
Abstract
Background
A significant trend towards increased fusidic acid (FA) resistance among Staphylococcus aureus with increased duration of use is of concern. The aim of the present study is to investigate the dissemination of fusidic acid resistance among Staphylococcus aureus clinical isolates.
Methods
The susceptibility of S. aureus isolates to antimicrobial agents was determined by disc-diffusion method. The minimal inhibitory concertrations(MICs) of fusidic acid and vacomycin for fusidic acid-resisitant isolates were determined by ager dillution method. FA resistance determinants were determined by PCR and DNA sequencing. SCCmec typing, spa typing and multi-locus sequence typing were used for the determination of molecular characteristics for S. aureus isolates.
Results
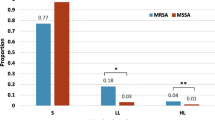
A total of 392 non-duplicate S. aureus isolates including 181 methicillin-resistant S. aureus (MRSA) isolates, which were isolated from the clinical specimens of patients at a Chinese tertiary hospital from January, 2012 to September, 2013, were collected for investigating FA resistance. Among 392 S. aureus clinical isolates tested, 56 (14.3 %) with FA MIC values ranging from 2 μg/ml to ≥128 μg/ml were resistant to FA. The proportions of FA resistance among MRSA and MSSA isolates were 27.1 % (49/181) and 3.3 % (7/211). There was a trend of rapidly increased FA resistance among S. aureus and MRSA isolates from 5.2 % and 8.9 % in 2012 to 24.9 % and 45.1 % in 2013. Acquired FA resistance gene, fusB, was present in 73.2 % (41/56) of FA-resistant S. aureus isolates. fusC and fusA mutation were not found in any of tested isolates. A total of 9 sequence types (STs) and 12 spa types were identified among the 56 FA-resistant S. aureus isolates. ST5 accounting for 66.1 % (37/56) was the most prevalent ST. The majority (92.9 %, 52/56) of the isolates tested belonged to clonal complex 5(CC5). t2460 was the most prevalent spa type, accounting for 67.9 % (38/56) . ST5-MRSA- II-t2460 was predominant clone, accounting for 75.5 % (37/49) of FA-resistant MRSA isolates and 66.1 % (37/56) of FA-resistant S. aureus isolates. Five of 7 FA-resistant MSSA isolates belonged to ST630-MSSA.
Conclusion
Increased FA resistance among S. aureus isolates was found in China. fusB was predominant FA resistance determinant. The spread of CC5 clone, especially novel ST5-MRSA- II-t2460 clone with high-level resistance to FA, was responsible for the increase of FA resistance.
Similar content being viewed by others


Background
Staphylococcus aureus, especially methicillin-resistant S. aureus (MRSA), is an important and often encountered human pathogen responsible for many infectious diseases from mild superficial skin infection to life-threatening diseases, including skin and soft tissue infections (SSTIs), foreign-body infections, pneumonia, septic arthritis, endovascular infections, osteomyelitis, sepsis, and bloodstream infections (BSI) in both hospitals and community settings [1]. Since the first European isolate of MRSA was detected in 1960s, MRSA with resistance to all available penicillins and other β-lactam antimicrobial agents has become a leading cause of nosocomial infections in hospitals and other healthcare facilities. In addition to β-lactam, MRSA isolates are often resistant to erythromycin, clindamycin, aminoglycosides and quinolones. Since the mid-1990s, however, MRSA isolates are found to be responsible for severe infections in individuals lacking risk factors for exposure to the health care system [1, 2]. MRSA infections has become a global concern [1, 3]. As vancomycin has relatively clean safety profile and durability against the development of resistance, it has been considered to be the optimal option for the treatment of invasive MRSA infections [4]. However, the emergence of vancomycin- hetero-resistant S. aureus (hVRSA), vancomycin-intermediate S. aureus (VISA) and vancomycin-resistant S. aureus (VRSA) limits the use of vancomycin for the treatment of serious MRSA infections [5–7]. In addition, the drawbacks of vancomycin, including poor tissue penetration, serious nephrotoxicity and neutropenia and unpredictable synergy with other antimicrobial agents, also limit the selection of the alternatives for the treatment of MRSA infections. Fusidic acid (FA), is an effective agent for the treatment of SSTIs, acute osteomyelitis, chronic osteomyelitis, vertebral infection, septic arthritis, and prosthetic and other device related infections caused by MRSA and methicillin-susceptible S. aureus (MSSA) [8–10]. FA dose of 250-500 mg every 8–12 h was recommended for the treatment of MRSA infections [8]. However, recent study reported that front-loaded dosing of ≥1200 mg every 12 h (q12h) has better activity against MRSA infections than non-front-loaded dosing (600 mg q12 h) [11]. Although FA has not been approved for use in the USA by the US FDA, this drug has been widely used in Europe, Canada, Australia and some Asian countries for decades [8]. FA is a valuable alternative to vancomycin for the treatment of MRSA infections. In the 1990s, the prevalence of resistance to FA in S. aureus remained low. But there is a concern that there is a significant trend towards increased FA resistance among S. aureus with increased duration of use. In a surveillance report on FA resistance in 13 European countries, the prevalence of resistance to FA among S. aureus isolates was 10.7 %, with the highest resistance rates of 62.4 % among the isolates from Greece [12]. Although many Asian countries have reported FA resistance among S. aureus isolates, these data in China are limited. The aim of the present study is to investigate the dissemination of fusidic acid resistance among S. aureus clinical isolates from a tertiary hospital between 2012 and 2013 in Wenzhou, east China.
Methods
Collection of S. aureus clinical isolates
From Jan. 2012 to Sep. 2013, a total of 392 non-duplicate S. aureus isolates (single isolate per patient) from the various specimens of patients at the first Affiliated Hospital of Wenzhou Medical University locating in Wenzhou, east China, were selected for investigating the prevalence of FA resistance. The numbers of S. aureus isolates included in 2012 and 2013 were 211 and 181. Isolates were identified as S. aureus by using Gram’s stain, positive catalase and coagulase test results, and Vitek microbiology analysis (BioMérieux, Marcy l’Etoile, France). S. aureus ATCC25923 was used as a control strain for the identification of bacteria. Because the present study focused on bacteria, the Ethics Committee of the first Affiliated Hospital of Wenzhou Medical University exempted this investigation from review. Informed consents were required from patients for sample collection.
Antimicrobial susceptibility testing
S. aureus susceptibilities to penicillin (10 μg), erythromycin (15 μg), clindamycin (2 μg), rifampicin (5 μg), tetracycline(30 μg), linezolid (30 μg), quinupristin/dalfopristin (15 μg), trimethoprim/sulfamethoxazole (1.25/23.75 μg), gentamicin (10 μg), ciprofloxacin (5 μg), chloramphenicol (30 μg), fusidic acid (5 μg) and nitrofurantoin (300 μg) were determined using the disk diffusion test recommended by the Clinical and Laboratory Standards Institute (CLSI) [13]. Every assay was replicated for three times. The mean value was applied for the assay. All disks were obtained from Oxoid Ltd. Vancomycin MICs for S. aureus isolates were determined by the agar dilution method in accordance with the CLSI guidelines [14]. Interpretive standards of antimicrobial susceptibility testing for S. aureus isolates tested was in accordance with the guidelines provided by CLSI [13]. Initial screening of FA resistance was performed by disk diffusion method with 5 μg FA containing disks. The isolates with an inhibition zone ≤17 mm in diameter were further determined for FA MIC values by an agar dilution method. The interpretive criterion of FA susceptibility is in accordance with the European Committee for Antimicrobial Susceptibility Testing (EUCAST)/ criteria (susceptible, MIC < 2 μg/ml; resistant, MIC ≥ 2 μg/ml). S. aureus ATCC 25923 and Escherichia coli ATCC25922 were used as reference strains for antimicrobial susceptibility testing.
DNA extraction
S. aureus isolates with FA resistance were cultured on blood agar overnight at 35 °C. Then, three to four bacterial colonies were suspended in 150 μl sterile distilled water with lysostaphin (1 mg/mL) (Sangon, Shanghai, China) and incubated at 37 °C for an hour. Finally, DNA was extracted following the instructions of the Genomic DNA Extraction kit (Sangon, Shanghai, China), stored at −20 °C and prepared for PCR assays.
MRSA identification
A simplex PCR was used for the detection of mecA using MRSA N315 as positive control strain. The isolates positive for mecA were identified as MRSA. And cefoxitin disk diffusion test was also used for the detection of MRSA in accordance with the guideline provided by the Clinical and Laboratory Standards Institute (CLSI) [13].
Detection of FA resistance determinants by PCR
Detection of fusA mutations and acquired FA resistance determinants including fusB and fusC was detected by PCR assays with primers and reaction conditions described previously [15]. DNA sequencing was used for the identification of genotypes of acquired FA resistance determinants. The fusA mutations were determined by sequencing entire fusA gene.
SCCmec typing
SCCmec typing of MRSA isolates was performed using a battery of multiplex PCRs as described previously [16]. The MRSA isolates with unanticipated fragments or lacking fragments by multiplex PCRs were defined as non-typeable (NT). MRSA NCTC 10442 (SCCmecI), MRSA N315 (SCCmec II), MRSA 85/2082 (SCCmec III), MRSA JCSC 4744 (SCCmec IV) and MRSA WZ153 (SCCmec V) were used as control strains for SCCmec typing.
spa typing
The spa variable repeat region from each S. aureus isolate was amplified using simplex PCR oligonucleotide primers as previously described [17, 18]. Following their purification and sequencing, spa types were assigned using the spa database website (http://spaserver.ridom.de).
Multilocus sequence typing (MLST)
MLST typing of S. aureus isolates was performed using amplification of internal fragments of the seven housekeeping genes(arcc, aroe, glpf, gmk, pta, tpi and yqil) of S. aureus as described previously [19]. Following purification and sequencing of these genes, the sequences were compared with the existing sequences available on the MLST website for S. aureus (http://saureus.mlst.net), and STs were assigned according to the allelic profiles.
Results and discussion
Prevalence of FA resistance among S. aureus isolates
Among 392 S. aureus isolates tested including 181 MRSA isolates, 56 (14.3 %) with FA MIC values ranging from 2 to ≥128 μg/ml, including 49 (87.5 %, 49/56) MRSA and 7 (12.5 %, 7/56) MSSA, were resistant to FA. The proportions of FA resistance among MRSA and MSSA isolates were 27.1 % (49/181) and 3.3 % (7/211). The sources for the isolation of the 56 FA-resistant isolates included sputum (42, 75 %), pus (10, 17.9 %), catheter (1, 1.8 %), blood (1, 1.8 %) and exudates (2, 3.6 %). Compared with previous reports from China with FA resistance rates of 3.5 % (4/116) from Shanghai and 2.2 % (4/186) from Chinese children with SSTIs [20, 21], the prevalence of FA resistance among S. aureus clinical isolates in the present study was very high. The resistance rates to FA among S. aureus isolates are different from country to country. The resistance rates of S. aureus isolates to FA were very low in the USA (0.3 %) and relatively high in Canada (7.0 %) and Australia (7.0 %) [22]. In an updated surveillance report on FA resistance in 13 European countries, Israel, Italy, Poland, Spain and Sweden had low rates (1.4–3.1 %), while Greece (62.4 %) and Ireland (19.9 %) had the highest resistance rates [12]. In most Asian countries, FA resistance rates are relatively low (<10 %) [8]. FA resistance was observed more frequently among MSSA isolates (0.6 %) than among MRSA isolates from the United States [22]. On the contrary, the prevalence of FA resistance among MRSA isolates was significantly higher than that among MSSA isolates in the present study. There is a significant trend towards increased FA resistance among S. aureus since FA has been used widely. In Kuwait, the rate of FA resistance increased rapidly from 22 % in 1994 to 92 % in 2004 [23]. There was a trend of rapidly increased FA resistance among S. aureus and MRSA isolates from 5.2 % and 8.9 % in 2012 to 24.9 % and 45.1 % in 2013 in this investigation. The MIC values of FA for 56 FA-resistant isolates were as follows: ≥128 μg/ml, 43; 16 μg/ml, 2; 8 μg/ml, 1; 4 μg/ml, 7; and 2 μg/ml, 3. In 2012, only 5 S. aureus isolates (45.5 %, 5/11) with FA MIC value of ≥128 μg/ml were identified. Surprisingly, the FA MIC values for 38 (84.4 %) of 45 FA-resistant isolates were ≥128 μg/ml in 2013. The resistance rates of FA-resistant S. aureus, MRSA and MSSA isolates to antimicrobials tested were showed in Table 1. All 56 FA-resistant isolates were susceptible to vancomycin, quinupristin-dalfopristin, linezolid, and nitrofurantoin. The antimicrobials with the resistance rates of more than 50 % among FA-resistant S. aureus isolates were penicillin (100 %), erythromycin (92.9 %), clindamycin (89.3 %), ciprofloxacin (82.1 %), and tetracycline (75.0 %), respectively. The resistance rates of the 56 isolates to gentamycin, rifampin, trimethoprim/sulfamethoxazole and chloramphenicol were relatively low. In contrast to the report from Taiwan among which the resistance rate to rimethoprim/sulfamethoxazole among FA-resistant MRSA isolates was high to 97 % [24], only 4.9 % (4/81) of FA-resistant MRSA isolates were resistant to this antimicrobial.
Prevalence of FA resistance determinants
Two major FA resistance mechanisms have been reported in S. aureus, including the alteration of the drug target site, which is due to mutations in fusA encoding elongation factor G (EF-G) or rplF encoding ribosome protein L6), and the protection of the drug target site by FusB family proteins including FusB, FusC, and FusD [8]. In staphylococci, high-level FA resistance is usually associated with mutations in fusA encoding EF-G, while low-level resistance is generally caused by the horizontally transferable genes including fusB, fusC and fusD [25]. The spontaneous mutations in fusA lead to modification of the drug target and reduced susceptibility [26]. The fusB-type resistance involves the recruitment of an exogenous resistance determinant whose product binds EF-G and protects it from FA [27]. In this study, the results from PCR and DNA sequencing demonstrated that acquired resistance gene, fusB was present in 73.2 % (41/56) of FA-resistant isolates tested. However, fusC and fusA mutations were not found in any of tested isolates. The previous investigation from China found that all 4 clinical isolates with FA resistance were positive for fusB and negative for fusC and fusA mutations [20]. These data indicated that fusB is the predominant determinant responsible for FA resistance among S. aureus in China. In Netherlands, fusB was also found to be predominant FA resistance determinant which was detected in about 90 % of FA-resistant S. aureus isolates [28]. On the contrary, a report from northern Taiwan found that 84 % of FA-resistant MRSA isolates had fusA mutations and another report from a Taiwanese hospital found that fusC was the most common FA resistance determinant [15, 24]. S. aureus isolates from Australia with FA MIC values ranging from 2 to 32 μg/ml were predominantly fusC positive [22]. In U.S. and European collections, fusC was also more prevalent than fusB in FA-resistant S. aureus isolates [12, 22]. In contrast to previous report from Taiwan among which the distribution of FA resistance determinants including fusA mutations, fusB and fusC was quite different between MRSA and MSSA groups and no MRSA isolates carried fusB [15].
Molecular characteristics of FA-resistant S. aureus isolates
The molecular characteristics of FA-resistant S. aureus isolates were listed in Table 2. A total of 9 STs were identified among the 56 FA-resistant S. aureus isolates, among which ST5 accounting for 66.1 % (37/56) was the most common ST, followed by ST630 (12.5 %, 7/56). Only 3 MRSA isolates belonged to ST239 which is the predominant ST in China. The STs for two isolates were not identified. The antimicrobial resistance profiles of ST5 and ST239 FA-resistant MRSA isolates were showed in Table 3. The clonal complex 5 (CC5) was the predominant CC, accounting for the majority (92.9 %, 52/56) of the isolates tested. Twelve spa types were identified among the 56 FA-resistant S. aureus isolates, among which t2460 (67.9 %, 38/56) was the most prevalent spa type. The spa types for 5 isolates were not identified. t2460 was identified among 43.9 % of MRSA isolates from 3 intensive care units at a Korean tertiary hospital in 2007–2008 [29]. Among the 49 MRSA isolates, 40, 7 and 2 harbored SCC mec types II, III and V, respectively.
In the present study, ST5-MRSA- II-t2460 was predominant clone, accounting for 75.5 % (37/49) of FA-resistant MRSA isolates and 66.1 % (37/56) of FA-resistant S. aureus isolates. ST5-MRSA- II is usually associated with t002 [30]. ST5-MRSA- II-t002 was one of two major clones in China [30]. However, ST5-MRSA- II was linked to t2460 in this study. This fairly unusual clone was only found among 6 MRSA isolates from Shanghai in China [30]. FA MIC values for 36 of 37 ST5-MRSA- II-t2460 isolates were ≥128 μg/ml. ST5-MRSA- II-t2460 clone first emerged in 2012. However, this newly emerging clone with FA resistance increased dramatically in 2013, accounting for 77.8 % (35/45) of FA-resistant isolates. The origin of 35 isolates belonged to ST5-MRSA- II-t2460 in 2013 was confined to only 4 wards including neurosurgery ward (28 isolates), orthopaedic ward (three isolates), intensive care unit (three isolates) and hematology ward (one isolate), indicating that outbreak of infections caused by this newly clone has occurred in our hospital, mainly in neurosurgery ward. Clonal spread is responsible for the increase in FA resistance in UK, France and other European countries [31, 32]. CC80-MRSA-IV clone carrying fusB has made a major contribution to the prevalence of FA resistance among MRSA isolates in Europe [25]. The highest prevalence of FA resistance in 2013 in this investigation was associated with the spread of ST5-MRSA- II-t2460 clone with high-level FA resistance. Among 7 FA-resistant MSSA isolates, 5 belonged to ST630-MSSA.
Conclusions
In conclusion, increased FA resistance among S. aureus isolates was found in China. fusB was predominant FA resistance determinant. The spread of CC5 clone, especially novel ST5-MRSA- II-t2460 clone with high-level resistance to FA, was responsible for the increase of FA resistance.
References
David MZ, Daum RS. Community-associated methicillin-resistant Staphylococcus aureus: epidemiology and clinical consequences of an emerging epidemic. Clin Microbiol Rev. 2010;23(3):616–87.
Said-Salim B, Mathema B, Kreiswirth BN. Community-acquired methicillin-resistant Staphylococcus aureus: an emerging pathogen. Infect Control Hosp Epidemiol. 2003;24(6):451–5.
Cohen PR. Community-acquired methicillin-resistant Staphylococcus aureus skin infections: implications for patients and practitioners. Am J Clin Dermatol. 2007;8(5):259–70.
Daschner FD, Kropec A. Glycopeptides in the treatment of staphylococcal infections. Eur J Clin Microbiol Infect Dis. 1995;14 Suppl 1:S12–17.
Centers for Disease Control and Prevention (CDC). Vancomycin-resistant Staphylococcus aureus--Pennsylvania, 2002. MMWR Morb Mortal Wkly Rep. 2002;51(40):902.
Hiramatsu K. The emergence of Staphylococcus aureus with reduced susceptibility to vancomycin in Japan. Am J Med. 1998;104(5A):7S–10S.
Hiramatsu K, Aritaka N, Hanaki H, Kawasaki S, Hosoda Y, Hori S, et al. Dissemination in Japanese hospitals of strains of Staphylococcus aureus heterogeneously resistant to vancomycin. Lancet. 1997;350(9092):1670–3.
Wang JL, Tang HJ, Hsieh PH, Chiu FY, Chen YH, Chang MC, et al. Fusidic acid for the treatment of bone and joint infections caused by meticillin-resistant Staphylococcus aureus. Int J Antimicrob Agents. 2012;40(2):103–7.
Schofer H, Simonsen L. Fusidic acid in dermatology: an updated review. Eur J Dermatol. 2010;20(1):6–15.
Whitby M. Fusidic acid in the treatment of methicillin-resistant Staphylococcus aureus. Int J Antimicrob Agents. 1999;12 Suppl 2:S67–71.
Tsuji BT, Okusanya OO, Bulitta JB, Forrest A, Bhavnani SM, Fernandez PB, et al. Application of pharmacokinetic-pharmacodynamic modeling and the justification of a novel fusidic acid dosing regimen: raising Lazarus from the dead. Clin Infect Dis. 2011;52 Suppl 7:S513–519.
Castanheira M, Watters AA, Mendes RE, Farrell DJ, Jones RN. Occurrence and molecular characterization of fusidic acid resistance mechanisms among Staphylococcus spp. from European countries (2008). J Antimicrob Chemother. 2010;65(7):1353–8.
CLSI. Performance standards for antimicrobial susceptibility testing, 18th informational supplement (M100-S22). Wayne, PA, USA: Clinical and Laboratory Standards Institute; 2012.
CLSI. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; approved standard-eighth edition. Wayne, PA, USA: Clinical and Laboratory Standards Institute; 2009.
Chen HJ, Hung WC, Tseng SP, Tsai JC, Hsueh PR, Teng LJ. Fusidic acid resistance determinants in Staphylococcus aureus clinical isolates. Antimicrob Agents Chemother. 2010;54(12):4985–91.
Kondo Y, Ito T, Ma XX, Watanabe S, Kreiswirth BN, Etienne J, et al. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: rapid identification system for mec, ccr, and major differences in junkyard regions. Antimicrob Agents Chemother. 2007;51(1):264–74.
Koreen L, Ramaswamy SV, Graviss EA, Naidich S, Musser JM, Kreiswirth BN. spa typing method for discriminating among Staphylococcus aureus isolates: implications for use of a single marker to detect genetic micro- and macrovariation. J Clin Microbiol. 2004;42(2):792–9.
Harmsen D, Claus H, Witte W, Rothganger J, Turnwald D, Vogel U. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J Clin Microbiol. 2003;41(12):5442–8.
Enright MC, Day NP, Davies CE, Peacock SJ, Spratt BG. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J Clin Microbiol. 2000;38(3):1008–15.
Huang J, Ye M, Ding H, Guo Q, Ding B, Wang M. Prevalence of fusB in Staphylococcus aureus clinical isolates. J Med Microbiol. 2013;62(Pt 8):1199–203.
Liu Y, Geng W, Yang Y, Wang C, Zheng Y, Shang Y, et al. Susceptibility to and resistance determinants of fusidic acid in Staphylococcus aureus isolated from Chinese children with skin and soft tissue infections. FEMS Immunol Med Microbiol. 2012;64(2):212–8.
Castanheira M, Watters AA, Bell JM, Turnidge JD, Jones RN. Fusidic acid resistance rates and prevalence of resistance mechanisms among Staphylococcus spp. isolated in North America and Australia, 2007–2008. Antimicrob Agents Chemother. 2010;54(9):3614–7.
Udo EE, Al-Sweih N, Mokaddas E, Johny M, Dhar R, Gomaa HH, et al. Antibacterial resistance and their genetic location in MRSA isolated in Kuwait hospitals, 1994–2004. BMC Infect Dis. 2006;6:168.
Chen CM, Huang M, Chen HF, Ke SC, Li CR, Wang JH, et al. Fusidic acid resistance among clinical isolates of methicillin-resistant Staphylococcus aureus in a Taiwanese hospital. BMC Microbiol. 2011;11:98.
McLaws FB, Larsen AR, Skov RL, Chopra I, O'Neill AJ. Distribution of fusidic acid resistance determinants in methicillin-resistant Staphylococcus aureus. Antimicrob Agents Chemother. 2011;55(3):1173–6.
Nagaev I, Bjorkman J, Andersson DI, Hughes D. Biological cost and compensatory evolution in fusidic acid-resistant Staphylococcus aureus. Mol Microbiol. 2001;40(2):433–9.
O'Neill AJ, Chopra I. Molecular basis of fusB-mediated resistance to fusidic acid in Staphylococcus aureus. Mol Microbiol. 2006;59(2):664–76.
Rijnders MI, Wolffs PF, Hopstaken RM, den Heyer M, Bruggeman CA, Stobberingh EE. Spread of the epidemic European fusidic acid-resistant impetigo clone (EEFIC) in general practice patients in the south of The Netherlands. J Antimicrob Chemother. 2012;67(5):1176–80.
Kim T, Yi J, Hong KH, Park JS, Kim EC. Distribution of virulence genes in spa types of methicillin-resistant Staphylococcus aureus isolated from patients in intensive care units. Korean J Lab Med. 2011;31(1):30–6.
Cheng H, Yuan W, Zeng F, Hu Q, Shang W, Tang D, et al. Molecular and phenotypic evidence for the spread of three major methicillin-resistant Staphylococcus aureus clones associated with two characteristic antimicrobial resistance profiles in China. J Antimicrob Chemother. 2013;68(11):2453–7.
O'Neill AJ, Larsen AR, Skov R, Henriksen AS, Chopra I. Characterization of the epidemic European fusidic acid-resistant impetigo clone of Staphylococcus aureus. J Clin Microbiol. 2007;45(5):1505–10.
Laurent F, Tristan A, Croze M, Bes M, Meugnier H, Lina G, et al. Presence of the epidemic European fusidic acid-resistant impetigo clone (EEFIC) of Staphylococcus aureus in France. J Antimicrob Chemother. 2009;63(2):420–1. author reply 421.
Acknowledgement
We are grateful to Pro. T. Ito for the kind gift of MRSA NCTC 10442 (SCCmec I), MRSA N315 (SCCmec II), MRSA85 ⁄ 2082 (SCCmec III), and MRSA JCSC4744 (SCCmec IV). This study was supported by grants from Natural Science fund of China (81271906H2002 and 81472011H2002) and the Program for Zhejiang Provincial Leading Team for S&T innovation (2011R50018-11).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
YLL, CHL, JNL, XQQ, YD and DL performed the laboratory measurements. FYY and LXW made substantial contributions to conception and design. FYY and LXW revised the manuscript critically for important intellectual content. XYH and LHH participated in experimental design and data analysis. FYY drafted the manuscript. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Yu, F., Liu, Y., Lu, C. et al. Dissemination of fusidic acid resistance among Staphylococcus aureus clinical isolates. BMC Microbiol 15, 210 (2015). https://doi.org/10.1186/s12866-015-0552-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12866-015-0552-z