduction in growth [54]. Root activity reflects the strength of all metabolic processes in the root system, including respiration, oxidation, and enzyme activities, and nutrient absorption and translocation from roots to shoots, which are closely related to all developmental processes [55]. Our results demonstrated that, LT treatment severely reduced root activity and bleeding rate, while these significantly increased in cucumber seedlings treated with exogenous EBR (Fig. 2). The correlation analysis suggests the positive correlation between root activity, bleeding rate with root ion flux, as presented in Table 2. These findings suggest that EBR plays an important role in alleviating the harmful effects of suboptimal RZT by regulating root activity and bleeding rate in cucumber seedlings. The results are consistent with those of earlier studies, which found that RZT significantly affected plant biomass, bleeding rate, and root activities [1, 55]. Moreover, suboptimal RZT both increases root oxygen demand and decreases oxygen concentration available in soil, which can result in hypoxic conditions and reduce root and shoot growth [56].
Nitrogen is an important constituent of basic nitrogen-containing compounds such as amino acids, proteins, chlorophylls, and nucleotides, which play important roles in plant growth and development [12, 17, 57, 51]. Therefore, understanding the physiological and molecular mechanisms of nitrogen metabolism and responses to suboptimal RZT is important for agronomic approaches to enhance nitrogen use efficiency in crops and reduce losses [13, 20, 58]. Most plants absorb inorganic nitrogen from soil as ammonium (NH4+) and nitrate (NO3−), which serve as N source [59]. NH4+ can be assimilated to glutamine by the GS and GOGAT enzymes [59, 60]. Plant roots absorb NO3− which is then converted into NH4+ by NR and NiR enzymes for synthesis of amino acids, proteins, and nucleotides [17], thus N accumulation in roots and shoots is important for plant growth and development [58]. A previous study revealed that RZT significantly reduced N accumulation in leaf and root tissues of cucumber, and suggested that N uptake depends on the temperature root zone [42]. Our results indicated that N contents (total N, nitrate, and ammonium contents) under suboptimal RZT were much lower than those of EBR treated seedlings, are suggested that EBR can reduce the harmful effects of suboptimal RZT, as presented in Fig. 6. Under suboptimal RZT, root activity and bleeding rate (Fig. 2) were significantly lower than in the EBR treatments, which indicated that EBR alleviates the harmful effects induced by suboptimal RZT and may explain why EBR increased N accumulation (Fig. 6). These findings are suggested that, suboptimal RZT reduce N accumulation, thus leads to reduce cucumber seedlings growth (Table 1). The results build upon those of previous studies in which RZT negatively affected plant growth through reduced nutrient accumulation [2, 6, 61].
Enzyme activities are very sensitive and reduce very quickly under abiotic stresses [42, 49, 52, 62]. Previous studies indicated that exogenous EBR application positively regulated the activities of enzymes involved in nitrogen metabolism (NR, NiR, GS, and GOGAT) [28, 63]. We investigated the activity of these enzymes involved in N metabolism, as presented in Fig. 3. Our results indicated that the activities of these enzymes (NR, NiR, GS and GOGAT) under suboptimal RZT were much lower than in the EBR treatment. We proposed that suboptimal RZT might have cause a reduction in enzyme activities (Fig. 3), thus leading to a significant reduction in nitrate and ammonium contents (Fig. 7). Our results suggested that the NR, NiR, GS and GOGAT enzymes activities, and assimilation of nitrate and ammonium were promoted after EBR application in cucumber, as exposed to stress. These findings are supported by an earlier study which reported that EBR enhanced the activity and expression levels of GS and GOGAT enzymes and genes in Arabidopsis and concluded that BZR1 and BES1 transcription factors might be involved in different pathways of BR-mediated nitrogen metabolism and uptake [14, 23, 52]. Therefore, exogenous EBR application regulated N metabolism under suboptimal RZT, thus leading to improved growth, as presented in Table 1.
Suboptimal RZT significantly reduced root activity and bleeding rate, both of which may affect nutrient and water uptakes in cucumber seedlings (Fig. 2). N acquisition in plants is primarily regulated by plant hormones [28, 64], which may activate nitrogen signaling pathway to promotes the flux rate of NH4+ and NO3− ion in roots [65]. Previous studies reported that ion flux is sensitive to external stimuli (abiotic stresses), which can cause a significant reduction in ion uptake/flux rate [1, 2]. Our previous results suggested, that suboptimal RZT severely reduced the enzyme activities involved in N metabolism (Fig. 3). Therefore, we speculated that EBR plays a role in NH4+ and NO3− flux rates in cucumber roots under suboptimal RZT. As predicted, NH4+ efflux and NO3− influx rate in cucumber roots under suboptimal RZT were significantly lower than in the EBR treatments (Figs. 4 & 5). Additionally, we compared NH4+ efflux and NO3− influx rates under the EBR treatment with those of the NT and BZR treatments to make clear the role of EBR. Our findings suggested that EBR increased NH4+ and NO3− flux rates and reduced harmful effects, thus leads to significant increment in nitrogen accumulation (Fig. 6). The earlier studies suggested that ammonium and nitrate flux rates are affected by abiotic stresses [8, 46, 66]. EBR is a steroid hormone and induces plant tolerance to a variety of stresses [51, 64, 67, 68]. These findings are suggested that, EBR reduces the negative effect of suboptimal RZT, through increasing NH4+ and NO3− flux rates under suboptimal RZT [28, 69, 70]. Additionally, NRT1s protein family plays an important role in nitrate absorption from soil and translocation to various plant tissues, and these proteins were significantly upregulated by exogenous EBR application, as presented in Fig. 8. Activation of the EBR signal transduction pathway may lead to upregulation of the AMT1, NRT1, and GS/GOGAT genes in Arabidopsis [23, 71, 72]. The positive correlation was reported between ion flux rate, root activity and bleeding rate (Table 2), are suggesting that EBR minimized the detrimental effects induced by suboptimal RZT, and could explain mechanism of NH4+ and NO3− influx rates, that’s significantly higher in EBR treated seedlings. These findings provide evidence that EBR enhanced NH4+ and NO3− acquisition capacity, which may have significantly increased nitrogen metabolism and cucumber seedling growth (Table 1).
Plants absorb nitrate and ammonium from soil through various transporters: NO3− is absorbed by NRT1 protein family members and incorporated into amino acids through the GS and GOGAT enzymes [2, 15, 73]. The NRT1s gene family is responsible for the overall mechanism of nitrate absorption and translocation in plants [12, 74]. However, suboptimal RZT is unfavorable for N acquisition and metabolism, which significantly reduces horticultural production [2, 7, 54]. We investigated the effect of EBR using transcript levels of NRT1 genes, which play specific roles in nitrate absorption and translocation in various plant tissue. Previous studies reported a positive correlation between the flux rates of nitrate and ammonium with the transcript levels of NRT1 genes [13]. The results of this study indicated that exogenous EBR application significantly induced the expression of CsNRT1 genes, which were downregulated by suboptimal RZT and exogenous BZR (Fig. 8). These findings suggest that exogenous EBR activated the expression levels of NRT1 genes, potentially contributing to the observed increase in nitrate (Fig. 4) and ammonium flux rates (Fig. 5), enzyme activities (Fig. 3), and N accumulation in leaf and root tissues of EBR treated seedlings (Fig. 6). The CsNRT1s gene upregulated after exogenous EBR application, are suggested that BZR1 and BES1 transcription factors might be involved directly in NRT1s regulation [14, 15, 23]. A previous study reported that CsNRT1 genes showed variable expression patterns across plant tissues and suggested that NRT1s proteins are primarily responsible for nitrate absorption and translocation. Among these, CsNRT1.1, CsNRT1.3, CsNRT1.4B, CsNRT1.5A and CsNRT1.8 regulate nitrate, whereas other members of this family (CsNRT1.9, CsNRT1.2 s, CsNRT1.4A, CsNRT1.5B, and CsNRT1.5C) also appear to play distinct physiological roles in plants [12, 16]. These variable roles help to explain why NTR1 genes showed different expression patterns, as presented in Fig. 8. The EBR signaling pathway is known to mediate AMT1 encoded genes and induce N-metabolism and uptake in Arabidopsis [23]. In our study, similar temporal levels of NRT1 genes were observed, indicating that suboptimal RZT down-regulated NRT1 genes and significantly reduced growth [2, 7]. In a recent study, EBR affected N metabolism by increasing nitrate and ammonium contents and enzyme activities (NR, NiR, GS, and GOGAT), which increased the tolerance of tomato to low light and temperature [23, 28, 49]. These findings are concluding that, exogenous EBR alleviates the adverse effect of suboptimal RZT by modulating nitrogen metabolism, thus leading to improved cucumber seedling growth.
Conclusion
In summary, suboptimal RZT caused harmful effects on cucumber seedlings and severely reduced plant growth, while exogenous EBR application reduced these detrimental effects of suboptimal RZT. Exogenous EBR application could effectively regulate nitrogen metabolism via increasing activity of enzymes involved in nitrogen metabolism (NR, NiR, GS, and GOGAT), and transcript levels of NRT1 genes, nitrate and ammonium influx rate, which might have a positive effect on growth of cucumber seedlings exposed to suboptimal RZT, as presented in Fig. 9. Our study provides the first evidence of the role of exogenous EBR in transcriptional regulation of NRT1 genes, NO3− and HN4+ ion flux rates under suboptimal RZT in cucumber. This study provides new insights into EBR signal transduction pathway and interactions with NRT1 family genes and responses under suboptimal RZT. Future studies will need to focus on the molecular mechanism of the interaction between EBR and NRT1 genes during signal transduction in plants.

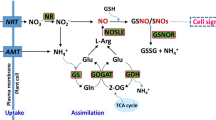
The regulatory mechanism of EBR to alleviate the deleterious effect of suboptimal RZT
Methods
Plant material and growth conditions
Cucumber (Cucumis sativus L. Cv. Zhongnong 26) seeds were obtained from; The Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences, Beijing, China. The detail method and treatments are same as previously described by Anwar et al. [75]
Measurement of growth
Plant height and root length were determined using a ruler, while fresh weight was determined using a digital balance [9]. The seedling index was used to calculate as (stem thickness / plant height + root FW / shoot FW) * FW of whole plant.
Determination of root activity and bleeding rate
The root activity and bleeding rate were investigated six days after exposure to suboptimal RZT. Eight seedlings per treatment were cut below the cotyledons and the incisions were quickly covered with absorbent cotton to collect bleeding sap for two hours. The bleeding rate was calculated from the weight increments of absorbent cotton after two hours. The root activity was determined using TTC (C19H15CIN4) [76].
Enzyme assays
Enzyme activities of nitrate reductase (NR), nitrite reductase (NiR), glutamine synthetase (GS), and glutamate synthase (GOGAT) were determined using assay kits (COMINBIO) with a UV-1800 spectrophotometer following the manufacturer’s instructions [77].
Determination of nitrate and ammonium contents
Nitrate (NO3−) and ammonium (NH4+) contents in the leaf were determined after seven days of exposure to suboptimal RZT, using the methods described by [28]. The OD was read at 410 nm and 625 nm, and contents were calculated using a standard curve [28].
Measurement of NO3 − and NH4 + flux rate at the root surface with NMT
Net NH4+ flux of cucumber seedlings roots was measured by using Non-invasive Micro-test Technology (NMT Physiolyzer®, Younger USA LLC, Amherst, MA 01002, USA) in Xuyue (Beijing) Sci. & Tech. Co., Ltd., Beijing, China [78].
The cucumber roots were fixed to the bottom of petri dish using resin blocks and filter paper strips, the root tip was exposed, then incubated in the testing solution (2.625 mM Ca (NO3)2, 0.1 mM MgSO4, 0.25 mM NH4NO3, 0.3 mM MES, pH 6.0) for 20 min. After that, roots were transferred to a petri dish containing 5 ml of fresh testing solution. Then placed the root sample on the detection platform, and the NH4+ flux microsensor (NH4+ liquid ion exchanger: XY-SJ-NH4; NH4+ flux microsensor: XY-CGQ-01; Xuyue (Beijing) Sci. &Tech. Co., Ltd., Beijing, China.) was positioned close to the root tip (root hair zone) of cucumber seeding. The tip of NH4+ flux microsensor was about 5 μm form the root surface without touched the root. 10 min for each sample and 6 replicates per group. Use imFluxes software (imfluxes.com, Xuyue (Beijing) Sci. & Tech. Co., Ltd., Beijing, China) to obtained NH4+ flux data and process them. NO3− flux detection steps are exactly the same as NH4+ [78].
Estimation of total nitrogen contents
The total nitrogen (N) contents in root and shoots were estimated using the method described by Anwar et al. [75, 79].
Endogenous EBR contents determination
EBR contents were determined using an enzyme-linked immunosorbent assay technology (ELISA) at the College of Agronomy and Biotechnology, China Agricultural University, Beijing, China [48].
Quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNA was isolated using RNAprep Pure Kit (TANGEN) and Fast Quant RT Kit (TANGEN) was used to synthesized first strand cDNA, as described by Anwar et al. [75]. Additionally, primers were designed by using Primer Premier 5 software (Additional file 1: Table S1).
Statistical analysis
Statistix 8.1 software (www.statistix.com) was used to analyze the difference between treatments. The figures were drown by using Graphpad Prism 5 (www.graphpad.com), as described by Anwar et al. [75].
Availability of data and materials
The supporting data are available within the article and additional files.
Change history
28 October 2019
In the original publication of this article [1], the author pointed out there is an error in Figs. 4 and 5.
Abbreviations
- AMT1:
-
Ammonium Transporter 1
- ANOVA:
-
Analysis of Variance
- BES1:
-
BRI1-EMS SUPPERSSOR 1
- BZR:
-
Brassinazole
- BZR1:
-
BRASSINAZOLE RESISTANT 1
- COR:
-
Cold Responsive Genes
- EBR:
-
24-Epibrassinolide
- GOGAT:
-
Glutamate Synthase
- GS:
-
Glutamine Synthetase
- HSP:
-
Heat Shock Proteins
- LSD:
-
Least Significant Difference
- N:
-
Nitrogen
- NH4 + :
-
Ammonium
- NiR:
-
Nitrite Reductase
- NMT:
-
Non-invasive Micro-test Technology
- NO3 − :
-
Nitrate
- NR:
-
Nitrate Reductase
- NRT1:
-
Nitrate Transporter 1
- RZT:
-
Root Zone Temperature
References
Wang X, Zhang W, Miao Y, Gao L. Root-zone warming differently benefits mature and newly unfolded leaves of Cucumis sativus L. seedlings under sub-optimal temperature stress. PLoS One. 2016;11(5):e0155298.
Bai L, Deng H, Zhang X, Yu X, Li Y. Gibberellin is involved in inhibition of cucumber growth and nitrogen uptake at suboptimal root-zone temperatures. PLoS One. 2016;11(5):e0156188.
Li L, Lu X, Ma H, Lyu D. Jasmonic acid regulates the ascorbate–glutathione cycle in Malus baccata Borkh. Roots under low root-zone temperature. Acta Physiolo Plant. 2017;39(8):174.
Hao HP, Jiang CD, Zhang SR, Tang YD, Shi L. Enhanced thermal-tolerance of photosystem II by elevating root zone temperature in Prunus mira Koehne seedlings. Plant Soil. 2012;353(1):367–78.
Li L, Lu X, Ma H, Lyu D. Comparative proteomic analysis reveals the roots response to low root-zone temperature in Malus baccata. J Plant Res. 2018;131:865.
He Y, Yang J, Zhu B. Zhu Z-j. low root zone temperature exacerbates the ion imbalance and photosynthesis inhibition and induces antioxidant responses in tomato plants under salinity. J Integr Agr. 2014;13(1):89–99.
Ding X, Jiang Y, He L, Zhou Q, Yu J, Hui D, Huang D. Exogenous glutathione improves high root-zone temperature tolerance by modulating photosynthesis, antioxidant and osmolytes systems in cucumber seedlings. Sci Rep. 2016;6:35424.
Yusuf M, Fariduddin Q, Ahmad I, Ahmad A. Brassinosteroid-mediated evaluation of antioxidant system and nitrogen metabolism in two contrasting cultivars of Vigna radiata under different levels of nickel. Physiol Mol Biol Plants. 2014;20(4):449–60.
Sakamoto M, Suzuki T. Effect of root-zone temperature on growth and quality of hydroponically grown red leaf lettuce (Lactuca sativa L. cv. Red wave). Am J Plant Sci. 2015;6(14):2350–60.
Gruszka D, Janeczko A, Dziurka M, Pociecha E, Oklestkova J, Szarejko I. Barley Brassinosteroid mutants provide an insight into Phytohormonal homeostasis in plant reaction to drought stress. Front Plant Sci. 2016;7:1824.
Li Y, Li J, Yan Y, Liu W, Zhang W, Gao L, Tian Y. Knock-down of CsNRT2.1, a cucumber nitrate transporter, reduces nitrate uptake, root length, and lateral root number at low external nitrate concentration. Front Plant Sci. 2018;9:722.
Yi-Fang C, Yi W, Wei-Hua W. Membrane transporters for nitrogen, phosphate and potassium uptake in plants. J Integr Plant Biol. 2008;50(7):835–48.
Glass ADM, Britto DT, Kaiser BN, Kinghorn JR, Kronzucker HJ, Kumar A, Okamoto M, Rawat S, Siddiqi MY, Unkles SE, et al. The regulation of nitrate and ammonium transport systems in plants. J Exp Bot. 2002;53(370):855–64.
Chen C-Z, Lv X-F, Li J-Y, Yi H-Y, Gong J-M. Arabidopsis NRT1.5 is another essential component in the regulation of nitrate reallocation and stress tolerance. Plant Physiol. 2012;159(4):1582–90.
Hsu P-K, Tsay Y-F. Two phloem nitrate transporters, NRT1.11 and NRT1.12, are important for redistributing xylem-borne nitrate to enhance plant growth. Plant Physiol. 2013;163(2):844–56.
Fan X, Feng H, Tan Y, Xu Y, Miao Q, Xu G. A putative 6-transmembrane nitrate transporter OsNRT1.1b plays a key role in rice under low nitrogen. J Integr Plant Biol. 2016;58(6):590–9.
De Jong F, Thodey K, Lejay LV, Bevan MW. Glucose elevates NITRATE TRANSPORTER 2.1 protein levels and nitrate transport activity independently of its HEXOKINASE1-mediated stimulation of NITRATE TRANSPORTER2.1 expression. Plant Physiol. 2014;164(1):308–20.
Lu J, Zhang L, Lewis RS, Bovet L, Goepfert S, Jack AM, Crutchfield JD, Ji H, Dewey RE. Expression of a constitutively active nitrate reductase variant in tobacco reduces tobacco-specific nitrosamine accumulation in cured leaves and cigarette smoke. Plant Biotech J. 2016;14(7):1500–10.
Huang L, Li M, Shao Y, Sun T, Li C, Ma F. Ammonium uptake increases in response to PEG-induced drought stress in Malus hupehensis Rehd. Environ Exp Bot. 2018;151:32–42 https://doi.org/10.1016/j.envexpbot.2018.04.007.
Orsel M, Chopin F, Leleu O, Smith SJ, Krapp A, Daniel-Vedele F, Miller AJ. Characterization of a two-component high-affinity nitrate uptake system in Arabidopsis. Physiology and protein-protein interaction. Plant Physiol. 2006;142(3):1304–17.
Møller ALB, Pedas P, Andersen B, Svensson B, Schjoerring JK, Finnie C. Responses of barley root and shoot proteomes to long-term nitrogen deficiency, short-term nitrogen starvation and ammonium. Plant Cell Environ. 2011;34(12):2024–37.
Anwar A, Liu Y, Dong R, Bai L, Yu X, Li Y. The physiological and molecular mechanism of brassinosteroid in response to stress: a review. Biol Res. 2018;51(1):46.
Zhao BT, Zhu XF, Jung JH, Xuan YH. Effect of brassinosteroids on ammonium uptake via regulation of ammonium transporter and N-metabolism genes in Arabidopsis. Biol Plant. 2016;60(3):563–71.
Sharma P, Kumar A, Bhardwaj R. Plant steroidal hormone epibrassinolide regulate–heavy metal stress tolerance in Oryza sativa L. by modulating antioxidant defense expression. Environ Exp Bot. 2016;122:1–9.
Choudhary SP, Yu JQ, Yamaguchi-Shinozaki K, Shinozaki K, Tran LS. Benefits of brassinosteroid crosstalk. Trends Plant Sci. 2012;17(10):594–05.
Wu L, Zhang Z, Zhang H, Wang XC, Huang R. Transcriptional modulation of ethylene response factor protein JERF3 in the oxidative stress response enhances tolerance of tobacco seedlings to salt, drought, and freezing. Plant Physiol. 2008;148(4):1953–63.
Sun S, An M, Han L, Yin S. Foliar application of 24-Epibrassinolide improved salt stress tolerance of perennial ryegrass. Horti sci. 2015;50(10):1518–23.
Shu S, Tang Y, Yuan Y, Sun J, Zhong M, Guo S. The role of 24-epibrassinolide in the regulation of photosynthetic characteristics and nitrogen metabolism of tomato seedlings under a combined low temperature and weak light stress. Plant Physiol Bioch. 2016;107:344.
Li J, Yang P, Kang J, Gan Y, Yu J, Calderon-Urrea A, Lyu J, Zhang G, Feng Z, Xie J. Transcriptome analysis of pepper (Capsicum annuum) revealed a role of 24-Epibrassinolide in response to chilling. Front Plant Sci. 2016;7:1281.
Wei LJ, Deng XG, Zhu T, Zheng T, Li PX, Wu JQ, Zhang DW, Lin HH. Ethylene is involved in Brassinosteroids induced alternative respiratory pathway in cucumber (Cucumis sativus L.) seedlings response to abiotic stress. Front Plant Sci. 2015;6:982.
Xia X-J, Huang L-F, Zhou Y-H, Mao W-H, Shi K, Wu J-X, Asami T, Chen Z, Yu J-Q. Brassinosteroids promote photosynthesis and growth by enhancing activation of rubisco and expression of photosynthetic genes in Cucumis sativus L. Planta. 2009;230(6):1185.
Bajguz A. Brassinosteroid enhanced the level of abscisic acid in Chlorella vulgaris subjected to short-term heat stress. J Plant Physiol. 2009;166(8):882–6.
Saini S, Sharma I, Pati PK. Versatile roles of brassinosteroid in plants in the context of its homoeostasis, signaling and crosstalks. Front Plant Sci. 2015;6:950.
Xia XJ, Wang YJ, Zhou YH, Yuan T, Mao WH, Kai S, Asami T, Chen ZX, Yu JQ. Reactive oxygen species are involved in brassinosteroid-induced stress tolerance in cucumber. Plant Physiol. 2009;150(2):801–14.
Wu W, Zhang Q, Ervin EH, Yang Z, Zhang X. Physiological mechanism of enhancing salt stress tolerance of perennial ryegrass by 24-Epibrassinolide. Front Plant Sci. 2017;8:1017.
Fariduddin Q, Khalil RR, Mir BA, Yusuf M, Ahmad A. 24-Epibrassinolide regulates photosynthesis, antioxidant enzyme activities and proline content of Cucumis sativus under salt and/or copper stress. Environ Monit Assess. 2013;185(9):7845–56.
Fariduddin Q, Yusuf M, Chalkoo S, Hayat S, Ahmad A. 28-homobrassinolide improves growth and photosynthesis in Cucumis sativus L. through an enhanced antioxidant system in the presence of chilling stress. Photosynthetica. 2011;49(1):55–64.
Talaat NB, Shawky BT. 24-Epibrassinolide ameliorates the saline stress and improves the productivity of wheat (Triticum aestivum L.). Environ Exp Bot. 2012;82:80–8.
Choe S, Dilkes BP, Fujioka S, Takatsuto S, Sakurai A, Feldmann KA. The DWF4 gene of Arabidopsis encodes a cytochrome P450 that mediates multiple 22alpha-hydroxylation steps in brassinosteroid biosynthesis. Plant Cell. 1998;10(2):231–43.
Clouse SD, Sasse JM. BRASSINOSTEROIDS: essential regulators of plant growth and development. Annu Rev Plant Physiol Plant Mol Biol. 1998;49(1):427–51.
Choe S, Schmitz RJ, Fujioka S, Takatsuto S, Lee M-O, Yoshida S, Feldmann KA, Tax FE. Arabidopsis Brassinosteroid-Insensitive dwarf12 mutants are Semidominant and defective in a glycogen synthase kinase 3β-like kinase. Plant Physiol. 2002;130(3):1506–15.
Yan Q, Duan Z, Mao J, Li X, Dong F. Effects of root-zone temperature and N, P, and K supplies on nutrient uptake of cucumber (Cucumis sativus L.) seedlings in hydroponics. Soil Sci Plant Nutr. 2012;58(6):707–17.
Nada K, He LX, Tachibana S. Impaired photosynthesis in cucumber (Cucumis sativus L.) by high root-zone temperature involves ABA-induced stomatal closure and reduction in Ribulose-1,5-bisphosphate carboxylase/oxygenase activity. Journal of the Japanese Society for Horticultural Science. 2008;72(6):504–10.
Ogweno JO, Song XS, Shi K, Hu WH, Mao WH, Zhou YH, Yu JQ, Nogués S. Brassinosteroids alleviate heat-induced inhibition of photosynthesis by increasing carboxylation efficiency and enhancing antioxidant systems in Lycopersicon esculentum. J Plant Growth Regul. 2008;27(1):49–57.
Hayat S, Hasan SA, Yusuf M, Hayat Q, Ahmad A. Effect of 28-homobrassinolide on photosynthesis, fluorescence and antioxidant system in the presence or absence of salinity and temperature in Vigna radiata. Environ Exp Bot. 2010;69(2):105–12.
Li J, Yang P, Kang J, Gan Y, Yu J, Calderón-Urrea A, Lyu J, Zhang G, Feng Z, Xie J. Transcriptome analysis of pepper (Capsicum annuum) revealed a role of 24-Epibrassinolide in response to chilling. Front Plant Sci. 2016;7:1281.
Dhaubhadel S, Browning KS, Gallie DR, Krishna P. Brassinosteroid functions to protect the translational machinery and heat-shock protein synthesis following thermal stress. Plant J. 2002;29(6):681–91.
Anwar A, Bai L, Miao L, Liu Y, Li S, Yu X, Li Y. 24-Epibrassinolide ameliorates endogenous hormone levels to enhance low-temperature stress tolerance in cucumber seedlings. Int J Mol Sci. 2018;19(9):2497.
He J, Qin L, Lee SK. Root-zone CO2 and root-zone temperature effects on photosynthesis and nitrogen metabolism of aeroponically grown lettuce (Lactuca sativa L.) in the tropics. Photosynthetica. 2013;51(3):330–40.
Adebooye OC, Schmitz-Eiberger M, Lankes C, Noga GJ. Inhibitory effects of sub-optimal root zone temperature on leaf bioactive components, photosystem II (PS II) and minerals uptake in Trichosanthes cucumerina L. Cucurbitaceae. Acta Physiol Plant. 2009;32(1):67.
Tanveer M, Shahzad B, Sharma A, Khan EA. 24-Epibrassinolide application in plants: An implication for improving drought stress tolerance in plants. Plant Physiol Bioch. 2019;135:295–303.
Tanveer M, Shahzad B, Sharma A, Biju S, Bhardwaj R. 24-Epibrassinolide; an active brassinolide and its role in salt stress tolerance in plants. A review. Plant Physiol Bioch. 2018;130:69–79.
Comas L, Becker S, Cruz VMV, Byrne PF, Dierig DA. Root traits contributing to plant productivity under drought. Front Plant Sci. 2013;4:442.
Q-y Y, Z-q D, J-d M, Li X, Dong F. Low root zone temperature limits nutrient effects on cucumber seedling growth and induces adversity physiological response. J Integr Agr. 2013;12(8):1450–60.
Fang S, Tao Y, Zhang Y, Kong F, Wang Y. Effects of metalaxyl enantiomers stress on root activity and leaf antioxidant enzyme activities in tobacco seedlings. Chirality. 2018;30(4):469–74.
Gonzalez-Fuentes JA, Shackel K, Lieth JH, Albornoz F, Benavides-Mendoza A, Evans RY. Diurnal root zone temperature variations affect strawberry water relations, growth, and fruit quality. Sci Hortic. 2016;203:169–77.
Xiaorong F, Huimin F, Yawen T, Yanling X, Qisong M, Guohua X. A putative 6-transmembrane nitrate transporter OsNRT1.1b plays a key role in rice under low nitrogen. J Integr Plant Biol. 2016;58(6):590–9.
Wu M, Liu M, Liu J. Li W-t, Jiang C-y, Li Z-p. optimize nitrogen fertilization location in root-growing zone to increase grain yield and nitrogen use efficiency of transplanted rice in subtropical China. J Integr Agr. 2017;16(9):2073–81.
O'Brien J, Vega A, Bouguyon E, Krouk G, Gojon A, Coruzzi G, Gutiérrez R. Nitrate transport, sensing, and responses in plants. Mol Plant. 2016;9(6):837–56.
Santiago JP, Tegeder M. Implications of nitrogen phloem loading for carbon metabolism and transport during Arabidopsis development. J Integr Plant Biol. 2017;59(6):409–21.
He J, Austin PT, Lee SK. Effects of elevated root zone CO2 and air temperature on photosynthetic gas exchange, nitrate uptake, and total reduced nitrogen content in aeroponically grown lettuce plants. J Exp Bot. 2010;61(14):3959.
Gill SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Bioch. 2010;48(12):909–30.
Chaiwanon J, Garcia Veder J, Cartwright H, Sun Y, Wang Z-Y. Immunophilin-like FKBP42/TWISTED DWARF1 interacts with the receptor kinase BRI1 to regulate Brassinosteroid signaling in Arabidopsis. Mol Plant. 2016;9(4):593–600.
Belkhadir Y, Wang X, Chory J. Brassinosteroid signaling pathway. Sci STKE. 2006.
Bartoli CG, Casalongué CA, Simontacchi M, Marquez-Garcia B, Foyer CH. Interactions between hormone and redox signalling pathways in the control of growth and cross tolerance to stress. Environ Exp Bot. 2013;94:73–88.
Huang L, Li M, Zhou K, Sun T, Hu L, Li C, Ma F. Uptake and metabolism of ammonium and nitrate in response to drought stress in Malus prunifolia. Plant Physiol Bioch. 2018;127:185–93.
Divi UK, Krishna P. Brassinosteroids confer stress tolerance. Plant stress biology; genomics goes systems biology. Edited by Hirt H. Weinheim: Wiley-VCH; 2009.
Divi UK, Krishna P. Overexpression of the Brassinosteroid biosynthetic gene AtDWF4 in Arabidopsis seeds overcomes abscisic acid-induced inhibition of germination and increases cold tolerance in transgenic seedlings. J Plant Growth Regul. 2010;29(4):385–93.
Dalio RJD, Pinheiro HP, Sodek L, Haddad CRB. 24-epibrassinolide restores nitrogen metabolism of pigeon pea under saline stress. Bot Stud. 2013;54(1):9.
Druege U, Franken P, Hajirezaei MR. Plant hormone homeostasis, signaling, and function during adventitious root formation in cuttings. Front Plant Sci. 2016;7(381).
Migocka M, Warzybok A, Kłobus G. The genomic organization and transcriptional pattern of genes encoding nitrate transporters 1 (NRT1) in cucumber. Plant Soil. 2013;364(1):245–60.
Gupta P, Srivastava S, Seth CS. 24-Epibrassinolide and sodium nitroprusside alleviate the salinity stress in Brassica juncea L. cv. Varuna through cross talk among proline, nitrogen metabolism and abscisic acid. Plant Soil. 2017;411(1):483–98.
Meng S, Su L, Li Y, Wang Y, Zhang C, Zhao Z. Nitrate and ammonium contribute to the distinct nitrogen metabolism of Populus simonii during moderate salt stress. PLoS One. 2016;11(3):e0150354.
Bai H, Euring D, Volmer K, Janz D, Polle A. The nitrate transporter (NRT) gene family in poplar. PLoS One. 2013;8(8):e72126.
Anwar A, Di Q, Yan Y, He C, Li Y, Yu X. Exogenous 24-epibrassinolide alleviates the detrimental effects of suboptimal root zone temperature in cucumber seedlings. Arch Agron Soil Sci. 2019:1–14.
Bai BZ, Jin JZ, Bai S, Huang LP. Improvement of TTC method determining root activity in corn. Maize Sci. 1994;2:44–7 in Chinese with english abstract.
Bai L, Liu Y, Mu Y, Anwar A, He C, Yan Y, Li Y, Yu X. Heterotrimeric G-protein γ subunit CsGG3.2 positively regulates the expression of CBF genes and chilling tolerance in cucumber. Front Plant Sci. 2018;9(488).
Yu Y, Xu T, Li X, Tang J, Ma D, Li Z, Sun J. NaCl-induced changes of ion homeostasis and nitrogen metabolism in two sweet potato (Ipomoea batatas L.) cultivars exhibit different salt tolerance at adventitious root stage. Environ Exp Bot. 2016;129:23–36.
Jiahui Z, Ning Z, Congcong L, Hao Y, Meiling L, Guirui Y, Kevin W, Qiang Y, Nianpeng H. C:N:P stoichiometry in China's forests: from organs to ecosystems. Funct Ecol. 2018;32(1):50–60.
Acknowledgments
Not applicable.
Funding
This work was supported by the National Key Research and Development Program of China (2016YFD0201006) to Yansu Li, the Earmarked fund for Modern Agro-industry Technology Research System (CARS-25-C-01) to Xianchang Yu, the Science and Technology Innovation Program of the Chinese Academy of Agricultural Sciences (CAAS-ASTIP-IVFCAAS) to Xianchang Yu, and the Key Laboratory of Horticultural Crop Biology and Germplasm Innovation, Ministry of Agriculture, China. The funders had no role in the study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Author information
Authors and Affiliations
Contributions
AA and XY, LY conceived and designed the study. AA perform the whole experiment and wrote the manuscript. YL and CH review the manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors have declared that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Additional files
Additional file 1:
Table S1. The list of specific primers for Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR). (XLSX 10 kb)
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Anwar, A., Li, Y., He, C. et al. 24-Epibrassinolide promotes NO3− and NH4+ ion flux rate and NRT1 gene expression in cucumber under suboptimal root zone temperature. BMC Plant Biol 19, 225 (2019). https://doi.org/10.1186/s12870-019-1838-3
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12870-019-1838-3